
Minor Changes in Erythrocyte Osmotic Fragility in Trace Amine-Associated Receptor 5 (TAAR5) Knockout Mice

 , ,
, , 

Abstract
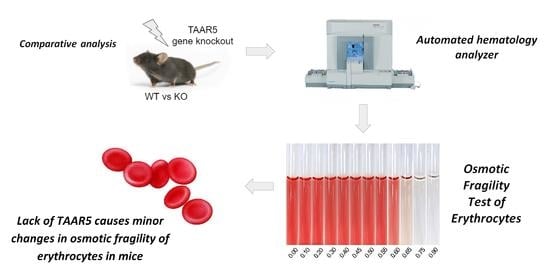
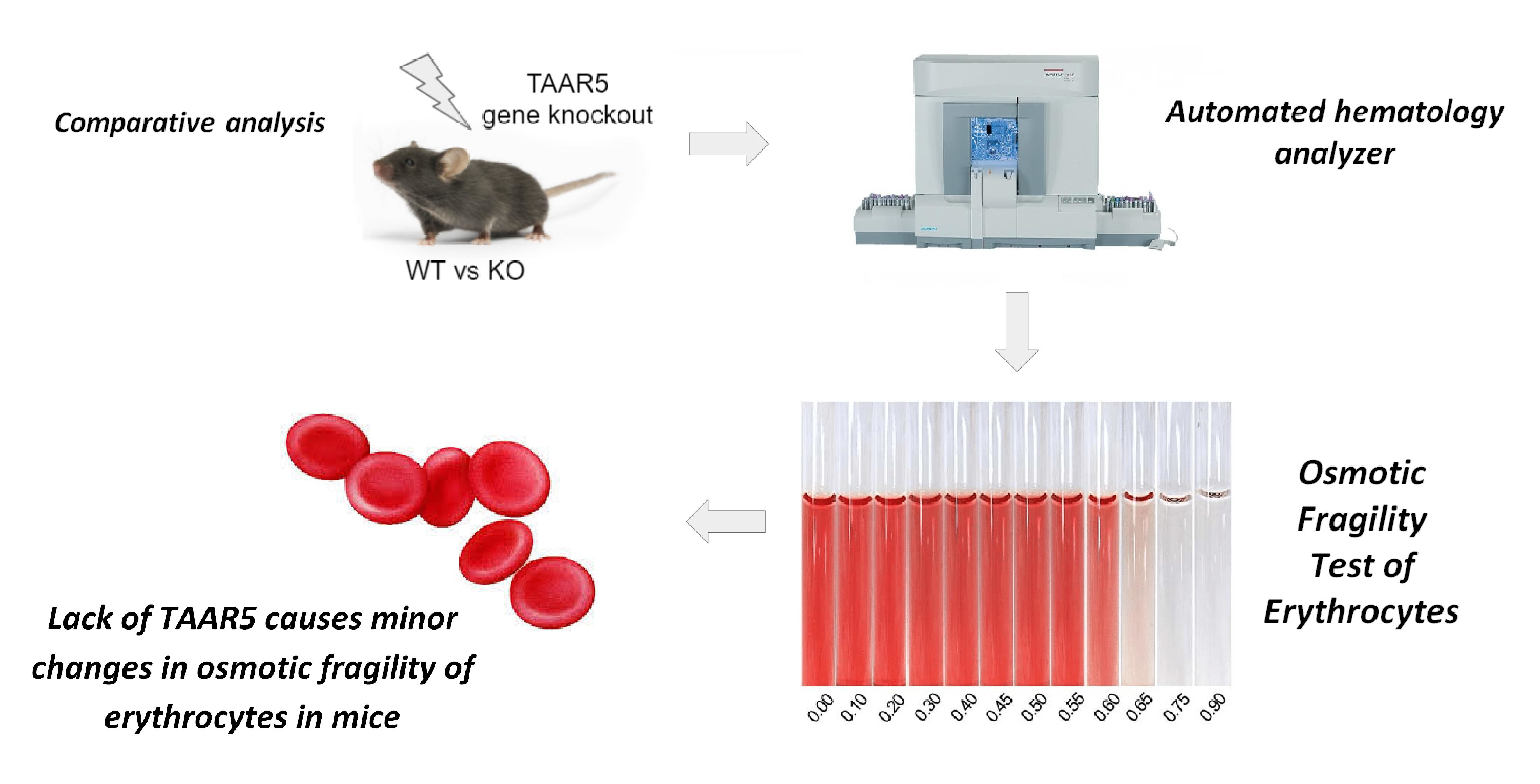{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
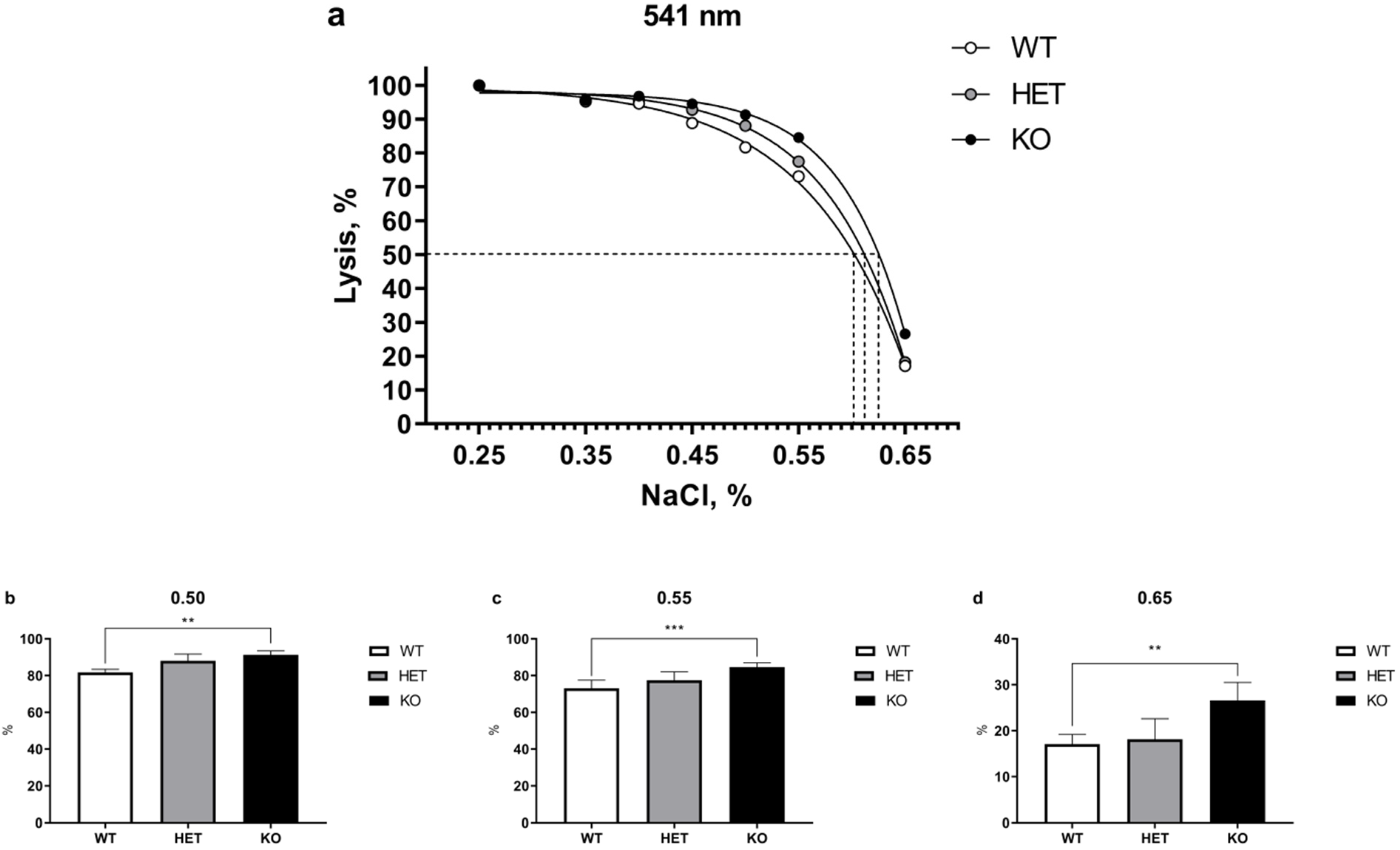
2.1. Lack of TAAR5 Gene Associated with Increased Osmotic Fragility in Mouse Erythrocytes
2.2. TAAR5 Gene Knockout Does Not Cause Significant Changes in Reticulocyte Levels
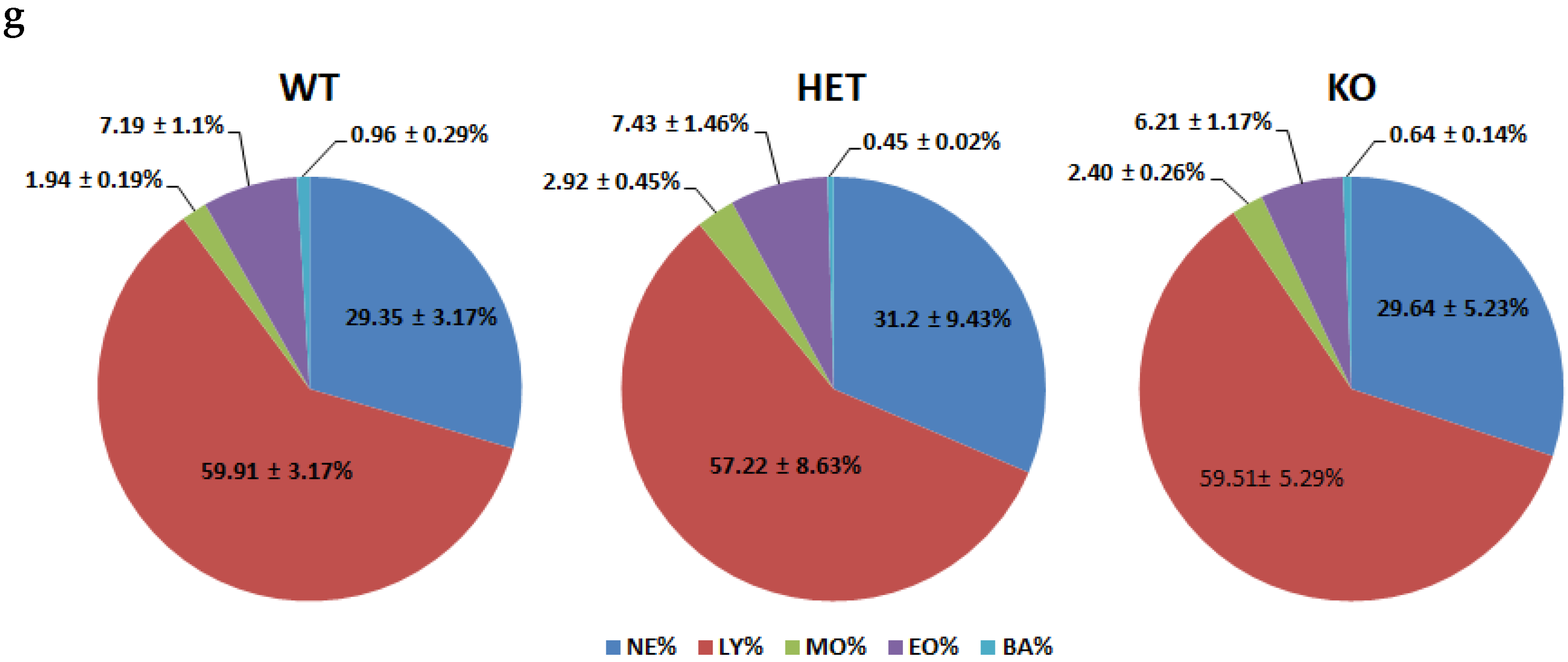
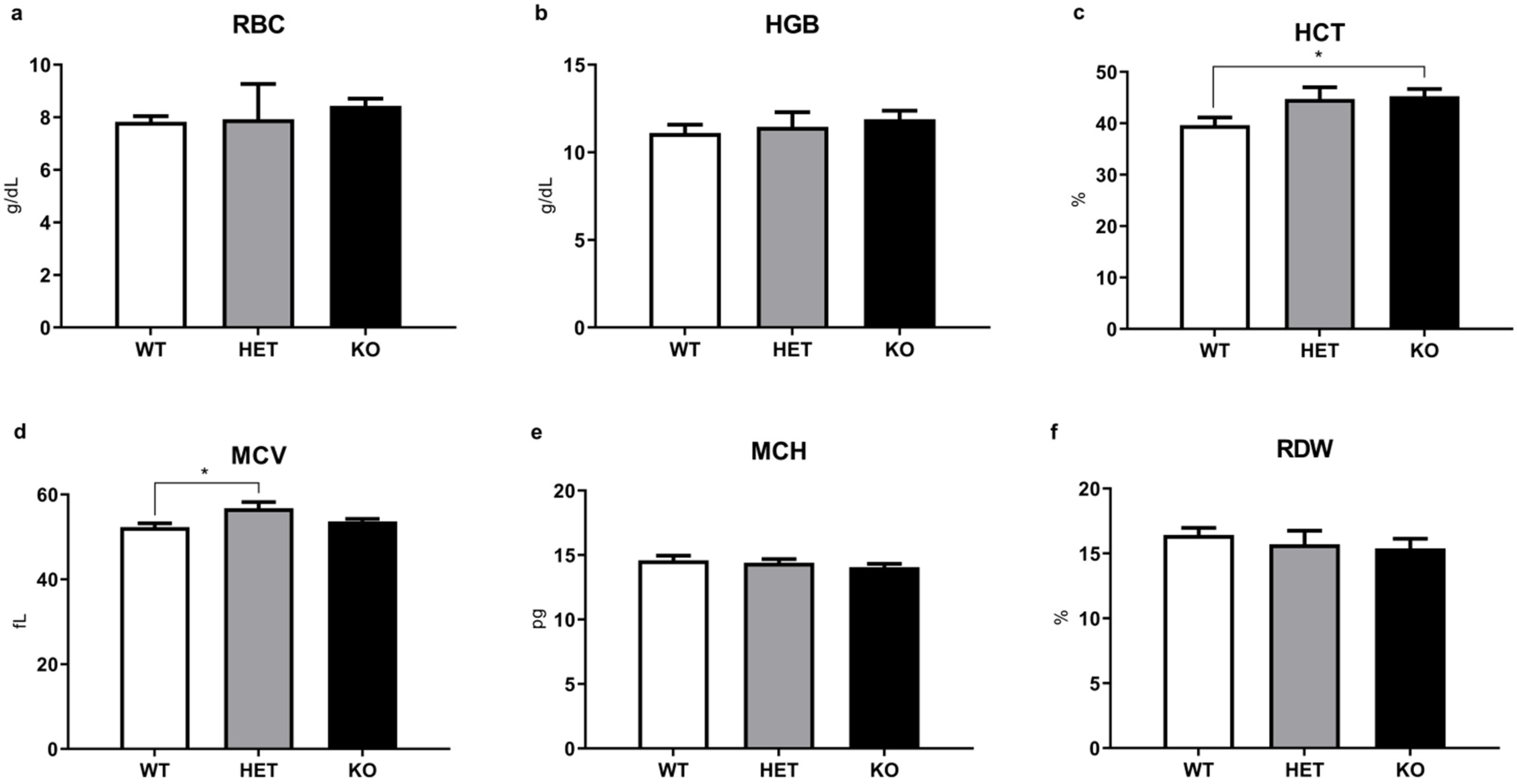
2.3. WBC and Other Routine Hematological Parameters Have Minimal Alterations in Mutant Mice
2.4. Erythrocyte TAAR5 mRNA Expression in Public Transcriptomic Datasets
3. Discussion
4. Materials and Methods
4.1. Subject
4.2. Sample Collection and Storage Methods
4.3. Osmotic Fragility Test of Erythrocytes
4.4. Measurement of Complete Blood Count Parameters
4.5. Interassay Repeatability
4.6. Statistical Analysis
4.7. Bioinformatic Analysis of RNA Seq Datasets
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boulton, A.A. Letter: Amines and theories in psychiatry. Lancet 1974, 2, 52–53. [Google Scholar] [CrossRef]
- Berry, M.D. Mammalian central nervous system trace amines. Pharmacologic amphetamines, physiologic neuromodulators. J. Neurochem. 2004, 90, 257–271. [Google Scholar] [CrossRef]
