
Are Changes in the Percentage of Specific Leukocyte Subpopulations Associated with Endogenous DNA Damage Levels in Testicular Cancer Patients?

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Results
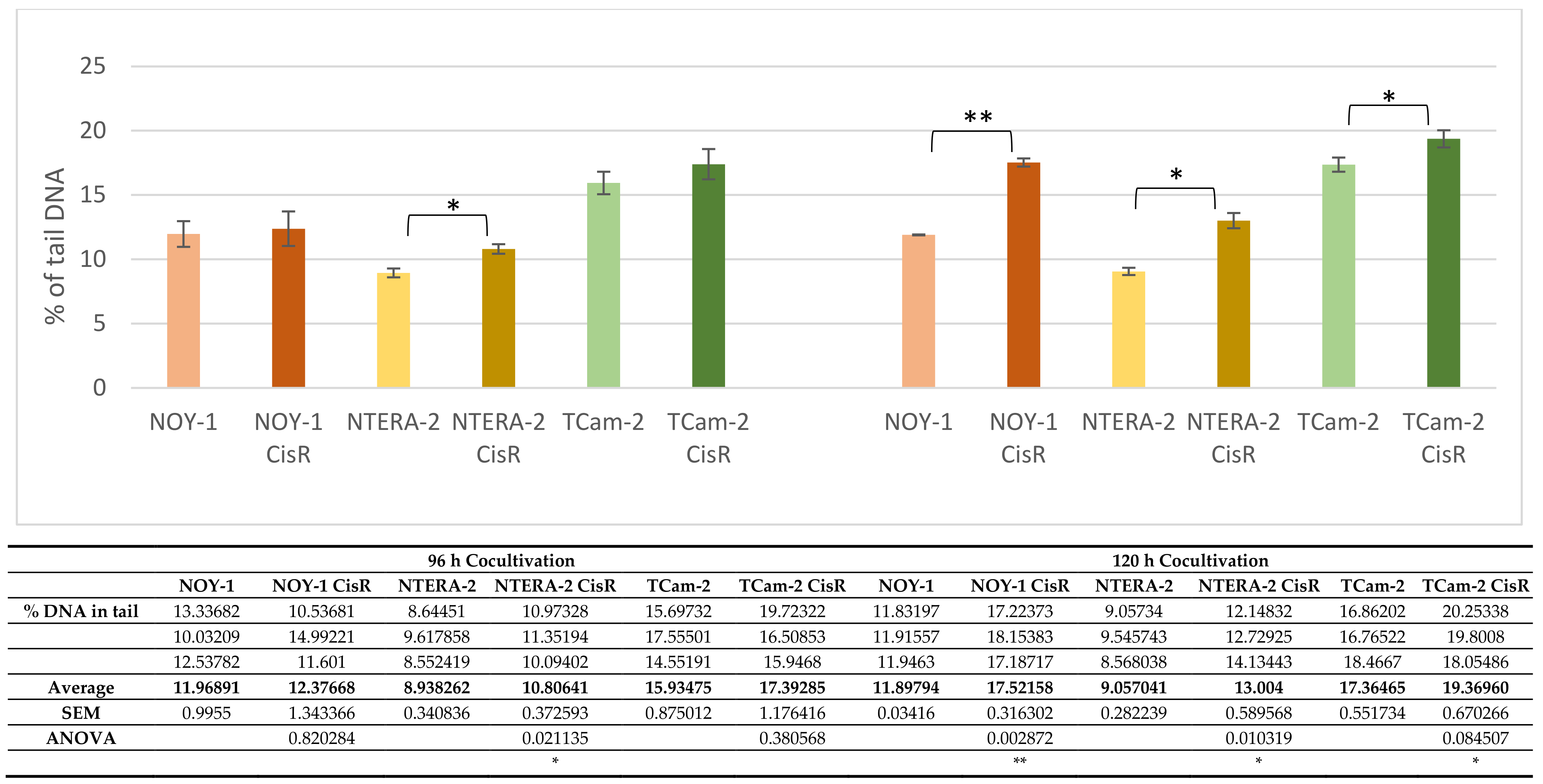
2.1. 2D Cocultivation of GCT Cell Lines with PBMCs
2.2. GCT Subjects’ Analyses
2.2.1. Patient’s Characteristics
2.2.2. Associations among the Clinicopathological Characteristics, Percentage of Different Leukocyte Subsets and Endogenous DNA Damage Level
2.2.3. Multivariate Associations among the Patients’ Clinicopathological Characteristics, Percentage of Different Leukocyte Subpopulations and Endogenous DNA Damage Level
3. Discussion
4. Material and Methods
4.1. Cell Lines
4.2. Patients
4.3. PB Sampling
4.4. PBMC Isolation by Density Gradient Centrifugation
4.5. Cocultivation of GCT Cell Lines with PBMCs
4.6. Determination of the DNA Damage Level in PBMCs
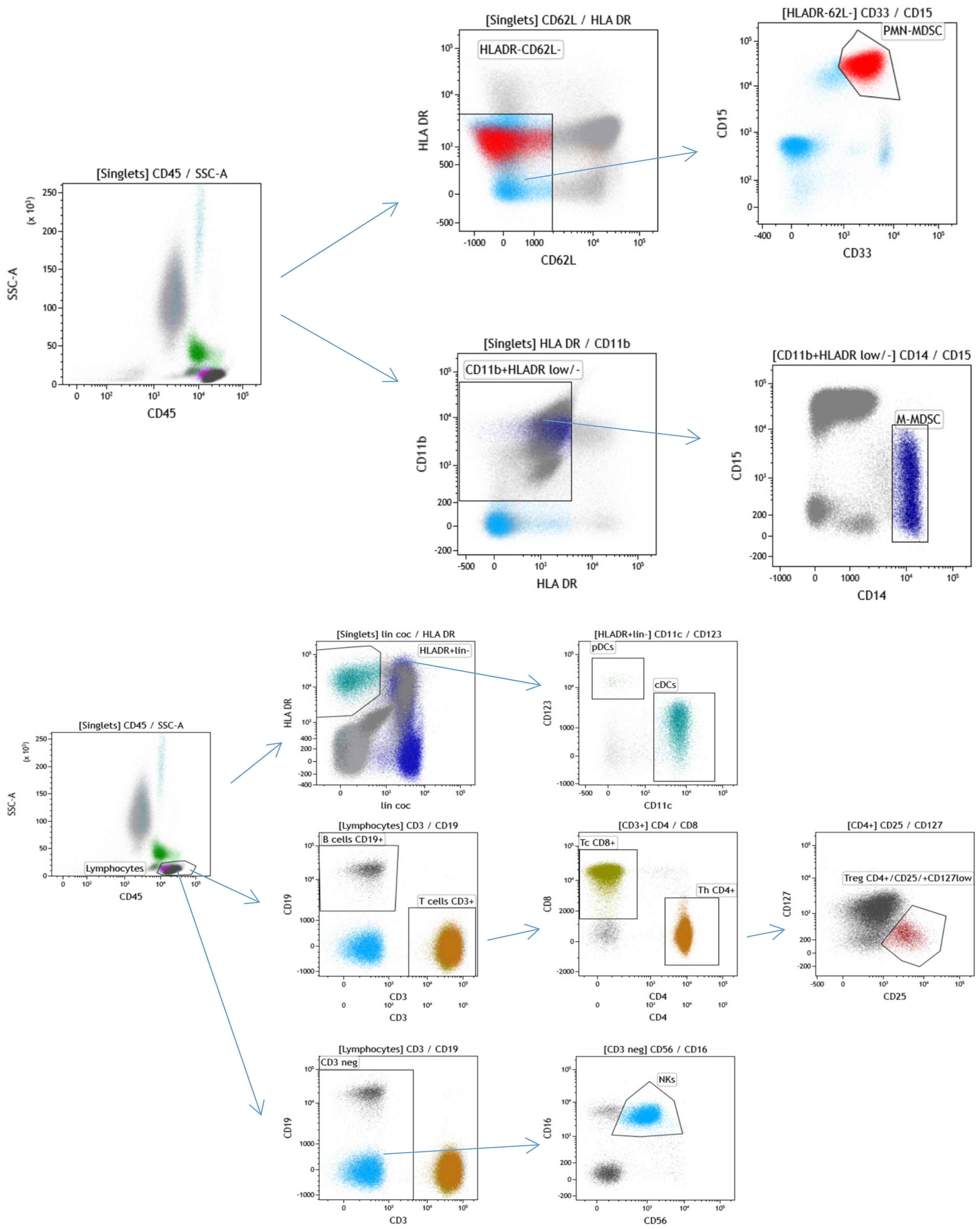
4.7. Immunophenotyping of Leukocytes Subpopulations
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, L.; Albers, P.; Berney, D.M.; Feldman, D.R.; Daugaard, G.; Gilligan, T.; Looijenga, L. Testicular cancer. Nat. Rev. Dis. Prim. 2018, 4, 29. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef]
- Fankhauser, C.D.; Curioni-Fontecedro, A.; Allmann, V.; Beyer, J.; Tischler, V.; Sulser, T.; Moch, H.; Bode, P. Frequent PD-L1 expression in testicular germ cell tumors. Br. J. Cancer 2015, 113, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Kalavska, K.; Kucerova, L.; Schmidtova, S.; Chovanec, M.; Mego, M. Cancer Stem Cell Niche and Immune-Active Tumor Microenvironment in Testicular Germ Cell Tumors. Adv. Exp. Med. Biol. 2020, 1226, 111–121. [Google Scholar] [CrossRef]
- Wang, M.; Fijak, M.; Hossain, H.; Markmann, M.; Nüsing, R.M.; Lochnit, G.; Hartmann, M.F.; Wudy, S.A.; Zhang, L.; Gu, H.; et al. Characterization of the Micro-Environment of the Testis that Shapes the Phenotype and Function of Testicular Macrophages. J. Immunol. 2017, 198, 4327–4340. [Google Scholar] [CrossRef] [PubMed]
- Loveland, K.L.; Klein, B.; Pueschl, D.; Indumathy, S.; Bergmann, M.; Loveland, B.E.; Hedger, M.P.; Schuppe, H.-C. Cytokines in Male Fertility and Reproductive Pathologies: Immunoregulation and Beyond. Front. Endocrinol. 2017, 8, 307. [Google Scholar] [CrossRef]
- Siska, P.J.; Johnpulle, R.A.N.; Zhou, A.; Bordeaux, J.; Kim, J.Y.; Dabbas, B.; Dakappagari, N.; Rathmell, J.C.; Rathmell, W.K.; Morgans, A.K.; et al. Deep exploration of the immune infiltrate and outcome prediction in testicular cancer by quantitative multiplexed immunohistochemistry and gene expression profiling. OncoImmunology 2017, 6, e1305535. [Google Scholar] [CrossRef]
