
Regulatory Mechanisms of Retinal Photoreceptors Development at Single Cell Resolution


Abstract
1. Introduction
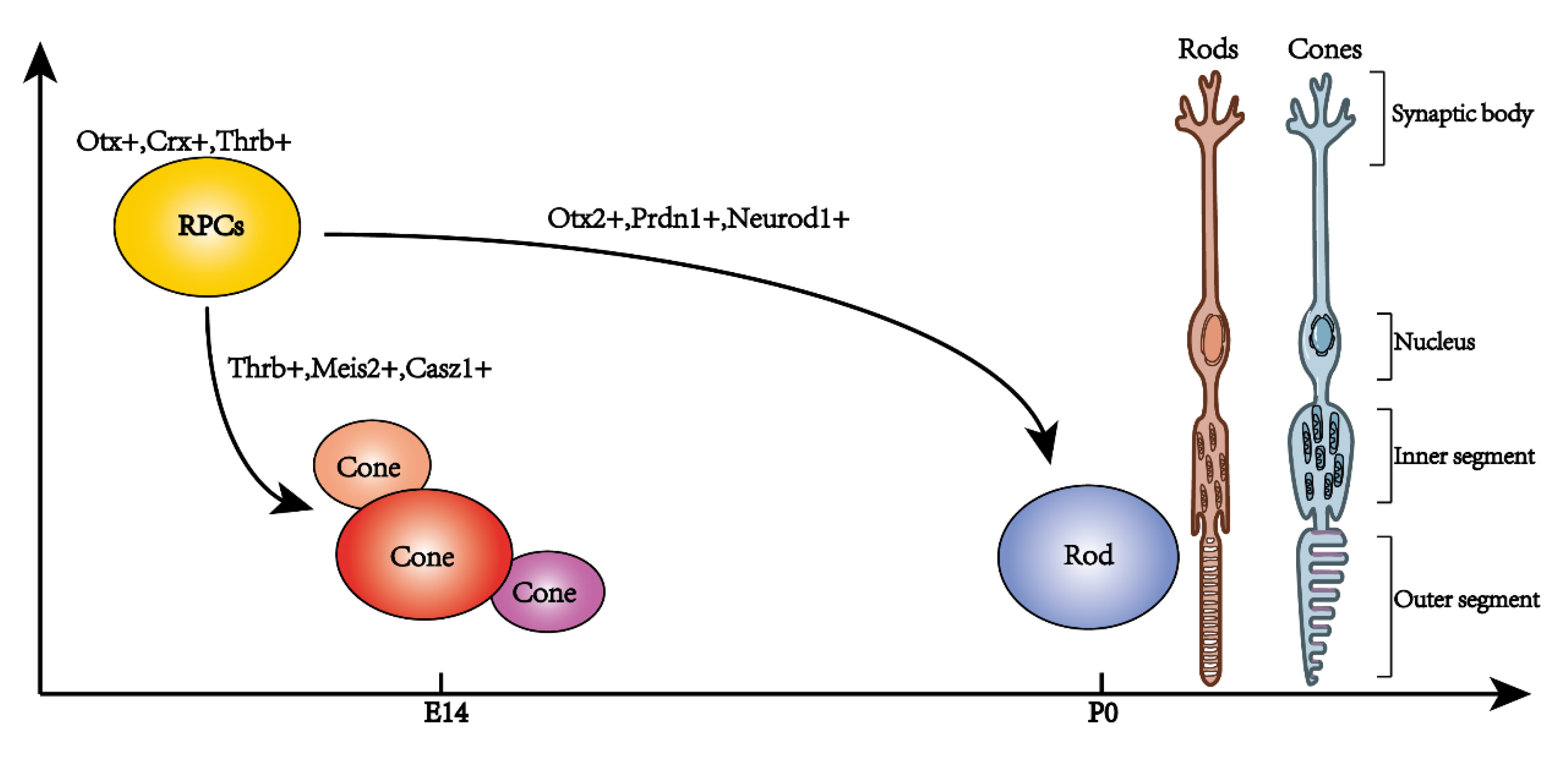
2. Single-Cell Sequencing Analysis in Mice Retinal Photoreceptor Development
3. Single-Cell Sequencing Analysis in Human Retinal Photoreceptor Developmental Process
3.1. Human Fetal Retina Development
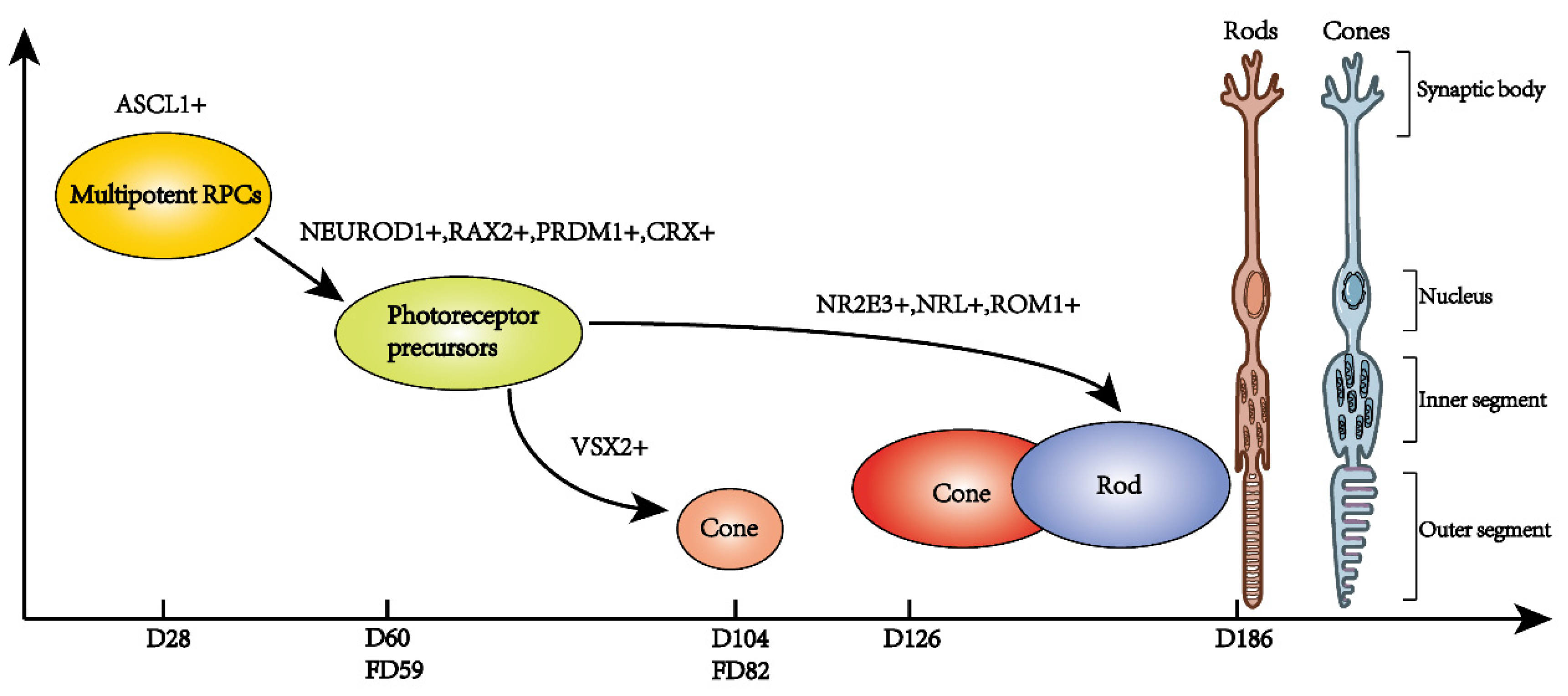
3.2. Human Retinal Organoids Development
3.3. Comparisons between Human Fetal Retina and Retinal Organoids Development
4. Comparisons between Mice Retina and Retinal Organoids Development
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| PCs | photoreceptors |
| BCs | bipolar cells |
| HCs | horizontal cells |
| ACs | amacrine cells |
| RGCs | retinal ganglion cells |
| ONL | outer nuclear layer |
| OPL | outer plexiform layer |
| INL | inner nuclear layer |
| IPL | inner plexiform layer |
| GCL | ganglion cell layer |
| ipRGCs | intrinsically photosensitive retinal ganglion cells |
| RPCs | retinal progenitor cells |
| ED | embryonic day |
| PD | postnatal day |
| LHX4 | LIM homeobox protein 4 |
| Rxrg | retinoic acid receptor RXR-gamma |
| Otx2 | orthodenticle homeobox 2 |
| SLC7A3 | solute carrier family 7 member 3 |
| SLC7A5 | solute carrier family 7 member 5 |
| Thrb | thyroid hormone receptor beta |
| Nrl | neural retina leucine zipper |
| t-SNE | t-distributed stochastic neighbor embedding |
| Crx | cone-rod homeobox |
| BEAM | branched expression analysis modeling |
| Casz1 | castor zinc finger 1 |
| Meis2 | Meis homeobox 2 |
| TFs | transcription factors |
| Prdm1 | PR domain containing 1, with ZNF domain |
| Neurod1 | neurogenic differentiation 1 |
| Ldhb | lactate dehydrogenase B |
| Ldha | lactate dehydrogenase A |
| Neurog2 | neurogenin 2 |
| Atoh7 | atonal bHLH transcription factor 7 |
| Olig2 | oligodendrocyte transcription factor 2 |
| NFI | nuclear factor I |
| Pax6 | paired box 6 |
| Rax | retina and anterior neural fold homeobox |
| Vsx2 | visual system homeobox 2 |
| Notch1 | notch receptor 1 |
| Fgf15 | fibroblast growth factor 15 |
| Cdc20 | cell division cycle 20 |
| Crym | crystallin, mu |
| Rbp3 | retinol binding protein 3, interstitial |
| PCW | weeks after conception |
| EYS | eyes shut homolog |
| SCENIC | single-cell regulatory network inference and clustering |
| RHO | rhodopsin |
| SAG | S-antigen visual arrestin |