- Eiden, L.E.; Weihe, E. VMAT2: A dynamic regulator of brain monoaminergic neuronal function interacting with drugs of abuse. Ann. N. Y. Acad. Sci. 2011, 1216, 86–98. [Google Scholar] [CrossRef]
- Berry, M.D.; Gainetdinov, R.R.; Hoener, M.C.; Shahid, M. Pharmacology of human trace amine-associated receptors: Therapeutic opportunities and challenges. Pharmacol. Ther. 2017, 180, 161–180. [Google Scholar] [CrossRef]
- Gainetdinov, R.R.; Hoener, M.C.; Berry, M.D. Trace Amines and Their Receptors. Pharmacol. Rev. 2018, 70, 549–620. [Google Scholar] [CrossRef]
- Babusyte, A.; Kotthoff, M.; Fiedler, J.; Krautwurst, D. Biogenic amines activate blood leukocytes via trace amine-associated receptors TAAR1 and TAAR2. J. Leukoc. Biol. 2013, 93, 387–394. [Google Scholar] [CrossRef]
- Adriaenssens, A.; Lam, B.Y.H.; Billing, L.; Skeffington, K.; Sewing, S.; Reimann, F.; Gribble, F. A Transcriptome-Led Exploration of Molecular Mechanisms Regulating Somatostatin-Producing D-Cells in the Gastric Epithelium. Endocrinology 2015, 156, 3924–3936. [Google Scholar] [CrossRef] [PubMed]
- Raab, S.; Wang, H.; Uhles, S.; Cole, N.; Alvarez-Sanchez, R.; Künnecke, B.; Ullmer, C.; Matile, H.; Bedoucha, M.; Norcross, R.D.; et al. Incretin-like effects of small molecule trace amine-associated receptor 1 agonists. Mol. Metab. 2016, 5, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Brix, K.; Führer, D.; Biebermann, H. Molecules important for thyroid hormone synthesis and action—Known facts and future perspectives. Thyroid Res. 2011, 4 (Suppl. 1), S9. [Google Scholar] [CrossRef] [PubMed]
