
Further Characterization of Intrastriatal Lipopolysaccharide Model of Parkinson’s Disease in C57BL/6 Mice




Abstract
:1. Introduction
2. Results
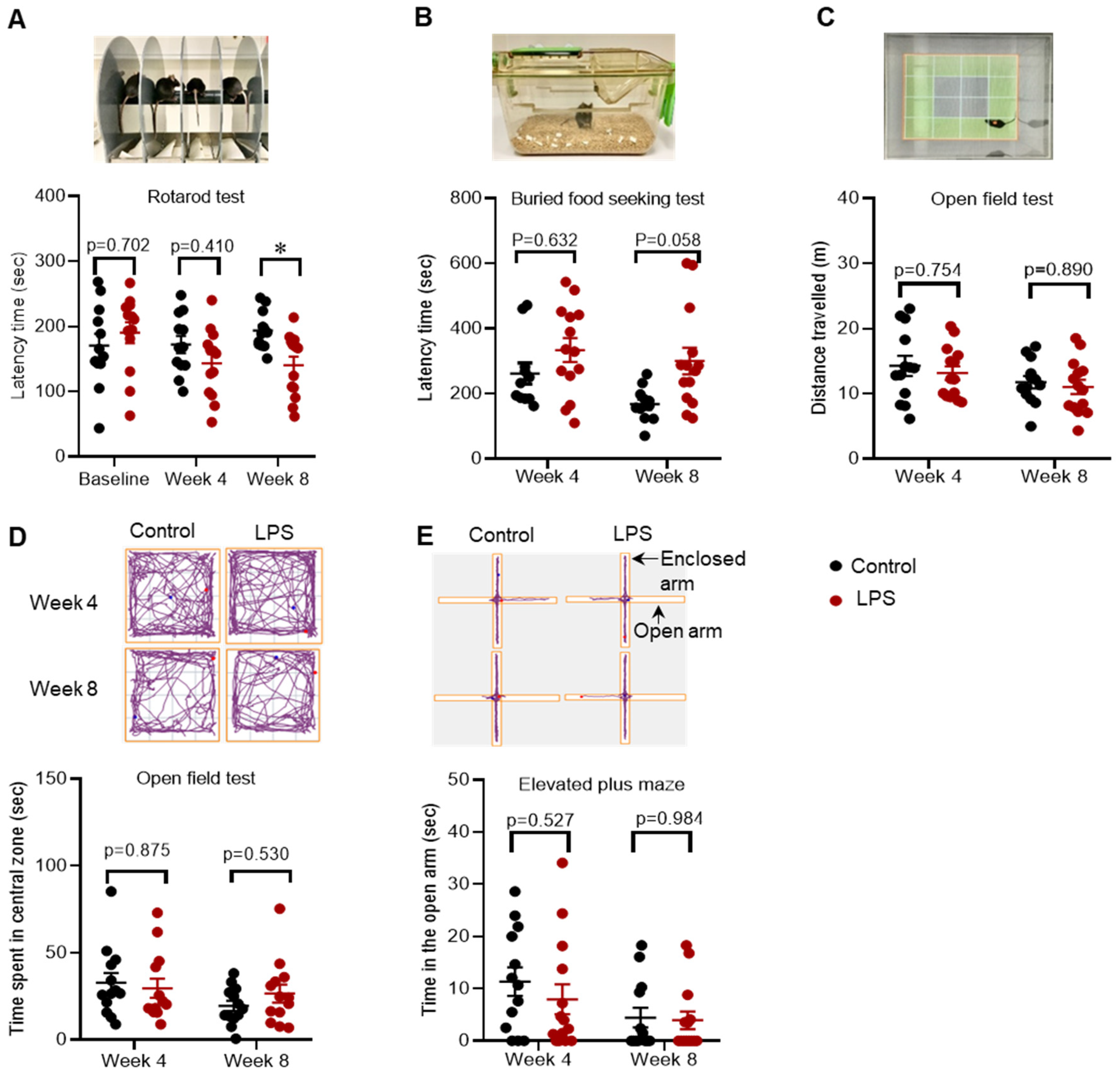
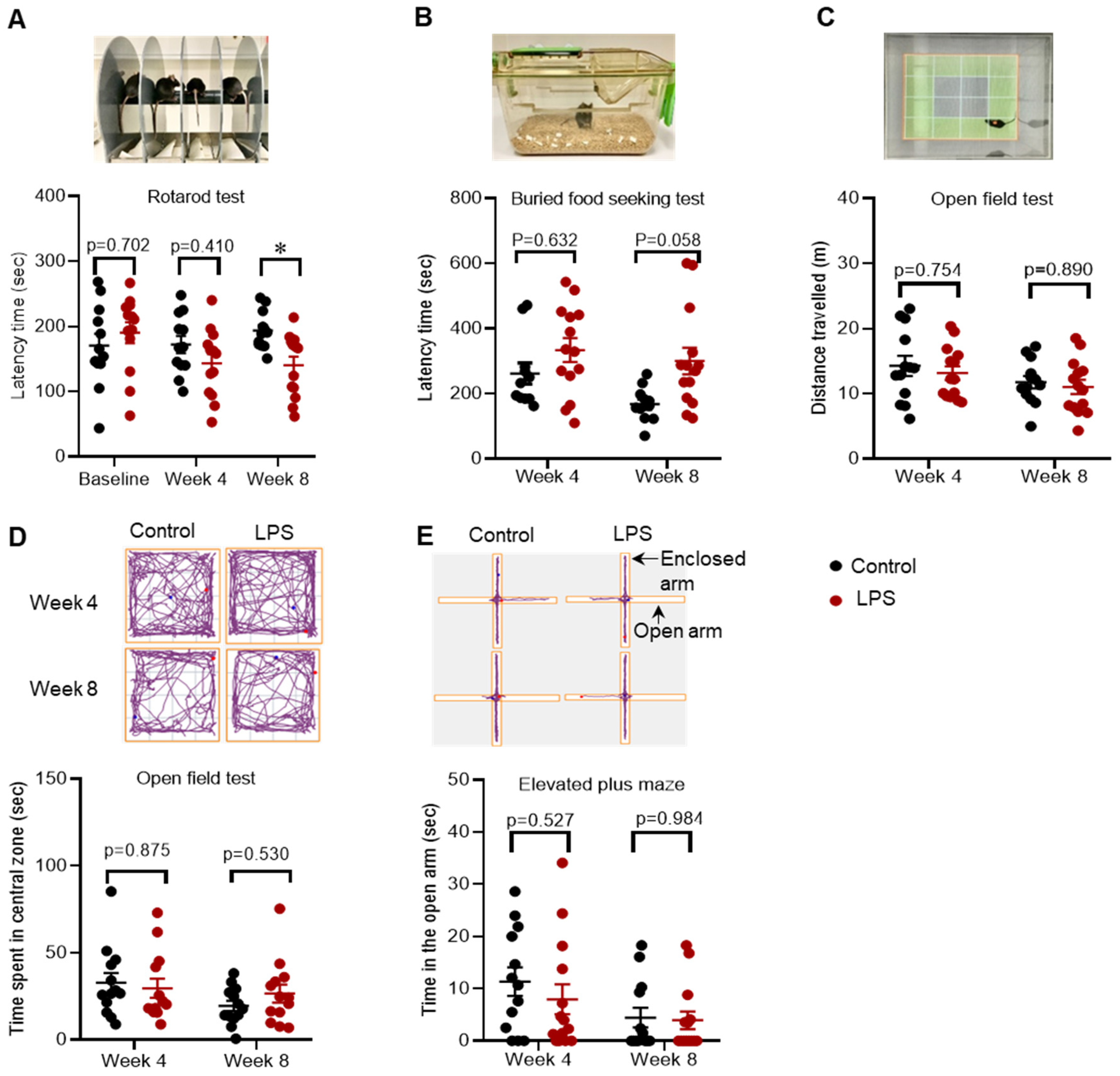
2.1. Effects of Intrastriatal Injection of LPS on Motor Behaviour, Olfactory Function and Mood in C57BL/6 Mice
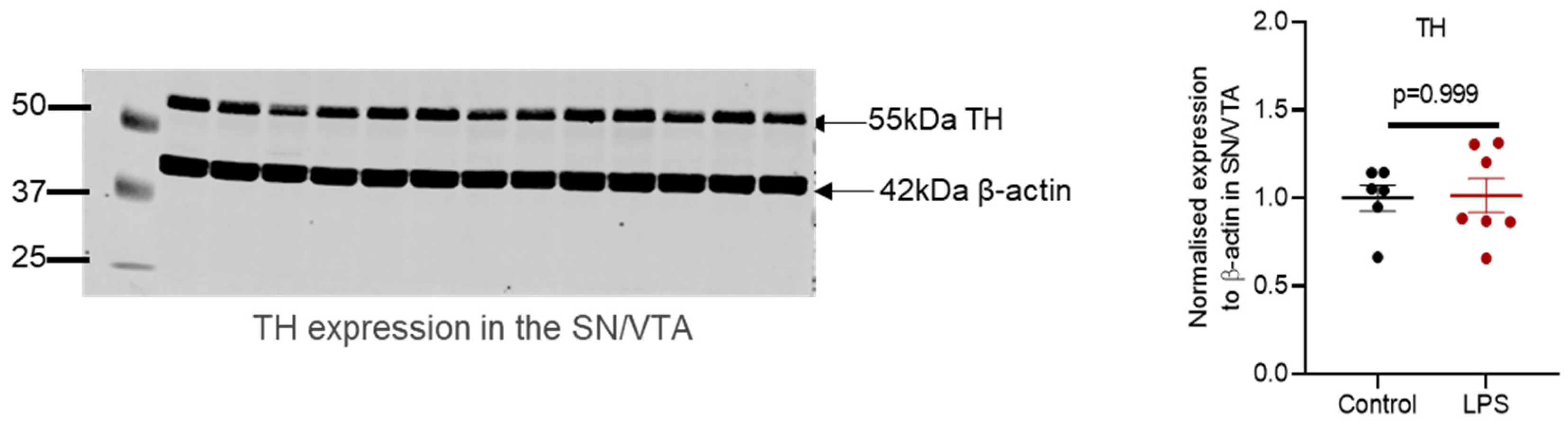
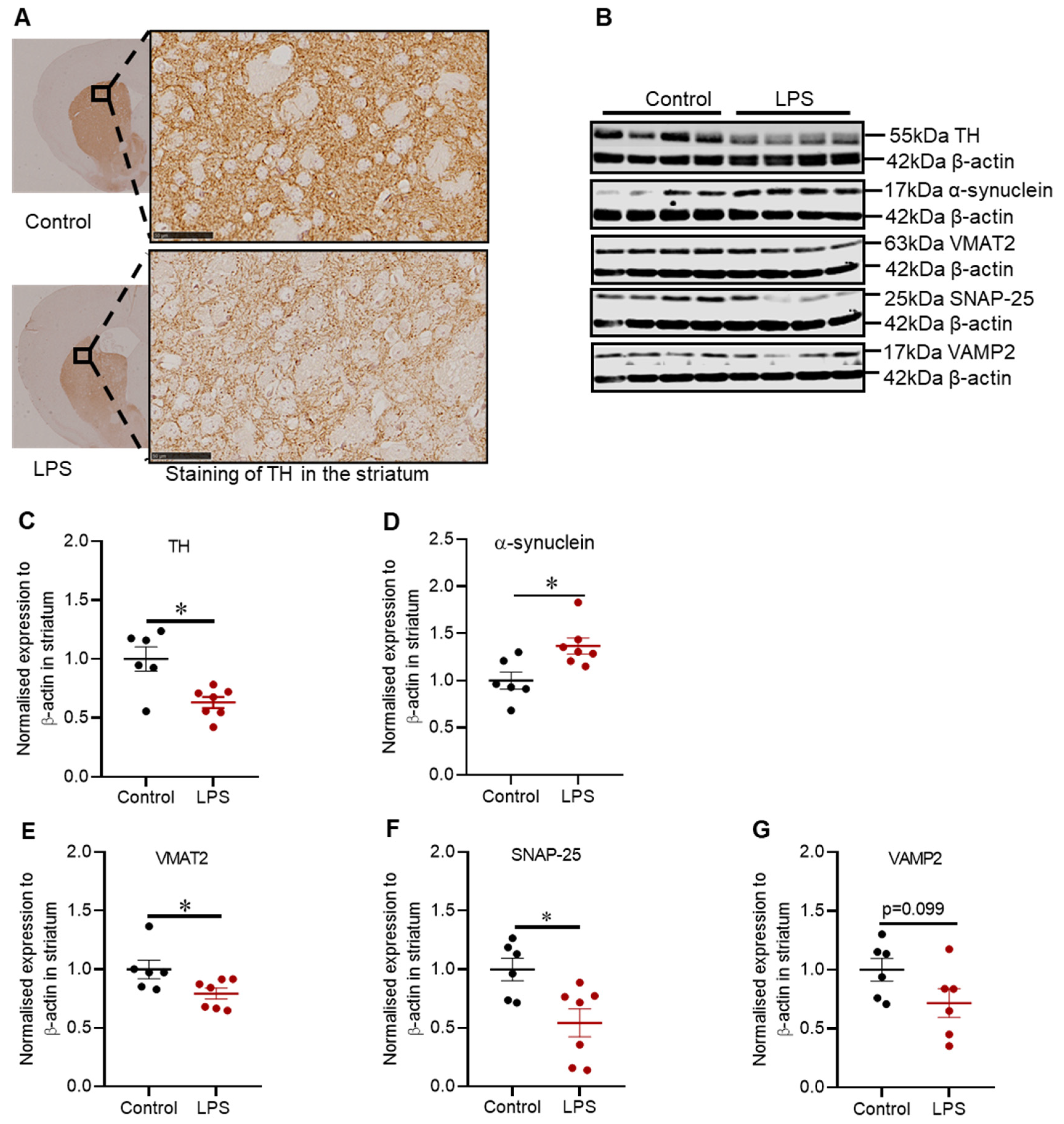
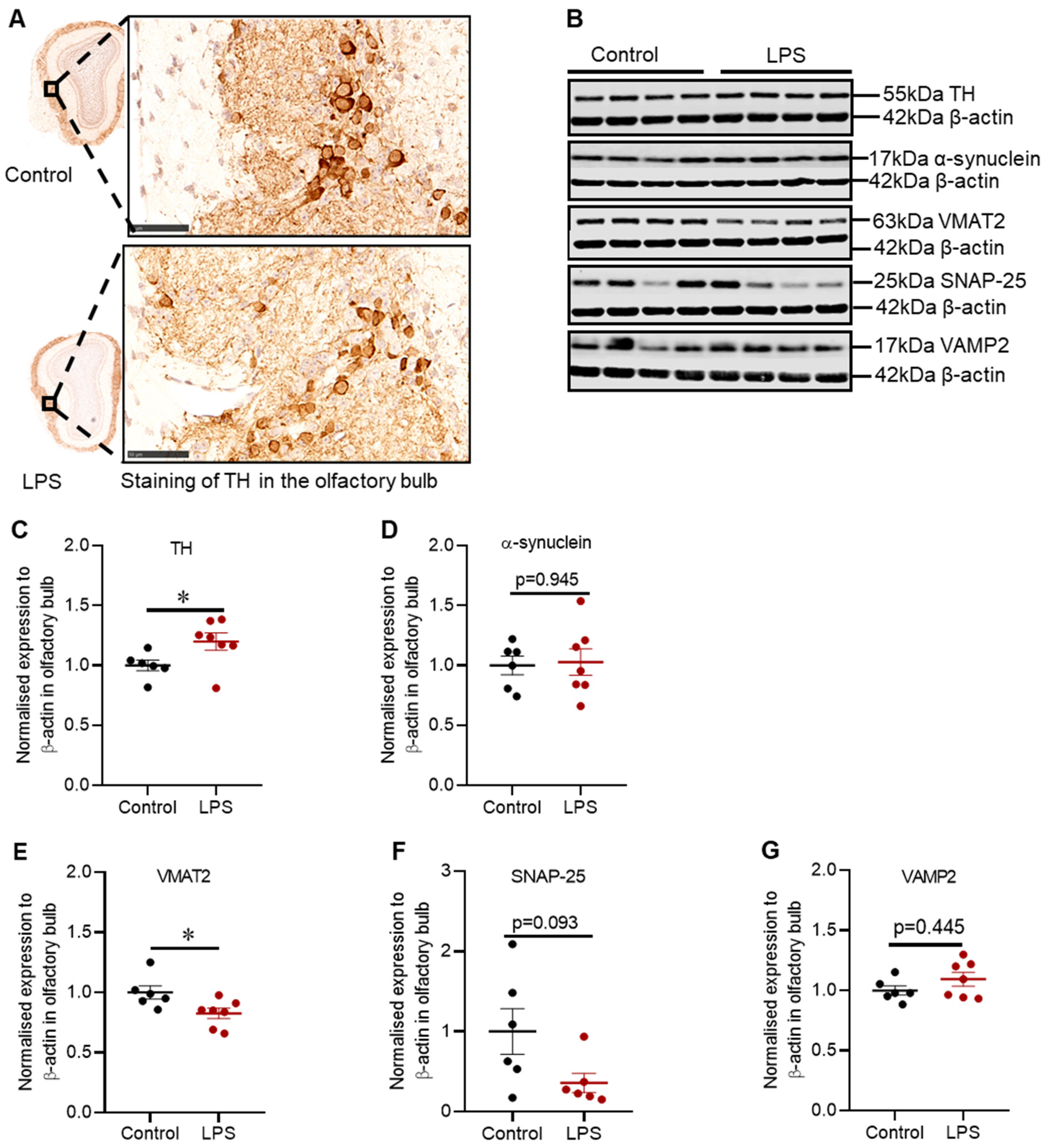
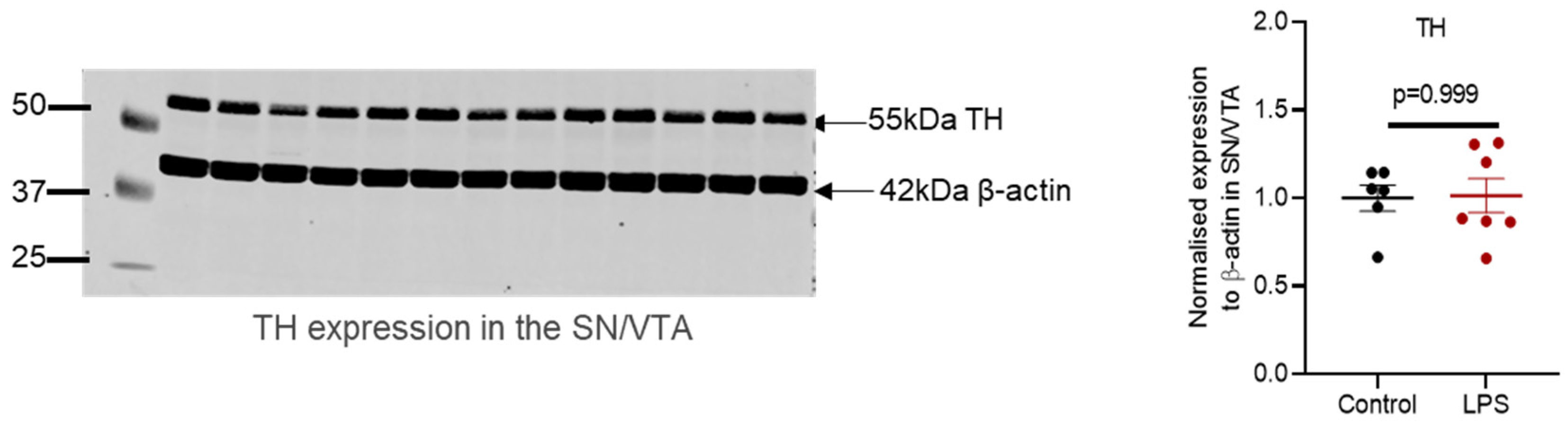
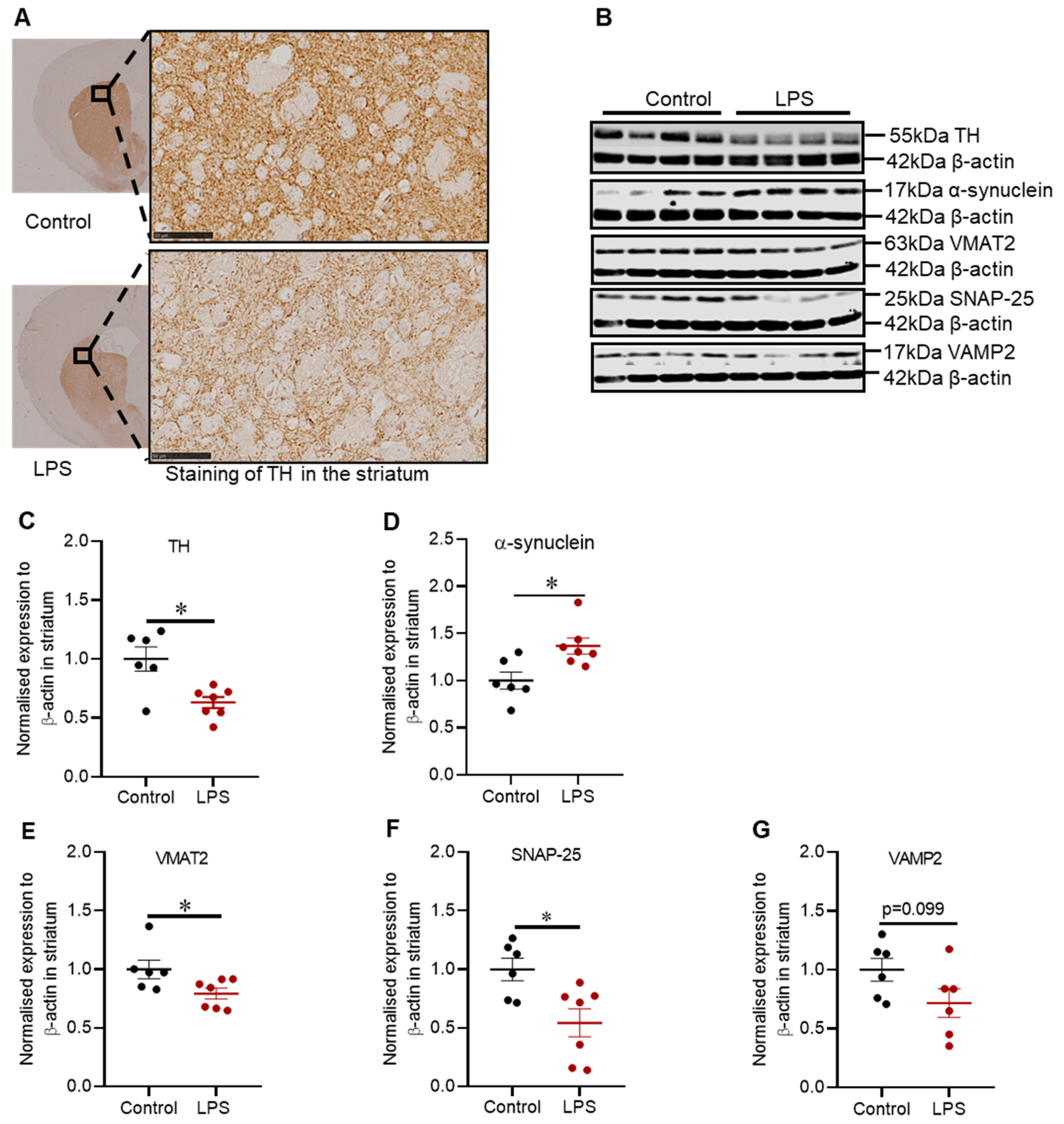
2.2. Administration of LPS Altered the Expression of TH, α-Synuclein and Synaptic Proteins in the Striatum of C57BL/6 Mice
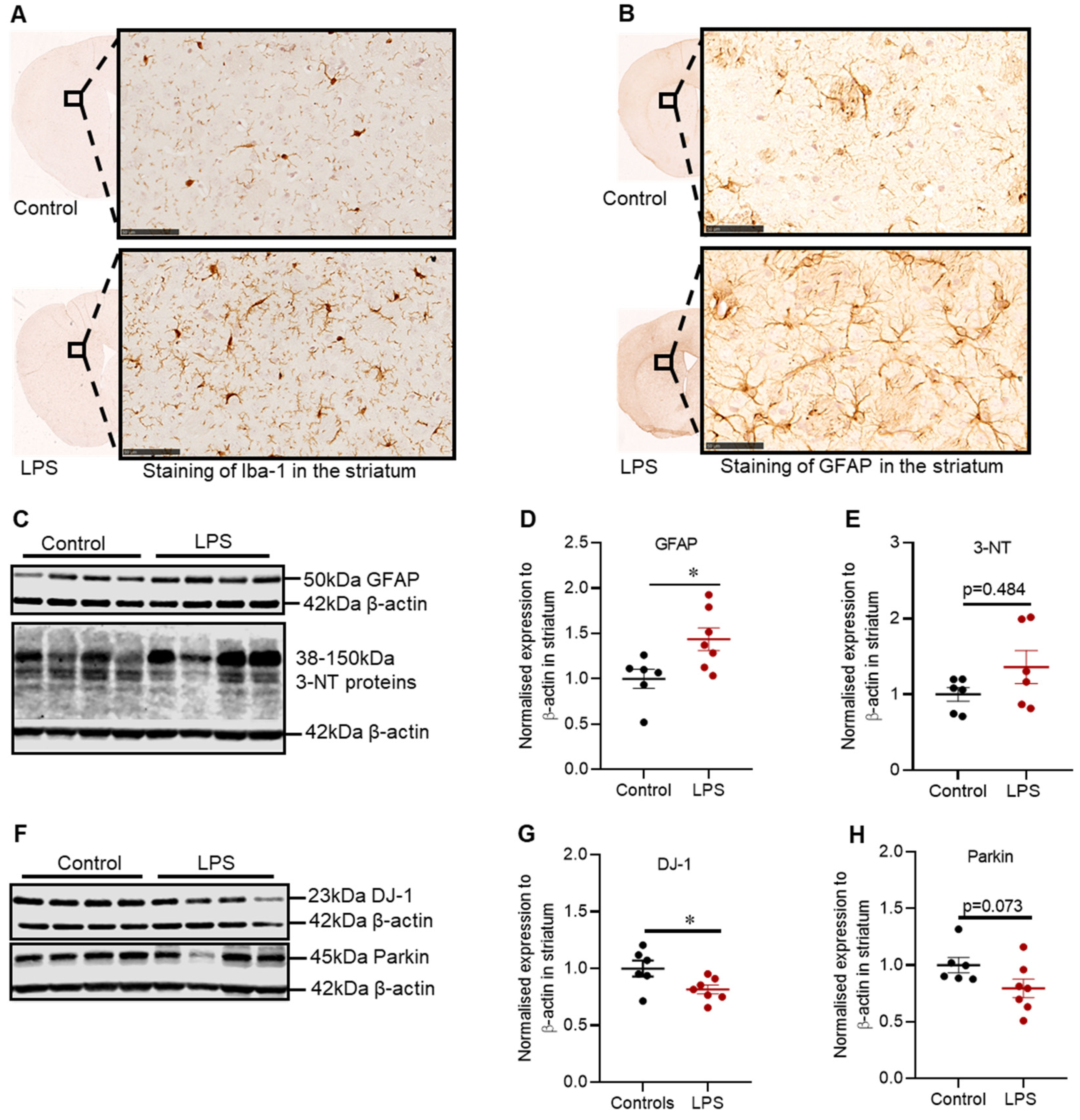
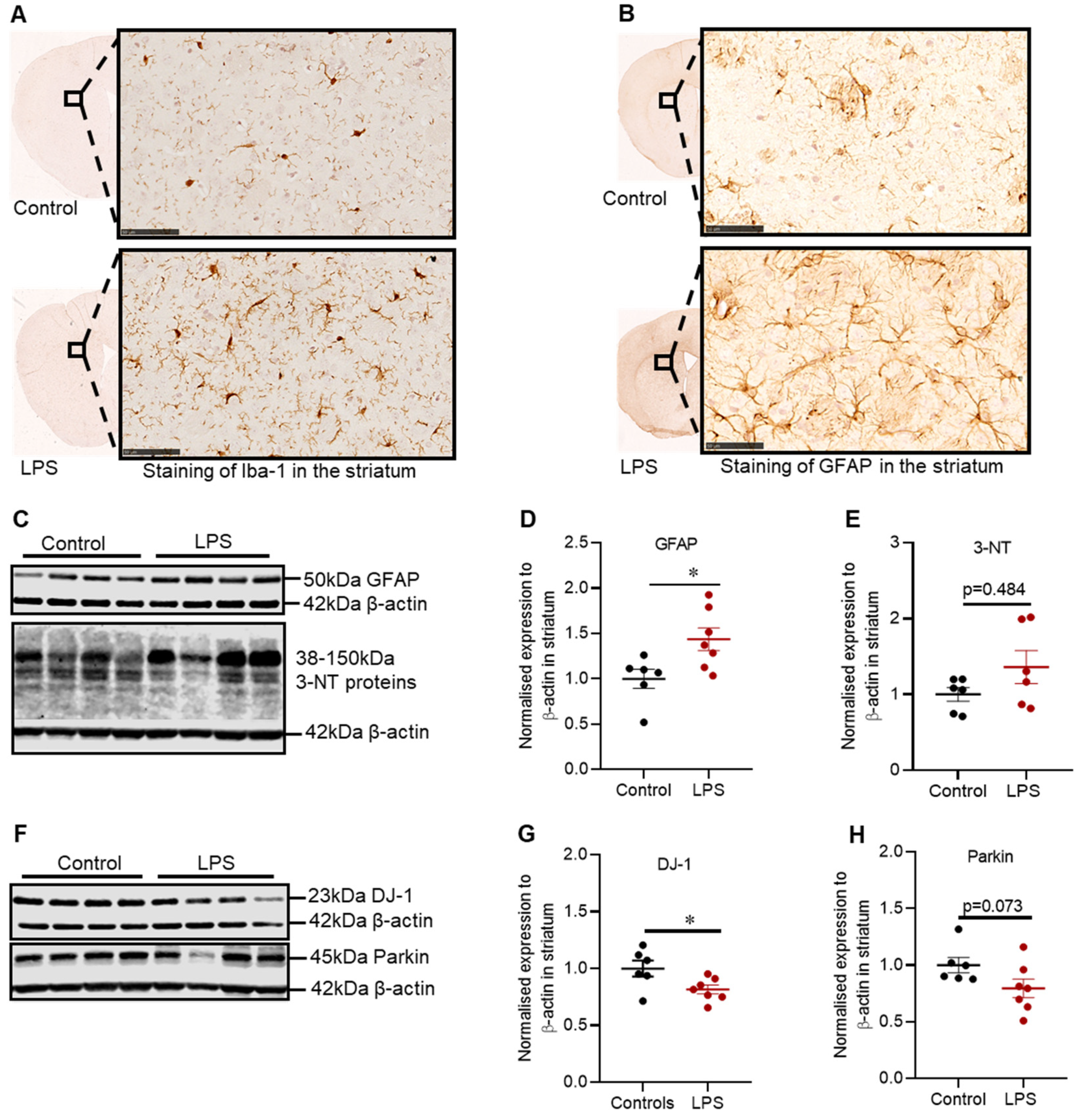
2.3. Effects of LPS on the Inflammatory and Oxidative Stress Markers, and Proteins Involved in Defence Mechanisms against Oxidative Stress in the Striatum of C57BL/6 Mice
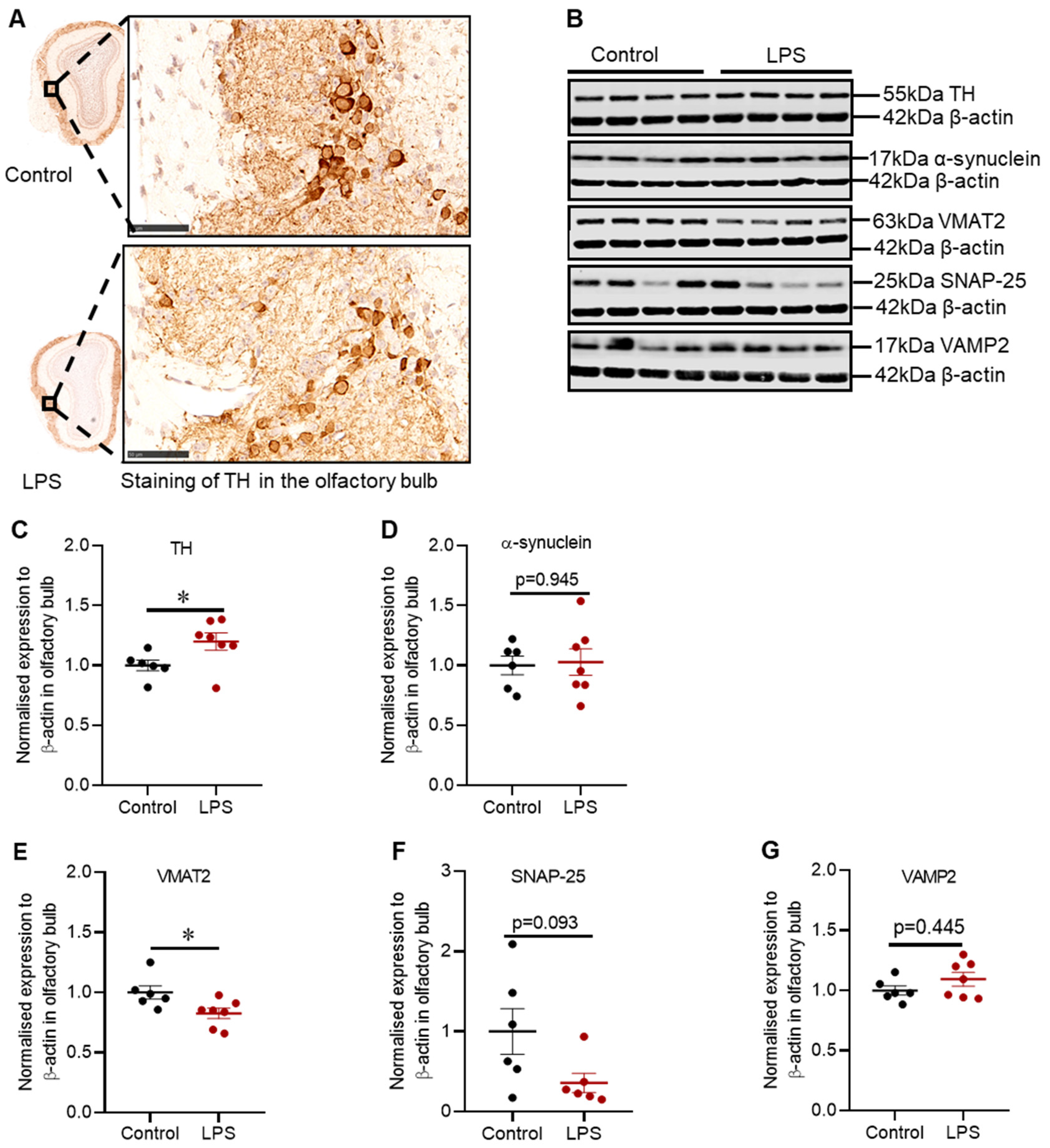
2.4. Intrastriatal Administration of LPS Altered the Expression of TH and Synaptic Proteins in the Olfactory Bulb of C57BL/6 Mice
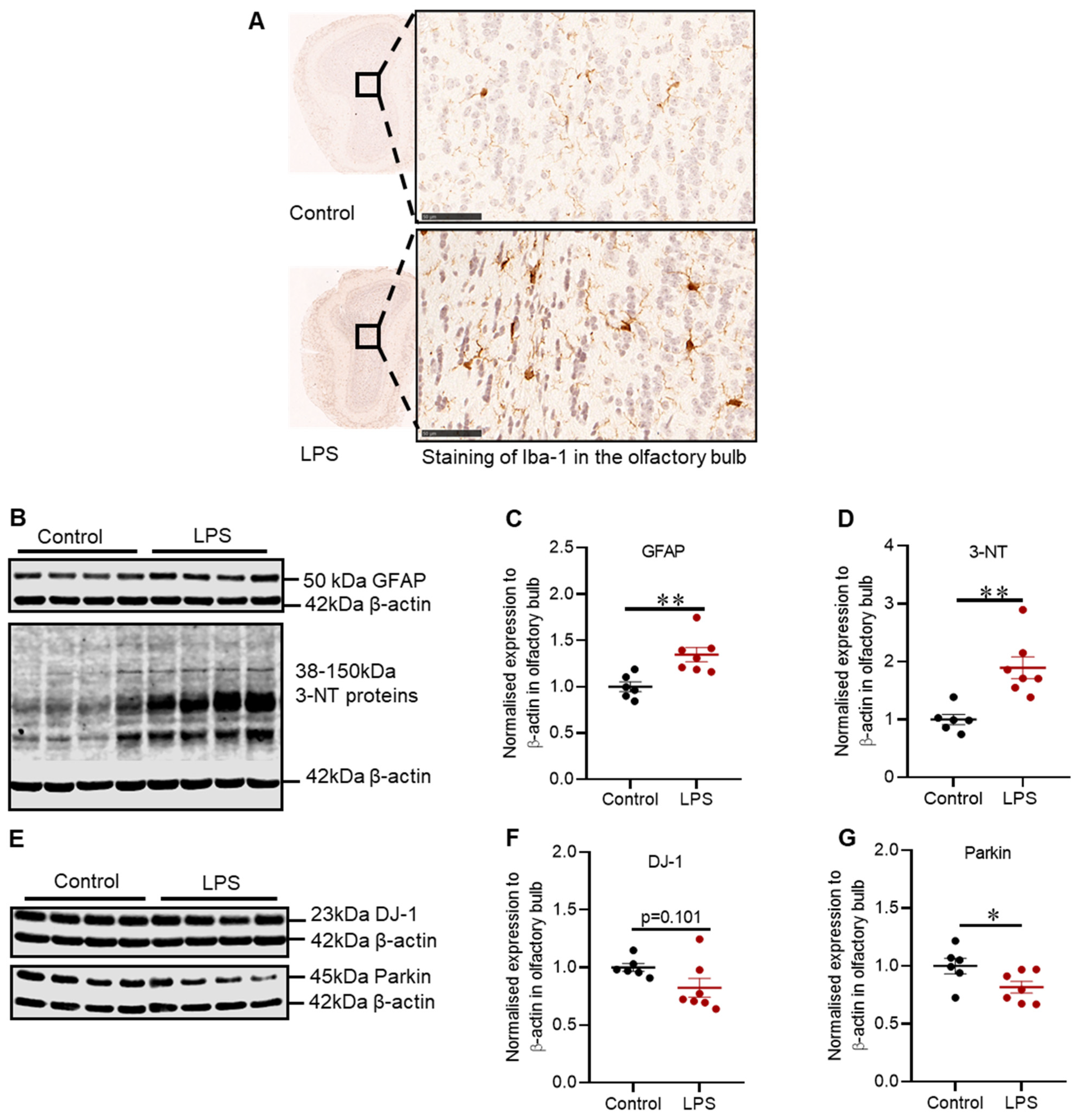
2.5. Effects of LPS on the Inflammatory and Oxidative Stress Markers, and Proteins Involved in Defence Mechanisms against Oxidative Stress in the Olfactory Bulb of C57BL/6 Mice
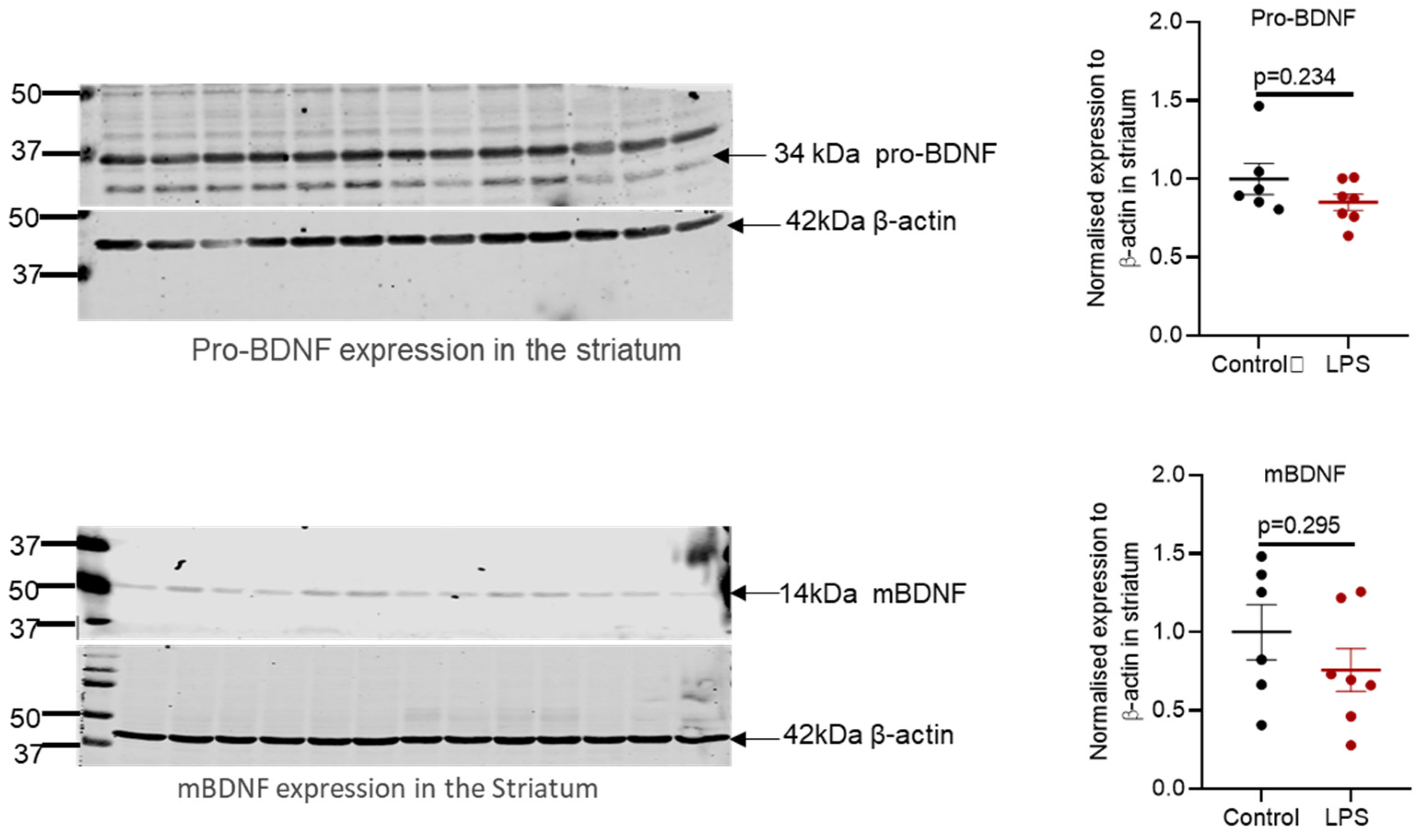
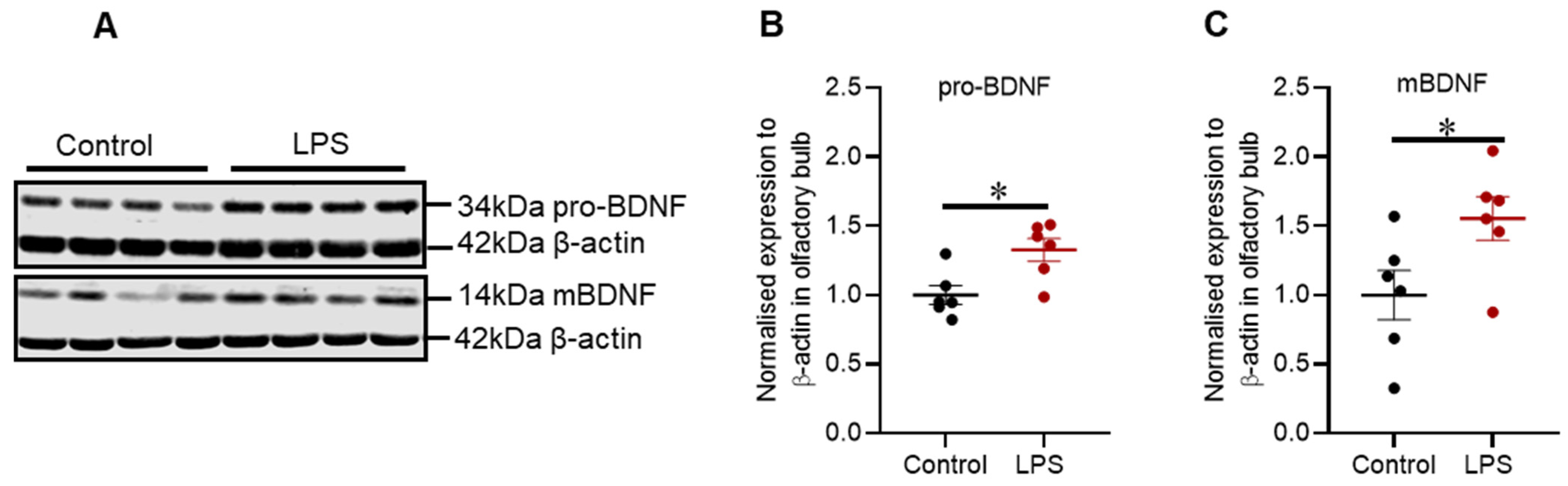
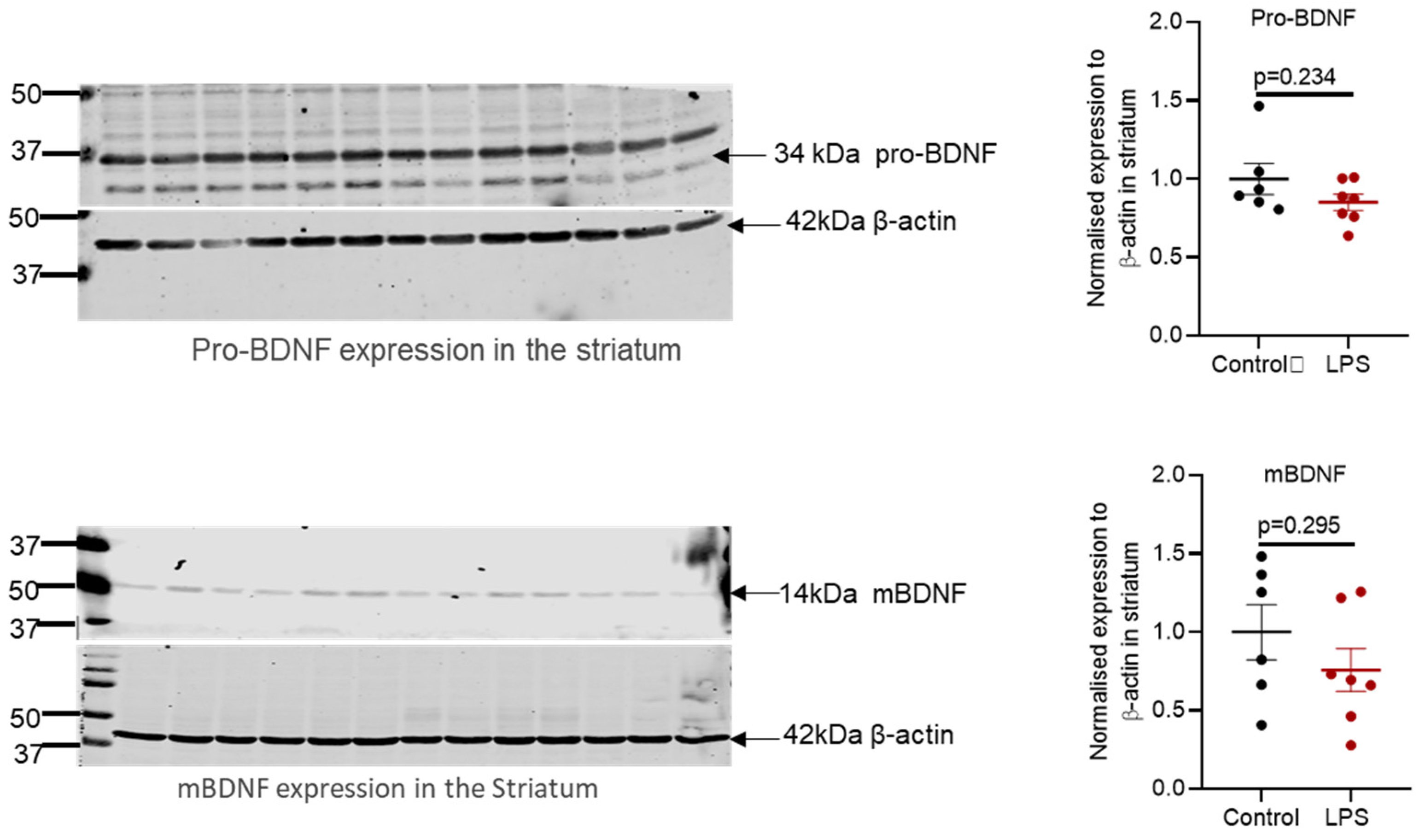
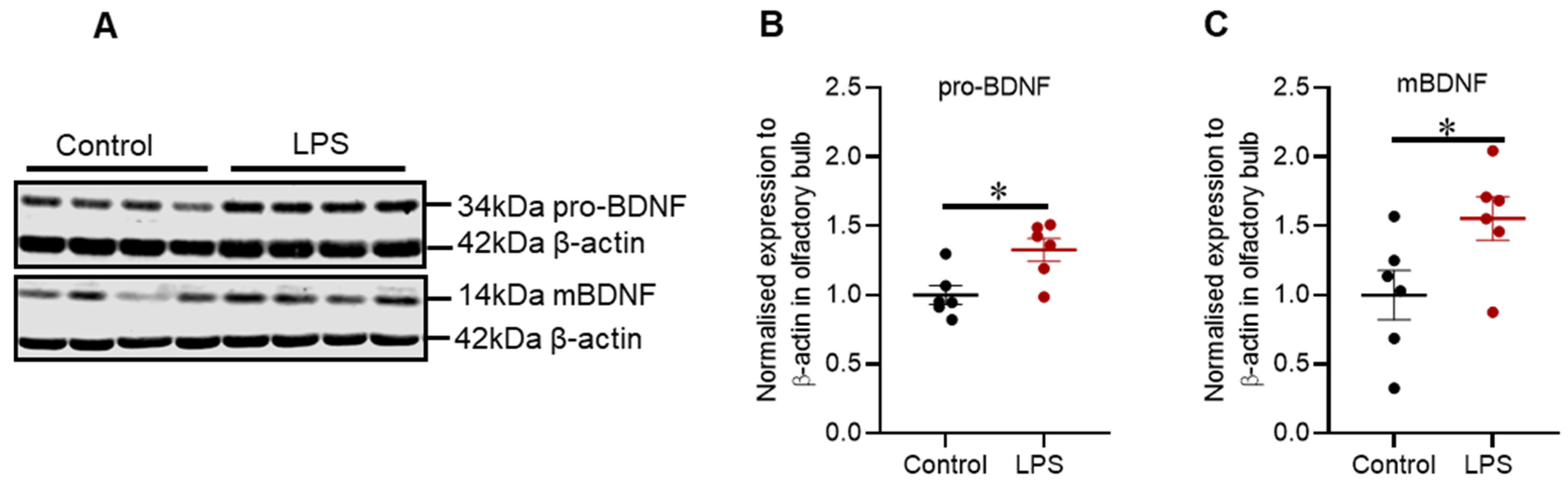
2.6. Intrastriatal Administration of LPS Altered the Expression of Brain-Derived Neurotrophic Factor Protein in the Olfactory Bulb but Not in the Striatum of C57BL/6 Mice
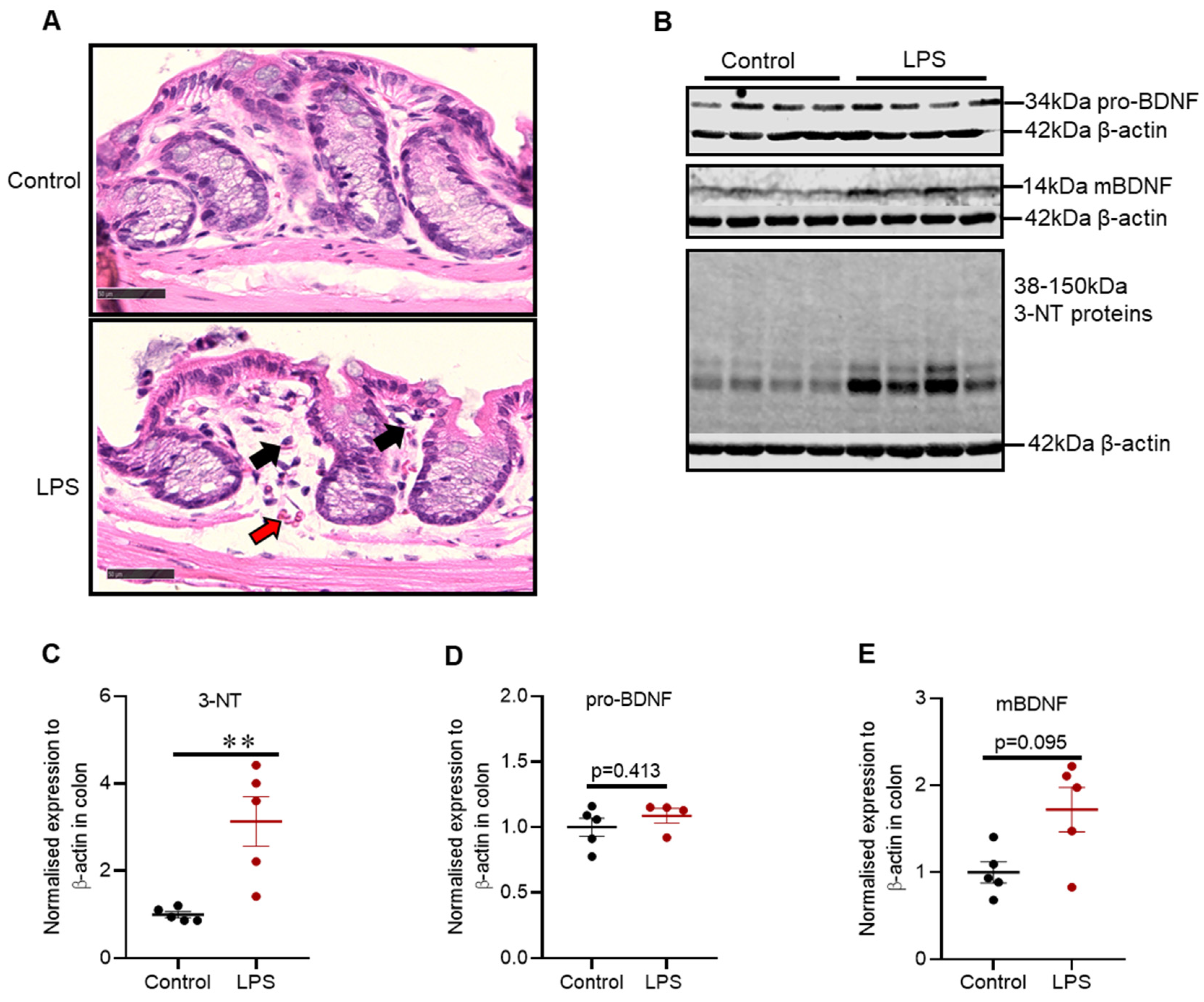
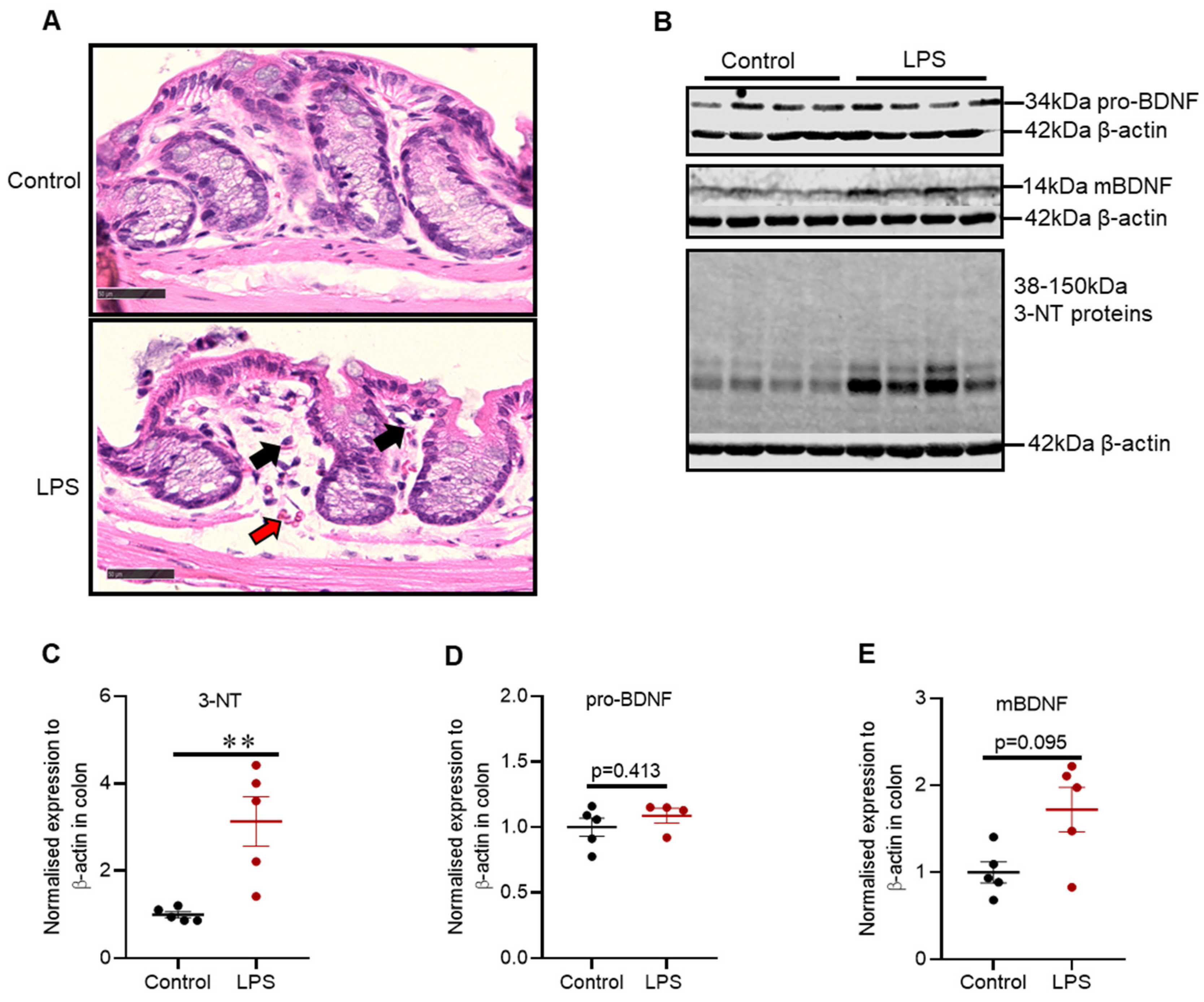
2.7. Intrastriatal Administration of LPS Induced Mild Inflammatory Changes and Alteration in Oxidative Stress Markers in the Distal Colon of C57BL/6 Mice
3. Discussion
3.1. Main Findings of the Study
3.2. Motor Function and Striatal Pathology
3.3. Olfactory Function and Pathology
3.4. Colonic Pathology
4. Materials and Methods
4.1. Animals
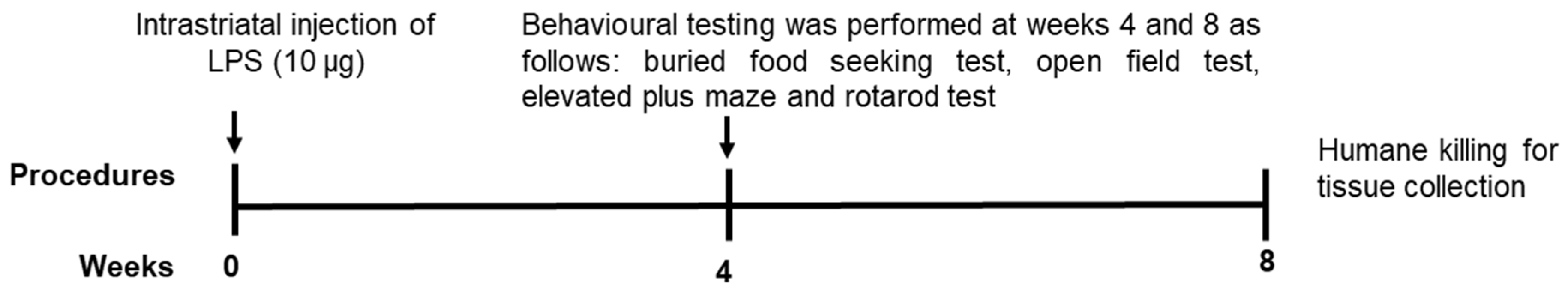
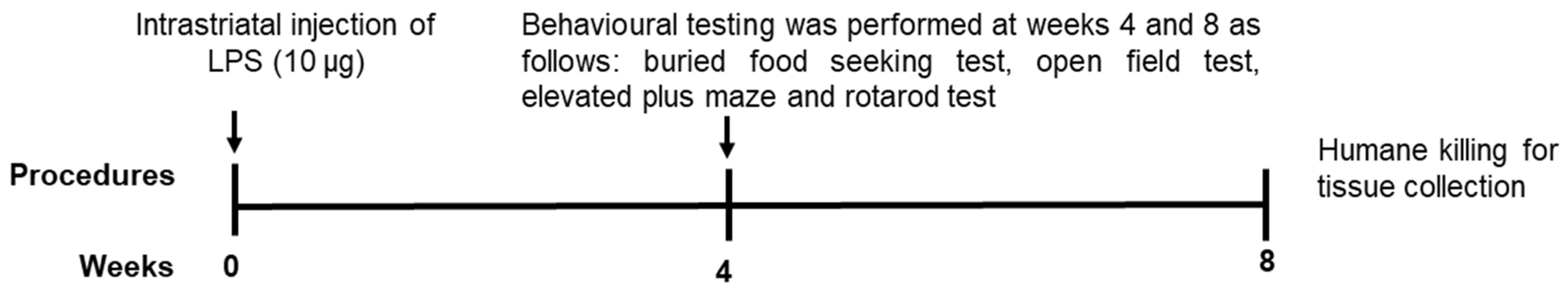
4.2. Experimental Design
4.3. Behavioural Testing
4.3.1. Buried Food-Seeking Test
4.3.2. Open Field Test
4.3.3. Elevated Plus Maze
4.3.4. Rotarod Test
4.4. Fresh Tissue Collection and Homogenisation
4.5. Western Blot
4.6. Immunohistochemistry
4.7. Hematoxylin and Eosin Staining for Distal Colon