| GNGT1 | G protein subunit gamma transducin 1 |
| CNGA1 | cyclic nucleotide gated channel subunit alpha 1 |
| MDK | midkine |
| PTN | pleiotrophin |
| PTPRZ1 | receptor protein tyrosine phosphatase type Z |
| hiPSCs | human induced pluripotent stem cells |
| hESCs | human embryonic stem cells |
| ASCL1 | achaete-scute family bHLH transcription factor 1 |
| NGN2 | NEUROG 2 |
| INSM1 | INSM transcriptional repressor 1 |
| BHLHE22 | basic helix-loop-helix family member e22 |
| ISLR2 | immunoglobulin superfamily containing leucine rich repeat 2 |
| CCND1 | cyclin D1 |
| FOXN4 | forkhead box N4 |
| NR2E3 | nuclear receptor subfamily 2 group E member 3 |
| HES1 | hes family bHLH transcription factor 1 |
| HMGA1 | high mobility group AT-hook 1 |
| BAZ2B | bromodomain adjacent to zinc finger domain 2B |
| MECOM | MDS1 and EVI1 complex locus |
| RCVRN | recoverin |
| OPN1MW | opsin 1, medium wave sensitive |
| OPN1LW | opsin 1, long wave sensitive |
| OPN1SW | opsin 1, short wave sensitive |
| FD | fetal day |
| DLL3 | delta like canonical Notch ligand 3 |
| NEFM | neurofilament medium |
| GAD1 | glutamate decarboxylase 1 |
| TFAP2A | transcription factor AP-2 alpha |
| INSR | insulin receptor |
| CALM2 | calmodulin 2 |
| PRPH | peripherin |
| Btg2 | BTG anti-proliferation factor 2 |
| Isl2 | insulin related protein 2 |
| LMO4 | LIM domain only 4 |
| CYP26A1 | cytochrome P450 family 26 subfamily A member 1 |
| DIO2 | iodothyronine deiodinase 2 |
| CDKN1A | cyclin dependent kinase inhibitor 1A |
| ANXA2 | annexin A2 |
| CTGF | cellular communication network factor 2 |
| FGF19 | fibroblast growth factor 19 |
| SALL3 | spalt like transcription factor 3 |
| DC7 | VPS29 retromer complex component |
| LHX9 | LIM homeobox 9 |
| NHLH1 | nescient helix-loop-helix 1 |
References
- Hoon, M.; Okawa, H.; Della Santina, L.; Wong, R.O. Functional architecture of the retina: Development and disease. Prog. Retin. Eye Res. 2014, 42, 44–84. [Google Scholar] [CrossRef]
- Mitchell, D.M.; Stevens, C.B.; Frey, R.A.; Hunter, S.S.; Ashino, R.; Kawamura, S.; Stenkamp, D.L. Retinoic Acid Signaling Regulates Differential Expression of the Tandemly-Duplicated Long Wavelength-Sensitive Cone Opsin Genes in Zebrafish. PLoS Genet. 2015, 11, e1005483. [Google Scholar] [CrossRef] [PubMed]
