
Benzo[a]pyrene-Induced Genotoxicity in Rats Is Affected by Co-Exposure to Sudan I by Altering the Expression of Biotransformation Enzymes

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
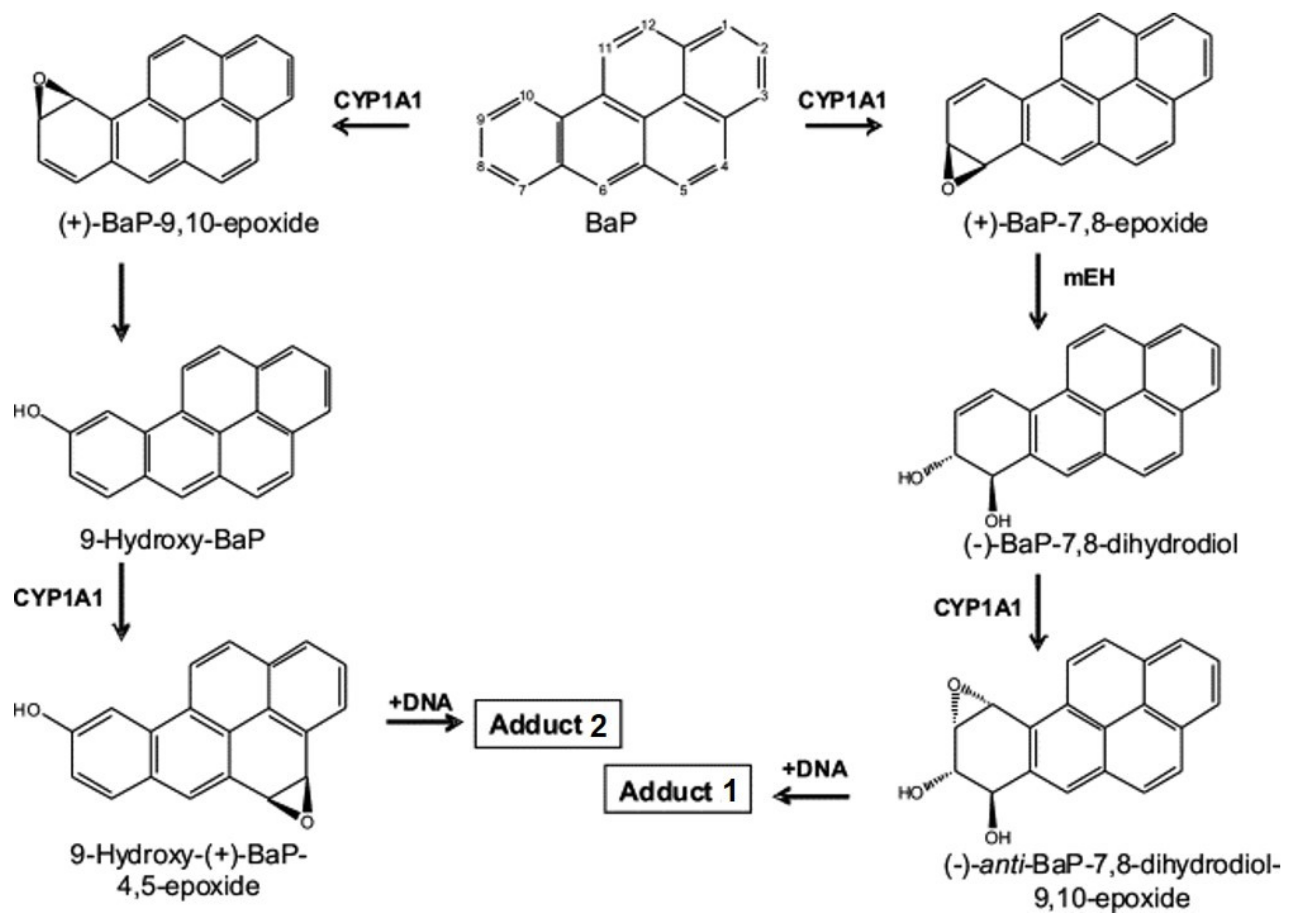
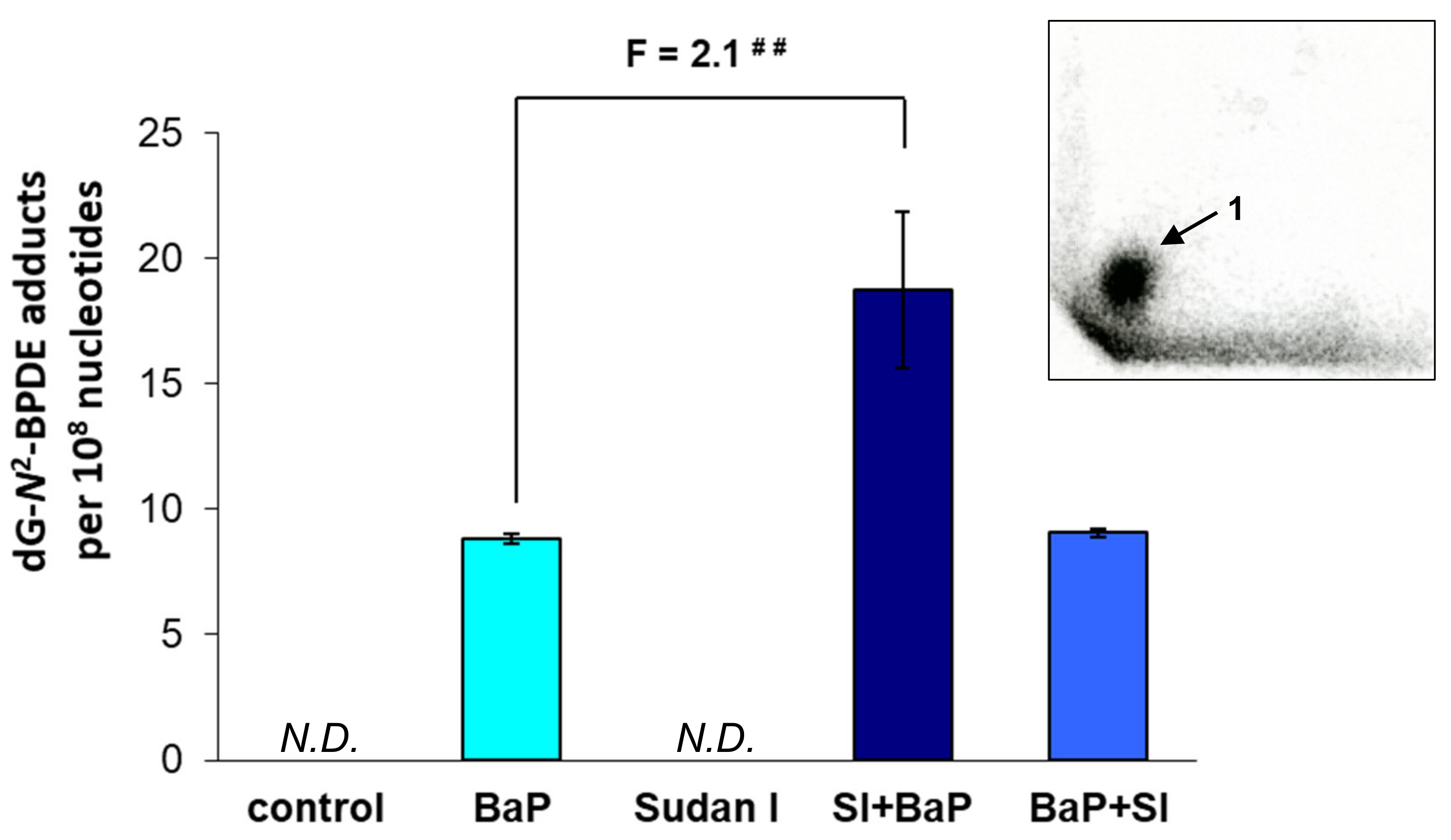
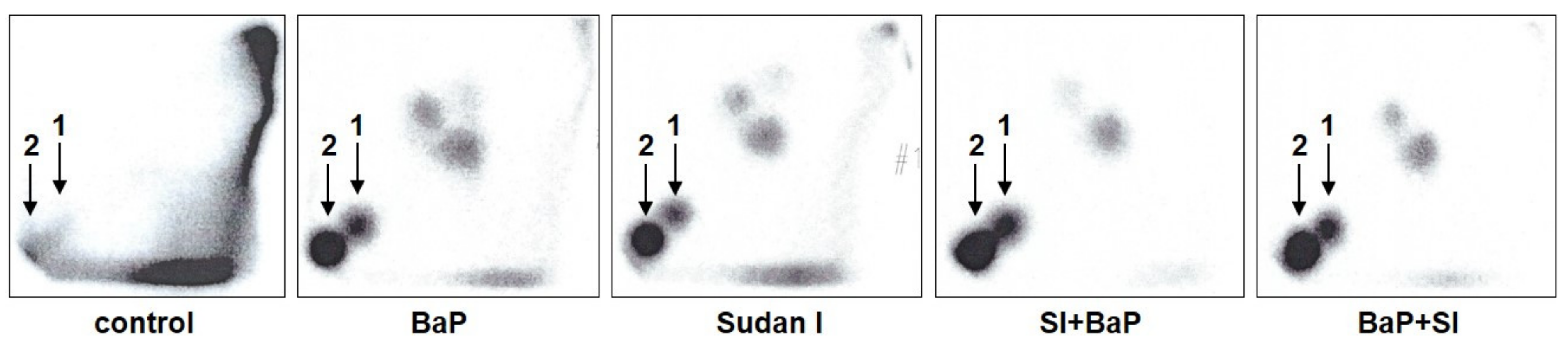
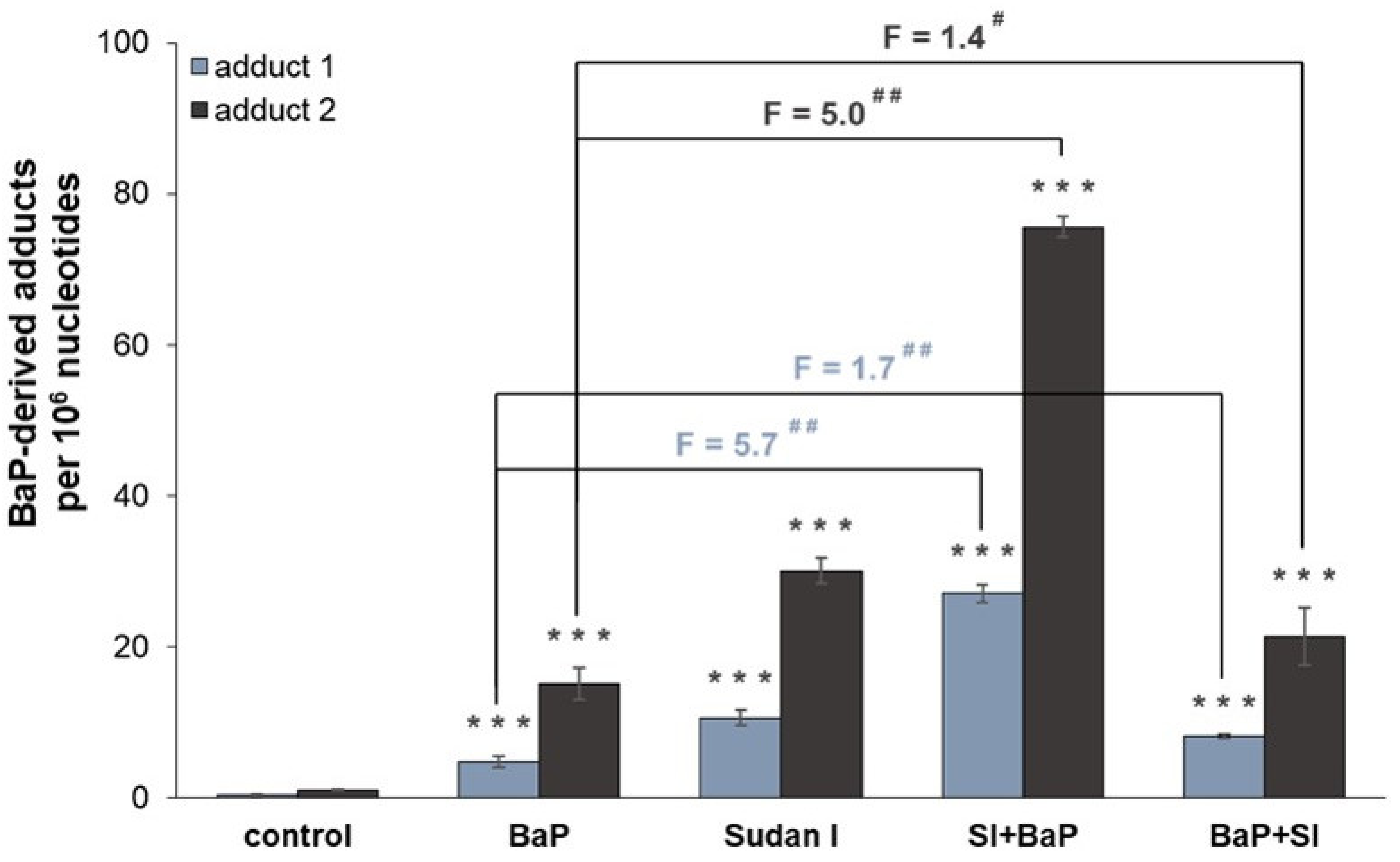
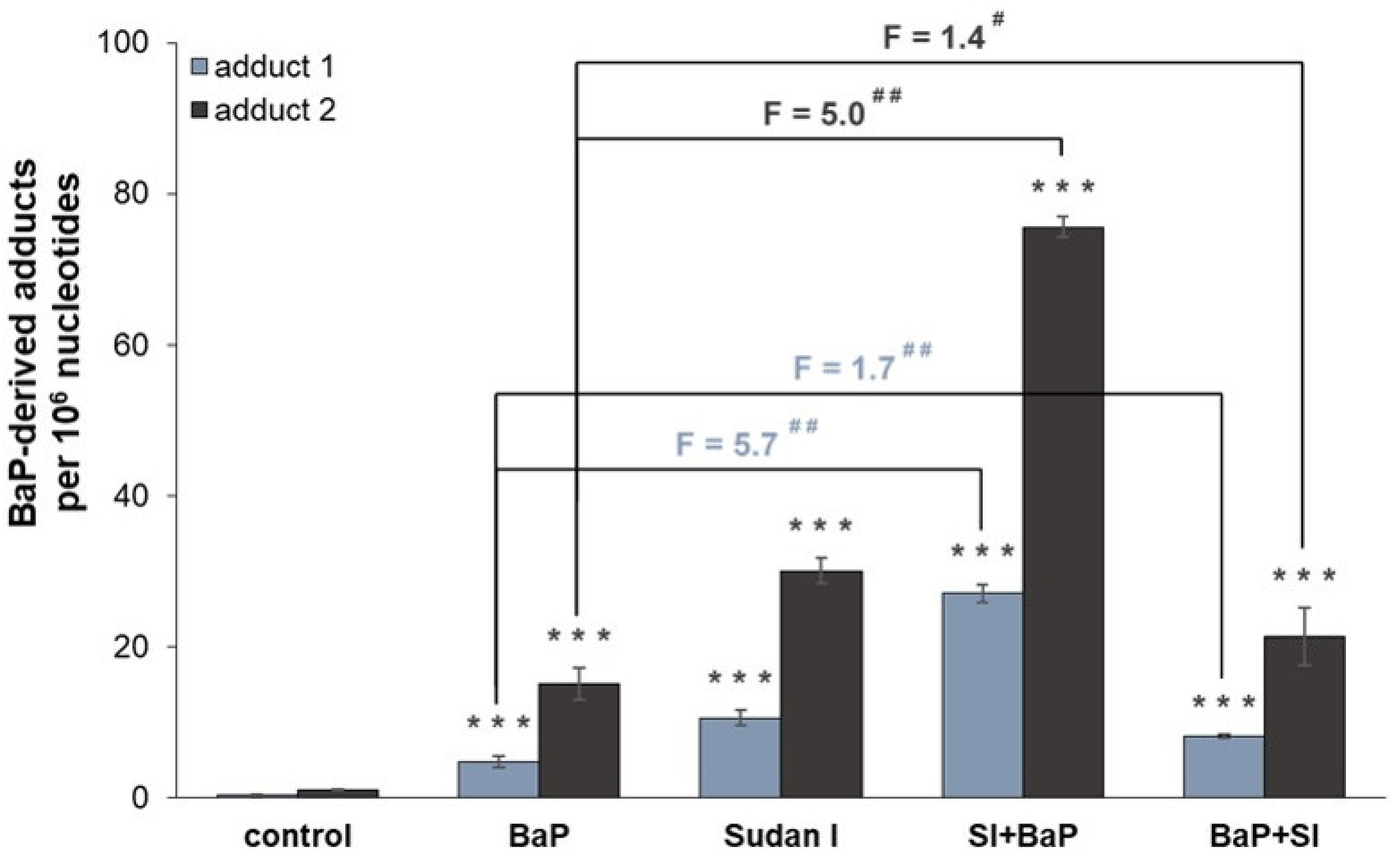
2.1. Formation of BaP-Derived DNA Adducts In Vivo in Rats Exposed to BaP Alone or in Combination with Sudan I
2.2. Formation of BaP-Derived DNA Adducts after Incubation of BaP with Rat Hepatic Microsomes In Vitro
2.3. Protein Expression of CYP1A1, CYP1A2 and CYP1B1 in Livers of Rats Exposed to BaP Alone or in Combination with Sudan I
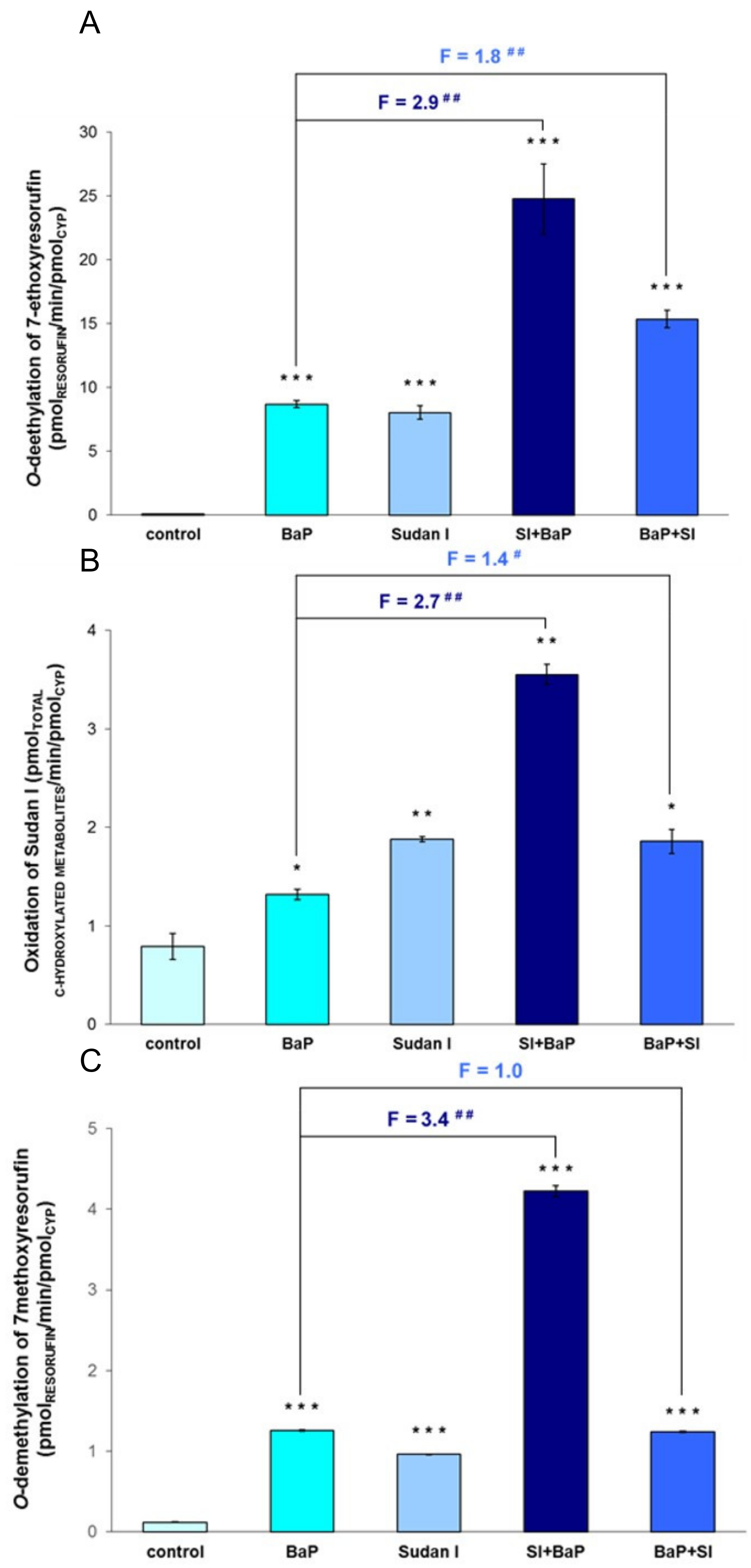
2.4. Enzymatic Activity of CYP1A1, CYP1A2 and CYP1B1 in Livers of Rats Exposed to BaP Alone or in Combination with Sudan I
2.5. Gene Expression of CYP1A1, CYP1A2 and CYP1B1 in Livers of Rats Exposed to BaP Alone or in Combination with Sudan I
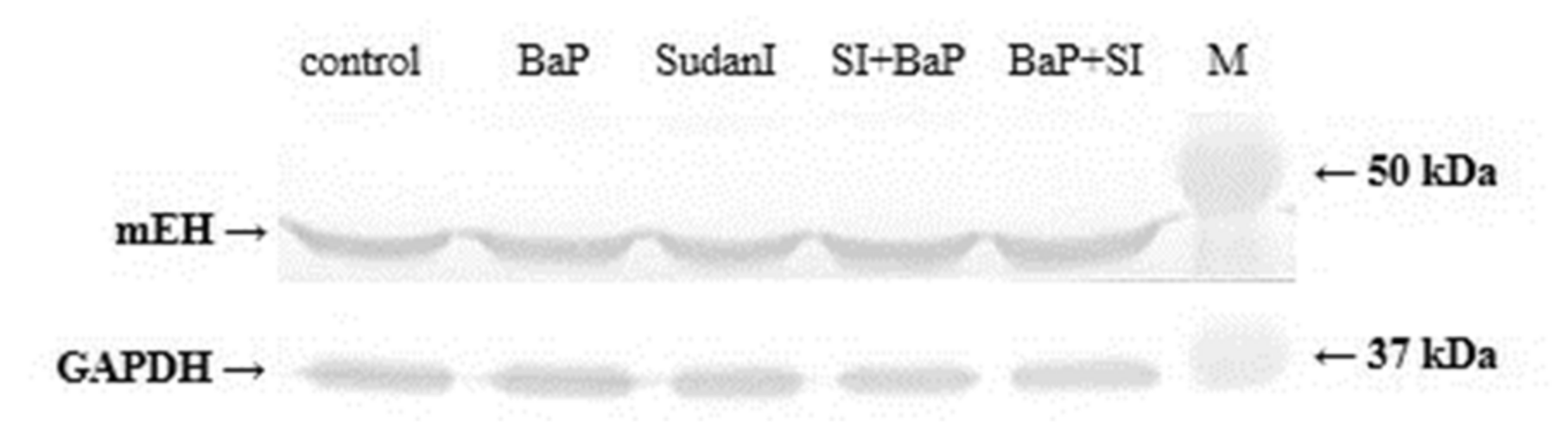
2.6. Protein and Gene Expression of mEH in Livers of Rats Exposed to BaP Alone or in Combination with Sudan I
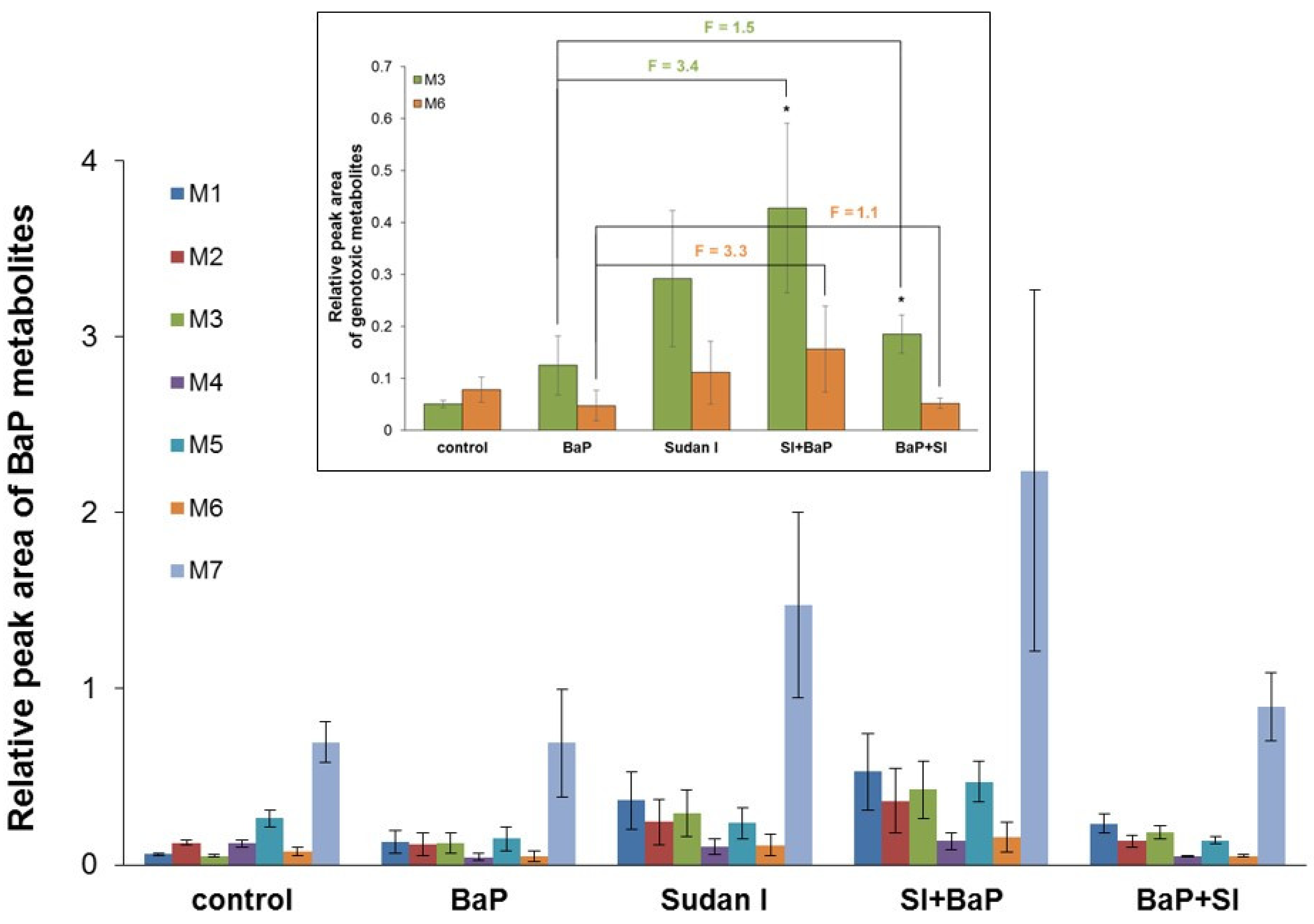
2.7. Metabolism of BaP by Hepatic Microsomes Isolated from Rats Exposed to BaP Alone or in Combinations with Sudan I
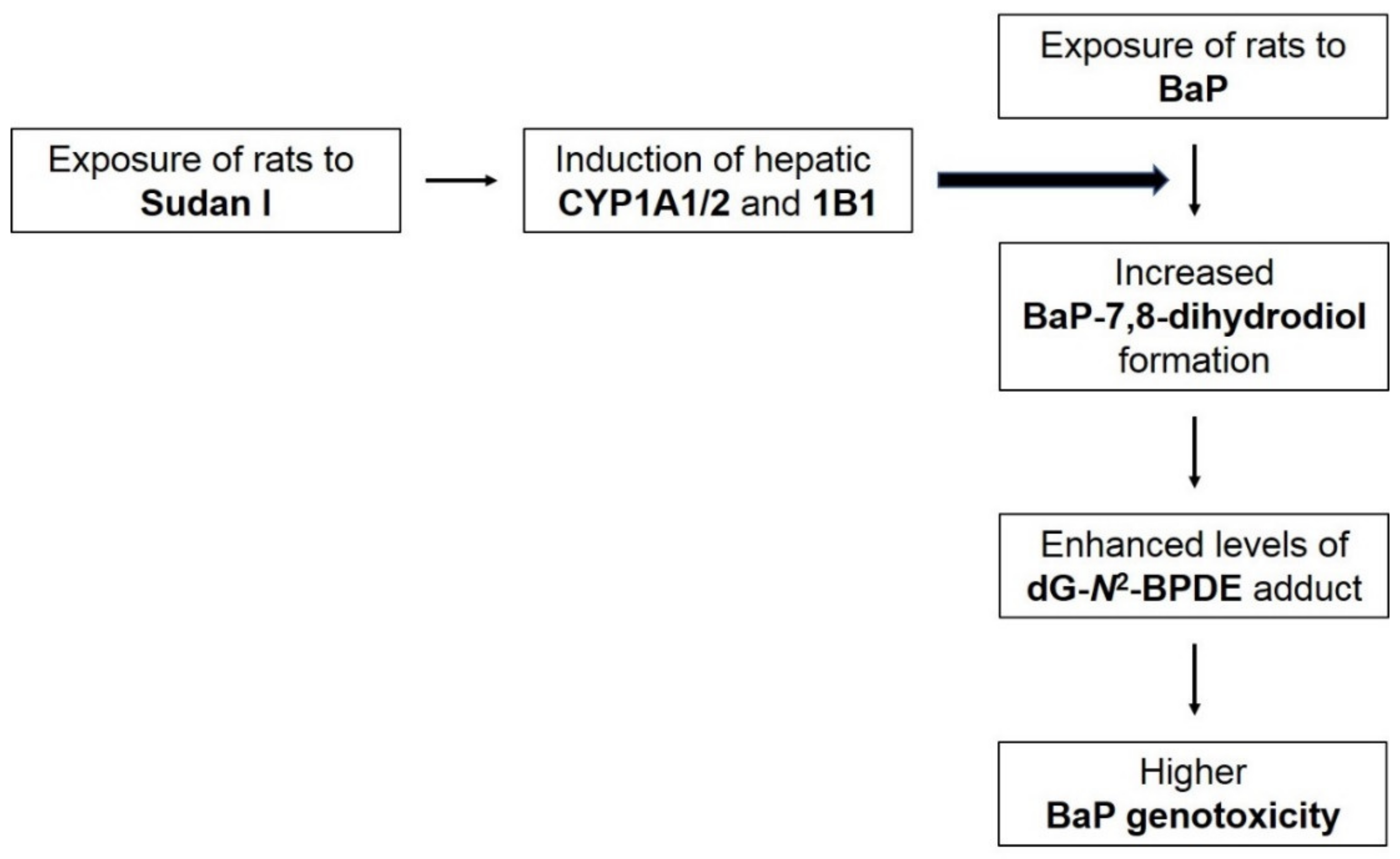
3. Discussion
4. Materials and Methods
4.1. Chemicals and Enzymes
4.2. Treatment of Rats
4.3. BaP-DNA Adduct Detection by 32P-Postlabelling Analysis
4.4. Preparation of Microsomes
4.5. Microsomal Incubations for BaP-DNA Adduct Formation
4.6. Western Blot Analysis
4.7. Enzyme Activity Assays
4.8. Relative Gene Expression Analysis in Rat Livers
4.9. Microsomal Incubations for BaP Metabolites Detection
4.10. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BaP | benzo[a]pyrene |
| BPDE | benzo[a]pyrene-7,8-dihydrodiol-9,10-epoxide |
| bw | body weight |
| CYP | cytochrome P450 |
| DMSO | dimethyl sulfoxide |
| dG-N2-BPDE | 10-(deoxyguanosin-N2-yl)-7,8,9-trihydroxy-7,8,9,10-tetrahydrobenzo[a]pyrene |
| EROD | 7-ethoxyresorufin O-deethylation |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| i.p. | intraperitoneal injection |
| mEH | microsomal epoxide hydrolase |
| MROD | 7-methoxyresorufin O-demethylation |
| r | correlation coefficient |
| SI | Sudan I |
| TLC | thin layer chromatography |
References
