
Activity of Selected Group of Monoterpenes in Alzheimer’s Disease Symptoms in Experimental Model Studies—A Non-Systematic Review



 ,
,  and
and
Abstract
1. Introduction
2. Pathophysiology of Alzheimer’s Disease
2.1. β-Amyloid
2.2. Tau Protein
2.3. Oxidative Stress
2.4. Inflammation
2.5. Apolipoprotein E
2.6. The Cholinergic System
2.7. Glutamatergic System
3. Therapeutic Potential of Monoterpenes for Alzheimer’s Disease
3.1. Outline of Mechanisms Involved in the Anti-AD Action of Monoterpenes and Monoterpenoids
3.2. Monoterpenes and Monoterpenoids in the Treatment of AD—Possible Mechanisms of Action
3.3. Iridoids in the Treatment of AD
3.4. Secoiridoids in the Treatment of AD
4. Memory and Learning Improvement
4.1. In Vitro and In Silico Studies Results
4.2. In Vivo Studies Results
5. Anxiolytic Effects
6. Insomnia
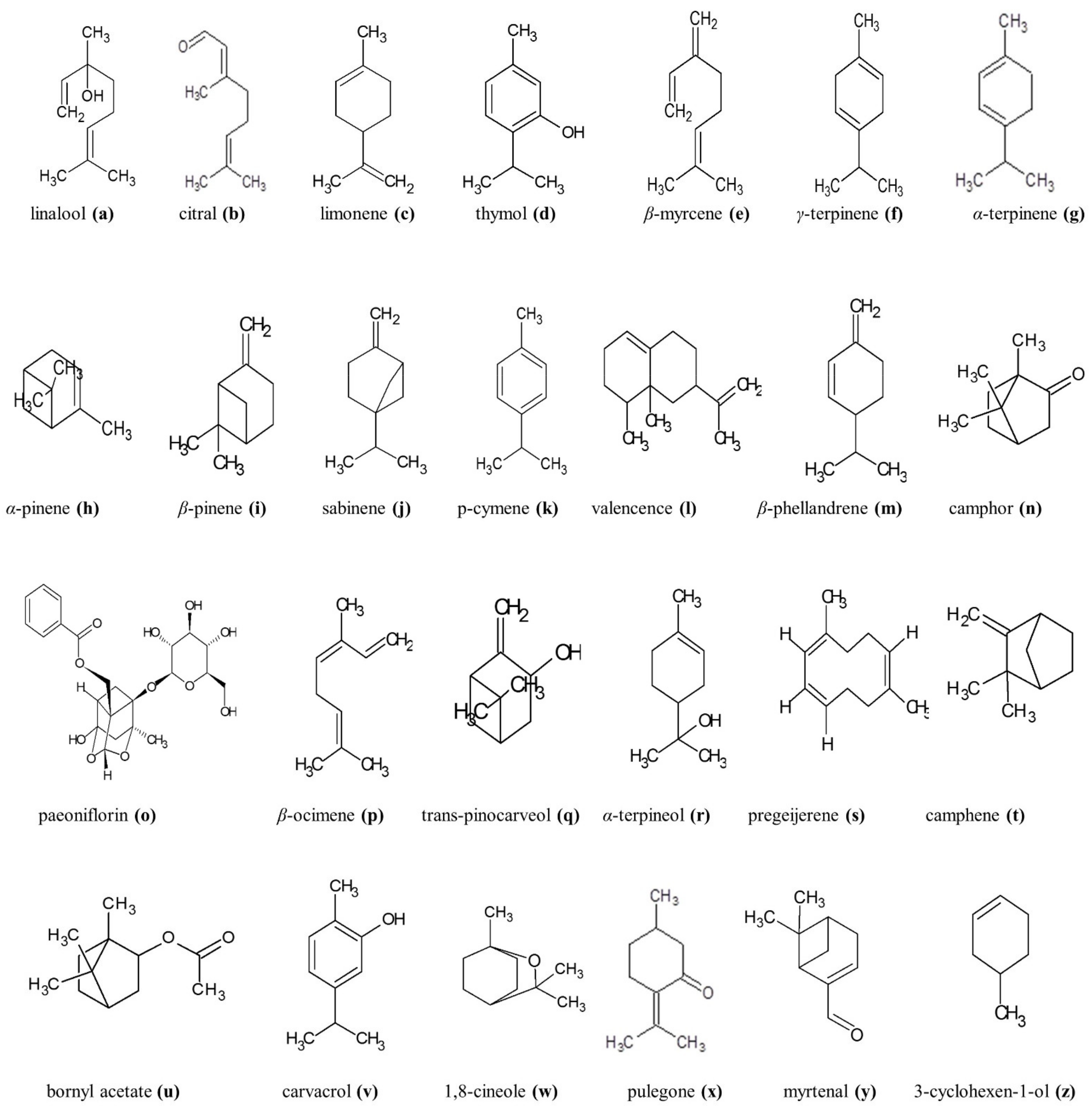
Monoterpenes
7. Outline of Polyphenols and Essential Oils Rich in Monoterpenes and Polyphenols Revealing Neuroprotective Effect
7.1. In Vitro Studies Results
7.2. In Vivo Studies Results
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Aβ | amyloid β |
| ABCA-1 | ATP-binding cassette transporter-A1 |
| ACh | acetylcholine |
| AChE | acetylcholinesterase |
| AD | Alzheimer’s disease |
| AICD | intracellular domain CTFβ |
| AMPK | AMP-activated protein kinase |
| ApoE | apolipoprotein E |
| APP | amyloid β precursor protein |
| BACE1 | β-secretase 1 |
| BBB | blood-brain barrier |
| BChE | butyrylcholinesterase |
| BDNF | brain-derived neurotrophic factor |
| ChAT | choline acetyltrasferase |
| COX | cyclooxygenase |
| ECE | endothelin converting enzymes |
| ER | endoplasmic reticulum |
| ERK | extracellular signal–regulated kinase |
| CREB | cAMP-response element binding protein |
| GAD | generalized anxiety disorder |
| GAP-43 | growth associated protein 43 |
| GFAP | glial fibrillary acidic protein |
| GLP-1R | glucagon-like peptide-1 receptor |
| GLT-1 | glutamine transporter-1 |
| GLUT-1 | glucose transporter-1 |
| GPx | glutathione peroxidase |
| GSHPx | glutathione peroxidase |
| GSH-ST | glutathione S-transferase |
| HFO | hogh fat diet |
| hptau | hyperphosphorylated tau protein |
| IDE | insulin degrading enzyme |
| IκB | nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor |
| IL-1 | interleukin-1 |
| JNK | c-Jun N-terminal kinase |
| LC3II | lysosomal cysteine protease 3II |
| LRP-1 | lipoprotein receptor-related protein 1 |
| mAChR | muscarinic acetylcholine receptors |
| MAPK | mitogen-activated protein kinase |
| mTOR | mammalian target of rapamycin |
| nAChR | nicotinic acetylcholine receptors |
| NEP | neprilysin |
| NF-κB | nuclear factor kappa light chain enhancer of activated B cells |
| NFTs | neurofibrillary tangles |
| NGF | nerve growth factor |
| NMDA | N-methyl-D-aspartic acid |
| NOS | nitric oxide synthase |
| PGE | prostaglandin |
| PHF | paired helical filaments |
| PKC | protein kinase C |
| PS | presenilin |
| PSD-95 | postsynaptic density protein 95 |
| PTEN | phosphatase and tensin homolog |
| RAGE | receptor for advanced glycation endproducts |
| ROS | reactive oxygen species |
| SERT | serotonin transporter |
| SNAP-25 | synaptosomal nerve-associated protein-25 |
| SOD | superoxide dismutase |
| SWS | slow wave sleep |
| TACE | α-secretase |
| TNF | tumor necrozing factor |
| TrkB | tropomyosin receptor kinase B |
References
- Wojtunik-Kulesza, K.; Oniszczuk, A.; Waksmundzka-Hajnos, M. An attempt to elucidate the role of iron and zinc ions in development of Alzheimer’s and Parkinson’s diseases. Biomed. Pharmacother. 2019, 111, 1277–1289. [Google Scholar] [CrossRef] [PubMed]
- Kulshreshtha, A.; Piplani, P. Current pharmacotherapy and putative disease-modifying therapy for Alzheimer’s disease. Neurol. Sci. Off. J. Ital. Neurol. Soc. Ital. Soc. Clin. Neurophysiol. 2016, 37, 1403–1435. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, L.; Achor, S.; Allen, B.; Bauchmire, N.; Dunnington, D.; Klisovic, R.B.; Naber, S.J.; Roblee, K.; Samczak, A.; Tomlinson-Pinkham, K.; et al. The effect of aromatherapy on insomnia and other common symptoms among patients with acute leukemia. Oncol. Nurs. Forum 2017, 44, E185–E193. [Google Scholar] [CrossRef]
- Da Silva, G.L.; Luft, C.; Lunardelli, A.; Amaral, R.H.; da Silva Melo, D.A.; Donadio, M.V.F.; Nunes, F.B.; de Azambuja, M.S.; Santana, J.C.; Moraes, C.M.B.; et al. Antioxidant, analgesic and anti-inflammatory effects of lavender essential oil. An. Acad. Bras. Cienc. 2015, 87, 1397–1408. [Google Scholar] [CrossRef]
- Popova, A.; Dalemska, Z.; Mihaylova, D.; Hristova, I.; Alexieva, I. Melissa officinalis L.- GC Profile and Antioxidant Activity. Int. J. Pharmacogn. Phytochem. Res. 2016, 4, 634–638. [Google Scholar]
- Wojtunik-Kulesza, K.A.; Kasprzak, K.; Oniszczuk, T.; Oniszczuk, A. Natural Monoterpenes: Much More than Only a Scent. Chem. Biodivers. 2019, 16, e1900434. [Google Scholar] [CrossRef]
- Maggio, A.; Rosselli, S.; Bruno, M. Essential oils and pure volatile compounds as potential drugs in Alzheimer’s disease therapy: An updated review of the literature. Curr. Pharm. Des. 2016, 22, 4011–4027. [Google Scholar] [CrossRef]
- Adorjan, B.; Buchbauer, G. Biological properties of essential oils: An updated review. Flavour Fragr. J. 2010, 25, 407–426. [Google Scholar] [CrossRef]
- Chen, J.; Jiang, Q.-D.; Chai, Y.-P.; Zhang, H.; Peng, P.; Yang, X.-X. Natural terpenes as penetration enhancers for transdermal drug delivery. Molecules 2016, 21, 1709. [Google Scholar] [CrossRef]
- Wippold, F.J., II; Cairns, N.; Vo, K.; Holtzman, D.M.; Morris, J.C. Neuropathology for the neuroradiologist: Plaques and tangles. Am. J. Neuroradiol. 2008, 29, 18–22. [Google Scholar] [CrossRef]
- Yankner, B.A.; Caceres, A.; Duffy, L.K. Nerve growth factor potentiates the neurotoxicity of β amyloid. Proc. Natl. Acad. Sci. USA 1990, 87, 9020–9023. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef]
- Duce, J.A.; Tsatsanis, A.; Cater, M.A.; James, S.A.; Robb, E.; Wikhe, K.; Leong, S.L.; Perez, K.; Johanssen, T.; Greenough, M.A.; et al. Iron-export ferroxidase activity of β-amyloid precursor protein is inhibited by zinc in Alzheimer’s disease. Cell 2010, 142, 857–867. [Google Scholar] [CrossRef] [PubMed]
- Priller, C.; Bauer, T.; Mitteregger, G.; Krebs, B.; Kretzschmar, H.A.; Herms, J. Synapse formation and function is modulated by the amyloid precursor protein. J. Neurosci. Off. J. Soc. Neurosci. 2006, 26, 7212–7221. [Google Scholar] [CrossRef]
- Isbert, S.; Wagner, K.; Eggert, S.; Schweitzer, A.; Multhaup, G.; Weggen, S.; Kins, S.; Pietrzik, C.U. APP dimer formation is initiated in the endoplasmic reticulum and differs between APP isoforms. Cell. Mol. Life Sci. 2012, 69, 1353–1375. [Google Scholar] [CrossRef][Green Version]
- Abeysinghe, G.; Kuchira, M.; Kudo, G.; Masuo, S.; Ninomiya, A.; Takahashi, K.; Utad, A.S.; Hagiwara, D.; Nomura, N.; Takaya, N.; et al. Fungal mycelia and bacterial thiamine establish a mutualistic growth mechanism. Life Sci. Alliance 2020, 3. [Google Scholar] [CrossRef]
- Godyń, J.; Jończyk, J.; Panek, D.; Malawska, B. Therapeutic strategies for Alzheimer’s disease in clinical trials. Pharmacol. Rep. 2016, 68, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.-S.; Dickson, D.W.; Malter, J.S. beta-Amyloid degradation and Alzheimer’s disease. J. Biomed. Biotechnol. 2006, 2006, 58406. [Google Scholar] [CrossRef]
- Winkler, E.; Kamp, F.; Scheuring, J.; Ebke, A.; Fukumori, A.; Steiner, H. Generation of Alzheimer disease-associated amyloid β42/43 peptide by γ-secretase can be inhibited directly by modulation of membrane thickness. J. Biol. Chem. 2012, 287, 21326–21334. [Google Scholar] [CrossRef] [PubMed]
- Lewczuk, P.; Kamrowski-Kruck, H.; Peters, O.; Heuser, I.; Jessen, F.; Popp, J.; Bürger, K.; Hampel, H.; Frölich, L.; Wolf, S.; et al. Soluble amyloid precursor proteins in the cerebrospinal fluid as novel potential biomarkers of Alzheimer’s disease: A multicenter study. Mol. Psychiatry 2010, 15, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Rabbito, A.; Dulewicz, M.; Kulczyńska-Przybik, A.; Mroczko, B. Biochemical Markers in Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21, 1989. [Google Scholar] [CrossRef]
- Haass, C.; Lemere, C.A.; Capell, A.; Citron, M.; Seubert, P.; Schenk, D.; Lannfelt, L.; Selkoe, D.J. The Swedish mutation causes early-onset Alzheimer’s disease by beta-secretase cleavage within the secretory pathway. Nat. Med. 1995, 1, 1291–1296. [Google Scholar] [CrossRef] [PubMed]