- Cook, T.; Desplan, C. Photoreceptor subtype specification: From flies to humans. Semin. Cell Dev. Biol. 2001, 12, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.M.; Chen, S.K.; Hattar, S. Intrinsically photosensitive retinal ganglion cells: Many subtypes, diverse functions. Trends Neurosci. 2011, 34, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Lazzerini Ospri, L.; Prusky, G.; Hattar, S. Mood, the Circadian System, and Melanopsin Retinal Ganglion Cells. Annu. Rev. Neurosci. 2017, 40, 539–556. [Google Scholar] [CrossRef] [PubMed]
- Do, M.T.H.; Yau, K.W. Intrinsically photosensitive retinal ganglion cells. Physiol. Rev. 2010, 90, 1547–1581. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.A.; Guido, W. Shedding light on class-specific wiring: Development of intrinsically photosensitive retinal ganglion cell circuitry. Mol. Neurobiol. 2011, 44, 321–329. [Google Scholar] [CrossRef][Green Version]
- Quattrochi, L.E.; Stabio, M.E.; Kim, I.; Ilardi, M.C.; Michelle Fogerson, P.; Leyrer, M.L.; Berson, D.M. The M6 cell: A small-field bistratified photosensitive retinal ganglion cell. J. Comp. Neurol. 2019, 527, 297–311. [Google Scholar] [CrossRef]
- Corso-Díaz, X.; Jaeger, C.; Chaitankar, V.; Swaroop, A. Epigenetic control of gene regulation during development and disease: A view from the retina. Prog. Retin. Eye Res. 2018, 65, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Hartong, D.T.; Berson, E.L.; Dryja, T.P. Retinitis pigmentosa. Lancet 2006, 368, 1795–1809. [Google Scholar] [CrossRef]
- Molday, R.S.; Zhang, K. Defective lipid transport and biosynthesis in recessive and dominant Stargardt macular degeneration. Prog. Lipid. Res. 2010, 49, 476–492. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Falk, M.J.; Zhang, Q.; Nakamaru-Ogiso, E.; Kannabiran, C.; Fonseca-Kelly, Z.; Chakarova, C.; Audo, I.; Mackay, D.S.; Zeitz, C.; Borman, A.D.; et al. NMNAT1 mutations cause Leber congenital amaurosis. Nat. Genet. 2012, 44, 1040–1045. [Google Scholar] [CrossRef]
- Roosing, S.; Thiadens, A.A.; Hoyng, C.B.; Klaver, C.C.; den Hollander, A.I.; Cremers, F.P. Causes and consequences of inherited cone disorders. Prog. Retin. Eye Res. 2014, 42, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Aldiri, I.; Xu, B.; Wang, L.; Chen, X.; Hiler, D.; Griffiths, L.; Valentine, M.; Shirinifard, A.; Thiagarajan, S.; Sablauer, A.; et al. The Dynamic Epigenetic Landscape of the Retina During Development, Reprogramming, and Tumorigenesis. Neuron 2017, 94, 550–568. [Google Scholar] [CrossRef]