- Bell, D.A.; Flotte, T.J.; Bhan, A.K. Immunohistochemical characterization of seminoma and its inflammatory cell infiltrate. Hum. Pathol. 1987, 18, 511–520. [Google Scholar] [CrossRef]
- Saint, F.; Leroy, X.; Graziana, J.P.; Moukassa, D.; Gosselin, B.; Biserte, J.; Chopin, D.; Rigot, J.M. Dendritic cell infiltration in a patient with seminomatous germ cell tumor of the testis: Is there a relationship with infertility and tumor stage? J. Urol. 2002, 167, 1643–1647. [Google Scholar] [CrossRef]
- Yakirevich, E.; Lefel, O.; Sova, Y.; Stein, A.; Cohen, O.; Izhak, O.B.; Resnick, M.B. Activated status of tumour-infiltrating lym-phocytes and apoptosis in testicular seminoma. J. Pathol. 2002, 196, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Chovanec, M.; Mego, M.; Cholujova, D.; Gronesova, P.; Miskovska, V.; Sycova-Mila, Z.; Usakova, V.; Svetlovska, D.; Bujdak, P.; Spanik, S.; et al. A cytokine and angiogenic factor (CAF) analysis in plasma in testicular germ cell tumor patients (TGCTs). J. Clin. Oncol. 2015, 33 (Suppl. 15), e15552. [Google Scholar] [CrossRef]
- Pearce, H.; Hutton, P.; Chaudhri, S.; Porfiri, E.; Patel, P.; Viney, R.; Moss, P. Spontaneous CD4+and CD8+T-cell responses directed against cancer testis antigens are present in the peripheral blood of testicular cancer patients. Eur. J. Immunol. 2017, 47, 1232–1242. [Google Scholar] [CrossRef]
- Chovanec, M.; Cierna, Z.; Miskovska, V.; Machalekova, K.; Kalavska, K.; Rejlekova, K.; Svetlovska, D.; Macak, D.; Spanik, S.; Kajo, K.; et al. Systemic immune-inflammation index in germ-cell tumours. Br. J. Cancer 2018, 118, 831–838. [Google Scholar] [CrossRef]
- Cursano, M.C.; Kopf, B.; Scarpi, E.; Menna, C.; Casadei, C.; Schepisi, G.; Lolli, C.; Altavilla, A.; Gallà, V.; Santini, D.; et al. Prognostic Role of Systemic Inflammatory Indexes in Germ Cell Tumors Treated with High-Dose Chemotherapy. Front. Oncol. 2020, 10, 1325. [Google Scholar] [CrossRef]
- Lama-Sherpa, T.; Shevde, L.A. An Emerging Regulatory Role for the Tumor Microenvironment in the DNA Damage Response to Double-Strand Breaks. Mol. Cancer Res. 2019, 18, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Sestakova, Z.; Kalavska, K.; Smolkova, B.; Miskovska, V.; Rejlekova, K.; Sycova-Mila, Z.; Palacka, P.; Obertova, J.; Holickova, A.; Hurbanova, L.; et al. DNA damage measured in blood cells predicts overall and progression-free survival in germ cell tumour patients. Mutat. Res. Toxicol. Environ. Mutagen. 2020, 854–855, 503200. [Google Scholar] [CrossRef]