- Szumska, J.; Qatato, M.; Rehders, M.; Führer, D.; Biebermann, H.; Grandy, D.K.; Köhrle, J.; Brix, K. Trace Amine-Associated Receptor 1 Localization at the Apical Plasma Membrane Domain of Fisher Rat Thyroid Epithelial Cells Is Confined to Cilia. Eur. Thyroid J. 2015, 4, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Qatato, M.; Szumska, J.; Skripnik, V.; Rijntjes, E.; Köhrle, J.; Brix, K. Canonical TSH Regulation of Cathepsin-Mediated Thyroglobulin Processing in the Thyroid Gland of Male Mice Requires Taar1 Expression. Front. Pharmacol. 2018, 9, 221. [Google Scholar] [CrossRef]
- Zhukov, I.S.; Kubarskaya, L.G.; Tissen, I.Y.; Kozlova, A.A.; Dagayev, S.G.; Kashuro, V.A.; Vlasova, O.L.; Sinitca, E.L.; Karpova, I.V.; Gainetdinov, R.R. Minimal Age-Related Alterations in Behavioral and Hematological Parameters in Trace Amine-Associated Receptor 1 (TAAR1) Knockout Mice. Cell. Mol. Neurobiol. 2020, 40, 273–282. [Google Scholar] [CrossRef]
- Burchett, S.A.; Hicks, T.P. The mysterious trace amines: Protean neuromodulators of synaptic transmission in mammalian brain. Prog. Neurobiol. 2006, 79, 223–246. [Google Scholar] [CrossRef]