- IARC. Chemical Agents and Related Occupations; IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 2012; Volume 100F, ISBN 978-92-832-1323-9. [Google Scholar]
- Miller, K.P.; Ramos, K.S. Impact of Cellular Metabolism on the Biological Effects of Benzo[a]Pyrene and Related Hydrocarbons. Drug Metab. Rev. 2001, 33, 1–35. [Google Scholar] [CrossRef]
- Baird, W.M.; Hooven, L.A.; Mahadevan, B. Carcinogenic Polycyclic Aromatic Hydrocarbon-DNA Adducts and Mechanism of Action. Environ. Mol. Mutagen. 2005, 45, 106–114. [Google Scholar] [CrossRef]
- Luch, A.; Baird, W.M. Metabolic Activation and Detoxification of Polycyclic Aromatic Hydrocarbons. In The Carcinogenic Effects of Polycyclic Aromatic Hydrocarbons; Imperial College Press: London, UK, 2005; pp. 19–96. ISBN 978-1-86094-417-8. [Google Scholar]
- Shimada, T.; Fujii-Kuriyama, Y. Metabolic Activation of Polycyclic Aromatic Hydrocarbons to Carcinogens by Cytochromes P450 1A1 and 1B1. Cancer Sci. 2004, 95, 1–6. [Google Scholar] [CrossRef]
- Stiborová, M.; Moserová, M.; Černá, V.; Indra, R.; Dračínský, M.; Šulc, M.; Henderson, C.J.; Wolf, C.R.; Schmeiser, H.H.; Phillips, D.H.; et al. Cytochrome B5 and Epoxide Hydrolase Contribute to Benzo[a]Pyrene-DNA Adduct Formation Catalyzed by Cytochrome P450 1A1 under Low NADPH:P450 Oxidoreductase Conditions. Toxicology 2014, 318, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Stansbury, K.H.; Walker, N.J.; Trush, M.A.; Strickland, P.T.; Sutter, T.R. Metabolism of Benzo[a]Pyrene and Benzo[a]Pyrene-7,8-Diol by Human Cytochrome P450 1B1. Carcinogenesis 1998, 19, 1847–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimada, T.; Oda, Y.; Gillam, E.M.; Guengerich, F.P.; Inoue, K. Metabolic Activation of Polycyclic Aromatic Hydrocarbons and Other Procarcinogens by Cytochromes P450 1A1 and P450 1B1 Allelic Variants and Other Human Cytochromes P450 in Salmonella typhimurium NM2009. Drug Metab. Dispos. Biol. Fate Chem. 2001, 29, 1176–1182. [Google Scholar] [PubMed]
- Šulc, M.; Indra, R.; Moserová, M.; Schmeiser, H.H.; Frei, E.; Arlt, V.M.; Stiborová, M. The Impact of Individual Cytochrome P450 Enzymes on Oxidative Metabolism of Benzo[a]Pyrene in Human Livers. Environ. Mol. Mutagen. 2016, 57, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Bauer, E.; Guo, Z.; Ueng, Y.F.; Bell, L.C.; Zeldin, D.; Guengerich, F.P. Oxidation of Benzo[a]Pyrene by Recombinant Human Cytochrome P450 Enzymes. Chem. Res. Toxicol. 1995, 8, 136–142. [Google Scholar] [CrossRef]