- Sestakova, Z.; Kalavska, K.; Hurbanova, L.; Jurkovicova, D.; Gursky, J.; Chovanec, M.; Svetlovska, D.; Miskovska, V.; Obertova, J.; Palacka, P.; et al. The prognostic value of DNA damage level in peripheral blood lymphocytes of chemotherapy-naïve patients with germ cell cancer. Oncotarget 2016, 7, 75996–76005. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hapakova, N.; Sestakova, Z.; Holickova, A.; Hurbanova, L.; Miskovska, V.; Chovanec, M.; Rejlekova, K.; Svetlovska, D.; Kalavska, K.; Obertova, J.; et al. High Endogenous DNA Damage Levels Predict Hematological Toxicity in Testicular Germ Cell Tumor Patients Treated with First-Line Chemotherapy. Clin. Genitourin. Cancer 2019, 17, e1020–e1025. [Google Scholar] [CrossRef]
- Mead, G.M.; Stenning, S.P.; Cook, P.; Fossa, S.D.; Horwich, A.; Kaye, S.B.; Oliver, R.T.D.; de Mulder, P.H.M.; de Wit, R.; Coppin, C.; et al. The International Germ Cell Consensus Classification: A prognostic factor based staging system for metastatic germ cell cancer. J. Clin. Oncol. 1997, 15, 594–603. [Google Scholar]
- Singh, R.; Fazal, Z.; Freemantle, S.J.; Spinella, M.J. Mechanisms of cisplatin sensitivity and resistance in testicular germ cell tumors. Cancer Drug Resist. 2019, 2, 580–594. [Google Scholar] [CrossRef]
- Lobo, J.; Jerónimo, C.; Henrique, R. Cisplatin Resistance in Testicular Germ Cell Tumors: Current Challenges from Various Perspectives. Cancers 2020, 12, 1601. [Google Scholar] [CrossRef] [PubMed]
- Testa, E.; Nardozi, D.; Antinozzi, C.; Faieta, M.; di Cecca, S.; Caggiano, C.; Fukuda, T.; Bonanno, E.; Zhenkun, L.; Maldonado, A.; et al. H2AFX and MDC1 promote maintenance of genomic integrity in male germ cells. J. Cell Sci. 2018, 131, jcs.214411. [Google Scholar] [CrossRef]
- Palla, V.V.; Karaolanis, G.; Katafigiotis, I.; Anastasiou, I.; Patapis, P.; Dimitroulis, D.; Perrea, D. gamma-H2AX: Can it be established as a classical cancer prognostic factor? Tumour Biol. 2017, 39, 1010428317695931. [Google Scholar] [CrossRef]
- Fromm, P.; Papadimitrious, M.; Hsu, J.; Larsen, S.R.; Gibson, J.; Bradstock, K.; Kupresanin, F.; Clark, G.; Hart, D.N. CD16+ Dendritic Cells Are a Unique Myeloid Antigen Presenting Cell Population. Blood 2016, 128, 4897. [Google Scholar] [CrossRef]
- Vivier, E.; Morin, P.; O’Brien, C.; Druker, B.; Schlossman, S.F.; Anderson, P. Tyrosine phosphorylation of the Fc gamma RIII(CD16): Zeta complex in human natural killer cells. Induction by antibody-dependent cytotoxicity but not by natural killing. J. Immunol. 1991, 146, 206–210. [Google Scholar]