- Buono, L.; Martinez-Morales, J.R. Retina Development in Vertebrates: Systems Biology Approaches to Understanding Genetic Programs: On the Contribution of Next-Generation Sequencing Methods to the Characterization of the Regulatory Networks Controlling Vertebrate Eye Development. Bioessays 2020, 42, e1900187. [Google Scholar] [CrossRef] [PubMed]
- Dharmat, R.; Kim, S.; Li, Y.; Chen, R. Single-Cell Capture, RNA-seq, and Transcriptome Analysis from the Neural Retina. Methods Mol. Biol. 2020, 2092, 159–186. [Google Scholar]
- Heavner, W.; Pevny, L. Eye development and retinogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008391. [Google Scholar] [CrossRef]
- Trimarchi, J.M.; Stadler, M.B.; Roska, B.; Billings, N.; Sun, B.; Bartch, B.; Cepko, C.L. Molecular heterogeneity of developing retinal ganglion and amacrine cells revealed through single cell gene expression profiling. J. Comp. Neurol. 2007, 502, 1047–1065. [Google Scholar] [CrossRef] [PubMed]
- Trimarchi, J.M.; Stadler, M.B.; Cepko, C.L. Individual retinal progenitor cells display extensive heterogeneity of gene expression. PLoS ONE 2008, 3, e1588. [Google Scholar] [CrossRef]
- Buenaventura, D.F.; Corseri, A.; Emerson, M.M. Identification of Genes With Enriched Expression in Early Developing Mouse Cone Photoreceptors. Invest. Ophthalmol. Vis. Sci. 2019, 60, 2787–2799. [Google Scholar] [CrossRef]
- Eldred, K.C.; Hadyniak, S.E.; Hussey, K.A.; Brenerman, B.; Zhang, P.W.; Chamling, X.; Sluch, V.M.; Welsbie, D.S.; Hattar, S.; Taylor, J.; et al. Thyroid hormone signaling specifies cone subtypes in human retinal organoids. Science 2018, 362. [Google Scholar] [CrossRef]
- Mo, A.; Luo, C.; Davis, F.P.; Mukamel, E.A.; Henry, G.L.; Nery, J.R.; Urich, M.A.; Picard, S.; Lister, R.; Eddy, S.R.; et al. Epigenomic landscapes of retinal rods and cones. Elife 2016, 5, e11613. [Google Scholar] [CrossRef] [PubMed]
- Welby, E.; Lakowski, J.; Di Foggia, V.; Budinger, D.; Gonzalez-Cordero, A.; Lun, A.T.; Epstein, M.; Patel, A.; Cuevas, E.; Kruczek, K.; et al. Isolation and Comparative Transcriptome Analysis of Human Fetal and iPSC-Derived Cone Photoreceptor Cells. Stem Cell Reports 2017, 9, 1898–1915. [Google Scholar] [CrossRef] [PubMed]
- Lo Giudice, Q.; Leleu, M.; La Manno, G.; Fabre, P.J. Single-cell transcriptional logic of cell-fate specification and axon guidance in early-born retinal neurons. Development 2019, 146, dev178103. [Google Scholar] [CrossRef]