- Leong, Y.Q.; Ng, K.Y.; Chye, S.M.; Ling, A.P.K.; Koh, R.Y. Mechanisms of action of amyloid-beta and its precursor protein in neuronal cell death. Metab. Brain Dis. 2020, 35, 11–30. [Google Scholar] [CrossRef] [PubMed]
- Nikolaev, A.; McLaughlin, T.; O’Leary, D.D.M.; Tessier-Lavigne, M. APP binds DR6 to trigger axon pruning and neuron death via distinct caspases. Nature 2009, 457, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Sennvik, K.; Bogdanovic, N.; Volkmann, I.; Fastbom, J.; Benedikz, E. Beta-secretase-cleaved amyloid precursor protein in Alzheimer brain: A morphologic study. J. Cell. Mol. Med. 2004, 8, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Fraser, P.E.; Yang, D.S.; Yu, G.; Lévesque, L.; Nishimura, M.; Arawaka, S.; Serpell, L.C.; Rogaeva, E.; St George-Hyslop, P. Presenilin structure, function and role in Alzheimer disease. Biochim. Biophys. Acta 2000, 1502, 1–15. [Google Scholar] [CrossRef]
- Suh, Y.-H.; Checler, F. Amyloid precursor protein, presenilins, and alpha-synuclein: Molecular pathogenesis and pharmacological applications in Alzheimer’s disease. Pharmacol. Rev. 2002, 54, 469–525. [Google Scholar] [CrossRef] [PubMed]
- Octave, J.N.; Essalmani, R.; Tasiaux, B.; Menager, J.; Czech, C.; Mercken, L. The role of presenilin-1 in the gamma-secretase cleavage of the amyloid precursor protein of Alzheimer’s disease. J. Biol. Chem. 2000, 275, 1525–1528. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J. Presenilin, Notch, and the genesis and treatment of Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2001, 98, 11039–11041. [Google Scholar] [CrossRef]
- Morley, J.E.; Farr, S.A.; Banks, W.A.; Johnson, S.N.; Yamada, K.A.; Xu, L. A physiological role for amyloid-beta protein: Enhancement of learning and memory. J. Alzheimer’s Dis. 2010, 19, 441–449. [Google Scholar] [CrossRef]
- Puzzo, D.; Privitera, L.; Leznik, E.; Fà, M.; Staniszewski, A.; Palmeri, A.; Arancio, O. Picomolar amyloid-beta positively modulates synaptic plasticity and memory in hippocampus. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 14537–14545. [Google Scholar] [CrossRef]
- Chen, G.; Xu, T.; Yan, Y.; Zhou, Y.; Jiang, Y.; Melcher, K.; Xu, H.E. Amyloid beta: Structure, biology and structure-based therapeutic development. Acta Pharmacol. Sinica. 2017, 38, 1205–1235. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.M.; Lomakin, A.; Benedek, G.B.; Condron, M.M.; Teplow, D.B. Amyloid β-protein fibrillogenesis: Detection of a protofibrillar intermediate. J. Biol. Chem. 1997, 272, 22364–22372. [Google Scholar] [CrossRef] [PubMed]
- Lesné, S.E.; Sherman, M.A.; Grant, M.; Kuskowski, M.; Schneider, J.A.; Bennett, D.A.; Ashe, K.H. Brain amyloid-β oligomers in ageing and Alzheimer’s disease. Brain 2013, 136, 1383–1398. [Google Scholar] [CrossRef] [PubMed]
- Ezeani, M.; Omabe, M. A New Perspective of Lysosomal Cation Channel-Dependent Homeostasis in Alzheimer’s Disease. Mol. Neurobiol. 2015, 53, 1672–1678. [Google Scholar] [CrossRef] [PubMed]
- Mark, R.J.; Hensley, K.; Butterfield, D.A.; Mattson, M.P. Amyloid beta-peptide impairs ion-motive ATPase activities: Evidence for a role in loss of neuronal Ca2+ homeostasis and cell death. J. Neurosci. Off. J. Soc. Neurosci. 1995, 15, 6239–6249. [Google Scholar] [CrossRef]
- Varadarajan, S.; Yatin, S.; Aksenova, M.; Butterfield, D.A. Review: Alzheimer’s amyloid beta-peptide-associated free radical oxidative stress and neurotoxicity. J. Struct. Biol. 2000, 130, 184–208. [Google Scholar] [CrossRef] [PubMed]
- Heneka, M.T.; Golenbock, D.T.; Latz, E. Innate immunity in Alzheimer’s disease. Nat. Immunol. 2015, 16, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Neniskyte, U.; Neher, J.J.; Brown, G.C. Neuronal death induced by nanomolar amyloid β is mediated by primary phagocytosis of neurons by microglia. J. Biol. Chem. 2011, 286, 39904–39913. [Google Scholar] [CrossRef]
- Weggen, S.; Eriksen, J.L.; Das, P.; Sagi, S.A.; Wang, R.; Pietrzik, C.U.; Findlay, K.A.; Smith, T.E.; Murphy, M.P.; Bulter, T.; et al. A subset of NSAIDs lower amyloidogenic Abeta42 independently of cyclooxygenase activity. Nature 2001, 414, 212–216. [Google Scholar] [CrossRef]
- Devi, L.; Prabhu, B.M.; Galati, D.F.; Avadhani, N.G.; Anandatheerthavarada, H.K. Accumulation of Amyloid Precursor Protein in the Mitochondrial Import Channels of Human Alzheimer’s Disease Brain Is Associated with Mitochondrial Dysfunction. J. Neurosci. 2006, 26, 9057–9068. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, C.G.; Averill, L.A.; Krystal, J.H.; Southwick, S.M.; Arnsten, A.F.T. Glutamate and Norepinephrine Interaction: Relevance to Higher Cognitive Operations and Psychopathology. Behav. Brain Sci. 2016, 39, e201. [Google Scholar] [CrossRef]
- Canevari, L.; Abramov, A.Y.; Duchen, M.R. Toxicity of amyloid beta peptide: Tales of calcium, mitochondria, and oxidative stress. Neurochem. Res. 2004, 29, 637–650. [Google Scholar] [CrossRef]
- Chiarini, A.; Dal Pra, I.; Whitfield, J.F.; Armato, U. The killing of neurons by beta-amyloid peptides, prions, and pro-inflammatory cytokines. Ital. J. Anat. Embryol. Arch. Ital. Anat. Ed. Embriol. 2006, 111, 221–246. [Google Scholar]
- Iwata, N.; Higuchi, M.; Saido, T.C. Metabolism of amyloid-beta peptide and Alzheimer’s disease. Pharmacol. Ther. 2005, 108, 129–148. [Google Scholar] [CrossRef]
- Eckman, E.A.; Reed, D.K.; Eckman, C.B. Degradation of the Alzheimer’s amyloid beta peptide by endothelin-converting enzyme. J. Biol. Chem. 2001, 276, 24540–24548. [Google Scholar] [CrossRef] [PubMed]
- Bertram, L.; Blacker, D.; Mullin, K.; Keeney, D.; Jones, J.; Basu, S.; Yhu, S.; McInnis, M.G.; Go, R.C.; Vekrellis, K.; et al. Evidence for genetic linkage of Alzheimer’s disease to chromosome 10q. Science 2000, 290, 2302–2303. [Google Scholar] [CrossRef]
- Tucker, H.M.; Kihiko, M.; Caldwell, J.N.; Wright, S.; Kawarabayashi, T.; Price, D.; Walker, D.; Scheff, S.; McGillis, J.P.; Rydel, R.E.; et al. The plasmin system is induced by and degrades amyloid-beta aggregates. J. Neurosci. Off. J. Soc. Neurosci. 2000, 20, 3937–3946. [Google Scholar] [CrossRef]
- Backstrom, J.R.; Lim, G.P.; Cullen, M.J.; Tökés, Z.A. Matrix metalloproteinase-9 (MMP-9) is synthesized in neurons of the human hippocampus and is capable of degrading the amyloid-beta peptide (1-40). J. Neurosci. Off. J. Soc. Neurosci. 1996, 16, 7910–7919. [Google Scholar] [CrossRef]
- Hu, J.; Igarashi, A.; Kamata, M.; Nakagawa, H. Angiotensin-converting enzyme degrades Alzheimer amyloid beta-peptide (A beta); retards A beta aggregation, deposition, fibril formation; and inhibits cytotoxicity. J. Biol. Chem. 2001, 276, 47863–47868. [Google Scholar] [CrossRef] [PubMed]
- McDermott, J.R.; Gibson, A.M. Degradation of Alzheimer’s beta-amyloid protein by human cathepsin D. Neuroreport 1996, 7, 2163–2166. [Google Scholar] [CrossRef] [PubMed]
- Deane, R.; Wu, Z.; Sagare, A.; Davis, J.; Du Yan, S.; Hamm, K.; Xu, F.; Parisi, M.; LaRue, B.; Hu, H.W.; et al. LRP/amyloid beta-peptide interaction mediates differential brain efflux of Abeta isoforms. Neuron 2004, 43, 333–344. [Google Scholar] [CrossRef]
- Deane, R.; Du Yan, S.; Submamaryan, R.K.; LaRue, B.; Jovanovic, S.; Hogg, E.; Welch, D.; Manness, L.; Lin, C.; Yu, J.; et al. RAGE mediates amyloid-beta peptide transport across the blood-brain barrier and accumulation in brain. Nat. Med. 2003, 9, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Shibata, M.; Yamada, S.; Kumar, S.R.; Calero, M.; Bading, J.; Frangione, B.; Holtzman, D.M.; Miller, C.A.; Strickland, D.K.; Ghiso, J.; et al. Clearance of Alzheimer’s amyloid-ss(1-40) peptide from brain by LDL receptor-related protein-1 at the blood-brain barrier. J. Clin. Investig. 2000, 106, 1489–1499. [Google Scholar] [CrossRef]
- Cirrito, J.R.; Deane, R.; Fagan, A.M.; Spinner, M.L.; Parsadanian, M.; Finn, M.B.; Jiang, H.; Prior, J.L.; Sagare, A.; Bales, K.R.; et al. P-glycoprotein deficiency at the blood-brain barrier increases amyloid-beta deposition in an Alzheimer disease mouse model. J. Clin. Investig. 2005, 115, 3285–3290. [Google Scholar] [CrossRef] [PubMed]
- Zlokovic, B.V.; Deane, R.; Sallstrom, J.; Chow, N.; Miano, J.M. Neurovascular pathways and Alzheimer amyloid beta-peptide. Brain Pathol. Zurich Switz. 2005, 15, 78–83. [Google Scholar] [CrossRef]