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibodies | Company | Dilution |
|---|---|---|
| Mouse anti-3-nitrotyrosine (3-NT) | Abcam | 5% milk/TBST (1:3000) |
| Mouse anti-parkin | Santa Cruz Biotechnology | 5% BSA/TBST (1:500) |
| Mouse anti-protein deglycase DJ-1 | Santa Cruz Biotechnology | 5% BSA/TBST (1:1000) |
| Mouse anti-synaptosomal associated protein 25 (SNAP-25) | Santa Cruz Biotechnology | 5% BSA/TBST (1:1000) |
| Mouse anti-tyrosine hydroxylase (TH) | Sigma | 5% BSA/TBST (1:7000) |
| Mouse anti-vesicular monoamine transporter 2 (VMAT2) | Santa Cruz Biotechnology | 5% BSA/TBST (1:1000) |
| Rabbit anti-α-synuclein C-20 | Santa Cruz Biotechnology | 5% BSA/TBST (1:300) |
| Rabbit anti-glial fibrillary acidic protein (GFAP) | Dako | 5% milk/TBST (1:2000) |
| Rabbit brain-derived neurotrophic factor (BDNF) | Osenses Australia | 5% BSA/azide (1:1500) |
| Rabbit precursor brain derived neurotrophic factor (pro-BDNF) | Osenses Australia | 5% BSA/azide (1:3000) |
| Rabbit anti-vesicle associated membrane protein 2 (VAMP2) | Osenses Australia | 5% BSA/TBST (1:1000) |
| Mouse β-actin | Abcam | TBST (1:15 000) |
| Secondary antibodies | ||
| Donkey anti-rabbit IgG | Li-Cor Biosciences | TBST (1:20,000) |
| Goat anti-mouse IgG | Li-Cor Biosciences | TBST (1:20,000) |
References
- Jankovic, J. Parkinson’s disease: Clinical features and diagnosis. J. Neurol. Neurosurg. Psychiatry 2008, 79, 368–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sveinbjornsdottir, S. The clinical symptoms of Parkinson’s disease. J. Neurochem. 2016, 139, 318–324. [Google Scholar] [CrossRef] [Green Version]
- Obeso, J.A.; Rodriguez-Oroz, M.C.; Goetz, C.G.; Marin, C.; Kordower, J.H.; Rodriguez, M.; Hirsch, E.C.; Farrer, M.; Schapira, A.H.; Halliday, G. Missing pieces in the Parkinson’s disease puzzle. Nat. Med. 2010, 16, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Dawson, T.M.; Dawson, V.L. Molecular pathways of neurodegeneration in Parkinson’s disease. Science 2003, 302, 819–822. [Google Scholar] [CrossRef] [PubMed]
- Picconi, B.; Piccoli, G.; Calabresi, P. Synaptic dysfunction in Parkinson’s disease. Adv. Exp. Med. Biol. 2012, 970, 553–572. [Google Scholar] [CrossRef] [PubMed]
- Emamzadeh, F.N. Alpha-synuclein structure, functions, and interactions. J. Res. Med. Sci. 2016, 21, 29. [Google Scholar] [CrossRef]
- Kim, K.; Wi, S.; Seo, J.H.; Pyo, S.; Cho, S.-R. Reduced interaction of aggregated α-synuclein and vamp2 by environmental enrichment alleviates hyperactivity and anxiety in a model of Parkinson’s disease. Genes 2021, 12, 392. [Google Scholar] [CrossRef]