- IARC Some Aromatic Azo Compounds; IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 1975; Volume 8, ISBN 978-92-832-1208-9.
- Federal Institute for Risk Assessment (BfR) Dyes Sudan I to IV in Food 2003. BfR Opinion 19 November 2003. Available online: https://www.bfr.bund.de/cm/349/dyes_sudan_I_IV.pdf (accessed on 23 June 2021).
- Møller, P.; Wallin, H. Genotoxic Hazards of Azo Pigments and Other Colorants Related to 1-Phenylazo-2-Hydroxynaphthalene. Mutat. Res. Mutat. Res. 2000, 462, 13–30. [Google Scholar] [CrossRef]
- Mazzetti, M.; Fascioli, R.; Mazzoncini, I.; Spinelli, G.; Morelli, I.; Bertoli, A. Determination of 1-Phenylazo-2-Naphthol (Sudan I) in Chilli Powder and in Chilli-Containing Food Products by GPC Clean-up and HPLC with LC/MS Confirmation. Food Addit. Contam. 2004, 21, 935–941. [Google Scholar] [CrossRef]
- Dixit, S.; Khanna, S.K.; Das, M. A Simple 2-Directional High-Performance Thin-Layer Chromatographic Method for the Simultaneous Determination of Curcumin, Metanil Yellow, and Sudan Dyes in Turmeric, Chili, and Curry Powders. J. AOAC Int. 2008, 91, 1387–1396. [Google Scholar] [CrossRef] [Green Version]
- Moreno-González, D.; Jáč, P.; Švec, F.; Nováková, L. Determination of Sudan Dyes in Chili Products by Micellar Electrokinetic Chromatography-MS/MS Using a Volatile Surfactant. Food Chem. 2020, 310, 125963. [Google Scholar] [CrossRef]
- Schwack, W.; Pellissier, E.; Morlock, G. Analysis of Unauthorized Sudan Dyes in Food by High-Performance Thin-Layer Chromatography. Anal. Bioanal. Chem. 2018, 410, 5641–5651. [Google Scholar] [CrossRef]
- Stiborová, M.; Martínek, V.; Rýdlová, H.; Hodek, P.; Frei, E. Sudan I Is a Potential Carcinogen for Humans: Evidence for Its Metabolic Activation and Detoxication by Human Recombinant Cytochrome P450 1A1 and Liver Microsomes. Cancer Res. 2002, 62, 5678–5684. [Google Scholar]
- Stiborova, M.; Schmeiser, H.H.; Frei, E.; Hodek, P.; Martinek, V. Enzymes Oxidizing the Azo Dye 1-Phenylazo-2-Naphthol (Sudan I) and Their Contribution to Its Genotoxicity and Carcinogenicity. Curr. Drug Metab. 2014, 15, 829–840. [Google Scholar] [CrossRef]
- Nebert, D.W.; Shi, Z.; Gálvez-Peralta, M.; Uno, S.; Dragin, N. Oral Benzo[a]Pyrene: Understanding Pharmacokinetics, Detoxication, and Consequences—Cyp1 Knockout Mouse Lines as a Paradigm. Mol. Pharmacol. 2013, 84, 304–313. [Google Scholar] [CrossRef] [Green Version]