- Mattson, M.P. Pathways Towards and Away from Alzheimer’s Disease. Nature 2004, 430, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Miyakawa, T.; Kimura, T.; Hirata, S.; Fujise, N.; Ono, T.; Ishizuka, K.; Nakabayashi, J. Role of blood vessels in producing pathological changes in the brain with Alzheimer’s disease. Ann. N. Y. Acad. Sci. 2000, 903, 46–54. [Google Scholar] [CrossRef]
- Cuchillo-Ibanez, I.; Seereeram, A.; Byers, H.L.; Leung, K.-Y.; Ward, M.A.; Anderton, B.H.; Hanger, D.P. Phosphorylation of tau regulates its axonal transport by controlling its binding to kinesin. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2008, 22, 3186–3195. [Google Scholar] [CrossRef]
- Dixit, R.; Ross, J.L.; Goldman, Y.E.; Holzbaur, E.L.F. Differential regulation of dynein and kinesin motor proteins by tau. Science 2008, 319, 1086–1089. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-C.; Jiang, Z.-F. Accumulated amyloid-beta peptide and hyperphosphorylated tau protein: Relationship and links in Alzheimer’s disease. J. Alzheimer’s Dis. 2009, 16, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Bloom, G.S. Amyloid-β and tau: The trigger and bullet in Alzheimer disease pathogenesis. JAMA Neurol. 2014, 71, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Lewczuk, P.; Lelental, N.; Lachmann, I.; Holzer, M.; Flach, K.; Brandner, S.; Engelborghs, S.; Teunissen, C.E.; Zetterberg, H.; Molinuevo, J.L.; et al. Non-phosphorylated tau as a potential biomarker of Alzheimer’s Disease: Analytical and diagnostic characterization. J. Alzheimer’s Dis. 2017, 55, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Hugon, J.; Mouton-Liger, F.; Cognat, E.; Dumurgier, J.; Paquet, C. Blood-Based Kinase Assessments in Alzheimer’s Disease. Front. Aging Neurosci. 2018, 10, 338. [Google Scholar] [CrossRef]
- Yuskaitis, C.J.; Jope, R.S. Glycogen synthase kinase-3 regulates microglial migration, inflammation, and inflammation-induced neurotoxicity. Cell. Signal. 2009, 21, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Hall, G.F.; Shea, T.B. Potentiation of tau aggregation by cdk5 and GSK3β. J. Alzheimer’s Dis. 2011, 26, 355–364. [Google Scholar] [CrossRef]
- Srebro, Z.; Wiliński, B.; Sura, P. Oxidative stress in Alzheimer’s disease. Folia Med. Cracov. 2000, 41, 165–170. [Google Scholar]
- Kaltschmidt, B.; Uherek, M.; Volk, B.; Baeuerle, P.A.; Kaltschmidt, C. Transcription factor NF-kappaB is activated in primary neurons by amyloid beta peptides and in neurons surrounding early plaques from patients with Alzheimer disease. Proc. Natl. Acad. Sci. USA 1997, 94, 2642–2647. [Google Scholar] [CrossRef]
- Salim, S. Oxidative Stress and the Central Nervous System. J. Pharmacol. Exp. Ther. 2017, 360, 201–205. [Google Scholar] [CrossRef]
- Akiyama, H.; Barger, S.; Barnum, S.; Bradt, B.; Bauer, J.; Cole, G.M.; Cooper, N.R.; Eikelenboom, P.; Emmerling, M.; Fiebich, B.L.; et al. Inflammation and Alzheimer’s disease. Neurobiol. Aging 2000, 21, 383–421. [Google Scholar] [CrossRef]
- Sayre, L.M.; Smith, M.A.; Perry, G. Chemistry and biochemistry of oxidative stress in neurodegenerative disease. Curr. Med. Chem. 2001, 8, 721–738. [Google Scholar] [CrossRef]
- Morales, I.; Guzmán-Martínez, L.; Cerda-Troncoso, C.; Farías, G.A.; Maccioni, R.B. Neuroinflammation in the pathogenesis of Alzheimer’s disease. A rational framework for the search of novel therapeutic approaches. Front. Cell. Neurosci. 2014, 8, 112. [Google Scholar] [CrossRef] [PubMed]
- Calsolaro, V.; Edison, P. Neuroinflammation in Alzheimer’s disease: Current evidence and future directions. Alzheimer’s Dement. J. Alzheimer’s Assoc. 2016, 12, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Domingues, C.; Cruz e Silva, O.A.B.; Henriques, A.G. Impact of Cytokines and Chemokines on Alzheimer’s Disease Neuro-pathological Hallmarks. Curr. Alzheimer Res. 2017, 14, 870–882. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Liu, C.-C.; Qiao, W.; Bu, G. Apolipoprotein E, Receptors, and Modulation of Alzheimer’s Disease. Biol. Psychiatry 2018, 83, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A.; Mattson, M.P. Apolipoprotein E and oxidative stress in brain with relevance to Alzheimer’s disease. Neurobiol. Dis. 2020, 138, 104795. [Google Scholar] [CrossRef]
- Woolf, N.J.; Butcher, L.L. Cholinergic systems mediate action from movement to higher consciousness. Behav. Brain Res. 2011, 221, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Picciotto, M.R.; Higley, M.J.; Mineur, Y.S. Acetylcholine as a neuromodulator: Cholinergic signaling shapes nervous system function and behavior. Neuron 2012, 76, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Bekdash, R.A. The Cholinergic System, the Adrenergic System and the Neuropathology of Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 1273. [Google Scholar] [CrossRef]
- Briggs, R.; Kennelly, S.P.; O’Neill, D. Drug treatments in Alzheimer’s disease. Clin. Med. 2016, 16, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Vieira, T.H.; Guimaraes, I.M.; Silva, F.R.; Ribeiro, F.M. Alzheimer’s disease: Targeting the Cholinergic System. Curr. Neuropharmacol. 2016, 14, 101–115. [Google Scholar] [CrossRef]
- Egea, J.; Buendia, I.; Parada, E.; Navarro, E.; León, R.; Lopez, M.G. Anti-inflammatory role of microglial alpha7 nAChRs and its role in neuroprotection. Biochem. Pharmacol. 2015, 97, 463–472. [Google Scholar] [CrossRef]
- Wang, H.Y.; Lee, D.H.; D’Andrea, M.R.; Peterson, P.A.; Shank, R.P.; Reitz, A.B. beta-Amyloid(1–42) binds to alpha7 nicotinic acetylcholine receptor with high affinity. Implications for Alzheimer’s disease pathology. J. Biol. Chem. 2000, 275, 5626–5632. [Google Scholar] [CrossRef]
- Lombardo, S.; Maskos, U. Role of the nicotinic acetylcholine receptor in Alzheimer’s disease pathology and treatment. Neuropharmacology 2015, 96, 255–262. [Google Scholar] [CrossRef]
- Hascup, K.N.; Hascup, E.R. Soluble Amyloid-β42 Stimulates Glutamate Release through Activation of the α7 Nicotinic Acetylcholine Receptor. J. Alzheimer’s Dis. 2016, 53, 337–347. [Google Scholar] [CrossRef]
- Jiang, S.; Li, Y.; Zhang, C.; Zhao, Y.; Bu, G.; Xu, H.; Zhang, Y.-W. M1 muscarinic acetylcholine receptor in Alzheimer’s disease. Neurosci. Bull. 2014, 30, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Caccamo, A.; Oddo, S.; Billings, L.M.; Green, K.N.; Martinez-Coria, H.; Fisher, A.; LaFerla, F.M. M1 receptors play a central role in modulating AD-like pathology in transgenic mice. Neuron 2006, 49, 671–682. [Google Scholar] [CrossRef]
- Bukke, V.N.; Archana, M.; Villani, R.; Romano, A.D.; Wawrzyniak, A.; Balawender, K.; Orkisz, S.; Beggiato, S.; Serviddio, G.; Cassano, T. The Dual Role of Glutamatergic Neurotransmission in Alzheimer’s Disease: From Pathophysiology to Pharmacotherapy. Int. J. Mol. Sci. 2020, 21, 7452. [Google Scholar] [CrossRef] [PubMed]
- Rupsingh, R.; Borrie, M.; Smith, M.; Wells, J.L.; Bartha, R. Reduced hippocampal glutamate in Alzheimer disease. Neurobiol. Aging 2011, 32, 802–810. [Google Scholar] [CrossRef] [PubMed]
- Jakob-Roetne, R.; Jacobsen, H. Alzheimer’s disease: From pathology to therapeutic approaches. Angew. Chem. Int. Ed. Engl. 2009, 48, 3030–3059. [Google Scholar] [CrossRef] [PubMed]
- Kothari, S.; Bala, N.; Patel, A.B.; Donovan, A.; Narayanaswami, V. The LDL receptor binding domain of apolipoprotein E directs the relative orientation of its C-terminal segment in reconstituted nascent HDL. Biochim. Biophys. Acta BBA-Biomembr. 2021, 1863, 183618. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Bezprozvanny, I. Differences in Recycling of Apolipoprotein E3 and E4—LDL Receptor Complexes—A Mechanistic Hypothesis. Int. J. Mol. Sci. 2021, 22, 5030. [Google Scholar] [CrossRef] [PubMed]
- Sotolongo, K.; Ghiso, J.; Rostagno, A. Nrf2 activation through the PI3K/GSK-3 axis protects neuronal cells from Aβ-mediated oxidative and metabolic damage. Alzheimer’s Res. Ther. 2020, 12, 13. [Google Scholar] [CrossRef]
- Hamilton, K.; Harvey, J. The Neuronal Actions of Leptin and the Implications for Treating Alzheimer’s Disease. Pharmaceuticals 2021, 14, 52. [Google Scholar] [CrossRef]
- Protective Effects of Carvacrol Against Diabetes-Induced Reproductive Damage in Male Rats: Modulation of Nrf2/HO-1 Signalling Pathway and Inhibition of Nf-kB-Mediated Testicular Apoptosis and Inflammation. Arkali. Andrologia. Wiley Online Library. 2021. Available online: https://onlinelibrary.wiley.com/doi/10.1111/and.13899 (accessed on 1 July 2021).