- Clark, B.S.; Stein-O’Brien, G.L.; Shiau, F.; Cannon, G.H.; Davis-Marcisak, E.; Sherman, T.; Santiago, C.P.; Hoang, T.V.; Rajaii, F.; James-Esposito, R.E.; et al. Single-Cell RNA-Seq Analysis of Retinal Development Identifies NFI Factors as Regulating Mitotic Exit and Late-Born Cell Specification. Neuron 2019, 102, 1111–1126. [Google Scholar] [CrossRef] [PubMed]
- Mizeracka, K.; Trimarchi, J.M.; Stadler, M.B.; Cepko, C.L. Analysis of gene expression in wild-type and Notch1 mutant retinal cells by single cell profiling. Dev. Dyn. 2013, 242, 1147–1159. [Google Scholar] [CrossRef] [PubMed]
- Lutty, G.A.; McLeod, D.S. Development of the hyaloid, choroidal and retinal vasculatures in the fetal human eye. Prog. Retin. Eye Res. 2018, 62, 58–76. [Google Scholar] [CrossRef] [PubMed]
- Mellough, C.B.; Bauer, R.; Collin, J.; Dorgau, B.; Zerti, D.; Dolan, D.W.; Jones, C.M.; Izuogu, O.G.; Yu, M.; Hallam, D.; et al. An integrated transcriptional analysis of the developing human retina. Development 2019, 146, dev169474. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, X.; Hu, B.; Mao, Y.; Chen, Y.; Yan, L.; Yong, J.; Dong, J.; Wei, Y.; Wang, W.; et al. Dissecting the transcriptome landscape of the human fetal neural retina and retinal pigment epithelium by single-cell RNA-seq analysis. PLoS Biol. 2019, 17, e3000365. [Google Scholar] [CrossRef] [PubMed]
- Collin, J.; Queen, R.; Zerti, D.; Dorgau, B.; Hussain, R.; Coxhead, J.; Cockell, S.; Lako, M. Deconstructing Retinal Organoids: Single Cell RNA-Seq Reveals the Cellular Components of Human Pluripotent Stem Cell-Derived Retina. Stem Cells 2019, 37, 593–598. [Google Scholar] [CrossRef]
- Mao, X.; An, Q.; Xi, H.; Yang, X.J.; Zhang, X.; Yuan, S.; Wang, J.; Hu, Y.; Liu, Q.; Fan, G. Single-Cell RNA Sequencing of hESC-Derived 3D Retinal Organoids Reveals Novel Genes Regulating RPC Commitment in Early Human Retinogenesis. Stem Cell Rep. 2019, 13, 747–760. [Google Scholar] [CrossRef]
- Bringmann, A.; Syrbe, S.; Görner, K.; Kacza, J.; Francke, M.; Wiedemann, P.; Reichenbach, A. The primate fovea: Structure, function and development. Prog. Retin. Eye Res. 2018, 66, 49–84. [Google Scholar] [CrossRef]
- Provis, J.M.; Dubis, A.M.; Maddess, T.; Carroll, J. Adaptation of the central retina for high acuity vision: Cones, the fovea and the avascular zone. Prog. Retin. Eye Res. 2013, 35, 63–81. [Google Scholar] [CrossRef]