- Mandelboim, O.; Malik, P.; Davis, D.; Jo, C.H.; Boyson, J.E.; Strominger, J.L. Human CD16 as a lysis receptor mediating direct natural killer cell cytotoxicity. Proc. Natl. Acad. Sci. USA 1999, 96, 5640–5644. [Google Scholar] [CrossRef]
- Randolph, G.J.; Sanchez-Schmitz, G.; Liebman, R.M.; Schäkel, K. The CD16+ (FcγRIII+) Subset of Human Monocytes Preferentially Becomes Migratory Dendritic Cells in a Model Tissue Setting. J. Exp. Med. 2002, 196, 517–527, Erratum in 2002, 196, 869. [Google Scholar] [CrossRef]
- Nastasi, C.; Mannarino, L.; D’Incalci, M. DNA Damage Response and Immune Defense. Int. J. Mol. Sci. 2020, 21, 7504. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, R.A.; von Andrian, U.H. How Tolerogenic Dendritic Cells Induce Regulatory T Cells. Adv. Immunol. 2010, 108, 111–165. [Google Scholar] [CrossRef]
- Chaudhary, B.; Al Samid, M.A.; Al-Ramadi, B.K.; Elkord, E. Phenotypic alterations, clinical impact and therapeutic potential of regulatory T cells in cancer. Expert Opin. Biol. Ther. 2014, 14, 931–945. [Google Scholar] [CrossRef]
- Nishikawa, H.; Sakaguchi, S. Regulatory T cells in cancer immunotherapy. Curr. Opin. Immunol. 2014, 27, 1–7. [Google Scholar] [CrossRef]
- Ohue, Y.; Nishikawa, H. Regulatory T (Treg) cells in cancer: Can Treg cells be a new therapeutic target? Cancer Sci. 2019, 110, 2080–2089. [Google Scholar] [CrossRef]
- Labani-Motlagh, A.; Ashja-Mahdavi, M.; Loskog, A. The Tumor Microenvironment: A Milieu Hindering and Obstructing Antitumor Immune Responses. Front. Immunol. 2020, 11, 940. [Google Scholar] [CrossRef]
- Melaiu, O.; Lucarini, V.; Cifaldi, L.; Fruci, D. Influence of the Tumor Microenvironment on NK Cell Function in Solid Tumors. Front. Immunol. 2020, 10. [Google Scholar] [CrossRef]
- López-Soto, A.; Huergo-Zapico, L.; Acebes-Huerta, A.; Villa-Alvarez, M.; Gonzalez, S. NKG2D signaling in cancer immunosurveillance. Int. J. Cancer 2014, 136, 1741–1750. [Google Scholar] [CrossRef]
- Alvarez, M.; Simonetta, F.; Baker, J.; Pierini, A.; Wenokur, A.; Morrison, A.R.; Murphy, W.J.; Negrin, R.S. Regulation of murine NK cell exhaustion through the activation of the DNA damage repair pathway. JCI Insight 2019, 4. [Google Scholar] [CrossRef]
- Slyskova, J.; Korenkova, V.; Collins, A.; Prochazka, P.; Vodickova, L.; Svec, J.; Lipska, L.; Levy, M.; Schneiderová, M.; Liška, V.; et al. Functional, Genetic, and Epigenetic Aspects of Base and Nucleotide Excision Repair in Colorectal Carcinomas. Clin. Cancer Res. 2012, 18, 5878–5887. [Google Scholar] [CrossRef]