- Ramos, A.G.B.; de Menezes, I.R.A.; da Silva, M.S.A.; Torres Pessoa, R.; de Lacerda Neto, L.J.; Rocha Santos Passos, F.; Melo Coutinho, H.D.; Iriti, M.; Quintans-Júnior, L.J. Antiedematogenic and Anti-Inflammatory Activity of the Monoterpene Isopulegol and Its β-Cyclodextrin (β-CD) Inclusion Complex in Animal Inflammation Models. Foods 2020, 9, 630. [Google Scholar] [CrossRef] [PubMed]
- Dinda, B.; Debnath, S.; Harigaya, Y. Naturally occurring secoiridoids and bioactivity of naturally occurring iridoids and secoiridoids. A review, part 2. Chem. Pharm. Bull. 2007, 55, 689–728. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. Iridoids and Other Monoterpenes in the Alzheimer’s Brain: Recent Development and Future Prospects. Molecules 2018, 23, 117. [Google Scholar] [CrossRef] [PubMed]
- Volcho, K.P.; Laev, S.S.; Ashraf, G.M.; Aliev, G.; Salakhutdinov, N.F. Application of monoterpenoids and their derivatives for treatment of neurodegenerative disorders. Curr. Med. Chem. 2018, 25, 5327–5346. [Google Scholar] [CrossRef]
- Stavrakov, G.; Philipova, I.; Zheleva-Dimitrova, D.; Valkova, I.; Salamanova, E.; Konstantinov, S.; Doytchinova, I. Docking-based design and synthesis of galantamine-camphane hybrids as inhibitors of acetylcholinesterase. Chem. Biol. Drug Des. 2017, 90, 709–718. [Google Scholar] [CrossRef]
- Silva Brum, L.F.; Emanuelli, T.; Souza, D.O.; Elisabetsky, E. Effects of linalool on glutamate release and uptake in mouse cortical synaptosomes. Neurochem. Res. 2001, 26, 191–194. [Google Scholar] [CrossRef]
- Sabogal-Guáqueta, A.M.; Osorio, E.; Cardona-Gómez, G.P. Linalool reverses neuropathological and behavioral impairments in old triple transgenic Alzheimer’s mice. Neuropharmacology 2016, 102, 111–120. [Google Scholar] [CrossRef]
- Zhong, Z.; Wang, B.; Dai, M.; Sun, Y.; Sun, Q.; Yang, G.; Bian, L. Carvacrol alleviates cerebral edema by modulating AQP4 expression after intracerebral hemorrhage in mice. Neurosci. Lett. 2013, 555, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wang, K.; Lu, C.; Dong, L.; Gao, L.; Yan, M.; Aibai, S.; Yang, Y.; Liu, X. Protective effects of linalool against amyloid beta-induced cognitive deficits and damages in mice. Life Sci. 2017, 174, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Vaibhav, K.; Javed, H.; Tabassum, R.; Ahmed, M.E.; Khan, M.M.; Khan, M.B.; Shrivastava, P.; Islam, F.; Siddiqui, M.S.; et al. 1,8-cineole (eucalyptol) mitigates inflammation in amyloid Beta toxicated PC12 cells: Relevance to Alzheimer’s disease. Neurochem. Res. 2014, 39, 344–352. [Google Scholar] [CrossRef]
- Dinda, B.; Dinda, M.; Kulsi, G.; Chakraborty, A.; Dinda, S. Therapeutic potentials of plant iridoids in Alzheimer’s and Parkinson’s diseases: A review. Eur. J. Med. Chem. 2019, 169, 185–199. [Google Scholar] [CrossRef]
- Liu, H.-C.; Hong, C.-J.; Liu, C.-Y.; Lin, K.-N.; Tsai, S.-J.; Liu, T.-Y.; Chi, C.-W.; Wang, P.-N. Association analysis of the 5-HT6 receptor polymorphism C267T with depression in patients with Alzheimer’s disease. Psychiatry Clin. Neurosci. 2001, 55, 427–429. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; He, Q.-J.; Zou, W.; Wang, H.-X.; Bao, Y.-M.; Liu, Y.-X.; An, L.-J. Catalpol increases hippocampal neuroplasticity and up-regulates PKC and BDNF in the aged rats. Brain Res. 2006, 1123, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-H.; Li, W.-T.; Yu, S.-T.; Xie, H.; Han, H.-R. Catalpol regulates function of hypothalamic-pituitary-adrenocortical-axis in an Alzheimer’s disease rat model. Pharmazie 2014, 69, 688–693. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Huang, X.; Zhao, P.; Zhao, L.; Wang, Z.-Y. Catalpol Inhibits Amyloid-β Generation Through Promoting α-Cleavage of APP in Swedish Mutant APP Overexpressed N2a Cells. Front. Aging Neurosci. 2018, 10. [Google Scholar] [CrossRef]
- Li, H.; Cao, L.; Ren, Y.; Jiang, Y.; Xie, W.; Li, D. GLP-1 receptor regulates cell growth through regulating IDE expression level in Aβ1-42-treated PC12 cells. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef]
- Liu, J.; Liu, Z.; Zhang, Y.; Yin, F. Leptin signaling plays a critical role in the geniposide-induced decrease of tau phosphorylation. Acta Biochim. Biophys. Sin. 2015, 47, 1018–1022. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-H.; Yin, F.; Guo, L.-X.; Deng, X.-H.; Hu, Y.-H. Neuroprotection of geniposide against hydrogen peroxide induced PC12 cells injury: Involvement of PI3 kinase signal pathway. Acta Pharmacol. Sin. 2009, 30, 159–165. [Google Scholar] [CrossRef]
- Flaherty, D.B.; Soria, J.P.; Tomasiewicz, H.G.; Wood, J.G. Phosphorylation of human tau protein by microtubule-associated kinases: GSK3beta and cdk5 are key participants. J. Neurosci. Res. 2000, 62, 463–472. [Google Scholar] [CrossRef]
- Lv, C.; Liu, X.; Liu, H.; Chen, T.; Zhang, W. Geniposide attenuates mitochondrial dysfunction and memory deficits in APP/PS1 transgenic mice. Curr. Alzheimer Res. 2014, 11, 580–587. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, C.; Lv, C.; Liu, X.; Du, S.; Li, Z.; Wang, Y.; Zhang, W. Geniposide Alleviates Amyloid-Induced Synaptic Injury by Protecting Axonal Mitochondrial Trafficking. Front. Cell. Neurosci. 2017, 10. [Google Scholar] [CrossRef]
- Lv, C.; Wang, L.; Liu, X.; Yan, S.; Yan, S.S.; Wang, Y.; Zhang, W. Multi-faced neuroprotective effects of geniposide depending on the RAGE-mediated signaling in an Alzheimer mouse model. Neuropharmacology 2015, 89, 175–184. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, Y.; Deng, X.; Yin, F. Geniposide decreases the level of Aβ1-42 in the hippocampus of streptozotocin-induced diabetic rats. Acta Biochim. Biophys. Sin. 2013, 45, 787–791. [Google Scholar] [CrossRef]
- Morrison, C.D. Leptin signaling in brain: A link between nutrition and cognition? Biochim. Biophys. Acta 2009, 1792, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Nam, K.N.; Choi, Y.-S.; Jung, H.-J.; Park, G.H.; Park, J.-M.; Moon, S.-K.; Cho, K.-H.; Kang, C.; Kang, I.; Oh, M.S.; et al. Genipin inhibits the inflammatory response of rat brain microglial cells. Int. Immunopharmacol. 2010, 10, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, M.; Sakura, N.; Chiba, K.; Mohri, T. Prevention of the neurotoxicity of the amyloid beta protein by genipin. Biol. Pharm. Bull. 2001, 24, 1454–1455. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Liu, Y.; Jiang, Y.; Ding, J.; Li, L. Geniposide ameliorates learning memory deficits, reduces tau phosphorylation and decreases apoptosis via GSK3β pathway in streptozotocin-induced alzheimer rat model. Brain Pathol. Zurich Switz. 2014, 24, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhang, H.; Xu, H.; Xue, R.; Zheng, Y.; Wu, T.; Lian, Y. Harpagoside Rescues the Memory Impairments in Chronic Cerebral Hypoperfusion Rats by Inhibiting PTEN Activity. J. Alzheimer’s Dis. 2018, 63, 445–455. [Google Scholar] [CrossRef]
- Cordero, J.G.; García-Escudero, R.; Avila, J.; Gargini, R.; García-Escudero, V. Benefit of Oleuropein Aglycone for Alzheimer’s Disease by Promoting Autophagy. Oxid. Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Batarseh, Y.S.; Mohamed, L.A.; Al Rihani, S.B.; Mousa, Y.M.; Siddique, A.B.; El Sayed, K.A.; Kaddoumi, A. Oleocanthal ameliorates amyloid-β oligomers’ toxicity on astrocytes and neuronal cells: In vitro studies. Neuroscience 2017, 352, 204–215. [Google Scholar] [CrossRef]
- Wojtunik-Kulesza, K.A.; Targowska-Duda, K.; Klimek, K.; Ginalska, G.; Jóźwiak, K.; Waksmundzka-Hajnos, M.; Cieśla, Ł. Volatile terpenoids as potential drug leads in Alzheimer’s disease. Open Chem. 2017, 15, 332–343. [Google Scholar] [CrossRef]
- Miyazawa, M.; Yamafuji, C. Inhibition of acetylcholinesterase activity by bicyclic monoterpenoids. J. Agric. Food Chem. 2005, 53, 1765–1768. [Google Scholar] [CrossRef]
- Marumoto, S.; Okuno, Y.; Miyazawa, M. Inhibition of β-Secretase Activity by Monoterpenes, Sesquiterpenes, and C13 Norisoprenoids. J. Oleo Sci. 2017, 66, 851–855. [Google Scholar] [CrossRef]
- Seifi Nahavandi, B.; Yaghmaei, P.; Ahmadian, S.; Ebrahim-Habibi, A.; Ghobeh, M. Effects of Terpinolene and Physical Activity on Memory and Learning in a Model of Alzheimer’s Disease among Rats. Qom Univ. Med. Sci. J. 2020, 14, 25–33. [Google Scholar]
- Sabogal-Guáqueta, A.M.; Hobbie, F.; Keerthi, A.; Oun, A.; Kortholt, A.; Boddeke, E.; Dolga, A. Linalool attenuates oxidative stress and mitochondrial dysfunction mediated by glutamate and NMDA toxicity. Biomed. Pharmacother. 2019, 118, 109295. [Google Scholar] [CrossRef]
- Celik, S.; Ozkaya, A. Effects of intraperitoneally administered lipoic acid, vitamin E, and linalool on the level of total lipid and fatty acids in guinea pig brain with oxidative stress induced by H2O2. J. Biochem. Mol. Biol. 2002, 35, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Coelho, V.R.; Gianesini, J.; Von Borowski, R.; Mazzardo-Martins, L.; Martins, D.F.; Picada, J.N.; Santos, A.R.S.; Brum, L.F.S.; Pereira, P. (−)-Linalool, a naturally occurring monoterpene compound, impairs memory acquisition in the object recognition task, inhibitory avoidance test and habituation to a novel environment in rats. Phytomed. Int. J. Phytother. Phytopharm. 2011, 18, 896–901. [Google Scholar] [CrossRef] [PubMed]
- Vianna, M.R.M.; Izquierdo, L.A.; Barros, D.M.; de Souza, M.M.; Rodrigues, C.; Sant’Anna, M.K.; Medina, J.H.; Izquierdo, I. Pharmacological differences between memory consolidation of habituation to an open field and inhibitory avoidance learning. Braz. J. Med. Biol. Res. 2001, 34, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Do Vale, T.G.; Furtado, E.C.; Santos, J.G.; Viana, G.S.B. Central effects of citral, myrcene and limonene, constituents of essential oil chemotypes from Lippia alba (Mill.) n.e. Brown. Phytomed. Int. J. Phytother. Phytopharm. 2002, 9, 709–714. [Google Scholar] [CrossRef]