- Provis, J.M.; Hendrickson, A.E. The foveal avascular region of developing human retina. Arch. Ophthalmol. 2008, 126, 507–511. [Google Scholar] [CrossRef]
- Hendrickson, A.; Possin, D.; Vajzovic, L.; Toth, C.A. Histologic development of the human fovea from midgestation to maturity. Am. J. Ophthalmol. 2012, 154, 767–778. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, S.; Cepko, C.L. Fgf8 Expression and Degradation of Retinoic Acid Are Required for Patterning a High-Acuity Area in the Retina. Dev. Cell 2017, 42, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Knoblich, J.A. Organogenesis in a dish: Modeling development and disease using organoid technologies. Science 2014, 345, 1247125. [Google Scholar] [CrossRef]
- Van Leeuwen, E.M.; Emri, E.; Merle, B.M.; Colijn, J.M.; Kersten, E.; Cougnard-Gregoire, A.; Dammeier, S.; Meester-Smoor, M.; Pool, F.M.; de Jong, E.K.; et al. A new perspective on lipid research in age-related macular degeneration. Prog. Retin. Eye Res 2018, 67, 56–86. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lowe, A.; Dharmat, R.; Lee, S.; Owen, L.A.; Wang, J.; Shakoor, A.; Li, Y.; Morgan, D.J.; Hejazi, A.A.; et al. Generation, transcriptome profiling, and functional validation of cone-rich human retinal organoids. Proc. Natl. Acad. Sci. USA 2019, 116, 10824–10833. [Google Scholar] [CrossRef]
- Sridhar, A.; Hoshino, A.; Finkbeiner, C.R.; Chitsazan, A.; Dai, L.; Haugan, A.K.; Eschenbacher, K.M.; Jackson, D.L.; Trapnell, C.; Bermingham-McDonogh, O.; et al. Single-Cell Transcriptomic Comparison of Human Fetal Retina, hPSC-Derived Retinal Organoids, and Long-Term Retinal Cultures. Cell Rep. 2020, 30, 1644–1659.e4. [Google Scholar] [CrossRef]
- Wang, S.; Poli, S.; Liang, X.; Peng, G.H. Longitudinal single-cell RNA-seq of hESCs-derived retinal organoids. Sci. China Life Sci. 2021, 1–16. [Google Scholar] [CrossRef]
- Lu, Y.; Shiau, F.; Yi, W.; Lu, S.; Wu, Q.; Pearson, J.D.; Kallman, A.; Zhong, S.; Hoang, T.; Zuo, Z.; et al. Single-Cell Analysis of Human Retina Identifies Evolutionarily Conserved and Species-Specific Mechanisms Controlling Development. Dev. Cell 2020, 53, 473–491. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Retinal Subtypes | Gene Markers | |
|---|---|---|
| PCs | Cones | Rxrg, Otx2, Lhx4 *, Gngt2, Gnb3, Opn1sw, FABP7 ** |
| Rods | Blimp1, Crx, Otx2, Nrl, Math3, Rbp3, Rax, Epha8, Neurod1, Nr2e3, Rom1, Rbp3, Rhodopsin **, ABCR **, Recoverin ** | |