- Yasuda, T.; Nakata, Y.; Choong, C.-J.; Mochizuki, H. Neurodegenerative changes initiated by presynaptic dysfunction. Transl. Neurodegener. 2013, 2, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Agliardi, C.; Meloni, M.; Guerini, F.R.; Zanzottera, M.; Bolognesi, E.; Baglio, F.; Clerici, M. Oligomeric α-syn and snare complex proteins in peripheral extracellular vesicles of neural origin are biomarkers for Parkinson’s disease. Neurobiol. Dis. 2021, 148, 105185. [Google Scholar] [CrossRef]
- Phan, J.-A.; Stokholm, K.; Zareba-Paslawska, J.; Jakobsen, S.; Vang, K.; Gjedde, A.; Landau, A.M.; Romero-Ramos, M. Early synaptic dysfunction induced by α-synuclein in a rat model of parkinson’s disease. Sci. Rep. 2017, 7, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, I.-T.; Weng, Y.-H.; Hsieh, C.-J.; Lin, W.-Y.; Wey, S.-P.; Kung, M.-P.; Yen, T.-C.; Lu, C.-S.; Lin, K.-J. Correlation of parkinson disease severity and 18f-dtbz positron emission tomography. JAMA Neurol. 2014, 71, 758–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohr, K.M.; Miller, G.W. Vmat2 and Parkinson’s disease: Harnessing the dopamine vesicle. Expert. Rev. Neurother. 2014, 14, 1115–1117. [Google Scholar] [CrossRef] [Green Version]
- Taylor, T.N.; Caudle, W.M.; Miller, G.W. Vmat2-deficient mice display nigral and extranigral pathology and motor and nonmotor symptoms of parkinson’s Disease. Parkinson’s Dis. 2011, 2011, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Reeve, A.K.; Grady, J.P.; Cosgrave, E.M.; Bennison, E.; Chen, C.; Hepplewhite, P.D.; Morris, C.M. Mitochondrial dysfunction within the synapses of substantia nigra neurons in parkinson’s disease. NPJ Parkinson’s Dis. 2018, 4, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Duty, S.; Jenner, P. Animal models of Parkinson’s disease: A source of novel treatments and clues to the cause of the disease. Br. J. Pharmacol. 2011, 164, 1357–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGeer, P.L.; Itagaki, S.; Boyes, B.E.; McGeer, E.G. Reactive microglia are positive for hla-dr in the substantia nigra of Parkinson’s and alzheimer’s disease brains. Neurol 1988, 38, 1285–1291. [Google Scholar] [CrossRef]
- Tansey, M.G.; Goldberg, M.S. Neuroinflammation in Parkinson’s disease: Its role in neuronal death and implications for therapeutic intervention. Neurobiol. Dis. 2010, 37, 510–518. [Google Scholar] [CrossRef] [Green Version]