- Yang, Z.; Xi, J.; Li, J.; Qu, W. Biphasic effect of citral, a flavoring and scenting agent, on spatial learning and memory in rats. Pharmacol. Biochem. Behav. 2009, 93, 391–396. [Google Scholar] [CrossRef]
- Chaudhary, S.C.; Siddiqui, M.S.; Athar, M.; Alam, M.S. D-Limonene modulates inflammation, oxidative stress and Ras-ERK pathway to inhibit murine skin tumorigenesis. Hum. Exp. Toxicol. 2012, 31, 798–811. [Google Scholar] [CrossRef]
- Murali, R.; Saravanan, R. Antidiabetic effect of d-limonene, a monoterpene in streptozotocin-induced diabetic rats. Biomed. Prev. Nutr. 2012, 2, 269–275. [Google Scholar] [CrossRef]
- Bigdeli, Y.; Asle-Rousta, M.; Rahnema, M. Effects of Limonene on Chronic Restraint Stress-Induced Memory Impairment and Anxiety in Male Rats. Neurophysiology 2019, 51, 107–113. [Google Scholar] [CrossRef]
- Asadbegi, M.; Yaghmaei, P.; Salehi, I.; Komaki, A.; Ebrahim-Habibi, A. Investigation of thymol effect on learning and memory impairment induced by intrahippocampal injection of amyloid beta peptide in high fat diet- fed rats. Metab. Brain Dis. 2017, 32, 827–839. [Google Scholar] [CrossRef]
- Fang, F.; Li, H.; Qin, T.; Li, M.; Ma, S. Thymol improves high-fat diet-induced cognitive deficits in mice via ameliorating brain insulin resistance and upregulating NRF2/HO-1 pathway. Metab. Brain Dis. 2017, 32, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; An, Z. Hesperidin attenuates learning and memory deficits in APP/PS1 mice through activation of Akt/Nrf2 signaling and inhibition of RAGE/NF-κB signaling. Arch. Pharm. Res. 2018, 41, 655–663. [Google Scholar] [CrossRef]
- Zhong, S.-Z.; Ge, Q.-H.; Li, Q.; Qu, R.; Ma, S.-P. Peoniflorin attentuates Aβ(1-42)-mediated neurotoxicity by regulating calcium homeostasis and ameliorating oxidative stress in hippocampus of rats. J. Neurol. Sci. 2009, 280, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Lan, Z.; Chen, L.; Fu, Q.; Ji, W.; Wang, S.; Liang, Z.; Qu, R.; Kong, L.; Ma, S. Paeoniflorin attenuates amyloid-beta peptide-induced neurotoxicity by ameliorating oxidative stress and regulating the NGF-mediated signaling in rats. Brain Res. 2013, 1498, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Niewiadomska, G.; Mietelska-Porowska, A.; Mazurkiewicz, M. The cholinergic system, nerve growth factor and the cytoskeleton. Behav. Brain Res. 2011, 221, 515–526. [Google Scholar] [CrossRef]
- Niewiadomska, G.; Komorowski, S.; Baksalerska-Pazera, M. Amelioration of cholinergic neurons dysfunction in aged rats depends on the continuous supply of NGF. Neurobiol. Aging 2002, 23, 601–613. [Google Scholar] [CrossRef]
- Azizi, Z.; Ebrahimi, S.; Saadatfar, E.; Kamalinejad, M.; Majlessi, N. Cognitive-enhancing activity of thymol and carvacrol in two rat models of dementia. Behav. Pharmacol. 2012, 23, 241–249. [Google Scholar] [CrossRef]
- Lee, B.; Yeom, M.; Shim, I.; Lee, H.; Hahm, D.-H. Inhibitory effect of carvacrol on lipopolysaccharide-induced memory impairment in rats. Korean J. Physiol. Pharmacol. 2020, 24, 27–37. [Google Scholar] [CrossRef]
- Zare Mehrjerdi, F.; Niknazar, S.; Yadegari, M.; Akbari, F.A.; Pirmoradi, Z.; Khaksari, M. Carvacrol reduces hippocampal cell death and improves learning and memory deficits following lead-induced neurotoxicity via antioxidant activity. Naunyn. Schmiedebergs Arch. Pharmacol. 2020, 393, 1229–1237. [Google Scholar] [CrossRef]
- Wang, X.; Li, G.; Shen, W. Protective effects of D-Limonene against transient cerebral ischemia in stroke-prone spontaneously hypertensive rats. Exp. Ther. Med. 2018, 15, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.-Y.; Lee, C.; Park, G.H.; Jang, J.-H. Amelioration of Scopolamine-Induced Learning and Memory Impairment by α-Pinene in C57BL/6 Mice. Evid. Based Complement. Altern. Med. 2017, 2017, 4926815. [Google Scholar] [CrossRef]
- Baldissera, M.D.; Souza, C.F.; Grando, T.H.; Sagrillo, M.R.; De Brum, G.F.; Nascimento, K.; Peres, D.S.; Maciel, M.F.; Silveira, S.O.; Da Luz, S.C.A.; et al. Memory deficit, toxic effects and activity of Na+, K+-ATPase and NTPDase in brain of Wistar rats submitted to orally treatment with alpha-terpinene. Environ. Toxicol. Pharmacol. 2016, 46, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Haskins, J.T. Generalized anxiety disorder. Epidemiology, impact of comorbidity, and natural history. Postgrad. Med. 1999, 106, 3–9. [Google Scholar] [CrossRef]
- Bilkei-Gorzo, A.; Ströhle, A. (Eds.) Anxiety and Anxiolytic Drugs; Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2005; ISBN 978-3-540-22568-3. [Google Scholar]
- Altunoz, U.; Kokurcan, A.; Kirici, S.; Bastug, G.; Ozel-Kizil, E.T. Clinical characteristics of generalized anxiety disorder: Older vs. young adults. Nord. J. Psychiatry 2018, 72, 97–102. [Google Scholar] [CrossRef]
- Pietrzak, R.H.; Lim, Y.Y.; Neumeister, A.; Ames, D.; Ellis, K.A.; Harrington, K.; Lautenschlager, N.T.; Restrepo, C.; Martins, R.N.; Masters, C.L.; et al. Amyloid-β, Anxiety, and Cognitive Decline in Preclinical Alzheimer Disease: A Multicenter, Prospective Cohort Study. JAMA Psychiatry 2015, 72, 284–291. [Google Scholar] [CrossRef]
- Porter, V.R.; Buxton, W.G.; Fairbanks, L.A.; Strickland, T.; O’Connor, S.M.; Rosenberg-Thompson, S.; Cummings, J.L. Frequency and Characteristics of Anxiety Among Patients with Alzheimer’s Disease and Related Dementias. J. Neuropsychiatry Clin. Neurosci. 2003, 15, 180–186. [Google Scholar] [CrossRef]
- Chemerinski, E.; Petracca, G.; Manes, F.; Leiguarda, R.; Starkstein, S.E. Prevalence and correlates of anxiety in Alzheimer’s disease. Depress. Anxiety 1998, 7, 166–170. [Google Scholar] [CrossRef]
- Li, S.; Wang, C.; Wang, W.; Tan, J. Trait anxiety, a personality risk factor associated with Alzheimer’s Disease. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 105, 110124. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-J.; Heinbockel, T. Essential Oils and Their Constituents Targeting the GABAergic System and Sodium Channels as Treatment of Neurological Diseases. Molecules 2018, 23, 1061. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, J.T.; Shropshire, B.C.; Nagy, T.R.; Chambers, K.D.; Li, Y.; Korach, K.S. Essential Oils and Health. Yale J. Biol. Med. 2020, 93, 291–305. [Google Scholar] [PubMed]
- Chioca, L.R.; Antunes, V.D.C.; Ferro, M.M.; Losso, E.M.; Andreatini, R. Anosmia does not impair the anxiolytic-like effect of lavender essential oil inhalation in mice. Life Sci. 2013, 92, 971–975. [Google Scholar] [CrossRef] [PubMed]
- Schuwald, A.M.; Nöldner, M.; Wilmes, T.; Klugbauer, N.; Leuner, K.; Müller, W.E. Lavender oil-potent anxiolytic properties via modulating voltage dependent calcium channels. PLoS ONE 2013, 8, e59998. [Google Scholar] [CrossRef]
- Takahashi, M.; Satou, T.; Ohashi, M.; Hayashi, S.; Sadamoto, K.; Koike, K. Interspecies comparison of chemical composition and anxiolytic-like effects of lavender oils upon inhalation. Nat. Prod. Commun. 2011, 6, 1769–1774. [Google Scholar] [CrossRef] [PubMed]
- Prusinowska, R.; Śmigielski, K.B. Composition, biological properties and therapeutic effects of lavender (Lavandula angustifolia L). A review. Herba Polonica 2014, 60, 56–66. [Google Scholar] [CrossRef]
- Nasiri Lari, Z.; Hajimonfarednejad, M.; Riasatian, M.; Abolhassanzadeh, Z.; Iraji, A.; Vojoud, M.; Heydari, M.; Shams, M. Efficacy of inhaled Lavandula angustifolia Mill. Essential oil on sleep quality, quality of life and metabolic control in patients with diabetes mellitus type II and insomnia. J. Ethnopharmacol. 2020, 251, 112560. [Google Scholar] [CrossRef] [PubMed]
- Smigielski, K.; Prusinowska, R.; Raj, A.; Sikora, M.; Woliñska, K.; Gruska, R. Effect of Drying on the Composition of Essential Oil from Lavandula angustifolia. J. Essent. Oil Bear. Plants 2011, 14, 532–542. [Google Scholar] [CrossRef]
- Kasper, S. An orally administered lavandula oil preparation (Silexan) for anxiety disorder and related conditions: An evidence based review. Int. J. Psychiatry Clin. Pract. 2013, 17, 15–22. [Google Scholar] [CrossRef]
- Melo, F.H.C.; Venâncio, E.T.; de Sousa, D.P.; de França Fonteles, M.M.; de Vasconcelos, S.M.M.; Viana, G.S.B.; de Sousa, F.C.F. Anxiolytic-like effect of Carvacrol (5-isopropyl-2-methylphenol) in mice: Involvement with GABAergic transmission. Fundam. Clin. Pharmacol. 2010, 24, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Pires, L.F.; Costa, L.M.; Silva, O.A.; de Almeida, A.A.C.; Cerqueira, G.S.; de Sousa, D.P.; de Freitas, R.M. Anxiolytic-like effects of carvacryl acetate, a derivative of carvacrol, in mice. Pharmacol. Biochem. Behav. 2013, 112, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Moreira, M.R.C.; da Silva Stiebbe Salvadori, M.G.; de Almeida, A.A.C.; de Sousa, D.P.; Jordán, J.; Satyal, P.; de Freitas, R.M.; de Almeida, R.N. Anxiolytic-like effects and mechanism of (−)-myrtenol: A monoterpene alcohol. Neurosci. Lett. 2014, 579, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Linck, V.M.; da Silva, A.L.; Figueiró, M.; Caramão, E.B.; Moreno, P.R.H.; Elisabetsky, E. Effects of inhaled Linalool in anxiety, social interaction and aggressive behavior in mice. Phytomedicine 2010, 17, 679–683. [Google Scholar] [CrossRef]
- Gomes, P.B.; Feitosa, M.L.; Silva, M.I.G.; Noronha, E.C.; Moura, B.A.; Venâncio, E.T.; Rios, E.R.V.; de Sousa, D.P.; de Vasconcelos, S.M.M.; de Franaç Fonteles, M.M. Anxiolytic-like effect of the monoterpene 1,4-cineole in mice. Pharmacol. Biochem. Behav. 2010, 96, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Satou, T.; Kasuya, H.; Maeda, K.; Koike, K. Daily inhalation of α-pinene in mice: Effects on behavior and organ accumulation. Phytother. Res. 2014, 28, 1284–1287. [Google Scholar] [CrossRef]
- Umezu, T.; Ito, H.; Nagano, K.; Yamakoshi, M.; Oouchi, H.; Sakaniwa, M.; Morita, M. Anticonflict effects of rose oil and identification of its active constituents. Life Sci. 2002, 72, 91–102. [Google Scholar] [CrossRef]
