
Vitamin K-Dependent Proteins in Skeletal Development and Disease

Abstract
1. Introduction
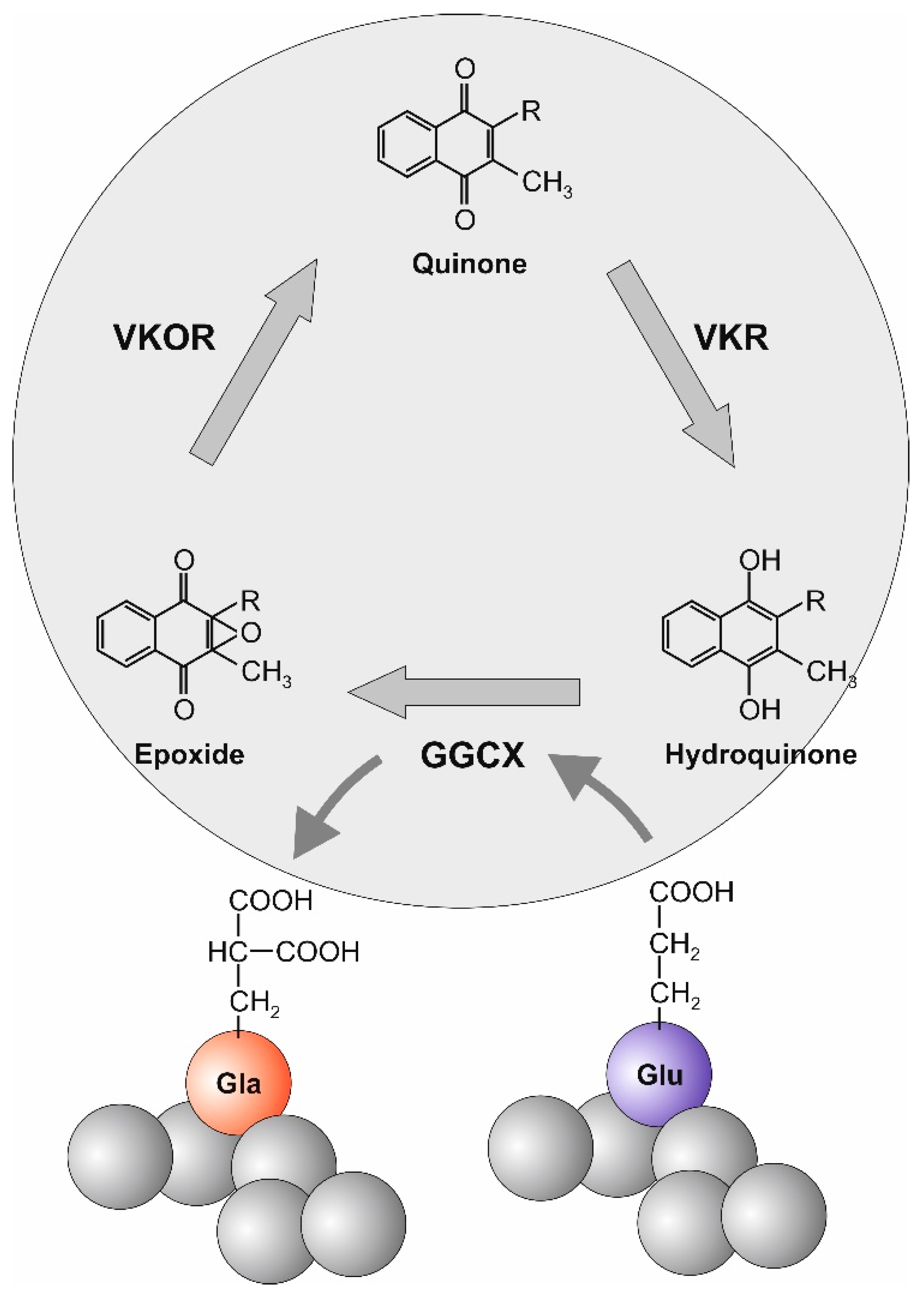
2. Vitamin K
3. Vitamin K in Skeletal Development and Disease
4. VDKP-Independent Effects of Vitamin K on Skeletal Biology
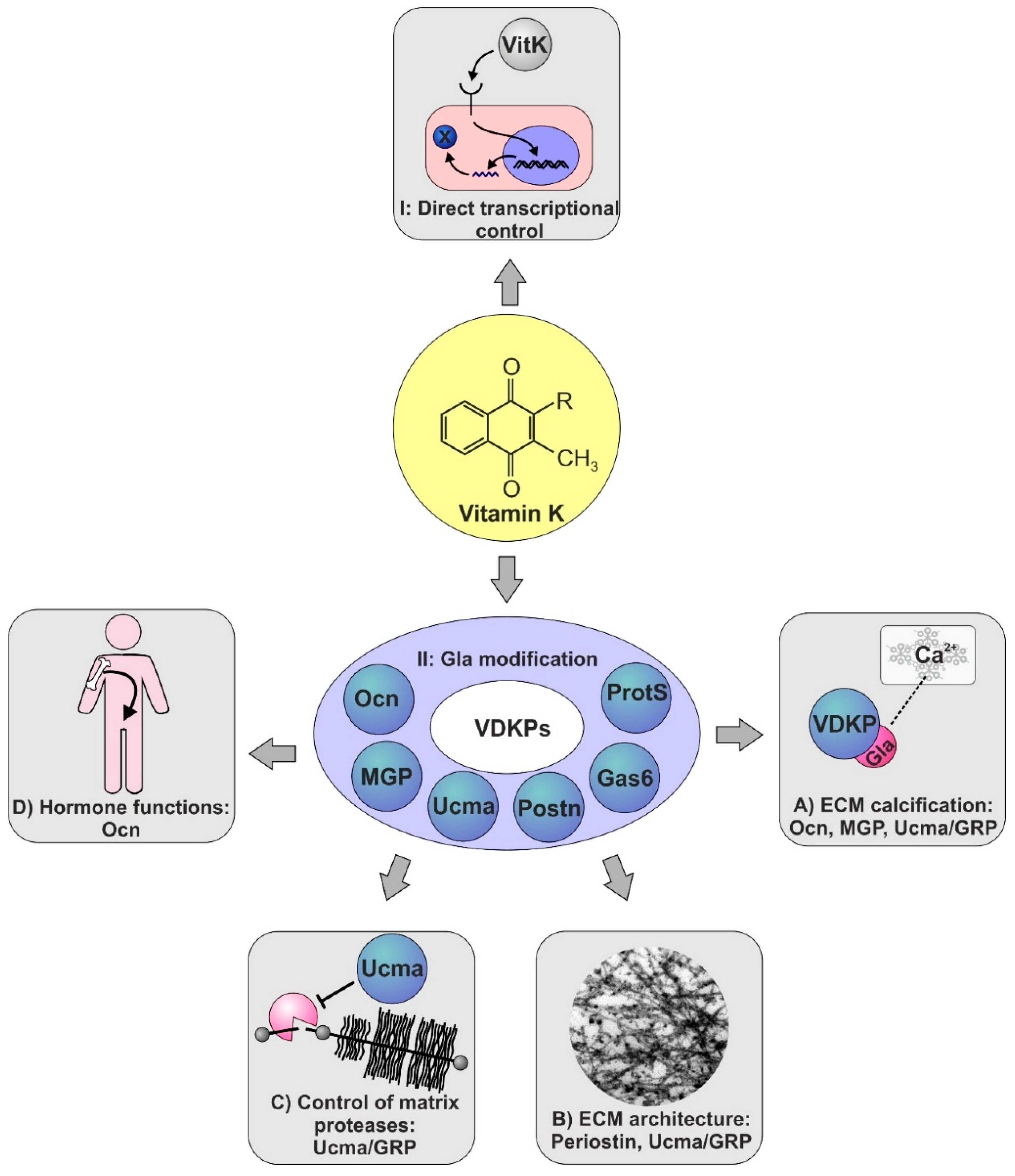
5. Vitamin K-Dependent Proteins in Skeletal Tissues
6. Vit K-Dependent Proteins in Skeletal Biology
6.1. Osteocalcin (OC; Bone-Gla-Protein)
6.2. Matrix Gla Protein (MGP)
6.3. UCMA/GRP
6.4. Periostin, Growth Arrest-Specific Protein 6 (Gas6), and Protein S
7. Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dam, H. The antihaemorrhagic vitamin of the chick. Biochem. J. 1935, 29, 1273–1285. [Google Scholar]
- Azuma, K.; Inoue, S. Multiple Modes of Vitamin K Actions in Aging-Related Musculoskeletal Disorders. Int. J. Mol. Sci. 2019, 20, 2844. [Google Scholar] [CrossRef] [PubMed]
- Sakagami, H.; Satoh, K.; Hakeda, Y.; Kumegawa, M. Apoptosis-inducing activity of vitamin C and vitamin K. Cell Mol. Biol. 2000, 46, 129–143. [Google Scholar] [PubMed]
- Vermeer, C.; Braam, L. Role of K vitamins in the regulation of tissue calcification. J. Bone Miner. Metab. 2001, 19, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Orlando, A.; Linsalata, M.; Tutino, V.; D’Attoma, B.; Notarnicola, M.; Russo, F. Vitamin K1 exerts antiproliferative effects and induces apoptosis in three differently graded human colon cancer cell lines. BioMed Res. Int. 2015, 2015, 296721. [Google Scholar] [CrossRef]
- Schwalfenberg, G.K. Vitamins K1 and K2: The Emerging Group of Vitamins Required for Human Health. J. Nutr. Metab. 2017, 2017, 6254836. [Google Scholar] [CrossRef] [PubMed]
- Harshman, S.G.; Shea, M.K. The Role of Vitamin K in Chronic Aging Diseases: Inflammation, Cardiovascular Disease, and Osteoarthritis. Curr. Nutr. Rep. 2016, 5, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Xv, F.; Chen, J.; Duan, L.; Li, S. Research progress on the anticancer effects of vitamin K2. Oncol. Lett. 2018, 15, 8926–8934. [Google Scholar] [CrossRef]
- Wang, K.; Wu, Q.; Li, Z.; Reger, M.K.; Xiong, Y.; Zhong, G.; Li, Q.; Zhang, X.; Li, H.; Foukakis, T.; et al. Vitamin K intake and breast cancer incidence and death: Results from a prospective cohort study. Clin. Nutr. 2021, 40, 3370–3378. [Google Scholar] [CrossRef] [PubMed]
- Pettifor, J.M.; Benson, R. Congenital malformations associated with the administration of oral anticoagulants during pregnancy. J. Pediatr. 1975, 86, 459–462. [Google Scholar]
- Price, P.A.; Williamson, M.K.; Haba, T.; Dell, R.B.; Jee, W.S. Excessive mineralization with growth plate closure in rats on chronic warfarin treatment. Proc. Natl. Acad. Sci. USA 1982, 79, 7734–7738. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; D’Souza, R.; Hogue, D.; Karsenty, G. The matrix Gla protein gene is a marker of the chondrogenesis cell lineage during mouse development. J. Bone Miner. Res. 1995, 10, 325–334. [Google Scholar] [CrossRef]
- Hauschka, P.V.; Lian, J.B.; Cole, D.E.; Gundberg, C.M. Osteocalcin and matrix Gla protein: Vitamin K-dependent proteins in bone. Physiol. Rev. 1989, 69, 990–1047. [Google Scholar] [CrossRef] [PubMed]
- Yagami, K.; Suh, J.Y.; Enomoto-Iwamoto, M.; Koyama, E.; Abrams, W.R.; Shapiro, I.M.; Pacifici, M.; Iwamoto, M. Matrix GLA protein is a developmental regulator of chondrocyte mineralization and, when constitutively expressed, blocks endochondral and intramembranous ossification in the limb. J. Cell Biol. 1999, 147, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Ducy, P.; Desbois, C.; Boyce, B.; Pinero, G.; Story, B.; Dunstan, C.; Smith, E.; Bonadio, J.; Goldstein, S.; Gundberg, C.; et al. Increased bone formation in osteocalcin-deficient mice. Nature 1996, 382, 448–452. [Google Scholar] [CrossRef]