- Sofroniew, M.V. Astrogliosis. Cold Spring Harb. Perspect. Biol. 2015, 7, a020420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boje, K.M.; Arora, P.K. Microglial-produced nitric oxide and reactive nitrogen oxides mediate neuronal cell death. Brain. Res. 1992, 587, 250–256. [Google Scholar] [CrossRef]
- Chao, C.C.; Hu, S.; Molitor, T.W.; Shaskan, E.G.; Peterson, P.K. Activated microglia mediate neuronal cell injury via a nitric oxide mechanism. J. Immunol. 1992, 149, 2736–2741. [Google Scholar] [PubMed]
- Giacobbo, B.L.; Doorduin, J.; Klein, H.C.; Dierckx, R.A.; Bromberg, E.; de Vries, E.F. Brain-derived neurotrophic factor in brain disorders: Focus on neuroinflammation. Mol. Neurobiol. 2019, 56, 3295–3312. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Yun, W.; Zhang, M.; Luo, W.; Zhou, X. Serum concentration and clinical significance of brain-derived neurotrophic factor in patients with parkinson’s disease or essential tremor. J. Int. Med. Res. 2018, 46, 1477–1485. [Google Scholar] [CrossRef] [Green Version]
- Devos, D.; Lebouvier, T.; Lardeux, B.; Biraud, M.; Rouaud, T.; Pouclet, H.; Coron, E.; des Varannes, S.B.; Naveilhan, P.; Nguyen, J.-M. Colonic inflammation in Parkinson’s disease. Neurobiol. Dis. 2013, 50, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Keshavarzian, A.; Green, S.J.; Engen, P.A.; Voigt, R.M.; Naqib, A.; Forsyth, C.B.; Mutlu, E.; Shannon, K.M. Colonic bacterial composition in Parkinson’s disease. Mov. Disord. 2015, 30, 1351–1360. [Google Scholar] [CrossRef] [PubMed]
- van IJzendoorn, S.C.; Derkinderen, P. The intestinal barrier in parkinson’s disease: Current state of knowledge. J. Parkinson’s Dis. 2019, 9, S323–S329. [Google Scholar] [CrossRef] [Green Version]
- Wakabayashi, K.; Takahashi, H.; Takeda, S.; Ohama, E.; Ikuta, F. Parkinson’s disease: The presence of lewy bodies in auerbach’s and meissner’s plexuses. Acta Neuropathol. 1988, 76, 217–221. [Google Scholar] [CrossRef]
- Campos-Acuña, J.; Elgueta, D.; Pacheco, R. T-cell-driven inflammation as a mediator of the gut-brain axis involved in Parkinson’s disease. Front. Immunol. 2019, 10, 239. [Google Scholar] [CrossRef] [Green Version]
- Villumsen, M.; Aznar, S.; Pakkenberg, B.; Jess, T.; Brudek, T. Inflammatory bowel disease increases the risk of parkinson’s disease: A danish nationwide cohort study 1977–2014. Gut 2019, 68, 18–24. [Google Scholar] [CrossRef]