- Marques, T.; Marques, M.L.B.G.C.B.; Lima, D.; Siqueira, H.; Neto, J.; Branco, M.S.B.G.C.; Souza, A.; Sousa, D.; Freitas, R. Evaluation of the neuropharmacological properties of nerol in mice. World J. Neurosci. 2013, 3, 32–38. [Google Scholar] [CrossRef]
- Satou, T.; Matsuura, M.; Takahashi, M.; Umezu, T.; Hayashi, S.; Sadamoto, K.; Koike, K. Anxiolytic-like effect of essential oil extracted from Abies sachalinensis. Flavour Fragr. J. 2011, 26, 416–420. [Google Scholar] [CrossRef]
- Radulović, N.S.; Dekić, M.S.; Ranđelović, P.J.; Stojanović, N.M.; Zarubica, A.R.; Stojanović-Radić, Z.Z. Toxic essential oils: Anxiolytic, antinociceptive and antimicrobial properties of the yarrow Achillea umbellata Sibth. et Sm. (Asteraceae) volatiles. Food Chem. Toxicol. 2012, 50, 2016–2026. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Matsuura, M.; Satou, T.; Hayashi, S.; Koike, K. Effects of the Essential Oil from Leaves of Alpinia zerumbet on Behavioral Alterations in Mice. Nat. Prod. Commun. 2009, 4, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.A.R.A.; Cury, T.C.; Cassettari, B.O.; Takahira, R.K.; Flório, J.C.; Costa, M. Citrus aurantium L. essential oil exhibits anxiolytic-like activity mediated by 5-HT1A-receptors and reduces cholesterol after repeated oral treatment. BMC Complement. Altern. Med. 2013, 13, 42. [Google Scholar] [CrossRef]
- Carvalho-Freitas, M.I.R.; Costa, M. Anxiolytic and Sedative Effects of Extracts and Essential Oil from Citrus aurantium L. Biol. Pharm. Bull. 2002, 25, 1629–1633. [Google Scholar] [CrossRef]
- Leite, M.P.; Fassin, J., Jr.; Baziloni, E.M.F.; Almeida, R.N.; Mattei, R.; Leite, J.R. Behavioral effects of essential oil of Citrus aurantium L. inhalation in rats. Rev. Bras. Farmacogn. 2008, 18, 661–666. [Google Scholar] [CrossRef]
- Lopes Campêlo, L.M.; Gonçalves e Sá, C.; de Almeida, A.A.C.; da Costa, J.P.; Marques, T.H.C.; Feitosa, C.M.; Saldanha, G.B.; de Freitas, R.M. Sedative, anxiolytic and antidepressant activities of Citrus limon (Burn) essential oil in mice. Pharm. 2011, 66, 623–627. [Google Scholar]
- Ceccarelli, I.; Lariviere, W.R.; Fiorenzani, P.; Sacerdote, P.; Aloisi, A.M. Effects of long-term exposure of lemon essential oil odor on behavioral, hormonal and neuronal parameters in male and female rats. Brain Res. 2004, 1001, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Faturi, C.B.; Leite, J.R.; Alves, P.B.; Canton, A.C.; Teixeira-Silva, F. Anxiolytic-like effect of sweet orange aroma in Wistar rats. Prog. Neuropsychopharmacol. Biol. Psychiatry 2010, 34, 605–609. [Google Scholar] [CrossRef] [PubMed]
- Dougnon, G.; Ito, M. Sedative effects of the essential oil from the leaves of Lantana camara occurring in the Republic of Benin via inhalation in mice. J. Nat. Med. 2020, 74, 159–169. [Google Scholar] [CrossRef]
- Takahashi, M.; Yoshino, A.; Yamanaka, A.; Asanuma, C.; Satou, T.; Hayashi, S.; Masuo, Y.; Sadamoto, K.; Koike, K. Effects of Inhaled Lavender Essential Oil on Stress-Loaded Animals: Changes in Anxiety-Related Behavior and Expression Levels of Selected mRNAs and Proteins. Nat. Prod. Commun. 2012, 7, 1934578X1200701132. [Google Scholar] [CrossRef]
- Chioca, L.R.; Ferro, M.M.; Baretta, I.P.; Oliveira, S.M.; Silva, C.R.; Ferreira, J.; Losso, E.M.; Andreatini, R. Anxiolytic-like effect of lavender essential oil inhalation in mice: Participation of serotonergic but not GABAA/benzodiazepine neurotransmission. J. Ethnopharmacol. 2013, 147, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Yamanaka, A.; Asanuma, C.; Asano, H.; Satou, T.; Koike, K. Anxiolytic-like Effect of Inhalation of Essential Oil from Lavandula officinalis: Investigation of Changes in 5-HT Turnover and Involvement of Olfactory Stimulation. Nat. Prod. Commun. 2014, 9, 1934578X1400900735. [Google Scholar] [CrossRef]
- Shaw, D.; Annett, J.M.; Doherty, B.; Leslie, J.C. Anxiolytic effects of lavender oil inhalation on open-field behaviour in rats. Phytomed. Int. J. Phytother. Phytopharm. 2007, 14, 613–620. [Google Scholar] [CrossRef]
- Bradley, B.F.; Starkey, N.J.; Brown, S.L.; Lea, R.W. Anxiolytic effects of Lavandula angustifolia odour on the Mongolian gerbil elevated plus maze. J. Ethnopharmacol. 2007, 111, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Hawken, P.A.R.; Fiol, C.; Blache, D. Genetic differences in temperament determine whether lavender oil alleviates or exacerbates anxiety in sheep. Physiol. Behav. 2012, 105, 1117–1123. [Google Scholar] [CrossRef]
- Woelk, H.; Schläfke, S. A multi-center, double-blind, randomised study of the Lavender oil preparation Silexan in comparison to Lorazepam for generalized anxiety disorder. Phytomedicine 2010, 17, 94–99. [Google Scholar] [CrossRef]
- Chen, C.-J.; Tseng, Y.-H.; Chu, F.-H.; Wen, T.-Y.; Cheng, W.-W.; Chen, Y.-T.; Tsao, N.-W.; Wang, S.-Y. Neuropharmacological activities of fruit essential oil from Litsea cubeba Persoon. J. Wood Sci. 2012, 58, 538–543. [Google Scholar] [CrossRef]
- Tankam, J.M.; Ito, M. Sedative, anxiolytic and antidepressant-like effects of inhalation of the essential oil of Ocimum gratissimum L. from Cameroon in mice. Ournal Pharmacogn. Phytochem. 2014, 2, 1–9. [Google Scholar]
- Siengsukon, C.F.; Nelson, E.; Williams-Cooke, C.; Ludwig, R.; Beck, E.S.; Vidoni, E.D.; Mahnken, J.D.; Stevens, S.; Drerup, M.; Bruce, J.; et al. Cognitive behavioral therapy for insomnia to enhance cognitive function and reduce the rate of Aβ deposition in older adults with symptoms of insomnia: A single-site randomized pilot clinical trial protocol. Contemp. Clin. Trials 2020, 99, 106190. [Google Scholar] [CrossRef] [PubMed]
- Fultz, N.E.; Bonmassar, G.; Setsompop, K.; Stickgold, R.A.; Rosen, B.R.; Polimeni, J.R.; Lewis, L.D. Coupled electrophysiological, hemodynamic, and cerebrospinal fluid oscillations in human sleep. Science 2019, 366, 628–631. [Google Scholar] [CrossRef]
- Xie, L.; Kang, H.; Xu, Q.; Chen, M.J.; Liao, Y.; Thiyagarajan, M.; O’Donnell, J.; Christensen, D.J.; Nicholson, C.; Iliff, J.J.; et al. Sleep drives metabolite clearance from the adult brain. Science 2013, 342, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Zhong, R.; Liu, H.; Zhang, F.; Li, S.; Le, W. Chronic Sleep Deprivation Exacerbates Learning-Memory Disability and Alzheimer’s Disease-Like Pathologies in AβPP(swe)/PS1(ΔE9) Mice. J. Alzheimer’s Dis. 2016, 50, 669–685. [Google Scholar] [CrossRef]
- Irwin, M.R.; Olmstead, R.; Carroll, J.E. Sleep Disturbance, Sleep Duration, and Inflammation: A Systematic Review and Meta-Analysis of Cohort Studies and Experimental Sleep Deprivation. Biol. Psychiatry 2016, 80, 40–52. [Google Scholar] [CrossRef]
- Xu, L.; Li, X.; Zhang, Y.; Ding, M.; Sun, B.; Su, G.; Zhao, Y. The effects of linalool acupoint application therapy on sleep regulation. RSC Adv. 2021, 11, 5896–5902. [Google Scholar] [CrossRef]
- Yang, H.; Woo, J.; Pae, A.N.; Um, M.Y.; Cho, N.-C.; Park, K.D.; Yoon, M.; Kim, J.; Lee, C.J.; Cho, S. α-Pinene, a Major Constituent of Pine Tree Oils, Enhances Non-Rapid Eye Movement Sleep in Mice through GABAA-benzodiazepine Receptors. Mol. Pharmacol. 2016, 90, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.; Yang, H.; Yoon, M.; Gadhe, C.G.; Pae, A.N.; Cho, S.; Lee, C.J. 3-Carene, a Phytoncide from Pine Tree Has a Sleep-enhancing Effect by Targeting the GABAA-benzodiazepine Receptors. Exp. Neurobiol. 2019, 28, 593–601. [Google Scholar] [CrossRef]
- Sobreira Dantas Nóbrega de Figuêiredo, F.R.; Monteiro, Á.B.; Alencar de Menezes, I.R.; Sales, V.D.S.; Petícia do Nascimento, E.; Kelly de Souza Rodrigues, C.; Bitu Primo, A.J.; da Cruz, L.P.; do Nascimento Amaro, É.; de Araújo Delmondes, G.; et al. Effects of the Hyptis Martiusii Benth. Leaf Essential Oil and 1,8-Cineole (Eucalyptol) on the Central Nervous System of Mice. Food Chem Toxicol 2019, 133, 110802. [Google Scholar] [CrossRef]
- De Sousa, D.P.; de Farias Nóbrega, F.F.; de Almeida, R.N. Influence of the chirality of (R)-(−)- and (S)-(+)-carvone in the central nervous system: A comparative study. Chirality 2007, 19, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, F.; Wink, M. Synergistic interactions of saponins and monoterpenes in HeLa cells, Cos7 cells and in erythrocytes. Phytomedicine 2011, 18, 1191–1196. [Google Scholar] [CrossRef]
- Gertsch, J. Botanical Drugs, Synergy, and Network Pharmacology: Forth and Back to Intelligent Mixtures. Planta Med. 2011, 77, 1086–1098. [Google Scholar] [CrossRef]
- Eid, S.Y.; El-Readi, M.Z.; Eldin, E.E.M.N.; Fatani, S.H.; Wink, M. Influence of combinations of digitonin with selected phenolics, terpenoids, and alkaloids on the expression and activity of P-glycoprotein in leukaemia and colon cancer cells. Phytomedicine 2013, 21, 47–61. [Google Scholar] [CrossRef]
- Jayamani, J.; Shanmugam, G. Gallic acid, one of the components in many plant tissues, is a potential inhibitor for insulin amyloid fibril formation. Eur. J. Med. Chem. 2014, 85, 352–358. [Google Scholar] [CrossRef]
- Yu, M.; Chen, X.; Liu, J.; Ma, Q.; Zhuo, Z.; Chen, H.; Zhou, L.; Yang, S.; Zheng, L.; Ning, C.; et al. Gallic acid disruption of Aβ1-42 aggregation rescues cognitive decline of APP/PS1 double transgenic mouse. Neurobiol. Dis. 2019, 124, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Ehrnhoefer, D.E.; Bieschke, J.; Boeddrich, A.; Herbst, M.; Masino, L.; Lurz, R.; Engemann, S.; Pastore, A.; Wanker, E.E. EGCG redirects amyloidogenic polypeptides into unstructured, off-pathway oligomers. Nat. Struct. Mol. Biol. 2008, 15, 558–566. [Google Scholar] [CrossRef]
- Huang, R.; Vivekanandan, S.; Brender, J.R.; Abe, Y.; Naito, A.; Ramamoorthy, A. NMR Characterization of Monomeric and Oligomeric Conformations of Human Calcitonin and Its Interaction with EGCG. J. Mol. Biol. 2012, 416, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Alternative Pathways of Human Islet Amyloid Polypeptide Aggregation Distinguished by 19F Nuclear Magnetic Resonance-Detected Kinetics of Monomer Consumption|Biochemistry. Available online: https://pubs.acs.org/doi/10.1021/bi3012548 (accessed on 4 July 2021).