- Shimada, T.; Sugie, A.; Shindo, M.; Nakajima, T.; Azuma, E.; Hashimoto, M.; Inoue, K. Tissue-Specific Induction of Cytochromes P450 1A1 and 1B1 by Polycyclic Aromatic Hydrocarbons and Polychlorinated Biphenyls in Engineered C57BL/6J Mice of Arylhydrocarbon Receptor Gene. Toxicol. Appl. Pharmacol. 2003, 187, 1–10. [Google Scholar] [CrossRef]
- Harrigan, J.A.; McGarrigle, B.P.; Sutter, T.R.; Olson, J.R. Tissue Specific Induction of Cytochrome P450 (CYP) 1A1 and 1B1 in Rat Liver and Lung Following in Vitro (Tissue Slice) and in Vivo Exposure to Benzo(a)Pyrene. Toxicol. Vitro Int. J. Publ. Assoc. BIBRA 2006, 20, 426–438. [Google Scholar] [CrossRef]
- Refat, N.A.G.A.; Ibrahim, Z.S.; Moustafa, G.G.; Sakamoto, K.Q.; Ishizuka, M.; Fujita, S. The Induction of Cytochrome P450 1A1 by Sudan Dyes. J. Biochem. Mol. Toxicol. 2008, 22, 77–84. [Google Scholar] [CrossRef]
- Stiborová, M.; Dračínská, H.; Martínek, V.; Svášková, D.; Hodek, P.; Milichovský, J.; Hejduková, Ž.; Brotánek, J.; Schmeiser, H.H.; Frei, E. Induced Expression of Cytochrome P450 1A and NAD(P)H:Quinone Oxidoreductase Determined at MRNA, Protein, and Enzyme Activity Levels in Rats Exposed to the Carcinogenic Azo Dye 1-Phenylazo-2-Naphthol (Sudan I). Chem. Res. Toxicol. 2013, 26, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Hodek, P.; Koblihová, J.; Kizek, R.; Frei, E.; Arlt, V.M.; Stiborová, M. The Relationship between DNA Adduct Formation by Benzo[a]Pyrene and Expression of Its Activation Enzyme Cytochrome P450 1A1 in Rat. Environ. Toxicol. Pharmacol. 2013, 36, 989–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aimová, D.; Poljaková, J.; Kotrbová, V.; Moserová, M.; Frei, E.; Arlt, V.M.; Stiborová, M. Ellipticine and Benzo(a)Pyrene Increase Their Own Metabolic Activation via Modulation of Expression and Enzymatic Activity of Cytochromes P450 1A1 and 1A2. Interdiscip. Toxicol. 2008, 1, 160–168. [Google Scholar] [CrossRef] [Green Version]
- Stiborová, M.; Indra, R.; Moserová, M.; Bořek-Dohalská, L.; Hodek, P.; Frei, E.; Kopka, K.; Schmeiser, H.H.; Arlt, V.M. Comparison of Human Cytochrome P450 1A1-Catalysed Oxidation of Benzo[a]Pyrene in Prokaryotic and Eukaryotic Expression Systems. Mon. Chem. 2017, 148, 1959–1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uno, S.; Dalton, T.P.; Derkenne, S.; Curran, C.P.; Miller, M.L.; Shertzer, H.G.; Nebert, D.W. Oral Exposure to Benzo[a]Pyrene in the Mouse: Detoxication by Inducible Cytochrome P450 Is More Important Than Metabolic Activation. Mol. Pharmacol. 