- Herrera, M.; Dominguez, G.; Garcia, J.M.; Peña, C.; Jimenez, C.; Silva, J.; Garcia, V.; Gomez, I.; Diaz, R.; Martinez, R.D.; et al. Differences in Repair of DNA Cross-links between Lymphocytes and Epithelial Tumor Cells from Colon Cancer Patients Measured In vitro with the Comet Assay. Clin. Cancer Res. 2009, 15, 5466–5472. [Google Scholar] [CrossRef]
- Chang, D.H.; Rutledge, J.R.; Patel, A.A.; Heerdt, B.G.; Augenlicht, L.H.; Korst, R.J. The Effect of Lung Cancer on Cytokine Expression in Peripheral Blood Mononuclear Cells. PLoS ONE 2013, 8, e64456. [Google Scholar] [CrossRef] [PubMed]
- Babini, G.; Morini, J.; Barbieri, S.; Baiocco, G.; Ivaldi, G.B.; Liotta, M.; de Fatis, P.T.; Ottolenghi, A. A Co-culture Method to Investigate the Crosstalk Between X-ray Irradiated Caco-2 Cells and PBMC. J. Vis. Exp. 2018, e56908. [Google Scholar] [CrossRef]
- Prieto-García, E.; Díaz-García, C.; Agudo-López, A.; Pardo-Marqués, V.; García-Consuegra, I.; Asensio-Peña, S.; Alonso-Riaño, M.; Pérez, C.; Gómez, C.; Adeva, J.; et al. Tumor–Stromal Interactions in a Co-Culture Model of Human Pancreatic Adenocarcinoma Cells and Fibroblasts and Their Connection with Tumor Spread. Biomedicines 2021, 9, 364. [Google Scholar] [CrossRef]
- Schmidtova, S.; Kalavska, K.; Gercakova, K.; Cierna, Z.; Miklikova, S.; Smolková, B.; Buocikova, V.; Miskovska, V.; Durinikova, E.; Burikova, M.; et al. Disulfiram Overcomes Cisplatin Resistance in Human Embryonal Carcinoma Cells. Cancers 2019, 11, 1224. [Google Scholar] [CrossRef] [PubMed]
- Schmidtova, S.; Dorssers, L.C.J.; Kalavska, K.; Gillis, A.J.M.; Oosterhuis, J.W.; Stoop, H.; Miklikova, S.; Kozovska, Z.; Burikova, M.; Gercakova, K.; et al. Napabucasin overcomes cisplatin resistance in ovarian germ cell tumor-derived cell line by inhibiting cancer stemness. Cancer Cell Int. 2020, 20, 1–16. [Google Scholar] [CrossRef]
- Edge, S.B.; Compton, C.C. The American Joint Committee on Cancer: The 7th Edition of the AJCC Cancer Staging Manual and the Future of TNM. Ann. Surg. Oncol. 2010, 17, 1471–1474. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variable | N | % |
|---|---|---|
| All patients | 74 | 100.0 |
| Age (years) | ||
| Median (range) | 35 (19–62) | |
| Sex | ||
| Male | 74 | 100.0 |
| Female | 0 | 0.0 |
| Histology | ||
| Seminoma | 20 | 27.0 |
| Nonseminoma | 54 | 73.0 |
| Primary tumor localization | ||
| Testicular | 71 | 91 |
| Extragonadal | 3 | 8.1 |
| IGCCCG risk group | ||
| Good risk | 43 | 58.1 |
| Intermediate risk | 7 | 9.5 |
| Poor risk | 11 | 14.9 |