- Luo, G.; Ducy, P.; McKee, M.D.; Pinero, G.J.; Loyer, E.; Behringer, R.R.; Karsenty, G. Spontaneous calcification of arteries and cartilage in mice lacking matrix GLA protein. Nature 1997, 386, 78–81. [Google Scholar] [CrossRef]
- Akbari, S.; Rasouli-Ghahroudi, A.A. Vitamin K and Bone Metabolism: A Review of the Latest Evidence in Preclinical Studies. BioMed Res. Int. 2018, 2018, 4629383. [Google Scholar] [CrossRef]
- Lombardi, G.; Perego, S.; Luzi, L.; Banfi, G. A four-season molecule: Osteocalcin. Updates in its physiological roles. Endocrine 2015, 48, 394–404. [Google Scholar] [CrossRef]
- Ichikawa, T.; Horie-Inoue, K.; Ikeda, K.; Blumberg, B.; Inoue, S. Steroid and xenobiotic receptor SXR mediates vitamin K2-activated transcription of extracellular matrix-related genes and collagen accumulation in osteoblastic cells. J. Biol. Chem. 2006, 281, 16927–16934. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Casey, S.C.; Ito, M.; Urano, T.; Horie, K.; Ouchi, Y.; Kirchner, S.; Blumberg, B.; Inoue, S. Pregnane X receptor knockout mice display osteopenia with reduced bone formation and enhanced bone resorption. J. Endocrinol. 2010, 207, 257–263. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Weitzmann, M.N. Vitamin K2 stimulates osteoblastogenesis and suppresses osteoclastogenesis by suppressing NF-kappaB activation. Int. J. Mol. Med. 2011, 27, 3–14. [Google Scholar] [PubMed]
- Surmann-Schmitt, C.; Dietz, U.; Kireva, T.; Adam, N.; Park, J.; Tagariello, A.; Onnerfjord, P.; Heinegard, D.; Schlotzer-Schrehardt, U.; Deutzmann, R.; et al. Ucma, a novel secreted cartilage-specific protein with implications in osteogenesis. J. Biol. Chem. 2008, 283, 7082–7093. [Google Scholar] [CrossRef] [PubMed]
- Tagariello, A.; Luther, J.; Streiter, M.; Didt-Koziel, L.; Wuelling, M.; Surmann-Schmitt, C.; Stock, M.; Adam, N.; Vortkamp, A.; Winterpacht, A. Ucma—A novel secreted factor represents a highly specific marker for distal chondrocytes. Matrix Biol. 2008, 27, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Viegas, C.S.; Simes, D.C.; Laize, V.; Williamson, M.K.; Price, P.A.; Cancela, M.L. Gla-rich Protein (GRP), A New Vitamin K-dependent Protein Identified from Sturgeon Cartilage and Highly Conserved in Vertebrates. J. Biol. Chem. 2008, 283, 36655–36664. [Google Scholar] [CrossRef] [PubMed]
- Neacsu, C.D.; Grosch, M.; Tejada, M.; Winterpacht, A.; Paulsson, M.; Wagener, R.; Tagariello, A. Ucmaa (Grp-2) is required for zebrafish skeletal development. Evidence for a functional role of its glutamate gamma-carboxylation. Matrix Biol. 2011, 30, 369–378. [Google Scholar] [CrossRef]
- Eitzinger, N.; Surmann-Schmitt, C.; Bosl, M.; Schett, G.; Engelke, K.; Hess, A.; von der Mark, K.; Stock, M. Ucma is not necessary for normal development of the mouse skeleton. Bone 2012, 50, 670–680. [Google Scholar] [CrossRef] [PubMed]
- Seuffert, F.; Weidner, D.; Baum, W.; Schett, G.; Stock, M. Upper zone of growth plate and cartilage matrix associated protein protects cartilage during inflammatory arthritis. Arthritis Res. Ther. 2018, 20, 88. [Google Scholar] [CrossRef]
- Stock, M.; Menges, S.; Eitzinger, N.; Gesslein, M.; Botschner, R.; Wormser, L.; Distler, A.; Schlotzer-Schrehardt, U.; Dietel, K.; Distler, J.; et al. A Dual Role of Upper Zone of Growth Plate and Cartilage Matrix-Associated Protein in Human and Mouse Osteoarthritic Cartilage: Inhibition of Aggrecanases and Promotion of Bone Turnover. Arthritis Rheumatol. 2017, 69, 1233–1245. [Google Scholar] [CrossRef]
- Nakamura, Y.S.; Hakeda, Y.; Takakura, N.; Kameda, T.; Hamaguchi, I.; Miyamoto, T.; Kakudo, S.; Nakano, T.; Kumegawa, M.; Suda, T. Tyro 3 receptor tyrosine kinase and its ligand, Gas6, stimulate the function of osteoclasts. Stem Cells 1998, 16, 229–238. [Google Scholar] [CrossRef]