2004, 65, 1225–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arlt, V.M.; Stiborová, M.; Henderson, C.J.; Thiemann, M.; Frei, E.; Aimová, D.; Singh, R.; Gamboa da Costa, G.; Schmitz, O.J.; Farmer, P.B.; et al. Metabolic Activation of Benzo[a]Pyrene in Vitro by Hepatic Cytochrome P450 Contrasts with Detoxification in Vivo: Experiments with Hepatic Cytochrome P450 Reductase Null Mice. Carcinogenesis 2008, 29, 656–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, L.; Arlt, V.M.; Phillips, D.H. The Role of Cytochrome P450 Enzymes in Carcinogen Activation and Detoxication: An in Vivo–in Vitro Paradox. Carcinogenesis 2018, 39, 851–859. [Google Scholar] [CrossRef]
- Fang, A.H.; Smith, W.A.; Vouros, P.; Gupta, R.C. Identification and Characterization of a Novel Benzo[a]Pyrene-Derived DNA Adduct. Biochem. Biophys. Res. Commun. 2001, 281, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Schoket, B.; Lévay, K.; Phillips, D.H.; Vincze, I. 32P-Postlabelling Analysis of DNA Adducts of Benzo[a]Pyrene Formed in Complex Metabolic Activation Systems in Vitro. Cancer Lett. 1989, 48, 67–75. [Google Scholar] [CrossRef]
- Ross, J.; Nelson, G.; Kligerman, A.; Erexson, G.; Bryant, M.; Earley, K.; Gupta, R.; Nesnow, S. Formation and Persistence of Novel Benzo(a)Pyrene Adducts in Rat Lung, Liver, and Peripheral Blood Lymphocyte DNA. Cancer Res. 1990, 50, 5088–5094. [Google Scholar]
- Vähäkangas, K.; Raunio, H.; Pasanen, M.; Sivonen, P.; Park, S.S.; Gelboin, H.V.; Pelkonen, O. Comparison of the Formation of Benzo[a]Pyrene Diolepoxide-DNA Adducts in Vitro by Rat and Human Microsomes: Evidence for the Involvement of P-450IA1 and P-450IA2. J. Biochem. Toxicol. 1989, 4, 79–86. [Google Scholar] [CrossRef]
- Indra, R. Mechanism of Enzymatic Activation of Carcinogens and Drugs by the System of Cytochrome P450; Charles University: Prague, Czech Republic, 2015. [Google Scholar]
- Burke, M.D.; Thompson, S.; Weaver, R.J.; Wolf, C.R.; Mayers, R.T. Cytochrome P450 Specificities of Alkoxyresorufin O-Dealkylation in Human and Rat Liver. Biochem. Pharmacol. 1994, 48, 923–936. [Google Scholar] [CrossRef]
