
Skin Wound Healing Rate in Fish Depends on Species and Microbiota

 ,
,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
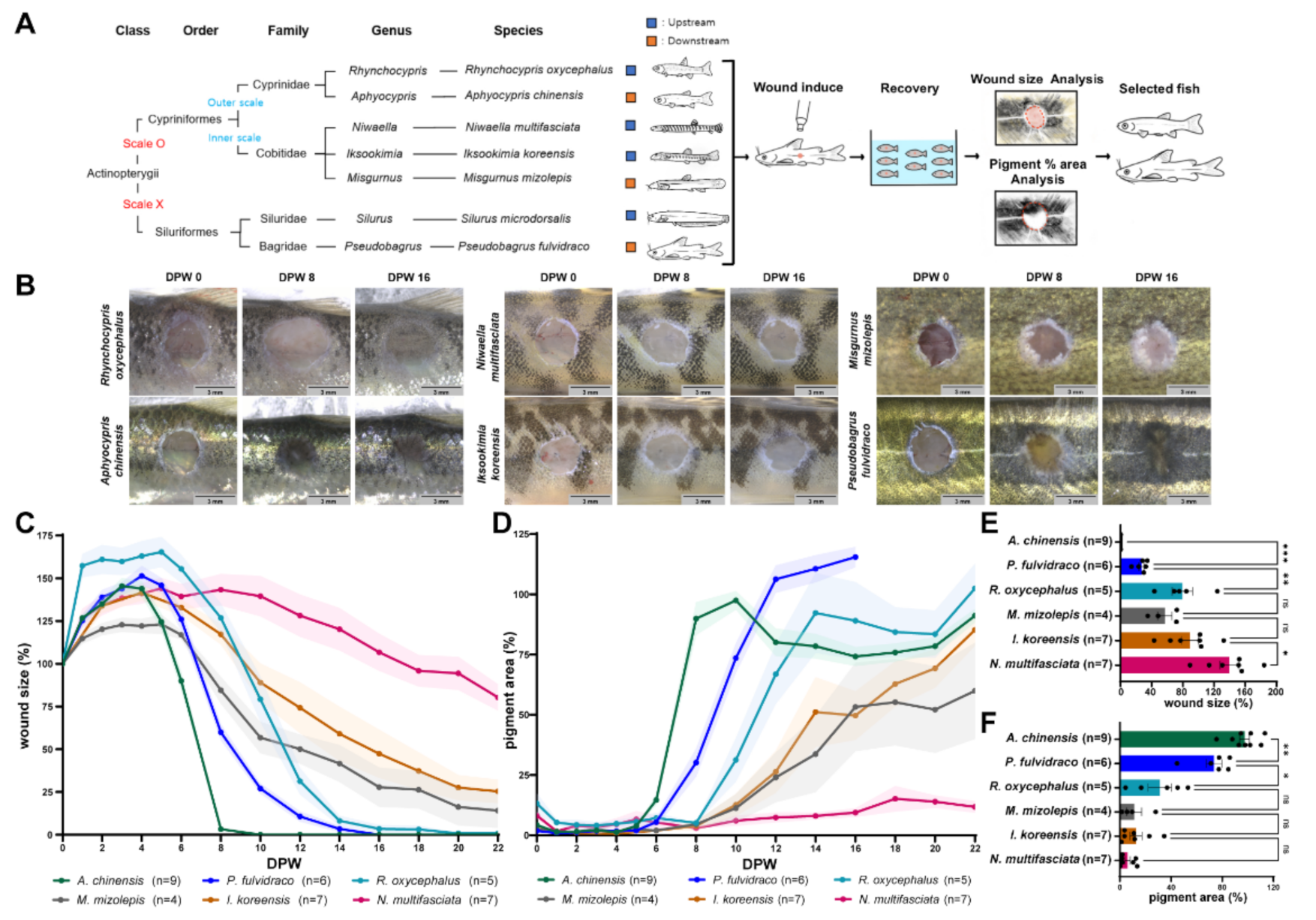
2.1. Korean Bullhead and Chinese Bleak Display Faster Wound Healing Rates
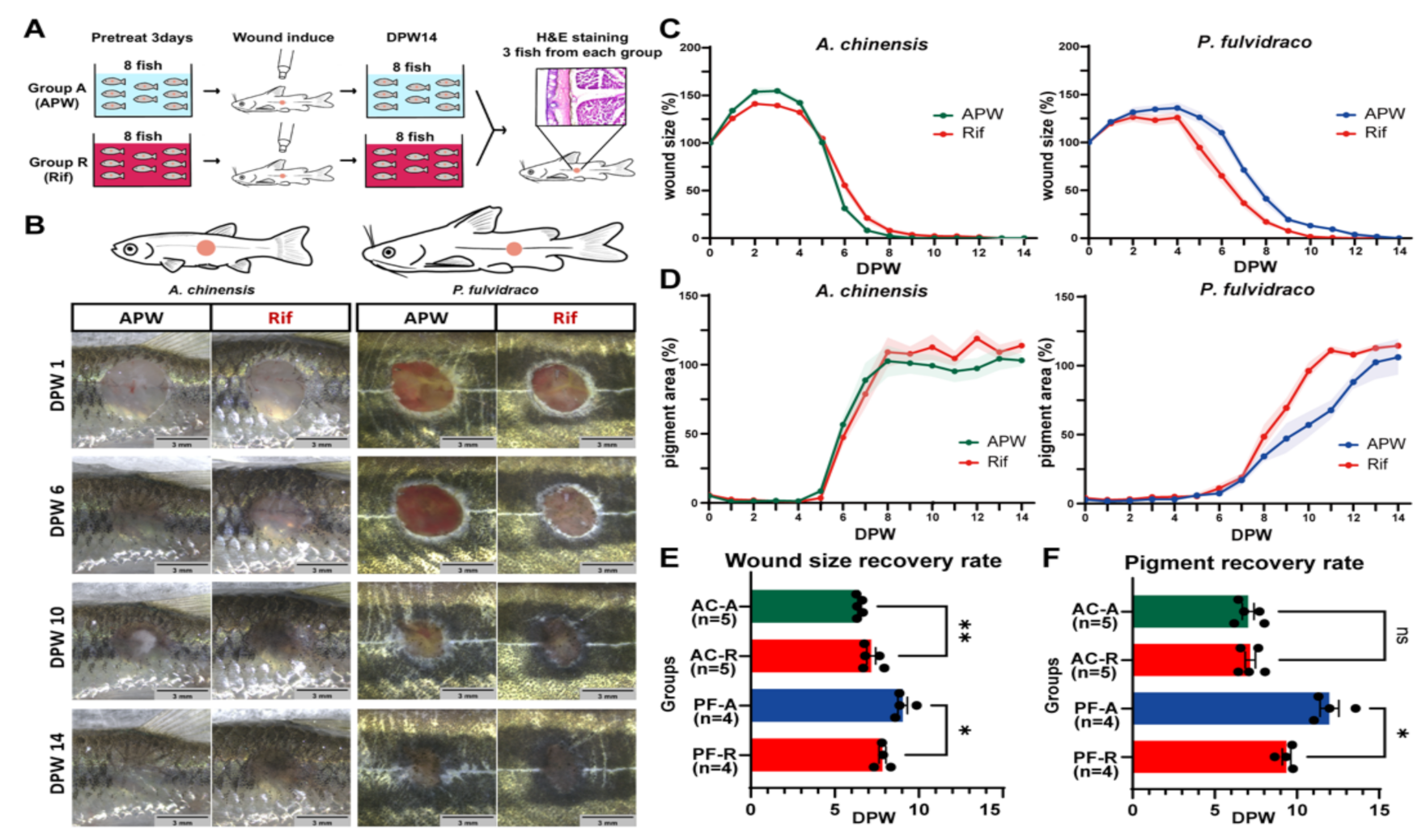
2.2. Rifampicin Treatment Induces Different Effects on the Wound Healing Rate
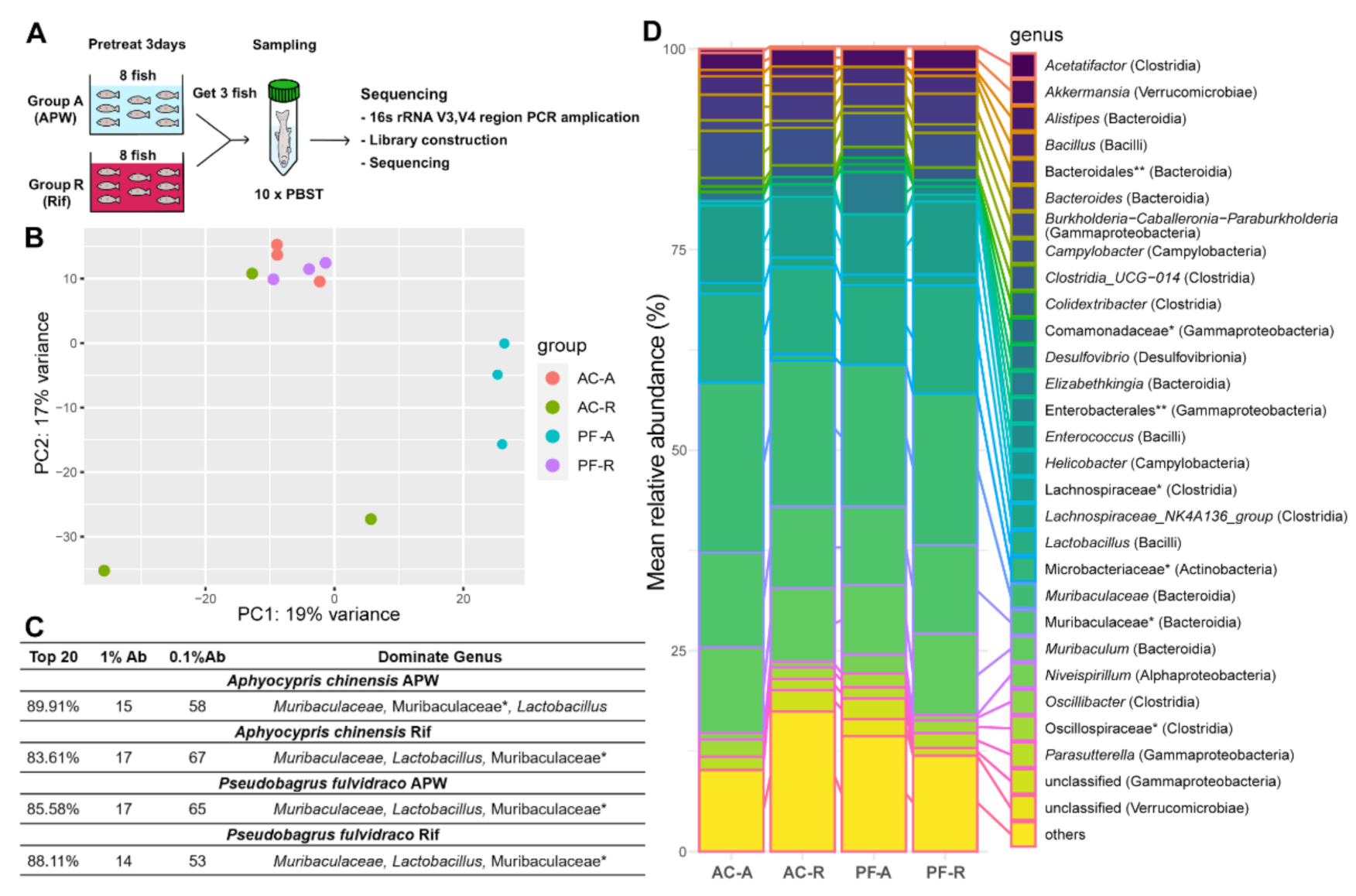
2.3. Microbiomes in Two Fish Species Respond Differently to Rifampicin Treatment
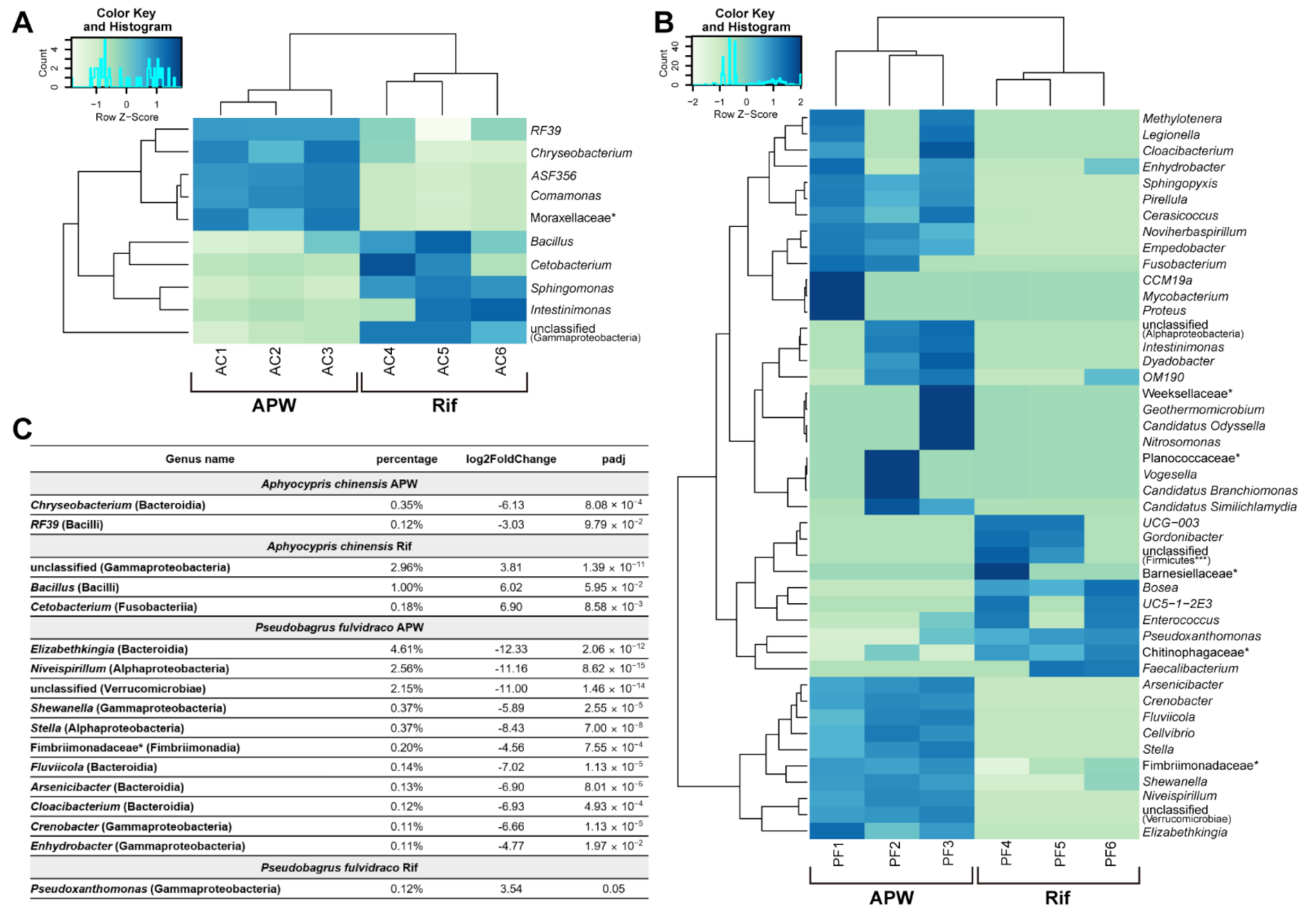
2.4. Upon Rifampicin Treatment, the Microbiome Changes with Some Correlation with the Wound Healing Rate
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moss, M.L. Vertebrate Dermis and Integumental Skeleton. Am. Zool 1972, 12, 27–34. [Google Scholar] [CrossRef][Green Version]
- Hawkes, J.W. The structure of fish skin. I. General organization. Cell Tissue Res. 1974, 149, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Grada, A.; Mervis, J.; Falanga, V. Research Techniques Made Simple: Animal Models of Wound Healing. J. Investig. Derm. 2018, 138, 2095–2105.e1. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.R.; Gomez, B.I.; McIntyre, M.K.; Dubick, M.A.; Christy, R.J.; Nicholson, S.E.; Burmeister, D.M. The Cutaneous Microbiome and Wounds: New Molecular Targets to Promote Wound Healing. Int. J. Mol. Sci. 2018, 19, 2699. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.B.; Dananjaya, S.H.S.; Nikapitiya, C.; Park, B.K.; Gooneratne, R.; Kim, T.Y.; Lee, J.; Kim, C.H.; De Zoysa, M. Silver nanoparticles enhance wound healing in zebrafish (Danio rerio). Fish Shellfish Immunol. 2017, 68, 536–545. [Google Scholar] [CrossRef] [PubMed]
- Richardson, R.; Slanchev, K.; Kraus, C.; Knyphausen, P.; Eming, S.; Hammerschmidt, M. Adult Zebrafish as a Model System for Cutaneous Wound-Healing Research. J. Investig. Dermatol. 2013, 133, 1655–1665. [Google Scholar] [CrossRef] [PubMed]
- Richardson, R.; Metzger, M.; Knyphausen, P.; Ramezani, T.; Slanchev, K.; Kraus, C.; Schmelzer, E.; Hammerschmidt, M. Re-epithelialization of cutaneous wounds in adult zebrafish combines mechanisms of wound closure in embryonic and adult mammals. Development 2016, 143, 2077–2088. [Google Scholar]
- Sveen, L.; Karlsen, C.; Ytteborg, E. Mechanical induced wounds in fish—A review on models and healing mechanisms. Rev. Aquacult. 2020, 12, 2446–2465. [Google Scholar] [CrossRef]
- Anderson, C.D.; Roberts, R.J. Comparison of Effects of Temperature on Wound-Healing in a Tropical and a Temperate Teleost. J. Fish Biol. 1975, 7, 173–182. [Google Scholar] [CrossRef]
- Jensen, L.B.; Wahli, T.; McGurk, C.; Eriksen, T.B.; Obach, A.; Waagbo, R.; Handler, A.; Tafalla, C. Effect of temperature and diet on wound healing in Atlantic salmon (Salmo salar L.). Fish Physiol. Biochem. 2015, 41, 1527–1543. [Google Scholar] [CrossRef]