- Coutu, D.L.; Wu, J.H.; Monette, A.; Rivard, G.E.; Blostein, M.D.; Galipeau, J. Periostin, a member of a novel family of vitamin K-dependent proteins, is expressed by mesenchymal stromal cells. J. Biol. Chem. 2008, 283, 17991–18001. [Google Scholar] [CrossRef]
- Rios, H.; Koushik, S.V.; Wang, H.; Wang, J.; Zhou, H.M.; Lindsley, A.; Rogers, R.; Chen, Z.; Maeda, M.; Kruzynska-Frejtag, A.; et al. Periostin null mice exhibit dwarfism, incisor enamel defects, and an early-onset periodontal disease-like phenotype. Mol. Cell. Biol. 2005, 25, 11131–11144. [Google Scholar] [CrossRef]
- Chijimatsu, R.; Kunugiza, Y.; Taniyama, Y.; Nakamura, N.; Tomita, T.; Yoshikawa, H. Expression and pathological effects of periostin in human osteoarthritis cartilage. BMC Musculoskelet. Disord. 2015, 16, 215. [Google Scholar] [CrossRef]
- Heo, S.C.; Shin, W.C.; Lee, M.J.; Kim, B.R.; Jang, I.H.; Choi, E.J.; Lee, J.S.; Kim, J.H. Periostin accelerates bone healing mediated by human mesenchymal stem cell-embedded hydroxyapatite/tricalcium phosphate scaffold. PLoS ONE 2015, 10, e0116698. [Google Scholar] [CrossRef]
- Maillard, C.; Berruyer, M.; Serre, C.M.; Dechavanne, M.; Delmas, P.D. Protein-S, a vitamin K-dependent protein, is a bone matrix component synthesized and secreted by osteoblasts. Endocrinology 1992, 130, 1599–1604. [Google Scholar]
- Schurgers, L.J.; Vermeer, C. Determination of phylloquinone and menaquinones in food. Effect of food matrix on circulating vitamin K concentrations. Haemostasis 2000, 30, 298–307. [Google Scholar] [PubMed]
- Nelsestuen, G.L.; Zytkovicz, T.H.; Howard, J.B. The mode of action of vitamin K. Identification of gamma-carboxyglutamic acid as a component of prothrombin. J. Biol. Chem. 1974, 249, 6347–6350. [Google Scholar] [CrossRef]
- Stenflo, J.; Fernlund, P.; Egan, W.; Roepstorff, P. Vitamin K dependent modifications of glutamic acid residues in prothrombin. Proc. Natl. Acad. Sci. USA 1974, 71, 2730–2733. [Google Scholar] [CrossRef] [PubMed]
- Czogalla, K.J.; Biswas, A.; Honing, K.; Hornung, V.; Liphardt, K.; Watzka, M.; Oldenburg, J. Warfarin and vitamin K compete for binding to Phe55 in human VKOR. Nat. Struct. Mol. Biol. 2017, 24, 77–85. [Google Scholar] [CrossRef]
- Feskanich, D.; Weber, P.; Willett, W.C.; Rockett, H.; Booth, S.L.; Colditz, G.A. Vitamin K intake and hip fractures in women: A prospective study. Am. J. Clin. Nutr. 1999, 69, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Caraballo, P.J.; Heit, J.A.; Atkinson, E.J.; Silverstein, M.D.; O’Fallon, W.M.; Castro, M.R.; Melton, L.J. Long-term use of oral anticoagulants and the risk of fracture. Arch. Intern. Med. 1999, 159, 1750–1756. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.Y. The Relationship between Vitamin K and Osteoarthritis: A Review of Current Evidence. Nutrients 2020, 12, 1208. [Google Scholar] [CrossRef]
- Boer, C.G.; Szilagyi, I.; Nguyen, N.L.; Neogi, T.; Meulenbelt, I.; Ikram, M.A.; Uitterlinden, A.G.; Bierma-Zeinstra, S.; Stricker, B.H.; van Meurs, J.B. Vitamin K antagonist anticoagulant usage is associated with increased incidence and progression of osteoarthritis. Ann. Rheum. Dis. 2021, 80, 598–604. [Google Scholar] [CrossRef]
- Okamoto, H. Vitamin K and rheumatoid arthritis. IUBMB Life 2008, 60, 355–361. [Google Scholar] [CrossRef]
- Ebina, K.; Shi, K.; Hirao, M.; Kaneshiro, S.; Morimoto, T.; Koizumi, K.; Yoshikawa, H.; Hashimoto, J. Vitamin K2 administration is associated with decreased disease activity in patients with rheumatoid arthritis. Mod. Rheumatol. 2013, 23, 1001–1007. [Google Scholar] [CrossRef]