- Deng, I.; Corrigan, F.; Zhai, G.; Zhou, X.-F.; Bobrovskaya, L. Lipopolysaccharide animal models of parkinson’s disease: Recent progress and relevance to clinical disease. Brain Behav. Immun. Health 2020, 4, 100060. [Google Scholar] [CrossRef]
- Wang, J.; He, C.; Wu, W.-Y.; Chen, F.; Wu, Y.-Y.; Li, W.-Z.; Chen, H.-Q.; Yin, Y.-Y. Biochanin a protects dopaminergic neurons against lipopolysaccharide-induced damage and oxidative stress in a rat model of Parkinson’s disease. Pharmacol. Biochem. Behav. 2015, 138, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.-Y.; Liu, M.; Hunter, R.L.; Cass, W.A.; Pandya, J.D.; Sullivan, P.G.; Shin, E.-J.; Kim, H.-C.; Gash, D.M.; Bing, G. Striatal neuroinflammation promotes parkinsonism in rats. PLoS ONE 2009, 4, e5482. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R.L.; Cheng, B.; Choi, D.Y.; Liu, M.; Liu, S.; Cass, W.A.; Bing, G. Intrastriatal lipopolysaccharide injection induces parkinsonism in c57/b6 mice. J. Neurosci. Res. 2009, 87, 1913–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Q.; Li, Y.h.; Guo, S.s.; Wang, Y.; Lin, W.; Zhang, Q.; Wang, J.; Ma, C.g.; Xiao, B.G. Inhibition of rho-kinase by fasudil protects dopamine neurons and attenuates inflammatory response in an intranasal lipopolysaccharide-mediated Parkinson’s model. Eur. J. Neurosci. 2016, 43, 41–52. [Google Scholar] [CrossRef]
- Beier, E.E.; Neal, M.; Alam, G.; Edler, M.; Wu, L.-J.; Richardson, J.R. Alternative microglial activation is associated with cessation of progressive dopamine neuron loss in mice systemically administered lipopolysaccharide. Neurobiol. Dis. 2017, 108, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Bodea, L.-G.; Wang, Y.; Linnartz-Gerlach, B.; Kopatz, J.; Sinkkonen, L.; Musgrove, R.; Kaoma, T.; Muller, A.; Vallar, L.; Di Monte, D.A.; et al. Neurodegeneration by activation of the microglial complement–phagosome pathway. J. Neurosci. 2014, 34, 8546–8556. [Google Scholar] [CrossRef] [Green Version]
- Nordström, U.; Beauvais, G.; Ghosh, A.; Sasidharan, B.C.P.; Lundblad, M.; Fuchs, J.; Joshi, R.L.; Lipton, J.W.; Roholt, A.; Medicetty, S.; et al. Progressive nigrostriatal terminal dysfunction and degeneration in the engrailed1 heterozygous mouse model of Parkinson’s disease. Neurobiol. Dis. 2015, 73, 70–82. [Google Scholar] [CrossRef] [Green Version]
- Hunter, R.L.; Dragicevic, N.; Seifert, K.; Choi, D.Y.; Liu, M.; Kim, H.-C.; Cass, W.A.; Sullivan, P.G.; Bing, G. Inflammation induces mitochondrial dysfunction and dopaminergic neurodegeneration in the nigrostriatal system. J. Neurochem. 2007, 100, 1375–1386. [Google Scholar] [CrossRef]