- Rakers, S.; Gebert, M.; Uppalapati, S.; Meyer, W.; Maderson, P.; Sell, A.F.; Kruse, C.; Paus, R. ‘Fish matters’: The relevance of fish skin biology to investigative dermatology. Exp. Derm. 2010, 19, 313–324. [Google Scholar] [CrossRef]
- Gomez, J.A.; Primm, T.P. A Slimy Business: The Future of Fish Skin Microbiome Studies. Microb. Ecol. 2021. [Google Scholar] [CrossRef]
- Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; Bouffard, G.G.; Blakesley, R.W.; Murray, P.R.; Green, E.D.; et al. Topographical and Temporal Diversity of the Human Skin Microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef] [PubMed]
- Di Domizio, J.; Belkhodja, C.; Chenuet, P.; Fries, A.; Murray, T.; Mondejar, P.M.; Demaria, O.; Conrad, C.; Homey, B.; Werner, S.; et al. The commensal skin microbiota triggers type I IFN-dependent innate repair responses in injured skin. Nat. Immunol. 2020, 21, 1034–1045. [Google Scholar] [CrossRef]
- Altoe, L.S.; Alves, R.S.; Sarandy, M.M.; Morais-Santos, M.; Novaes, R.D.; Goncalves, R.V. Does antibiotic use accelerate or retard cutaneous repair? A systematic review in animal models. PLoS ONE 2019, 14, e0223511. [Google Scholar] [CrossRef]
- Hjelmstedt, P.; Sundh, H.; Brijs, J.; Ekstrom, A.; Sundell, K.S.; Berg, C.; Sandblom, E.; Bowman, J.; Morgenroth, D.; Grans, A. Effects of prophylactic antibiotic-treatment on post-surgical recovery following intraperitoneal bio-logger implantation in rainbow trout. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Guerra, R.R.; Santos, N.P.; Cecarelli, P.; Silva, J.R.M.C.; Hernandez-Blazquez, F.J. Healing of skin wounds in the African catfish Clarias gariepinus. J. Fish Biol. 2008, 73, 572–583. [Google Scholar] [CrossRef]