- Kim, M.; Na, W.; Sohn, C. Vitamin K1 (phylloquinone) and K2 (menaquinone-4) supplementation improves bone formation in a high-fat diet-induced obese mice. J. Clin. Biochem. Nutr. 2013, 53, 108–113. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Igarashi, M.; Yogiashi, Y.; Mihara, M.; Takada, I.; Kitagawa, H.; Kato, S. Vitamin K induces osteoblast differentiation through pregnane X receptor-mediated transcriptional control of the Msx2 gene. Mol. Cell. Biol. 2007, 27, 7947–7954. [Google Scholar] [CrossRef]
- Derer, A.; Bohm, C.; Grotsch, B.; Grun, J.R.; Grutzkau, A.; Stock, M.; Bohm, S.; Sehnert, B.; Gaipl, U.; Schett, G.; et al. Rsk2 controls synovial fibroblast hyperplasia and the course of arthritis. Ann. Rheum. Dis. 2016, 75, 413–421. [Google Scholar] [CrossRef] [PubMed]
- McInnes, I.B.; Schett, G. The pathogenesis of rheumatoid arthritis. N. Engl. J. Med. 2011, 365, 2205–2219. [Google Scholar] [CrossRef] [PubMed]
- Criddle, D.N.; Gillies, S.; Baumgartner-Wilson, H.K.; Jaffar, M.; Chinje, E.C.; Passmore, S.; Chvanov, M.; Barrow, S.; Gerasimenko, O.V.; Tepikin, A.V.; et al. Menadione-induced reactive oxygen species generation via redox cycling promotes apoptosis of murine pancreatic acinar cells. J. Biol. Chem. 2006, 281, 40485–40492. [Google Scholar] [CrossRef] [PubMed]
- McInnes, I.B.; Schett, G. Cytokines in the pathogenesis of rheumatoid arthritis. Nat. Rev. Immunol. 2007, 7, 429–442. [Google Scholar] [CrossRef]
- Katagiri, M.; Hakeda, Y.; Chikazu, D.; Ogasawara, T.; Takato, T.; Kumegawa, M.; Nakamura, K.; Kawaguchi, H. Mechanism of stimulation of osteoclastic bone resorption through Gas6/Tyro 3, a receptor tyrosine kinase signaling, in mouse osteoclasts. J. Biol. Chem. 2001, 276, 7376–7382. [Google Scholar] [CrossRef]
- Gundberg, C.M.; Clough, M.E. The osteocalcin propeptide is not secreted in vivo or in vitro. J. Bone Miner. Res. 1992, 7, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Poser, J.W.; Esch, F.S.; Ling, N.C.; Price, P.A. Isolation and sequence of the vitamin K-dependent protein from human bone. Undercarboxylation of the first glutamic acid residue. J. Biol. Chem. 1980, 255, 8685–8691. [Google Scholar] [CrossRef]
- Price, P.A. Gla-containing proteins of bone. Connect. Tissue Res. 1989, 21, 51–57; discussion 57–60. [Google Scholar] [CrossRef]
- Hauschka, P.V.; Wians, F.H., Jr. Osteocalcin-hydroxyapatite interaction in the extracellular organic matrix of bone. Anat. Rec. 1989, 224, 180–188. [Google Scholar] [CrossRef]
- Hoang, Q.Q.; Sicheri, F.; Howard, A.J.; Yang, D.S. Bone recognition mechanism of porcine osteocalcin from crystal structure. Nature 2003, 425, 977–980. [Google Scholar] [CrossRef] [PubMed]
- Boskey, A.L.; Gadaleta, S.; Gundberg, C.; Doty, S.B.; Ducy, P.; Karsenty, G. Fourier transform infrared microspectroscopic analysis of bones of osteocalcin-deficient mice provides insight into the function of osteocalcin. Bone 1998, 23, 187–196. [Google Scholar] [CrossRef]
- Fratzl, P.; Paris, O.; Klaushofer, K.; Landis, W.J. Bone mineralization in an osteogenesis imperfecta mouse model studied by small-angle X-ray scattering. J. Clin. Investig. 1996, 97, 396–402. [Google Scholar] [CrossRef][Green Version]
- Brown, J.P.; Delmas, P.D.; Malaval, L.; Edouard, C.; Chapuy, M.C.; Meunier, P.J. Serum bone Gla-protein: A specific marker for bone formation in postmenopausal osteoporosis. Lancet 1984, 1, 1091–1093. [Google Scholar] [CrossRef]
- Plantalech, L.; Guillaumont, M.; Vergnaud, P.; Leclercq, M.; Delmas, P.D. Impairment of gamma carboxylation of circulating osteocalcin (bone gla protein) in elderly women. J. Bone Miner. Res. 1991, 6, 1211–1216. [Google Scholar] [CrossRef]
- Knapen, M.H.; Eisenwiener, H.G.; Vermeer, C. Osteocalcin detection in aging serum and whole blood: Stability of different osteocalcin fractions. Clin. Chim. Acta Int. J. Clin. Chem. 1996, 256, 151–164. [Google Scholar] [CrossRef]