- Zhao, X.; Zhai, S.; An, M.-S.; Wang, Y.-H.; Yang, Y.-F.; Ge, H.-Q.; Liu, J.-H.; Pu, X.-P. Neuroprotective Effects of Protocatechuic Aldehyde against Neurotoxin-Induced Cellular and Animal Models of Parkinson’s Disease. PLoS ONE 2013, 8, e78220. [Google Scholar] [CrossRef] [PubMed]
- Valle-Leija, P.; Drucker-Colín, R. Unilateral olfactory deficit in a hemiparkinson’s disease mouse model. NeuroReport 2014, 25, 948–953. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Han, C.; Zhang, G.; Guo, X.; Xia, Y.; Wan, F.; Yin, S.; Kou, L.; Liu, L.; Huang, J.; et al. Reduced VMAT2 expression exacerbates the hyposmia in the MPTP model of Parkinson’s disease. Biochem. Biophys. Res. Commun. 2019, 513, 306–312. [Google Scholar] [CrossRef]
- Xu, L.; Pu, J. Alpha-Synuclein in Parkinson’s Disease: From Pathogenetic Dysfunction to Potential Clinical Application. Park. Dis. 2016, 2016, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Shi, M.; Chung, K.A.; Zabetian, C.P.; Leverenz, J.B.; Berg, D.; Srulijes, K.; Trojanowski, J.Q.; Lee, V.M.-Y.; Siderowf, A.D.; et al. Phosphorylated α-synuclein in parkinson’s disease. Sci. Transl. Med. 2012, 4, 120–121. [Google Scholar] [CrossRef] [Green Version]
- Doty, R.L.; Deems, D.A.; Stellar, S. Olfactory dysfunction in parkinsonism: A general deficit unrelated to neurologic signs, disease stage, or disease duration. Neurol 1988, 38, 1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huisman, E.; Uylings, H.B.; Hoogland, P.V. A 100% increase of dopaminergic cells in the olfactory bulb may explain hyposmia in Parkinson’s disease. Mov. Disord. Off. J. Mov. Disord. Soc. 2004, 19, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Pignatelli, A.; Kobayashi, K.; Okano, H.; Belluzzi, O. Functional properties of dopaminergic neurones in the mouse olfactory bulb. J. Physiol. 2005, 564, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Mundinano, I.-C.; Caballero, M.-C.; Ordóñez, C.; Hernandez, M.; DiCaudo, C.; Marcilla, I.; Erro, M.-E.; Tuñon, M.-T.; Luquin, M.-R. Increased dopaminergic cells and protein aggregates in the olfactory bulb of patients with neurodegenerative disorders. Acta Neuropathol. 2011, 122, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Winner, B.; Desplats, P.; Hagl, C.; Klucken, J.; Aigner, R.; Ploetz, S.; Lämke, J.; Karl, A.; Aigner, L.; Masliah, E.; et al. Dopamine receptor activation promotes adult neurogenesis in an acute Parkinson model. Exp. Neurol. 2009, 219, 543–552. [Google Scholar] [CrossRef] [Green Version]