- Stiborová, M.; Martínek, V.; Rýdlová, H.; Koblas, T.; Hodek, P. Expression of Cytochrome P450 1A1 and Its Contribution to Oxidation of a Potential Human Carcinogen 1-Phenylazo-2-Naphthol (Sudan I) in Human Livers. Cancer Lett. 2005, 220, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Safe, S. Molecular Biology of the Ah Receptor and Its Role in Carcinogenesis. Toxicol. Lett. 2001, 120, 1–7. [Google Scholar] [CrossRef]
- Stiborová, M.; Indra, R.; Moserová, M.; Frei, E.; Schmeiser, H.H.; Kopka, K.; Philips, D.H.; Arlt, V.M. NADH:Cytochrome B5 Reductase and Cytochrome B5 Can Act as Sole Electron Donors to Human Cytochrome P450 1A1-Mediated Oxidation and DNA Adduct Formation by Benzo[a]Pyrene. Chem. Res. Toxicol. 2016, 29, 1325–1334. [Google Scholar] [CrossRef] [Green Version]
- Skupinska, K.; Misiewicz, I.; Kasprzycka-Guttman, T. A Comparison of the Concentration-Effect Relationships of PAHs on CYP1A Induction in HepG2 and Mcf7 Cells. Arch. Toxicol. 2007, 81, 183–200. [Google Scholar] [CrossRef]
- Arlt, V.M.; Krais, A.M.; Godschalk, R.W.; Riffo-Vasquez, Y.; Mrizova, I.; Roufosse, C.A.; Corbin, C.; Shi, Q.; Frei, E.; Stiborova, M.; et al. Pulmonary Inflammation Impacts on CYP1A1-Mediated Respiratory Tract DNA Damage Induced by the Carcinogenic Air Pollutant Benzo[a]Pyrene. Toxicol. Sci. Off. J. Soc. Toxicol. 2015, 146, 213–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nesnow, S.; Ross, J.; Nelson, G.; Holden, K.; Erexson, G.; Kligerman, A.; Gupta, R.C. Quantitative and Temporal Relationships between DNA Adduct Formation in Target and Surrogate Tissues: Implications for Biomonitoring. Environ. Health Perspect. 1993, 101 (Suppl. S3), 37–42. [Google Scholar] [CrossRef]
- Stansbury, K.H.; Flesher, J.W.; Gupta, R.C. Mechanism of Aralkyl-DNA Adduct Formation from Benzo[a]Pyrene in Vivo. Chem. Res. Toxicol. 1994, 7, 254–259. [Google Scholar] [CrossRef]
- Lee, B.M.; Shim, G.A. Dietary Exposure Estimation of Benzo[a]Pyrene and Cancer Risk Assessment. J. Toxicol. Environ. Health A 2007, 70, 1391–1394. [Google Scholar] [CrossRef]
- Alomirah, H.; Al-Zenki, S.; Al-Hooti, S.; Zaghloul, S.; Sawaya, W.; Ahmed, N.; Kannan, K. Concentrations and Dietary Exposure to Polycyclic Aromatic Hydrocarbons (PAHs) from Grilled and Smoked Foods. Food Control 2011, 22, 2028–2035. [Google Scholar] [CrossRef]
- Public Health England Polycyclic Aromatic Hydrocarbons (Benzo[a]Pyrene) Toxicological Overview 2018. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/737017/PAH_TO_PHE_240818.pdf (accessed on 16 July 2021).