- Franko, A.; Rodriguez Camargo, D.C.; Böddrich, A.; Garg, D.; Rodriguez Camargo, A.; Rathkolb, B.; Janik, D.; Aichler, M.; Feuchtinger, A.; Neff, F.; et al. Epigallocatechin gallate (EGCG) reduces the intensity of pancreatic amyloid fibrils in human islet amyloid polypeptide (hIAPP) transgenic mice. Sci. Rep. 2018, 8, 1116. [Google Scholar] [CrossRef]
- Ferreira, N.; Cardoso, I.; Domingues, M.R.; Vitorino, R.; Bastos, M.; Bai, G.; Saraiva, M.J.; Almeida, M.R. Binding of epigallocatechin-3-gallate to transthyretin modulates its amyloidogenicity. FEBS Lett. 2009, 583, 3569–3576. [Google Scholar] [CrossRef]
- Ferreira, N.; Pereira-Henriques, A.; Almeida, M.R. Transthyretin chemical chaperoning by flavonoids: Structure-activity insights towards the design of potent amyloidosis inhibitors. Biochem. Biophys. Rep. 2015, 3, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, N.; Saraiva, M.J.; Almeida, M.R. Natural polyphenols inhibit different steps of the process of transthyretin (TTR) amyloid fibril formation. FEBS Lett. 2011, 585, 2424–2430. [Google Scholar] [CrossRef]
- Garcia-Alloza, M.; Borrelli, L.A.; Rozkalne, A.; Hyman, B.T.; Bacskai, B.J. Curcumin labels amyloid pathology in vivo, disrupts existing plaques, and partially restores distorted neurites in an Alzheimer mouse model. J. Neurochem. 2007, 102, 1095–1104. [Google Scholar] [CrossRef]
- Ono, K.; Yamada, M. Antioxidant compounds have potent anti-fibrillogenic and fibril-destabilizing effects for alpha-synuclein fibrils in vitro. J. Neurochem. 2006, 97, 105–115. [Google Scholar] [CrossRef]
- Ghajarbeygi, P.; Hajhoseini, A.; Hosseini, M.-S.; Sharifan, A. An In Vitro and In Vivo Cholinesterase Inhibitory Activity of Pistacia khinjuk and Allium sativum Essential Oils. J. Pharmacopuncture 2019, 22, 231–238. [Google Scholar] [CrossRef]
- Azimi, M.; Sharifan, A.; Tarzi, B.G. The Use of Pistacia khinjuk Essential Oil to Modulate Shelf-Life and Organoleptic Traits of Mechanically Deboned Chicken Meat. J. Food Process. Preserv. 2017, 41, e12814. [Google Scholar] [CrossRef]
- Hawrył, A.; Hawrył, M.; Waksmundzka-Hajnos, M. Liquid chromatography fingerprint analysis and antioxidant activity of selected lavender species with chemometric calculations. PLoS ONE 2019, 14, e0218974. [Google Scholar] [CrossRef]
- Amer Tawfeeq, A.; Faisal, M.; Abaas, I.; Alwan, A. Isolation, quantification, and identification of rosmarinic acid, gas chromatography-mass spectrometry analysis of essential oil, cytotoxic effect, and antimicrobial investigation of rosmarinus officinalis leaves. Asian J. Pharm. Clin. Res. 2018, 11, 126. [Google Scholar] [CrossRef]
- Boiangiu, R.S.; Brinza, I.; Hancianu, M.; Erdogan Orhan, I.; Eren, G.; Gündüz, E.; Ertas, H.; Hritcu, L.; Cioanca, O. Cognitive Facilitation and Antioxidant Effects of an Essential Oil Mix on Scopolamine-Induced Amnesia in Rats: Molecular Modeling of In Vitro and In Vivo Approaches. Molecules 2020, 25, 1519. [Google Scholar] [CrossRef]
- Armijos, C.; Matailo, A.; Bec, N.; Salinas, M.; Aguilar, G.; Solano, N.; Calva, J.; Ludeña, C.; Larroque, C.; Vidari, G. Chemical composition and selective BuChE inhibitory activity of the essential oils from aromatic plants used to prepare the traditional Ecuadorian beverage horchata lojana. J. Ethnopharmacol. 2020, 263. [Google Scholar] [CrossRef]
- Bailon-Moscoso, N.; Tinitana, F.; Martínez-Espinosa, R.; Jaramillo-Velez, A.; Palacio-Arpi, A.; Aguilar-Hernandez, J.; Romero-Benavides, J.C. Cytotoxic, antioxidative, genotoxic and antigenotoxic effects of Horchata, beverage of South Ecuador. BMC Complement. Altern. Med. 2017, 17, 539. [Google Scholar] [CrossRef]
- Tene, V.; Malagon, O.; Vita-Finzi, P.; Vidari, G.; Armijos, C.; Zaragoza, T. An ethnobotanical survey of medicinal plants used in Loja and Zamora-Chinchipe, Ecuador. J. Ethnopharmacol. 2007, 111, 63–81. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H. Multitarget therapy--the future of treatment for more than just functional dyspepsia. Phytomed. Int. J. Phytother. Phytopharm. 2006, 13, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.O.; Scholey, A.B.; Wesnes, K.A. Differential, dose dependent changes in cognitive performance following acute administration of a Ginkgo biloba/Panax ginseng combination to healthy young volunteers. Nutr. Neurosci. 2001, 4, 399–412. [Google Scholar] [CrossRef]
- Butterweck, V.; Petereit, F.; Winterhoff, H.; Nahrstedt, A. Solubilized hypericin and pseudohypericin from Hypericum perforatum exert antidepressant activity in the forced swimming test. Planta Med. 1998, 64, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.; Ulrich-Merzenich, G. Synergy research: Approaching a new generation of phytopharmaceuticals. Phytomed. Int. J. Phytother. Phytopharm. 2009, 16, 97–110. [Google Scholar] [CrossRef]
- Rezai-Zadeh, K.; Shytle, D.; Sun, N.; Mori, T.; Hou, H.; Jeanniton, D.; Ehrhart, J.; Townsend, K.; Zeng, J.; Morgan, D.; et al. Green tea epigallocatechin-3-gallate (EGCG) modulates amyloid precursor protein cleavage and reduces cerebral amyloidosis in Alzheimer transgenic mice. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 8807–8814. [Google Scholar] [CrossRef]
- Ferreira, N.; Saraiva, M.J.; Almeida, M.R. Epigallocatechin-3-gallate as a potential therapeutic drug for TTR-related amyloidosis: “in vivo” evidence from FAP mice models. PLoS ONE 2012, 7, e29933. [Google Scholar] [CrossRef] [PubMed]
- Bagci, E.; Aydin, E.; Ungureanu, E.; Hritcu, L. Anthriscus nemorosa essential oil inhalation prevents memory impairment, anxiety and depression in scopolamine-treated rats. Biomed. Pharmacother. 2016, 84, 1313–1320. [Google Scholar] [CrossRef] [PubMed]
- Maleki, H.; Norouzi, R.; Shahi, A. Essential oil components, phenolic content and antioxidant activity of Anthriscus cerefolium and Anthriscus sylvestris from Iran. J. Hortic. Postharvest Res. 2020, 3, 355–366. [Google Scholar] [CrossRef]
- Areias, F.; Valentão, P.; Andrade, P.; Moreira, M.; Amaral, J.; Seabra, R. HPLC/DAD analysis of phenolic compounds from lavender and its application to quality control. J. Liq. Chromatogr. Relat. Technol. 2000, 23, 2563–2572. [Google Scholar] [CrossRef]
- López, V.; Nielsen, B.; Solas, M.; Ramírez, M.J.; Jäger, A.K. Exploring Pharmacological Mechanisms of Lavender (Lavandula angustifolia) Essential Oil on Central Nervous System Targets. Front. Pharmacol. 2017, 8. [Google Scholar] [CrossRef]
- Ko, L.-W.; Su, C.-H.; Yang, M.-H.; Liu, S.-Y.; Su, T.-P. A pilot study on essential oil aroma stimulation for enhancing slow-wave EEG in sleeping brain. Sci. Rep. 2021, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Rodríguez, R.; Pérez-Hernández, M.G.; Maturano-Melgoza, J.A.; Hilerio-López, Á.G.; Monroy-Rojas, A.; Arana-Gómez, B.; Vásquez, C. The effect of aromatherapy with lavender (LLavandula angustifoliaL) on serum melatonin levels. Complement. Ther. Med. 2019, 47, 102208. [Google Scholar] [CrossRef] [PubMed]
- Sadiki, F.Z.; Idrissi, M.E.; Cioanca, O.; Trifan, A.; Hancianu, M.; Hritcu, L.; Postu, P.A. Tetraclinis articulata essential oil mitigates cognitive deficits and brain oxidative stress in an Alzheimer’s disease amyloidosis model. Phytomedicine 2019, 56, 57–63. [Google Scholar] [CrossRef]
- Harouak, H.; Ibijbijen, J.; Nassiri, L. Chemical profile of Tetraclinis articulata (Vahl) Masters, and Juglans regia L. and Olea europaea L. var. Sylvestris used against oral diseases: In Vitro analysis between polyphenolic content and aqueous extraction optimization. Heliyon 2021, 7, e07118. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Monoterpene | In Vivo Model | Type of Test | Administration of Compound | Observations | Possible Mechanism of Action | References |
|---|---|---|---|---|---|---|
| thymol | Wistar Rats impaired with intrahippocampal injection of amyloid or intraperitoneal injection of scopolamine | Morris water maze task | intraperitoneally; 0.5, 1, or 2 mg/kg | Improvement cognitive functions; reversed negative effect of Aβ and scopolamine | Antioxidant and anti-inflammatory activity of thymol | [146] |
| carvacrol | Wistar Rats impaired with intrahippocampal injection of amyloid or intraperineal injection of scopolamine | Morris water maze task | intraperitoneally; 0.5, 1, or 2 mg/kg | Improvement cognitive functions; reversed negative effect of Aβ and scopolamine | Antioxidant and anti-inflammatory activity of carvacrol | [146] |
| Sprague-Dawley rats with memory impairment and neuroinflammation gained by lipopolysaccharide injected into the lateral ventricle | Object recognition task; Morris water maze test | Intraperitoneally; 25 mg/kg, 100 mg/kg | Memory improvement; significantly attenuation of induced defects in cognitive functions | Attenuation of expression of inflammatory factors in the brain (interleukin-1β, taumor necrosis factor-α, cyclooxygenase-2). Increasing expression of brain-derived neurotropic factor mRNA and decrease expression of Toll-like receptor 4 mRNA. | [147] | |
| Male Wistar rats with memory impairment obtained by lead acetate drinking water | Morris water maze test | Orally; 25,50, and 100 mg/kg | Improvement of memory and learning | Decreasing lipid peroxidation and oxidation stress by improvement of activity of antioxidant enzymes (SOD, CAT, GPx) | [148] | |
| linalool | C57BL/6J mice model anesthetized and stereotypically injected aggregated Aβ into hippocampus | Morris water maze test; passive avoidance test | intraperitoneally; 100 mg/kg | Cognitive improvement effect without negative influence on locomotor activity | Decreasing of apoptosis and oxidative stress induced by Aβ depending on Nrf2/HO-1 pathway | [104] |
| Homozygous triple transgenic AD model (3xTg-AD) and nontransgenic (Non-Tg) mice | Morris water maze test | Orally, 25 mg/kg | reversion of cognitive and emotional deficits | antioxidant activity; decreasing of β-amyloid level | [102] | |
| limonene | WKY and SHRsp rats with ischemia induced by occlusion of right middle cerebral artery—evaluation of influence of limonene on memory after stroke | Morris water maze test; novel object recognition test | Intraperitoneally; 20 mg/kg | protection against memory impairment | antioxidant effects through increasing SOD and CAT activities, decreasing MDA levels, increasing GSH levels and decreasing ROS levels. | [149] |
| α-pinene | scopolamine-induced memory deficit in C57BL/6 mice | Morris water maze test; passive avoidance test | Intraperitoneally; 10 mg/kg | memory enhancing activity and learning improvement | regulation of expression of proteins related to synthesis of acetylcholine and antioxidant defense system | [150] |
| α-terpinene | female Wistar rats | inhibitory avoidance task; | Orally; 0.5, 0.75 and 1.0 mL/kg | memory impairment | altering activity of enzymes responsible for neuronal plasticity and hydrolysis of ADP and ATP | [151] |