- Murtazina, R.Z.; Zhukov, I.S.; Korenkova, O.M.; Popova, E.A.; Kuvarzin, S.R.; Efimova, E.V.; Kubarskaya, L.G.; Batotsyrenova, E.G.; Zolotoverkhaya, E.A.; Vaganova, A.N.; et al. Genetic deletion of trace-amine associated receptor 9 (TAAR9) in rats leads to decreased blood cholesterol levels. Int. J. Mol. Sci. 2021, 22, 2942. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, S.; Sukhanov, I.; Efimova, E.V.; Kozlova, A.; Antonova, K.A.; Illiano, P.; Leo, D.; Merkulyeva, N.; Kalinina, D.; Musienko, P.; et al. Trace Amine-Associated Receptor 5 Provides Olfactory Input Into Limbic Brain Areas and Modulates Emotional Behaviors and Serotonin Transmission. Front. Mol. Neurosci. 2020, 13. [Google Scholar] [CrossRef] [PubMed]
- Efimova, E.V.; Kozlova, A.A.; Razenkova, V.; Katolikova, N.V.; Antonova, K.A.; Sotnikova, T.D.; Merkulyeva, N.S.; Veshchitskii, A.S.; Kalinina, D.S.; Korzhevskii, D.E.; et al. Increased dopamine transmission and adult neurogenesis in trace amine-associated receptor 5 (TAAR5) knockout mice. Neuropharmacology 2021, 182. [Google Scholar] [CrossRef] [PubMed]
- Dewey, M.J.; Brown, J.L.; Nallaseth, F.S. Genetic differences in red cell osmotic fragility: Analysis in allophenic mice. Blood 1982, 59, 986–989. [Google Scholar] [CrossRef]
- Bolliger, A.P.; Everds, N. Haematology of the Mouse. In The Laboratory Mouse; Elsevier Ltd.: Amsterdam, The Netherlands, 2012; pp. 331–347. ISBN 9780123820082. [Google Scholar]
- Kahn, S.E.; Watkins, B.F.; Bermes, E.W. An evaluation of a spectrophotometric scanning technique for measurement of plasma hemoglobin. Ann. Clin. Lab. Sci. 1981, 11, 126–131. [Google Scholar] [PubMed]