- Ding, Y.S.; Ward, J.; Hammond, D.; Watson, C.H. Mouth-Level Intake of Benzo[a]Pyrene from Reduced Nicotine Cigarettes. Int. J. Environ. Res. Public. Health 2014, 11, 11898–11914. [Google Scholar] [CrossRef] [Green Version]
- Shimada, T. Xenobiotic-Metabolizing Enzymes Involved in Activation and Detoxification of Carcinogenic Polycyclic Aromatic Hydrocarbons. Drug Metab. Pharmacokinet. 2006, 21, 257–276. [Google Scholar] [CrossRef] [Green Version]
- Stiborova, M.; Schmeiser, H.H.; Breuer, A.; Frei, E. P-32-Postlabelling Analysis of DNA Adducts with 1-(Phenylazo)-2-Naphthol (Sudan I, Solvent Yellow 14) Formed in Vivo in Fisher 344 Rats. Collect. Czechoslov. Chem. Commun. 1999, 64, 1335–1347. [Google Scholar] [CrossRef]
- Arlt, V.M.; Poirier, M.C.; Sykes, S.E.; John, K.; Moserova, M.; Stiborova, M.; Wolf, C.R.; Henderson, C.J.; Phillips, D.H. Exposure to Benzo[a]Pyrene of Hepatic Cytochrome P450 Reductase Null (HRN) and P450 Reductase Conditional Null (RCN) Mice: Detection of Benzo[a]Pyrene Diol Epoxide-DNA Adducts by Immunohistochemistry and 32P-Postlabelling. Toxicol. Lett. 2012, 213, 160–166. [Google Scholar] [CrossRef] [Green Version]
- Sims, P.; Grover, P.L.; Swaisland, A.; Pal, K.; Hewer, A. Metabolic Activation of Benzo(a)Pyrene Proceeds by a Diol-Epoxide. Nature 1974, 252, 326–328. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.; Mrizova, I.; Barta, F.; Indra, R.; Moserova, M.; Kopka, K.; Schmeiser, H.H.; Wolf, C.R.; Henderson, C.J.; Stiborova, M.; et al. Cytochrome B5impacts on Cytochrome P450-Mediated Metabolism of Benzo[a]Pyrene and Its DNA Adduct Formation: Studies in Hepatic Cytochrome B5/P450 Reductase Null (HBRN) Mice. Arch. Toxicol. 2018, 92, 1625–1638. [Google Scholar] [CrossRef] [Green Version]
- Beal, M.A.; Meier, M.J.; Williams, A.; Rowan-Carroll, A.; Gagné, R.; Lindsay, S.J.; Fitzgerald, T.; Hurles, M.E.; Marchetti, F.; Yauk, C.L. Paternal Exposure to Benzo(a)Pyrene Induces Genome-Wide Mutations in Mouse Offspring. Commun. Biol. 2019, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Reed, L.; Jarvis, I.W.H.; Phillips, D.H.; Arlt, V.M. Enhanced DNA Adduct Formation by Benzo[a]Pyrene in Human Liver Cells Lacking Cytochrome P450 Oxidoreductase. Mutat. Res. Toxicol. Environ. Mutagen. 2020, 852, 503162. [Google Scholar] [CrossRef]
- Meier, S.; Karlsen, Ø.; Le Goff, J.; Sørensen, L.; Sørhus, E.; Pampanin, D.M.; Donald, C.E.; Fjelldal, P.G.; Dunaevskaya, E.; Romano, M.; et al. DNA Damage and Health Effects in Juvenile Haddock (Melanogrammus aeglefinus) Exposed to PAHs Associated with Oil-Polluted Sediment or Produced Water. PLoS ONE 2020, 15, e0240307. [Google Scholar] [CrossRef]
- Kishino, Y.; Hasegawa, T.; Yamoto, T.; Mori, K. Species Differences in Micronucleus Induction of the Clastogenic Compounds Associated with Drug Metabolic Profile. J. Toxicol. Sci. 2019, 44, 701–709. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, A.; Knuckles, M.E. Dose-Dependent Benzo(a)Pyrene [B(a)P]–DNA Adduct Levels and Persistence in F-344 Rats Following Subchronic Dietary Exposure to B(a)P. Cancer Lett. 2006, 240, 268–278. [Google Scholar] [CrossRef]
- Denison, M.S.; Whitlock, J.P. Xenobiotic-Inducible Transcription of Cytochrome P450 Genes. J. Biol. Chem. 1995, 270, 18175–18178. [Google Scholar] [CrossRef] [Green Version]
- Stiborová, M.; Dračínská, H.; Bořek-Dohalská, L.; Klusoňová, Z.; Holecová, J.; Martínková, M.; Schmeiser, H.H.; Arlt, V.M. Exposure to Endocrine Disruptors 17alpha-Ethinylestradiol and Estradiol Influences Cytochrome P450 1A1-Mediated Genotoxicity of Benzo[a]Pyrene and Expression of This Enzyme in Rats. Toxicology 2018, 400–401, 48–56. [Google Scholar] [CrossRef]
- Endo, K.; Uno, S.; Seki, T.; Ariga, T.; Kusumi, Y.; Mitsumata, M.; Yamada, S.; Makishima, M. Inhibition of Aryl Hydrocarbon Receptor Transactivation and DNA Adduct Formation by CYP1 Isoform-Selective Metabolic Deactivation of Benzo[a]Pyrene. Toxicol. Appl. Pharmacol. 2008, 230, 135–143. [Google Scholar] [CrossRef]
- Dickins, M. Induction of Cytochromes P450. Curr. Top. Med. Chem. 2004, 4, 1745–1766. [Google Scholar] [CrossRef] [PubMed]
- Floreani, M.; Gabbia, D.; Barbierato, M.; DE Martin, S.; Palatini, P. Differential Inducing Effect of Benzo[a]Pyrene on Gene Expression and Enzyme Activity of Cytochromes P450 1A1 and 1A2 in Sprague-Dawley and Wistar Rats. Drug Metab. Pharmacokinet. 2012, 27, 640–652. [Google Scholar] [CrossRef]
- Wang, K.; Feng, C.; Li, C.; Yao, J.; Xie, X.; Gong, L.; Luan, Y.; Xing, G.; Zhu, X.; Qi, X.; et al. Baicalin Protects Mice from Aristolochic Acid I-Induced Kidney Injury by Induction of CYP1A through the Aromatic Hydrocarbon Receptor. Int. J. Mol. Sci. 2015, 16, 16454–16468. [Google Scholar] [CrossRef] [Green Version]
- Yoshinari, K.; Ueda, R.; Kusano, K.; Yoshimura, T.; Nagata, K.; Yamazoe, Y. Omeprazole Transactivates Human CYP1A1 and CYP1A2 Expression through the Common Regulatory Region Containing Multiple Xenobiotic-Responsive Elements. Biochem. Pharmacol. 2008, 76, 139–145. [Google Scholar] [CrossRef]
- Lin, J.H. CYP Induction-Mediated Drug Interactions: In Vitro Assessment and Clinical Implications. Pharm. Res. 2006, 23, 1089–1116. [Google Scholar] [CrossRef]