- Sala, G.; Brighina, L.; Saracchi, E.; Fermi, S.; Riva, C.; Carrozza, V.; Pirovano, M.; Ferrarese, C. Vesicular monoamine transporter 2 mRNA levels are reduced in platelets from patients with Parkinson’s disease. J. Neural Transm. 2010, 117, 1093–1098. [Google Scholar] [CrossRef]
- Taylor, T.N.; Greene, J.G.; Miller, G.W. Behavioral phenotyping of mouse models of Parkinson’s disease. Behav. Brain Res. 2010, 211, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Stoker, T.B.; Greenland, J.C. Parkinson’s Disease: Pathogenesis and Clinical Aspects; Codon Publications: Singapore, 2018. [Google Scholar] [CrossRef]
- Parashar, A.; Udayabanu, M. Gut microbiota: Implications in Parkinson’s disease. Park. Relat. Disord. 2017, 38, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Gil, P.; Rodriguez-Perez, A.I.; Dominguez-Meijide, A.; Guerra, M.J.; Labandeira-Garcia, J.L. Bidirectional Neural Interaction Between Central Dopaminergic and Gut Lesions in Parkinson’s Disease Models. Mol. Neurobiol. 2018, 55, 7297–7316. [Google Scholar] [CrossRef]
- Chen, Q.-Q.; Haikal, C.; Li, W.; Li, J.-Y. Gut Inflammation in Association with Pathogenesis of Parkinson’s Disease. Front. Mol. Neurosci. 2019, 12, 218. [Google Scholar] [CrossRef] [Green Version]
- Konturek, T.J.; Martinez, C.; Niesler, B.; Van Der Voort, I.; Mönnikes, H.; Stengel, A.; Goebel-Stengel, M. The Role of Brain-Derived Neurotrophic Factor in Irritable Bowel Syndrome. Front. Psychiatry 2021, 11, 1385. [Google Scholar] [CrossRef] [PubMed]
- Titova, N.; Schapira, A.H.; Chaudhuri, K.R.; Qamar, M.A.; Katunina, E.; Jenner, P. Nonmotor Symptoms in Experimental Models of Parkinson’s Disease. Int. Rev. Neurobiol. 2017, 133, 63–89. [Google Scholar] [CrossRef] [PubMed]
- Machado, C.F.; Reis-Silva, T.M.; Lyra, C.S.; Felicio, L.F.; Malnic, B. Buried food-seeking test for the assessment of olfactory detection in mice. Bioprotocol 2018, 8, e2897. [Google Scholar]
- Zhang, W.; Sun, C.; Shao, Y.; Zhou, Z.; Hou, Y.; Li, A. Partial depletion of dopaminergic neurons in the substantia nigra impairs olfaction and alters neural activity in the olfactory bulb. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Prut, L.; Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: A review. Eur. J. Pharmacol. 2003, 463, 3–33. [Google Scholar] [CrossRef]