- Szulc, P.; Chapuy, M.C.; Meunier, P.J.; Delmas, P.D. Serum undercarboxylated osteocalcin is a marker of the risk of hip fracture in elderly women. J. Clin. Investig. 1993, 91, 1769–1774. [Google Scholar] [CrossRef]
- Booth, S.L.; Tucker, K.L.; Chen, H.; Hannan, M.T.; Gagnon, D.R.; Cupples, L.A.; Wilson, P.W.; Ordovas, J.; Schaefer, E.J.; Dawson-Hughes, B.; et al. Dietary vitamin K intakes are associated with hip fracture but not with bone mineral density in elderly men and women. Am. J. Clin. Nutr. 2000, 71, 1201–1208. [Google Scholar] [CrossRef]
- Booth, S.L.; Broe, K.E.; Gagnon, D.R.; Tucker, K.L.; Hannan, M.T.; McLean, R.R.; Dawson-Hughes, B.; Wilson, P.W.; Cupples, L.A.; Kiel, D.P. Vitamin K intake and bone mineral density in women and men. Am. J. Clin. Nutr. 2003, 77, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Pullig, O.; Weseloh, G.; Ronneberger, D.; Kakonen, S.; Swoboda, B. Chondrocyte differentiation in human osteoarthritis: Expression of osteocalcin in normal and osteoarthritic cartilage and bone. Calcif. Tissue Int. 2000, 67, 230–240. [Google Scholar] [CrossRef]
- Campion, G.V.; Delmas, P.D.; Dieppe, P.A. Serum and synovial fluid osteocalcin (bone gla protein) levels in joint disease. Br. J. Rheumatol. 1989, 28, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Hilal, G.; Martel-Pelletier, J.; Pelletier, J.P.; Ranger, P.; Lajeunesse, D. Osteoblast-like cells from human subchondral osteoarthritic bone demonstrate an altered phenotype in vitro: Possible role in subchondral bone sclerosis. Arthritis Rheum. 1998, 41, 891–899. [Google Scholar] [CrossRef]
- Naito, K.; Watari, T.; Obayashi, O.; Katsube, S.; Nagaoka, I.; Kaneko, K. Relationship between serum undercarboxylated osteocalcin and hyaluronan levels in patients with bilateral knee osteoarthritis. Int. J. Mol. Med. 2012, 29, 756–760. [Google Scholar] [CrossRef][Green Version]
- Seriolo, B.; Ferretti, V.; Sulli, A.; Caratto, E.; Fasciolo, D.; Cutolo, M. Serum osteocalcin levels in premenopausal rheumatoid arthritis patients. Ann. N. Y. Acad. Sci. 2002, 966, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Calvo, M.S.; Eyre, D.R.; Gundberg, C.M. Molecular basis and clinical application of biological markers of bone turnover. Endocr. Rev. 1996, 17, 333–368. [Google Scholar]
- van den Berg, W.B. Osteoarthritis year 2010 in review: Pathomechanisms. Osteoarthr. Cartil. 2011, 19, 338–341. [Google Scholar] [CrossRef] [PubMed]
- Oury, F.; Sumara, G.; Sumara, O.; Ferron, M.; Chang, H.; Smith, C.E.; Hermo, L.; Suarez, S.; Roth, B.L.; Ducy, P.; et al. Endocrine regulation of male fertility by the skeleton. Cell 2011, 144, 796–809. [Google Scholar] [CrossRef] [PubMed]
- Oury, F.; Khrimian, L.; Denny, C.A.; Gardin, A.; Chamouni, A.; Goeden, N.; Huang, Y.Y.; Lee, H.; Srinivas, P.; Gao, X.B.; et al. Maternal and offspring pools of osteocalcin influence brain development and functions. Cell 2013, 155, 228–241. [Google Scholar] [CrossRef]
- Lee, N.K.; Sowa, H.; Hinoi, E.; Ferron, M.; Ahn, J.D.; Confavreux, C.; Dacquin, R.; Mee, P.J.; McKee, M.D.; Jung, D.Y.; et al. Endocrine regulation of energy metabolism by the skeleton. Cell 2007, 130, 456–469. [Google Scholar] [CrossRef]
- Ferron, M.; Hinoi, E.; Karsenty, G.; Ducy, P. Osteocalcin differentially regulates beta cell and adipocyte gene expression and affects the development of metabolic diseases in wild-type mice. Proc. Natl. Acad. Sci. USA 2008, 105, 5266–5270. [Google Scholar] [CrossRef]
- Price, P.A.; Poser, J.W.; Raman, N. Primary structure of the gamma-carboxyglutamic acid-containing protein from bovine bone. Proc. Natl. Acad. Sci. USA 1976, 73, 3374–3375. [Google Scholar] [CrossRef] [PubMed]