| ACs | Tfap2a *, Tfap2b *, Gad1, Glyt1, Onecut2, Prox1, Dlx1, Pax6, Pcdh17, Pou3f1 | |
| BCs | Og9x, Lhx3, Car8, Car10, Nfasc, Otx2 **, Lhx4 | |
| HCs | Onecut1, Lhx1, Onecut2, Prox1 | |
| RGCs | Sox11, Atoh7, NF68, Ebf3, Isl1, Pou4f2, Pou6f2, Elavl4, Pou4f1, Islr2, Syt4, Ebf1/3, L1cam, Brn3b | |
| Müller glia | Oaz1, Pebp1, Apoe, clusterin, Sox2, μ-crystallin, Dkk3 | |
| RPCs | Primary RPCs | Sox2, Fos, Hes1, Pax6, Vsx2, Lhx2, Ccnd1, Cdk4 |
| Neurogenic RPCs | Atoh7, Olig2, Neurog2, Sox11, Onecut1/2, Dlx1/2, Prdm1, Otx2, Ascl1, Hes6 | |
| Retinal Subtypes | Gene Markers | |
|---|---|---|
| PCs | Cones | ARR3, GUCA1C, PDE6C, PDE6H, ISL2, ATOH7, THRB, LMO4, SALL3, DC7, CRX, OPN1LW, OPN1SW, OPN1MW |
| Rods | GNGT1, CNGB1, GNAT1, NRL, NR2E3, PDE6A, RHO, PDE6B, HKR1, CRX, ROM1, SLC24A1 | |
| ACs | MEIS2, GAD1, GAD2, TFAP2A, PAX6, SLC6A1, ATP1B1 | |
| BCs | VSX1, VSX2, CHN2, SCN3A, TRPM1, LRTM1, TMEM215, PLXDC1, NETO1, CA10, ST18, SLC4A10, KCNMA1, ISL1, GRM6, OTX2 | |
| HCs | ONECUT1, ONECUT2, ONECUT3, NTRK1, SEPT4, TPM3, NDRG1, SEPT7 | |
| RGCs | HUC/D, GAP43, SNCG, RBPMS NEFL, TUBB3, POU4F1, SLC17A6 | |
| RPEs | SERPINF1, TYR, MITF, RPE65, BEST1, TTR, PMEL | |
| Müller glia | PLP1, TF, SOX2, GFAP, S100B, SLC1A3, RLBP1, HES1, GLUL, CLU | |
| Microglia cells | CX3CR1, C1QA, C1QB, C1QC | |
| RPCs | Primary RPCs | SFRP2, MKI67, SOX2, HES5, FZD5, ASCL1, PAX6, RAX, NESTIN, TOP2A |
| Neurogenic RPCs | ATOH7, PRDM1, ONECUT1, PROX1, VSX2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, M.; Peng, G.-H. Regulatory Mechanisms of Retinal Photoreceptors Development at Single Cell Resolution. Int. J. Mol. Sci. 2021, 22, 8357. https://doi.org/10.3390/ijms22168357
Zhao M, Peng G-H. Regulatory Mechanisms of Retinal Photoreceptors Development at Single Cell Resolution. International Journal of Molecular Sciences. 2021; 22(16):8357. https://doi.org/10.3390/ijms22168357
Chicago/Turabian StyleZhao, Meng, and Guang-Hua Peng. 2021. "Regulatory Mechanisms of Retinal Photoreceptors Development at Single Cell Resolution" International Journal of Molecular Sciences 22, no. 16: 8357. https://doi.org/10.3390/ijms22168357
APA StyleZhao, M., & Peng, G.-H. (2021). Regulatory Mechanisms of Retinal Photoreceptors Development at Single Cell Resolution. International Journal of Molecular Sciences, 22(16), 8357. https://doi.org/10.3390/ijms22168357