- Alhassan, J.M.; Thomson, M.; Criddle, K.R.; Summers, B.; Criddle, R.S. Catfish Epidermal Secretions in Response to Threat or Injury—A Novel Defense Response. Mar. Biol. 1985, 88, 117–123. [Google Scholar] [CrossRef]
- Carlson, J.M.; Leonard, A.B.; Hyde, E.R.; Petrosino, J.F.; Primm, T.P. Microbiome disruption and recovery in the fish Gambusia affinis following exposure to broad-spectrum antibiotic. Infect. Drug Resist. 2017, 10, 143–154. [Google Scholar] [CrossRef]
- Pandey, S.; Stockwell, C.A.; Snider, M.R.; Wisenden, B.D. Epidermal Club Cells in Fishes: A Case for Ecoimmunological Analysis. Int. J. Mol. Sci. 2021, 22, 1440. [Google Scholar] [CrossRef] [PubMed]
- Blazer, V.S.; Fabacher, D.L.; Little, E.E.; Ewing, M.S.; Kocan, K.M. Effects of ultraviolet-B radiation on fish: Histologic comparison of a UVB-sensitive and a UVB-tolerant species. J. Aquat. Anim. Health 1997, 9, 132–143. [Google Scholar] [CrossRef]
- Oh, M.K.; Park, J.Y. Seasonal Change of Skin Mucus Cells of Misgurnus mizolep (Cobitidae) Dwelling in a natural stream in Korea. Korean J. Ichthyol. 2010, 22, 230–237. [Google Scholar]
- Park, J. Morphology and histochemistry of the skin of the Korean spined loach, Iksookimia koreensis (Cobitidae), in relation to respiration. Folia Zool. 2002, 51, 241–247. [Google Scholar]