- Price, P.A.; Otsuka, A.A.; Poser, J.W.; Kristaponis, J.; Raman, N. Characterization of a gamma-carboxyglutamic acid-containing protein from bone. Proc. Natl. Acad. Sci. USA 1976, 73, 1447–1451. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.E.; Nishimoto, S.K. Matrix Gla protein binding to hydroxyapatite is dependent on the ionic environment: Calcium enhances binding affinity but phosphate and magnesium decrease affinity. Bone 2002, 31, 296–302. [Google Scholar] [CrossRef]
- Murshed, M.; Schinke, T.; McKee, M.D.; Karsenty, G. Extracellular matrix mineralization is regulated locally; different roles of two gla-containing proteins. J. Cell Biol. 2004, 165, 625–630. [Google Scholar] [CrossRef]
- Wallin, R.; Cain, D.; Hutson, S.M.; Sane, D.C.; Loeser, R. Modulation of the binding of matrix Gla protein (MGP) to bone morphogenetic protein-2 (BMP-2). Thromb. Haemost. 2000, 84, 1039–1044. [Google Scholar]
- Wallin, R.; Schurgers, L.J.; Loeser, R.F. Biosynthesis of the vitamin K-dependent matrix Gla protein (MGP) in chondrocytes: A fetuin-MGP protein complex is assembled in vesicles shed from normal but not from osteoarthritic chondrocytes. Osteoarthr. Cartil. 2010, 18, 1096–1103. [Google Scholar] [CrossRef]
- Xue, W.; Wallin, R.; Olmsted-Davis, E.A.; Borras, T. Matrix GLA protein function in human trabecular meshwork cells: Inhibition of BMP2-induced calcification process. Investig. Ophthalmol. Vis. Sci. 2006, 47, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Misra, D.; Booth, S.L.; Crosier, M.D.; Ordovas, J.M.; Felson, D.T.; Neogi, T. Matrix Gla protein polymorphism, but not concentrations, is associated with radiographic hand osteoarthritis. J. Rheumatol. 2011, 38, 1960–1965. [Google Scholar] [CrossRef] [PubMed]
- Silaghi, C.N.; Fodor, D.; Cristea, V.; Craciun, A.M. Synovial and serum levels of uncarboxylated matrix Gla-protein (ucMGP) in patients with arthritis. Clin. Chem. Lab. Med. 2011, 50, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Bing, W.; Feng, L. Attenuate Synovial Fluid Uncarboxylated Matrix Gla-Protein (ucMGP) Concentrations Are Linked with Radiographic Progression in Knee Psteoarthritis. Adv. Clin. Exp. Med. 2015, 24, 1013–1017. [Google Scholar] [CrossRef]
- Wei, F.F.; Trenson, S.; Verhamme, P.; Vermeer, C.; Staessen, J.A. Vitamin K-Dependent Matrix Gla Protein as Multifaceted Protector of Vascular and Tissue Integrity. Hypertension 2019, 73, 1160–1169. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, Z.; Yan, K.; Wang, Y.; Yang, Y.; Wu, X. Matrix Gla Protein Promotes the Bone Formation by Up-Regulating Wnt/β-Catenin Signaling Pathway. Front. Endocrinol. 2019, 10, 891. [Google Scholar] [CrossRef]
- Lee, Y.J.; Park, S.Y.; Lee, S.J.; Boo, Y.C.; Choi, J.Y.; Kim, J.E. Ucma, a direct transcriptional target of Runx2 and Osterix, promotes osteoblast differentiation and nodule formation. Osteoarthr. Cartil. 2015, 23, 1421–1431. [Google Scholar] [CrossRef]
- Le Jeune, M.; Tomavo, N.; Tian, T.V.; Flourens, A.; Marchand, N.; Camuzeaux, B.; Mallein-Gerin, F.; Duterque-Coquillaud, M. Identification of four alternatively spliced transcripts of the Ucma/GRP gene, encoding a new Gla-containing protein. Exp. Cell Res. 2010, 316, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Rafael, M.S.; Cavaco, S.; Viegas, C.S.; Santos, S.; Ramos, A.; Willems, B.A.; Herfs, M.; Theuwissen, E.; Vermeer, C.; Simes, D.C. Insights into the association of Gla-rich protein and osteoarthritis, novel splice variants and gamma-carboxylation status. Mol. Nutr. Food Res. 2014, 58, 1636–1646. [Google Scholar] [CrossRef]
- Onnerfjord, P.; Heathfield, T.F.; Heinegard, D. Identification of tyrosine sulfation in extracellular leucine-rich repeat proteins using mass spectrometry. J. Biol. Chem. 2004, 279, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Leyte, A.; van Schijndel, H.B.; Niehrs, C.; Huttner, W.B.; Verbeet, M.P.; Mertens, K.; van Mourik, J.A. Sulfation of Tyr1680 of human blood coagulation factor VIII is essential for the interaction of factor VIII with von Willebrand factor. J. Biol. Chem. 1991, 266, 740–746. [Google Scholar] [CrossRef]