- Vettore, L.; Zanella, A.; Molaro, L.; De Matteis, C.; Pavesi, M.; Mariani, M. A New Test for the Laboratory Diagnosis of Spherocytosis. Acta Haematol. 1984, 72, 258–263. [Google Scholar] [CrossRef]
- O’Connell, K.E.; Mikkola, A.M.; Stepanek, A.M.; Vernet, A.; Hall, C.D.; Sun, C.C.; Yildirim, E.; Staropoli, J.F.; Lee, J.T.; Brown, D.E. Practical murine hematopathology: A comparative review and implications for research. Comp. Med. 2015, 65, 96–113. [Google Scholar]
- Kishimoto, S.; Maruhashi, T.; Kajikawa, M.; Matsui, S.; Hashimoto, H.; Takaeko, Y.; Harada, T.; Yamaji, T.; Han, Y.; Kihara, Y.; et al. Hematocrit, hemoglobin and red blood cells are associated with vascular function and vascular structure in men. Sci. Rep. 2020, 10, 11467. [Google Scholar] [CrossRef] [PubMed]
- Walski, T.; Chludzińska, L.; Komorowska, M.; Witkiewicz, W. Individual osmotic fragility distribution: A new parameter for determination of the osmotic properties of human red blood cells. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Rocha, S.; Costa, E.; Rocha-Pereira, P.; Ferreira, F.; Cleto, E.; Barbot, J.; Quintanilha, A.; Belo, L.; Santos-Silva, A. Complementary markers for the clinical severity classification of hereditary spherocytosis in unsplenectomized patients. Blood Cells Mol. Dis. 2011, 46, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Danise, P.; Amendola, G.; Di Concilio, R.; Cillari, E.; Gioia, M.; Di Palma, A.; Avino, D.; Rigano, P.; Maggio, A. Nucleated red blood cells and soluble transferrin receptor in thalassemia syndromes: Relationship with global and ineffective erythropoiesis. Clin. Chem. Lab. Med. 2009, 47, 1539–1542. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Barcellini, W.; Fattizzo, B. Clinical Applications of Hemolytic Markers in the Differential Diagnosis and Management of Hemolytic Anemia. Dis. Markers 2015, 2015. [Google Scholar] [CrossRef]
- Hattangadi, S.M.; Wong, P.; Zhang, L.; Flygare, J.; Lodish, H.F. From stem cell to red cell: Regulation of erythropoiesis at multiple levels by multiple proteins, RNAs, and chromatin modifications. Blood 2011, 118, 6258–6268. [Google Scholar] [CrossRef]
- Reinhart, W.H.; Chien, S. Echinocyte-stomatocyte transformation and shape control of human red blood cells: Morphological aspects. Am. J. Hematol. 1987, 24, 1–14. [Google Scholar] [CrossRef]
- Bosman, G.J.C.G.M.; Willekens, F.L.A.; Werre, J.M. Erythrocyte aging: A more than superficial resemblance to apoptosis? Cell. Physiol. Biochem. 2005, 16, 1–8. [Google Scholar] [CrossRef]
- Föller, M.; Huber, S.M.; Lang, F. Erythrocyte programmed cell death. IUBMB Life 2008, 60, 661–668. [Google Scholar] [CrossRef]