- Wang, G.; Sweren, E.; Liu, H.; Wier, E.; Alphonse, M.P.; Chen, R.; Islam, N.; Li, A.; Xue, Y.; Chen, J.; et al. Bacteria induce skin regeneration via IL-1beta signaling. Cell Host Microbe 2021. [Google Scholar] [CrossRef] [PubMed]
- Canesso, M.C.C.; Vieira, A.T.; Castro, T.B.R.; Schirmer, B.G.A.; Cisalpino, D.; Martins, F.S.; Rachid, M.A.; Nicoli, J.R.; Teixeira, M.M.; Barcelos, L.S. Skin Wound Healing Is Accelerated and Scarless in the Absence of Commensal Microbiota. J. Immunol. 2014, 193, 5171–5180. [Google Scholar] [CrossRef]
- Chiarello, M.; Villeger, S.; Bouvier, C.; Bettarel, Y.; Bouvier, T. High diversity of skin-associated bacterial communities of marine fishes is promoted by their high variability among body parts, individuals and species. FEMS Microbiol. Ecol. 2015, 91. [Google Scholar] [CrossRef] [PubMed]
- Kim, I. A Review of the Spined Loaches, Family Cobitidae (Cypriniformes) in Korea. Korean J. Ichthyol. 2009, 21, 7–28. [Google Scholar]
- Kobayakawa, M. Systematic Revision of the Catfish Genus Silurus, with Description of a New Species from Thailand and Burma. Jpn. J. Ichthyol. 1989, 36, 155–186. [Google Scholar] [CrossRef]
- Lim, S.K.; Han, C.H. Annual Reproductive Cycle of the Banded Catfish, Pseudobagrus fulvidraco. Korean J. Fish Aquat. Sci. 1997, 30, 823–833. [Google Scholar]