- Michou, L.; Conceicao, N.; Morissette, J.; Gagnon, E.; Miltenberger-Miltenyi, G.; Siris, E.S.; Brown, J.P.; Cancela, M.L. Genetic association study of UCMA/GRP and OPTN genes (PDB6 locus) with Paget’s disease of bone. Bone 2012, 51, 720–728. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Willems, B.A.; Furmanik, M.; Caron, M.M.J.; Chatrou, M.L.L.; Kusters, D.H.M.; Welting, T.J.M.; Stock, M.; Rafael, M.S.; Viegas, C.S.B.; Simes, D.C.; et al. Ucma/GRP inhibits phosphate-induced vascular smooth muscle cell calcification via SMAD-dependent BMP signalling. Sci. Rep. 2018, 8, 4961. [Google Scholar] [CrossRef] [PubMed]
- Viegas, C.S.B.; Santos, L.; Macedo, A.L.; Matos, A.A.; Silva, A.P.; Neves, P.L.; Staes, A.; Gevaert, K.; Morais, R.; Vermeer, C.; et al. Chronic Kidney Disease Circulating Calciprotein Particles and Extracellular Vesicles Promote Vascular Calcification: A Role for GRP (Gla-Rich Protein). Arterioscler. Thromb. Vasc. Biol. 2018, 38, 575–587. [Google Scholar] [CrossRef]
- Viegas, C.S.; Cavaco, S.; Neves, P.L.; Ferreira, A.; Joao, A.; Williamson, M.K.; Price, P.A.; Cancela, M.L.; Simes, D.C. Gla-rich protein is a novel vitamin K-dependent protein present in serum that accumulates at sites of pathological calcifications. Am. J. Pathol. 2009, 175, 2288–2298. [Google Scholar] [CrossRef]
- Silva, A.P.; Viegas, C.S.; Mendes, F.; Macedo, A.; Guilherme, P.; Tavares, N.; Dias, C.; Rato, F.; Santos, N.; Faisca, M.; et al. Gla-Rich Protein (GRP) as an Early and Novel Marker of Vascular Calcification and Kidney Dysfunction in Diabetic Patients with CKD: A Pilot Cross-Sectional Study. J. Clin. Med. 2020, 9, 635. [Google Scholar] [CrossRef]
- Cavaco, S.; Viegas, C.S.; Rafael, M.S.; Ramos, A.; Magalhaes, J.; Blanco, F.J.; Vermeer, C.; Simes, D.C. Gla-rich protein is involved in the cross-talk between calcification and inflammation in osteoarthritis. Cell. Mol. Life Sci. CMLS 2016, 73, 1051–1065. [Google Scholar] [CrossRef]
- Okuyan, H.M.; Terzi, M.Y.; Karaboğa, İ.; Doğan, S.; Kalacı, A. In vivo protective effects of upper zone of growth plate and cartilage matrix associated protein against cartilage degeneration in a monosodium iodoacetate induced osteoarthritis model. Can. J. Physiol. Pharmacol. 2020, 98, 763–770. [Google Scholar] [CrossRef]
- Okuyan, H.M.; Terzi, M.Y.; Ozcan, O.; Kalaci, A. Association of UCMA levels in serum and synovial fluid with severity of knee osteoarthritis. Int. J. Rheum. Dis. 2019, 22, 1884–1890. [Google Scholar] [CrossRef]
- Horiuchi, K.; Amizuka, N.; Takeshita, S.; Takamatsu, H.; Katsuura, M.; Ozawa, H.; Toyama, Y.; Bonewald, L.F.; Kudo, A. Identification and characterization of a novel protein, periostin, with restricted expression to periosteum and periodontal ligament and increased expression by transforming growth factor beta. J. Bone Miner. Res. 1999, 14, 1239–1249. [Google Scholar] [CrossRef]
- Bonnet, N.; Standley, K.N.; Bianchi, E.N.; Stadelmann, V.; Foti, M.; Conway, S.J.; Ferrari, S.L. The matricellular protein periostin is required for sost inhibition and the anabolic response to mechanical loading and physical activity. J. Biol. Chem. 2009, 284, 35939–35950. [Google Scholar] [CrossRef] [PubMed]
- Norris, R.A.; Damon, B.; Mironov, V.; Kasyanov, V.; Ramamurthi, A.; Moreno-Rodriguez, R.; Trusk, T.; Potts, J.D.; Goodwin, R.L.; Davis, J.; et al. Periostin regulates collagen fibrillogenesis and the biomechanical properties of connective tissues. J. Cell. Biochem. 2007, 101, 695–711. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, J.C.; Sornay-Rendu, E.; Bertholon, C.; Garnero, P.; Chapurlat, R. Serum periostin is associated with prevalent knee osteoarthritis and disease incidence/progression in women: The OFELY study. Osteoarthr. Cartil. 2015, 23, 1736–1742. [Google Scholar] [CrossRef] [PubMed]