- Seibenhener, M.L.; Wooten, M.C. Use of the open field maze to measure locomotor and anxiety-like behavior in mice. J. Vis. Exp. 2015, 96, e52434. [Google Scholar] [CrossRef] [Green Version]
- Walf, A.A.; Frye, A.C. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef] [Green Version]
- Thornton, E.; Vink, R. Treatment with a substance P receptor antagonist is neuroprotective in the intrastriatal 6-hydroxydopamine model of early Parkinson’s disease. PLoS ONE 2012, 7, e34138. [Google Scholar]
- Rozas, G.; Guerra, M.J.; Labandeira-Garcı́a, J.L. An automated rotarod method for quantitative drug-free evaluation of overall motor deficits in rat models of parkinsonism. Brain Res. Protoc. 1997, 2, 75–84. [Google Scholar] [CrossRef]
- Johnson, M.E.; Zhou, X.-F.; Bobrovskaya, L. The effects of rotenone on TH, BDNF and BDNF-related proteins in the brain and periphery: Relevance to early Parkinson’s disease. J. Chem. Neuroanat. 2019, 97, 23–32. [Google Scholar] [CrossRef]

| Changes in Protein Expression | |||
|---|---|---|---|
| Proteins | Striatum | Olfactory Bulb | Colon |
| TH | ↓ | ↑ | - |
| α-syn | ↑ | ↔ | - |
| VMAT2 | ↓ | ↓ | - |
| SNAP-25 | ↓ | ↔ | - |
| VAMP2 | ↔ | ↔ | - |
| GFAP | ↑ | ↑↑ | - |
| DJ-1 | ↓ | ↔ | - |
| Parkin | ↔ | ↓ | - |
| 3-NT proteins | ↔ | ↑↑ | ↑↑ |
| Pro-BDNF | ↔ | ↑ | ↔ |
| mBDNF | ↔ | ↑ | ↔ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, I.; Corrigan, F.; Garg, S.; Zhou, X.-F.; Bobrovskaya, L. Further Characterization of Intrastriatal Lipopolysaccharide Model of Parkinson’s Disease in C57BL/6 Mice. Int. J. Mol. Sci. 2021, 22, 7380. https://doi.org/10.3390/ijms22147380
Deng I, Corrigan F, Garg S, Zhou X-F, Bobrovskaya L. Further Characterization of Intrastriatal Lipopolysaccharide Model of Parkinson’s Disease in C57BL/6 Mice. International Journal of Molecular Sciences. 2021; 22(14):7380. https://doi.org/10.3390/ijms22147380
Chicago/Turabian StyleDeng, Isaac, Frances Corrigan, Sanjay Garg, Xin-Fu Zhou, and Larisa Bobrovskaya. 2021. "Further Characterization of Intrastriatal Lipopolysaccharide Model of Parkinson’s Disease in C57BL/6 Mice" International Journal of Molecular Sciences 22, no. 14: 7380. https://doi.org/10.3390/ijms22147380
APA StyleDeng, I., Corrigan, F., Garg, S., Zhou, X.-F., & Bobrovskaya, L. (2021). Further Characterization of Intrastriatal Lipopolysaccharide Model of Parkinson’s Disease in C57BL/6 Mice. International Journal of Molecular Sciences, 22(14), 7380. https://doi.org/10.3390/ijms22147380