- Lang, K.S.; Lang, P.A.; Bauer, C.; Duranton, C.; Wieder, T.; Huber, S.M.; Lang, F. Mechanisms of suicidal erythrocyte death. Cell. Physiol. Biochem. 2005, 15, 195–202. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhukov, I.S.; Kubarskaya, L.G.; Karpova, I.V.; Vaganova, A.N.; Karpenko, M.N.; Gainetdinov, R.R. Minor Changes in Erythrocyte Osmotic Fragility in Trace Amine-Associated Receptor 5 (TAAR5) Knockout Mice. Int. J. Mol. Sci. 2021, 22, 7307. https://doi.org/10.3390/ijms22147307
Zhukov IS, Kubarskaya LG, Karpova IV, Vaganova AN, Karpenko MN, Gainetdinov RR. Minor Changes in Erythrocyte Osmotic Fragility in Trace Amine-Associated Receptor 5 (TAAR5) Knockout Mice. International Journal of Molecular Sciences. 2021; 22(14):7307. https://doi.org/10.3390/ijms22147307
Chicago/Turabian StyleZhukov, Ilya S., Larisa G. Kubarskaya, Inessa V. Karpova, Anastasia N. Vaganova, Marina N. Karpenko, and Raul R. Gainetdinov. 2021. "Minor Changes in Erythrocyte Osmotic Fragility in Trace Amine-Associated Receptor 5 (TAAR5) Knockout Mice" International Journal of Molecular Sciences 22, no. 14: 7307. https://doi.org/10.3390/ijms22147307
APA StyleZhukov, I. S., Kubarskaya, L. G., Karpova, I. V., Vaganova, A. N., Karpenko, M. N., & Gainetdinov, R. R. (2021). Minor Changes in Erythrocyte Osmotic Fragility in Trace Amine-Associated Receptor 5 (TAAR5) Knockout Mice. International Journal of Molecular Sciences, 22(14), 7307. https://doi.org/10.3390/ijms22147307