- Honsawek, S.; Wilairatana, V.; Udomsinprasert, W.; Sinlapavilawan, P.; Jirathanathornnukul, N. Association of plasma and synovial fluid periostin with radiographic knee osteoarthritis: Cross-sectional study. Jt. Bone Spine 2015, 82, 352–355. [Google Scholar] [CrossRef]
- Attur, M.; Duan, X.; Cai, L.; Han, T.; Zhang, W.; Tycksen, E.D.; Samuels, J.; Brophy, R.H.; Abramson, S.B.; Rai, M.F. Periostin loss-of-function protects mice from post-traumatic and age-related osteoarthritis. Arthritis Res. Ther. 2021, 23, 104. [Google Scholar] [CrossRef] [PubMed]
- Pan, E.Y.; Gomperts, E.D.; Millen, R.; Gilsanz, V. Bone mineral density and its association with inherited protein S deficiency. Thromb. Res. 1990, 58, 221–231. [Google Scholar] [CrossRef]
- Stanton, H.; Rogerson, F.M.; East, C.J.; Golub, S.B.; Lawlor, K.E.; Meeker, C.T.; Little, C.B.; Last, K.; Farmer, P.J.; Campbell, I.K.; et al. ADAMTS5 is the major aggrecanase in mouse cartilage in vivo and in vitro. Nature 2005, 434, 648–652. [Google Scholar] [CrossRef] [PubMed]
- Song, R.H.; Tortorella, M.D.; Malfait, A.M.; Alston, J.T.; Yang, Z.; Arner, E.C.; Griggs, D.W. Aggrecan degradation in human articular cartilage explants is mediated by both ADAMTS-4 and ADAMTS-5. Arthritis Rheum. 2007, 56, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Glasson, S.S.; Askew, R.; Sheppard, B.; Carito, B.; Blanchet, T.; Ma, H.L.; Flannery, C.R.; Peluso, D.; Kanki, K.; Yang, Z.; et al. Deletion of active ADAMTS5 prevents cartilage degradation in a murine model of osteoarthritis. Nature 2005, 434, 644–648. [Google Scholar] [CrossRef]
- Larkin, J.; Lohr, T.A.; Elefante, L.; Shearin, J.; Matico, R.; Su, J.L.; Xue, Y.; Liu, F.; Genell, C.; Miller, R.E.; et al. Translational development of an ADAMTS-5 antibody for osteoarthritis disease modification. Osteoarthr. Cartil. 2015, 23, 1254–1266. [Google Scholar] [CrossRef] [PubMed]
- Durham, T.B.; Klimkowski, V.J.; Rito, C.J.; Marimuthu, J.; Toth, J.L.; Liu, C.; Durbin, J.D.; Stout, S.L.; Adams, L.; Swearingen, C.; et al. Identification of potent and selective hydantoin inhibitors of aggrecanase-1 and aggrecanase-2 that are efficacious in both chemical and surgical models of osteoarthritis. J. Med. Chem. 2014, 57, 10476–10485. [Google Scholar] [CrossRef] [PubMed]
- Didangelos, A.; Mayr, U.; Monaco, C.; Mayr, M. Novel role of ADAMTS-5 protein in proteoglycan turnover and lipoprotein retention in atherosclerosis. J. Biol. Chem. 2012, 287, 19341–19345. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Vitamin K1 (Phylloquinone) | Vitamin K2 (Menaquinone) | Vitamin K3 (Menadione) | |
|---|---|---|---|
| Synonym | MK-n | ||
| Molecular Structure |  |  |  |
| Origin | Vegetables | Fermented foods | synthetic |
| Usage | Human diet | Human diet | Widely used in animal food but not for human diet |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stock, M.; Schett, G. Vitamin K-Dependent Proteins in Skeletal Development and Disease. Int. J. Mol. Sci. 2021, 22, 9328. https://doi.org/10.3390/ijms22179328
Stock M, Schett G. Vitamin K-Dependent Proteins in Skeletal Development and Disease. International Journal of Molecular Sciences. 2021; 22(17):9328. https://doi.org/10.3390/ijms22179328
Chicago/Turabian StyleStock, Michael, and Georg Schett. 2021. "Vitamin K-Dependent Proteins in Skeletal Development and Disease" International Journal of Molecular Sciences 22, no. 17: 9328. https://doi.org/10.3390/ijms22179328
APA StyleStock, M., & Schett, G. (2021). Vitamin K-Dependent Proteins in Skeletal Development and Disease. International Journal of Molecular Sciences, 22(17), 9328. https://doi.org/10.3390/ijms22179328