| Stage IA and IB (adjuvant therapy) | 13 | 17.6 |
| Sites of metastases | ||
| Retroperitoneum | 53 | 71.6 |
| Mediastinum | 10 | 13.5 |
| Lungs | 14 | 18.9 |
| Liver | 6 | 8.1 |
| Brain | 2 | 2.7 |
| Other | 1 | 1.4 |
| Visceral nonpulmonary metastases | 8 | 10.8 |
| No. of metastatic site(s) | ||
| 0 to 1 | 53 | 71.6 |
| >2 | 21 | 28.4 |
| Staging (UICC) | ||
| IA | 2 | 2.7 |
| IB | 11 | 14.9 |
| IS | 5 | 6.8 |
| IIA | 5 | 6.8 |
| IIB | 15 | 20.3 |
| IIC | 6 | 8.1 |
| IIIA | 8 | 10.8 |
| IIIB | 10 | 13.5 |
| IIIC | 12 | 16.2 |
| The DNA Damage Level ≤6.34 >6.34 | |||||
|---|---|---|---|---|---|
| Variable | N | % | N | % | p Value b |
| All patients | 55 | 74.3 | 19 | 25.7 | NA |
| Histology | |||||
| Seminoma | 13 | 65.0 | 7 | 35.0 | 0.37 |
| Nonseminoma | 42 | 77.8 | 12 | 22.2 | |
| Tumor primary | |||||
| Primary GCTs | 54 | 76.1 | 17 | 23.9 | 0.16 |
| Extragonadal GCTs | 1 | 33.3 | 2 | 66.7 | |
| IGCCCG risk group | |||||
| Good risk + adjuvant therapy | 32 | 74.4 | 11 | 25.6 | 0.54 |
| Intermediate + poor risk | 2 | 28.6 | 5 | 71.4 | |
| No. of metastatic site(s) | |||||
| 0 | 14 | 77.8 | 4 | 22.2 | 0.11 |
| 1 to 2 | 35 | 79.5 | 9 | 20.5 | |
| >3 | 6 | 50.0 | 6 | 50.0 | |
| Retroperitoneal lymph node metastases | |||||
| Absent | 17 | 81.0 | 4 | 19.0 | 0.56 |
| Present | 38 | 71.7 | 15 | 28.3 | |
| Mediastinal lymph node metastases | |||||
| Absent | 50 | 78.1 | 14 | 21.9 | 0.11 |
| Present | 5 | 50.0 | 5 | 50.0 | |
| Lung metastases | |||||
| Absent | 46 | 76.7 | 14 | 23.3 | 0.33 |
| Present | 9 | 64.3 | 5 | 35.7 | |
| Liver metastases | |||||
| Absent | 50 | 73.5 | 18 | 26.5 | 1.00 |
| Present | 5 | 83.3 | 1 | 16.7 | |
| Non-pulmonary visceral metastases | |||||
| Absent | 48 | 72.7 | 18 | 27.3 | 0.67 |
| Present | 7 | 87.5 | 1 | 12.5 | |
| S stage a | |||||
| 0 | 21 | 80.8 | 5 | 19.2 | 0.61 |
| 1 | 19 | 76.0 | 6 | 24.0 | |
| 2 | 8 | 61.5 | 5 | 38.5 | |
| 3 | 7 | 70.0 | 3 | 30.0 | |
| Variable | % of Individual Leukocytes Subpopulation | |||||
|---|---|---|---|---|---|---|
| N | Mean | SEM | Median | p Value b | p Value c | |
| Neutrophils percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 55 | 61.7 | 1.9 | 61.1 | 0.462 | |
| endogenous DNA damage level > 6.34 | 19 | 63.9 | 3.2 | 63.4 | ||
| Lymphocytes percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 54 | 27.5 | 1.7 | 26.6 | 0.232 | |
| endogenous DNA damage level > 6.34 | 19 | 23.6 | 2.9 | 20.9 | ||
| Monocytes percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 55 | 9.3 | 0.4 | 9.2 | 0.347 | |
| endogenous DNA damage level > 6.34 | 19 | 10.2 | 0.7 | 9.6 | ||