| Essential Oils and Main Chemical Compounds | Method of Administration | Dosage | In Vivo Model | Anxiety Model | Observed Effect, Main Results | Reference |
|---|---|---|---|---|---|---|
| α-pinene, camphene, β-pinene, β-phellandrene (Abies sachalinensis) | i.p. Inhalation | 0.6 g/kg body weight 3.6 mg/l air | Mouse | Elevated plus-maze test (EPM) | i.p. administration did not produce an anxiolytic-like effect, albeit, a significant anxiolytic-like effect was observed after inhalation. The influence of the way of administration on the distribution of the main compounds was then analyzed. The anxiolytical effect is suspected to be due to the achievement of adequate levels in the brain after inhalation | [176] |
| fragranol, fragranyl acetate, γ-terpinene, thymol, eugenol (Achillea umbellata) | p.o. | 50, 100 and 150 mg/kg | Mouse | Light/Dark transition test | The applied EO showed antianxiety properties (increased time spent in the light compartment and decreased time spent in the dark in a dose-dependent manner), but these are very likely signs of intoxication and not of possible beneficial effects of the plant volatiles. | [177] |
| p-cymene, 1,8-cineole, terpinen-4-ol, limonene (Alpinia zerumbet) | Inhalation | 0.087 ppm, 0.87 ppm and 8.7 ppm for 90 min | Mouse | Elevated plus-maze test | 0.087 and 8.7 ppm of EO obviously showed anxiolytic-like activity. | [178] |
| limonene, β-pinene, β-myrcene, sabinene (Citrus aurantium) | p.o. | 1, 5, 10, 50 mg/kg for 30 min or 14-day repeated (1, 5, 10 mg/kg/day) | Mouse | Light/Dark Box Test | The anxiolytic-like activity observed after acute (5 mg/kg) or 14-day repeated (1 mg/kg/day) dosing was mediated by the serotonergic system (5-HT1A receptors). The obtained results suggest that the EO does not function through the GABA-benzodiazepine receptor complex | [179] |
| limonene, β-pinene, β-myrcene, sabinene (Citrus aurantium) | p.o. | 0.5 and 1.0 g/kg | Mouse | Elevated plus-maze test | EPM results suggest the anxiolytic effect of the EO. | [180] |
| limonene, β-pinene, β-myrcene, sabinene (Citrus aurantium) | p.o. | Acutely pretreated at 1, 5, 10 or 50 mg/kg and 1, 5 or 10 mg/kg/day for 14 days | Mouse | Light/Dark Box test | The EO possesses a significant anxiolytic-like effect, and the present results strongly suggest the involvement of 5- HT1A-receptors. | [179] |
| limonene, β-pinene, β-myrcene, sabinene (Citrus aurantiu) | Inhalation | 1.0%, 2.5% and 5.0% for 7 min | Rat | Open-field test behavioral Social interaction test Elevated plus-maze test | The concentration of 2.5% increased both the time of the animals spent in the open arms of the EPM and the time of social interactions for rats. This was longer than that of the diazepam group (1.5 mg/kg i.p) and testifies to an anxiolytic-like effect. Results suggest a possible central action of EO | [181] |
| limonene, geranyl acetate and trans-limonene-oxide (Citrus limon) | p.o. | 50, 100 and 150 mg/kg/day for 30 days | Mouse | Elevated plus-maze test | Dosage of EO was able to significantly increase the time of permanence and the number of entrances in the open arms, indicating a positive response. The study suggests anxiolytic effects of the used EO that might involve an action on benzodiazepine-type receptors. | [182] |
| limonene, geranyl acetate and trans-limonene-oxide (Citrus limon) | Inhalation | 150 µL once a day for 2 weeks | Rat | Elevated plus-maze test | Long-term EO exposure (both male and female rats) increased anxiety in the plus-maze test, i.e., it reduced the time spent in the open arms. The obtained results indicate an anxiogenic-like effect. | [183] |
| limonene, myrcene (Citrus sinensis) | Inhalation | 100, 200, 400 μL for 5 min | Rat | Elevated plus-maze test Light/Dark Box Test | All the used doses of the EO showed anxiolytic effects. To exclude the possibility that this result was due to non-specific effects of odor exposure, the rats’ response to Melaleuca alternifolia EO was also assessed. | [184] |
| sabinene, 1,8-cineole, limonene, myrcene (Lantana camara) | Inhalation | 0.0004 and 0.04 mg for 60 min | Mouse | Open-field test | The administered EO significantly decreased locomotor activity in a dose-dependent manner. The reduction of locomotor activity may indicate anxiolytic effects. | [185] |
| linalol, linalyl acetate, terpinene-4-ol, ocimene, cineole (Lavandula angustifolia) | Inhalation | 0.5%, 2.5%, 5.0% for 15 min | Mouse | Marble-burying test | The result of the study suggests that the perception of smells is not crucial for inducing the anxiolytic-like effect in normal and anosmic mice. | [161] |
| linalol, linalyl acetate, terpinene-4-ol, ocimene, cineole (Lavandula angustifolia) | Inhalation | 4 µL/L air for 90 min | Mouse | Elevated plus-maze test | A similar anxiolytic-like effect was noticed in stressed and non-stressed mice | [186] |
| linalol, linalyl acetate, terpinene-4-ol, ocimene, cineole (Lavandula angustifolia) | Inhalation | 1–5% for 15 min | Mouse | Marble-burying test Elevated plus-maze test | The anxiolytic-like effect of lavender oil may be mediated by serotonergic transmission, and the participation of the GABAA/BDZ complex is unlikely. Administering the oil by inhalation induced rats to reduce the number of marbles buried. This occurred at a dose that did not affect locomotor activity. 5% lavender EO attenuated the serotonin syndrome induced by 40 mg/kg fluoxetine plus 80 mg/kg 5-hydroxytryptophan. | [187] |
| linalol, linalyl acetate, terpinene-4-ol, ocimene, cineole (Lavandula angustifolia) | Inhalation | Vaporized oil for 90 min | Mouse | Elevated plus-maze test | Olfactory stimulation is not necessarily required for lavender EO to show an anxiolytic action. An increase in hippocampal serotonin turn-over after lavender inhalation was, however, noticed. This enhanced the proposal of a serotonergic mediation of the anxiolytic-like effects of EO. | [188] |
| linalol, linalyl acetate, terpinene-4-ol, ocimene, cineole (Lavandula angustifolia) | Inhalation | 0.1–1.0 mL for 30 min or 1 h | Rat | Open-field test | An anxiolytic-like effect was established. Higher doses had some effects on behavior similar to those of 10 mg/kg body weight chlordiazepoxide (CDP). Doses 0.5 mL–1.0 mL increased immobility. A sedative effect can also occur at the upper doses. | [189] |
| linalol, linalyl acetate, terpinene-4-ol, ocimene, cineole (Lavandula angustifolia) | Inhalation | Refreshed three times daily with four drops of the EO by 24 h and 14 days | Gerbil | Elevated plus-maze test | Lavender EO inhalations have an anxiolytic profile in gerbils that is similar to that of the anxiolytic diazepam (1 mg/kg). Sex differences in protected head-dip (an ethological indicator of anxiety) was indicated–females showed a significant decrease in response to this test. Two-week exposure to the scent of lavender increased exploratory behavior in female gerbils, indicating a further decrease in anxiety in this sex. | [190] |
| linalol, linalyl acetate, terpinene-4-ol, ocimene, cineole (Lavandula angustifolia) | Inhalation | 1 mL of 10% EO | Sheep | Reaction to stress (isolation) | Genetic differences in temperament determined whether lavender oil alleviates or exacerbates the behavioral and/or endocrine correlates of anxiety in sheep. Exposure of calm sheep to lavender oil i.a. decreased the agitation score and the plasma concentrations of cortisol prior to isolation, compared to control, and thus they exhibited anxiolytic-like effects. The parameters of the above-mentioned tests carried out on nervous sheep were opposite, lavender oil showed an anxiogenic effect. | [191] |
| linalol, linalyl acetate, terpinene-4-ol, ocimene, cineole (Lavandula angustifolia) | p.o. | 1–30 mg/kg/day for 3 days | Mouse | Elevated plus-maze test | Anxiolytic-like effect was verified because the test mice showed an increase in open arms exploration in the elevated plus-maze compared to positive control (diazepam and pregablin). The results of the experiment suggest non-selective inhibition of voltage operated calcium channels as a mechanism of anxiolytic effect. | [162] |
| linalol, linalyl acetate, terpinene-4-ol, ocimene, cineole (Lavandula angustifolia) | p.o. | 6-week-intake 80 mg/day | Human (adults with GAD) | Hamilton Anxiety Rating Scale (HAMA) total score, Double-blind, double-dummy, randomized, reference-controlled multicentre trial | The results suggest that silexan effectively ameliorates generalized anxiety at a comparable degree to that of lorazepam (0.5 mg/day) administration. | [192] |
| linalol, linalyl acetate, terpinene-4-ol, ocimene, cineole (Lavandula angustifolia) | p.o. | 10-weeks-intake 80 mg/day | Human | Hamilton Anxiety Scale (HAMA), Double-blind, randomized, placebo-controlled multicentre trial | An anxiolytic effect was established. Symptoms of anxiety were reduced more explicitly in patients taking Silexan than that of those in the placebo group. Additionally, there was an improvement in the patients’ mental condition. | [167] |
| geranial, neral, d-limonene, β-myrcene, α-terpinyl acetate (Litsea cubeba) | p.o. | 100, 300 and 500 mg/kg/day for 7 days | Mouse | Elevated plus-maze test | An anxiolytic effect was established. L. cubeba oil has a potent effect on the central nervous system of mice. | [193] |
| thymol, γ-terpinene, trans-caryophyllene, β-selinene, p-cymene (Ocimum gratissimum) | Inhalation | 4.0 × 10−10 −4.0 × 10−1 mg for 60 min | Mouse | Light/Dark transition test | Inhalation of EO (at a concentration of 4.0 × 10−4 mg) showed anxiolytic potential and did not cause any deleterious effects on motor coordination. | [194] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojtunik-Kulesza, K.; Rudkowska, M.; Kasprzak-Drozd, K.; Oniszczuk, A.; Borowicz-Reutt, K. Activity of Selected Group of Monoterpenes in Alzheimer’s Disease Symptoms in Experimental Model Studies—A Non-Systematic Review. Int. J. Mol. Sci. 2021, 22, 7366. https://doi.org/10.3390/ijms22147366
Wojtunik-Kulesza K, Rudkowska M, Kasprzak-Drozd K, Oniszczuk A, Borowicz-Reutt K. Activity of Selected Group of Monoterpenes in Alzheimer’s Disease Symptoms in Experimental Model Studies—A Non-Systematic Review. International Journal of Molecular Sciences. 2021; 22(14):7366. https://doi.org/10.3390/ijms22147366
Chicago/Turabian StyleWojtunik-Kulesza, Karolina, Monika Rudkowska, Kamila Kasprzak-Drozd, Anna Oniszczuk, and Kinga Borowicz-Reutt. 2021. "Activity of Selected Group of Monoterpenes in Alzheimer’s Disease Symptoms in Experimental Model Studies—A Non-Systematic Review" International Journal of Molecular Sciences 22, no. 14: 7366. https://doi.org/10.3390/ijms22147366
APA StyleWojtunik-Kulesza, K., Rudkowska, M., Kasprzak-Drozd, K., Oniszczuk, A., & Borowicz-Reutt, K. (2021). Activity of Selected Group of Monoterpenes in Alzheimer’s Disease Symptoms in Experimental Model Studies—A Non-Systematic Review. International Journal of Molecular Sciences, 22(14), 7366. https://doi.org/10.3390/ijms22147366