- Tompkins, L.M.; Wallace, A.D. Mechanisms of Cytochrome P450 Induction. J. Biochem. Mol. Toxicol. 2007, 21, 176–181. [Google Scholar] [CrossRef]
- Corcos, L.; Marc, N.; Wein, S.; Fautrel, A.; Guillouzo, A.; Pineau, T. Phenobarbital Induces Cytochrome P4501A2 HnRNA, MRNA and Protein in the Liver of C57BL/6J Wild Type and Aryl Hydrocarbon Receptor Knock-out Mice. FEBS Lett. 1998, 425, 293–297. [Google Scholar] [CrossRef]
- Ryu, D.Y.; Levi, P.E.; Hodgson, E. Regulation of Hepatic CYP1A Isozymes by Piperonyl Butoxide and Acenaphthylene in the Mouse. Chem. Biol. Interact. 1997, 105, 53–63. [Google Scholar] [CrossRef]
- Silver, G.; Krauter, K.S. Expression of Cytochromes P-450c and P-450d MRNAs in Cultured Rat Hepatocytes. 3-Methylcholanthrene Induction Is Regulated Primarily at the Post-Transcriptional Level. J. Biol. Chem. 1988, 263, 11802–11807. [Google Scholar] [CrossRef]
- Yoshinari, K.; Yoda, N.; Toriyabe, T.; Yamazoe, Y. Constitutive Androstane Receptor Transcriptionally Activates Human CYP1A1 and CYP1A2 Genes through a Common Regulatory Element in the 5’-Flanking Region. Biochem. Pharmacol. 2010, 79, 261–269. [Google Scholar] [CrossRef]
- Lubet, R.A.; Connolly, G.; Kouri, R.E.; Nebert, D.W.; Bigelow, S.W. Biological Effects of the Sudan Dyes. Role of the Ah Cytosolic Receptor. Biochem. Pharmacol. 1983, 32, 3053–3058. [Google Scholar] [CrossRef]
- Schmeiser, H.H.; Stiborova, M.; Arlt, V.M. 32P-Postlabeling Analysis of DNA Adducts. In Genotoxicity Assessment: Methods and Protocols; Dhawan, A., Bajpayee, M., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2013; pp. 389–401. ISBN 978-1-62703-529-3. [Google Scholar]
- Kimura, T.; Kodama, M.; Nagata, C. N-Hydroxylation Enzymes of Carcinogenic Aminoazo Dyes: Possible Involvement of Cytochrome P-448. Gan 1982, 73, 55–62. [Google Scholar]
- Omura, T.; Sato, R. The carbon monoxide-binding pigment of liver microsomes. I. Evidence for its hemoprotein nature. J. Biol. Chem. 1964, 239, 2370–2378. [Google Scholar] [CrossRef]
- Stiborová, M.; Dračínská, H.; Mizerovská, J.; Frei, E.; Schmeiser, H.H.; Hudeček, J.; Hodek, P.; Phillips, D.H.; Arlt, V.M. The Environmental Pollutant and Carcinogen 3-Nitrobenzanthrone Induces Cytochrome P450 1A1 and NAD(P)H:Quinone Oxidoreductase in Rat Lung and Kidney, Thereby Enhancing Its Own Genotoxicity. Toxicology 2008, 247, 11–22. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mRNA | CYP1A1 | CYP1A2 | CYP1B1 | |||
|---|---|---|---|---|---|---|
| ΔcT ± S.D. | Fold Change (to Control/BaP) | ΔcT ± S.D. | Fold Change (to Control/BaP) | ΔcT ± S.D. | Fold Change (to Control/BaP) | |
| control | 7.49 ± 0.122 | - | −0.17 ± 0.007 | - | 11.36 ± 0.691 | - |
| BaP | −4.06 ± 0.183 | 2990 *** | −4.96 ± 0.175 | 27.7 *** | −0.42 ± 0.071 | 3520 *** |
| Sudan I | −3.22 ± 0.526 | 1680 *** | −4.66 ± 0.461 | 22.5 *** | −1.76 ± 0.286 | 8860 *** |
| SI + BaP | −3.07 ± 0.187 | 1510 ***/0.5 # | −4.45 ± 0.083 | 19.5 */0.7 # | −1.40 ± 0.161 | 6920 ***/2.0 # |
| BaP + SI | −4.25 ± 0.163 | 3420 ***/1.1 | −4.89 ± 0.106 | 26.4 */1.0 | −2.06 ± 0.109 | 10,900 ***/3.1 # |
| mRNA | mEH | |
|---|---|---|
| ΔcT ± S.D. | Fold Change (to Control/BaP) | |
| control | −1.97 ± 0.244 | - |
| BaP | −2.65 ± 0.361 | 1.6 * |
| Sudan I | −3.04 ± 0.406 | 2.1 *** |
| SI + BaP | −2.58 ± 0.379 | 1.5 */0.9 |
| BaP + SI | −3.26 ± 0.286 | 2.5/1.6 |
| Group Name | Treatment | Day 1 | Day 2 |
|---|---|---|---|
| control | untreated | Sunflower oil (i.p.) | Sunflower oil (gavage) |
| BaP | BaP | Sunflower oil (i.p.) | BaP (gavage) |
| Sudan I | Sudan I | Sunflower oil (gavage) | Sudan I (i.p.) |
| SI + BaP | Sudan I + BaP | Sudan I (i.p.) | BaP (gavage) |
| BaP + SI | BaP + Sudan I | BaP (gavage) | Sudan I (i.p.) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dračínská, H.; Indra, R.; Jelínková, S.; Černá, V.; Arlt, V.M.; Stiborová, M. Benzo[a]pyrene-Induced Genotoxicity in Rats Is Affected by Co-Exposure to Sudan I by Altering the Expression of Biotransformation Enzymes. Int. J. Mol. Sci. 2021, 22, 8062. https://doi.org/10.3390/ijms22158062
Dračínská H, Indra R, Jelínková S, Černá V, Arlt VM, Stiborová M. Benzo[a]pyrene-Induced Genotoxicity in Rats Is Affected by Co-Exposure to Sudan I by Altering the Expression of Biotransformation Enzymes. International Journal of Molecular Sciences. 2021; 22(15):8062. https://doi.org/10.3390/ijms22158062
Chicago/Turabian StyleDračínská, Helena, Radek Indra, Sandra Jelínková, Věra Černá, Volker Manfred Arlt, and Marie Stiborová. 2021. "Benzo[a]pyrene-Induced Genotoxicity in Rats Is Affected by Co-Exposure to Sudan I by Altering the Expression of Biotransformation Enzymes" International Journal of Molecular Sciences 22, no. 15: 8062. https://doi.org/10.3390/ijms22158062
APA StyleDračínská, H., Indra, R., Jelínková, S., Černá, V., Arlt, V. M., & Stiborová, M. (2021). Benzo[a]pyrene-Induced Genotoxicity in Rats Is Affected by Co-Exposure to Sudan I by Altering the Expression of Biotransformation Enzymes. International Journal of Molecular Sciences, 22(15), 8062. https://doi.org/10.3390/ijms22158062