| Eosinophils percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 55 | 2.5 | 0.4 | 2.7 | 0.334 | |
| endogenous DNA damage level > 6.34 | 19 | 3.0 | 0.6 | 2.7 | ||
| Basophils percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 55 | 0.6 | 0.04 | 0.3 | 0.064 | |
| endogenous DNA damage level > 6.34 | 19 | 0.8 | 0.07 | 0.4 | ||
| B cells percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 55 | 12.6 | 0.7 | 11.8 | 0.00058 | 0.60005 |
| endogenous DNA damage level > 6.34 | 19 | 8.3 | 1.2 | 8.0 | ||
| T cells percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 55 | 72.9 | 1.2 | 73.6 | 0.848 | |
| endogenous DNA damage level > 6.34 | 19 | 71.9 | 2.1 | 74.4 | ||
| T helper cells percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 54 | 45.4 | 1.2 | 45.8 | 0.530 | |
| endogenous DNA damage level > 6.34 | 19 | 43.3 | 2.0 | 43.6 | ||
| T cytotoxic cells percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 55 | 25.6 | 1.0 | 24.5 | 0.669 | |
| endogenous DNA damage level > 6.34 | 19 | 26.2 | 1.6 | 26.9 | ||
| NKT cells percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 53 | 1.7 | 0.4 | 2.4 | 0.057 | |
| endogenous DNA damage level > 6.34 | 19 | 3.1 | 0.6 | 3.3 | ||
| CD4+ NKT cells percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 24 | 0.2 | 0.1 | 0.3 | 0.155 | |
| endogenous DNA damage level > 6.34 | 11 | 0.7 | 0.2 | 0.9 | ||
| CD8+ NKT cells percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 25 | 1.7 | 0.4 | 1.5 | 0.243 | |
| endogenous DNA damage level > 6.34 | 11 | 2.7 | 0.5 | 2.4 | ||
| NK cells percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 55 | 11.3 | 1.1 | 9.6 | 0.008 | 0.00063 |
| endogenous DNA damage level > 6.34 | 19 | 17.3 | 1.9 | 13.6 | ||
| Tregs percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 55 | 3.8 | 0.2 | 3.8 | 0.03937 | 0.01588 |
| endogenous DNA damage level > 6.34 | 19 | 4.5 | 0.3 | 4.4 | ||
| Classical monocytes percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 41 | 87.6 | 1.1 | 87.9 | 0.256 | |
| endogenous DNA damage level > 6.34 | 17 | 85.7 | 1.7 | 86.5 | ||
| Intermediate monocytes percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 24 | 4.7 | 0.5 | 4.3 | 0.546 | |
| endogenous DNA damage level > 6.34 | 11 | 5.2 | 0.8 | 3.7 | ||
| Nonclassical monocytes percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 40 | 4.5 | 0.5 | 4.8 | 0.714 | |
| endogenous DNA damage level > 6.34 | 17 | 5.9 | 0.7 | 4.6 | ||
| Polymorphonuclear leukocytes (PNMs) percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 36 | 1.0 | 0.5 | 0.3 | 0.0753 | |