- Watanabe, K.; Tabata, R.; Nakajima, J.; Kobayakawa, M.; Matsuda, M.; Takaku, K.; Hosoya, K.; Ohara, K.; Takagi, M.; Jang-Liaw, N.H. Large-scale hybridization of Japanese populations of Hinamoroko, Aphyocypris chinensis, with A. kikuchiiintroduced from Taiwan. Ichthyol. Res. 2020, 67, 361–374. [Google Scholar] [CrossRef]
- Yu, D.; Chen, M.; Zhou, Z.C.; Eric, R.; Tang, Q.Y.; Liu, H.Z. Global climate change will severely decrease potential distribution of the East Asian coldwater fish Rhynchocypris oxycephalus (Actinopterygii, Cyprinidae). Hydrobiologia 2013, 700, 23–32. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yun, T.; Shin, S.; Bang, K.; Lee, M.; Cho, J.-A.; Baek, M. Skin Wound Healing Rate in Fish Depends on Species and Microbiota. Int. J. Mol. Sci. 2021, 22, 7804. https://doi.org/10.3390/ijms22157804
Yun T, Shin S, Bang K, Lee M, Cho J-A, Baek M. Skin Wound Healing Rate in Fish Depends on Species and Microbiota. International Journal of Molecular Sciences. 2021; 22(15):7804. https://doi.org/10.3390/ijms22157804
Chicago/Turabian StyleYun, Tery, Soeun Shin, Kyungwon Bang, Mugeun Lee, Jung-Ah Cho, and Myungin Baek. 2021. "Skin Wound Healing Rate in Fish Depends on Species and Microbiota" International Journal of Molecular Sciences 22, no. 15: 7804. https://doi.org/10.3390/ijms22157804
APA StyleYun, T., Shin, S., Bang, K., Lee, M., Cho, J.-A., & Baek, M. (2021). Skin Wound Healing Rate in Fish Depends on Species and Microbiota. International Journal of Molecular Sciences, 22(15), 7804. https://doi.org/10.3390/ijms22157804