| endogenous DNA damage level > 6.34 | 15 | 1.1 | 0.8 | 0.2 | ||
| Dendritic cells (cDCs) percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 44 | 0.8 | 0.07 | 0.8 | 0.614 | |
| endogenous DNA damage level > 6.34 | 13 | 1.0 | 0.1 | 0.7 | ||
| Plasmocytoid dendritic cells (pDCs) percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 43 | 0.1 | 0.01 | 0.1 | 0.938 | |
| endogenous DNA damage level > 6.34 | 13 | 0.2 | 0.02 | 0.1 | ||
| CD16+ within DCs percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 32 | 41.5 | 2.8 | 44.4 | 0.00574 | 0.00010 |
| endogenous DNA damage level > 6.34 | 6 | 64.1 | 6.5 | 64.8 | ||
| CD1c+ within DCs percentage | ||||||
| endogenous DNA damage level ≤ 6.34 | 44 | 23.1 | 1.3 | 22.9 | 0.066 | |
| endogenous DNA damage level > 6.34 | 13 | 18.1 | 2.4 | 17.3 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalavska, K.; Sestakova, Z.; Mlcakova, A.; Kozics, K.; Gronesova, P.; Hurbanova, L.; Miskovska, V.; Rejlekova, K.; Svetlovska, D.; Sycova-Mila, Z.; et al. Are Changes in the Percentage of Specific Leukocyte Subpopulations Associated with Endogenous DNA Damage Levels in Testicular Cancer Patients? Int. J. Mol. Sci. 2021, 22, 8281. https://doi.org/10.3390/ijms22158281
Kalavska K, Sestakova Z, Mlcakova A, Kozics K, Gronesova P, Hurbanova L, Miskovska V, Rejlekova K, Svetlovska D, Sycova-Mila Z, et al. Are Changes in the Percentage of Specific Leukocyte Subpopulations Associated with Endogenous DNA Damage Levels in Testicular Cancer Patients? International Journal of Molecular Sciences. 2021; 22(15):8281. https://doi.org/10.3390/ijms22158281
Chicago/Turabian StyleKalavska, Katarina, Zuzana Sestakova, Andrea Mlcakova, Katarína Kozics, Paulina Gronesova, Lenka Hurbanova, Viera Miskovska, Katarina Rejlekova, Daniela Svetlovska, Zuzana Sycova-Mila, and et al. 2021. "Are Changes in the Percentage of Specific Leukocyte Subpopulations Associated with Endogenous DNA Damage Levels in Testicular Cancer Patients?" International Journal of Molecular Sciences 22, no. 15: 8281. https://doi.org/10.3390/ijms22158281
APA StyleKalavska, K., Sestakova, Z., Mlcakova, A., Kozics, K., Gronesova, P., Hurbanova, L., Miskovska, V., Rejlekova, K., Svetlovska, D., Sycova-Mila, Z., Obertova, J., Palacka, P., Mardiak, J., Chovanec, M., Chovanec, M., & Mego, M. (2021). Are Changes in the Percentage of Specific Leukocyte Subpopulations Associated with Endogenous DNA Damage Levels in Testicular Cancer Patients? International Journal of Molecular Sciences, 22(15), 8281. https://doi.org/10.3390/ijms22158281