
Cytokine/Chemokine Release Patterns and Transcriptomic Profiles of LPS/IFNγ-Activated Human Macrophages Differentiated with Heat-Killed Mycobacterium obuense, M-CSF, or GM-CSF


 , ,
, ,
Abstract
1. Introduction
2. Results
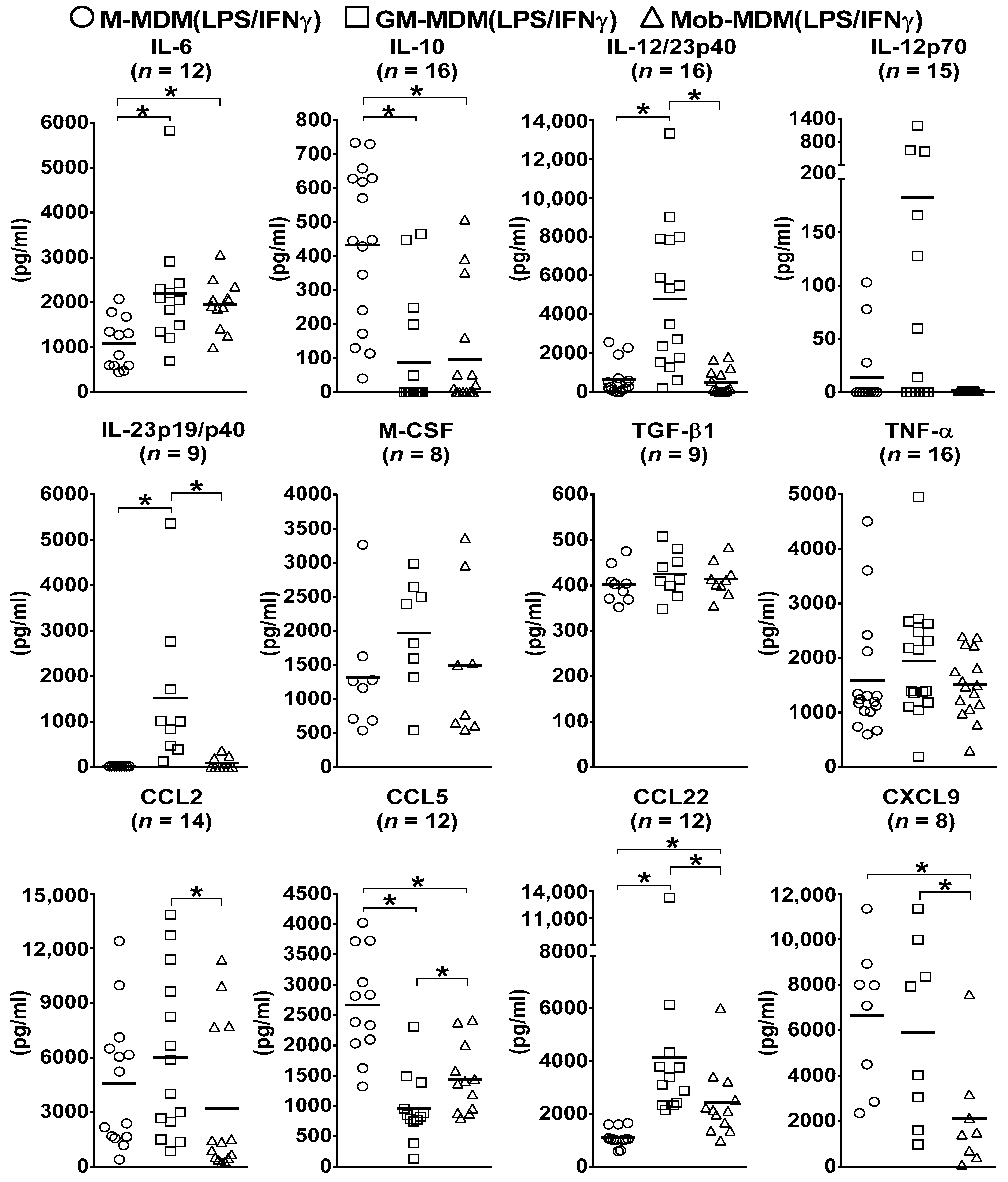
2.1. Patterns of Cytokine and Chemokine Secretion by M-MDM(LPS/IFNγ), GM-MDM(LPS/IFNγ), and Mob-MDM(LPS/IFNγ)
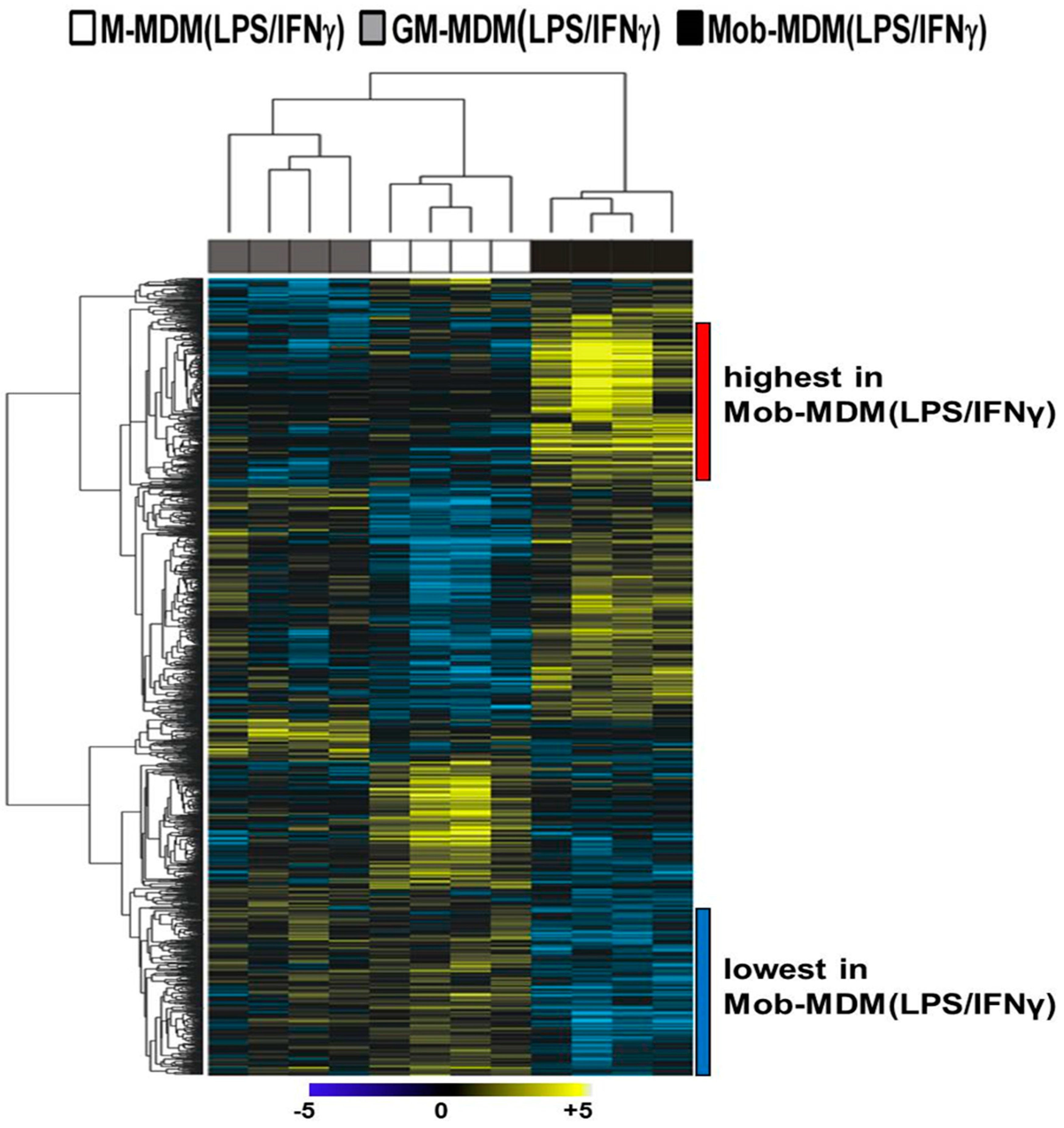
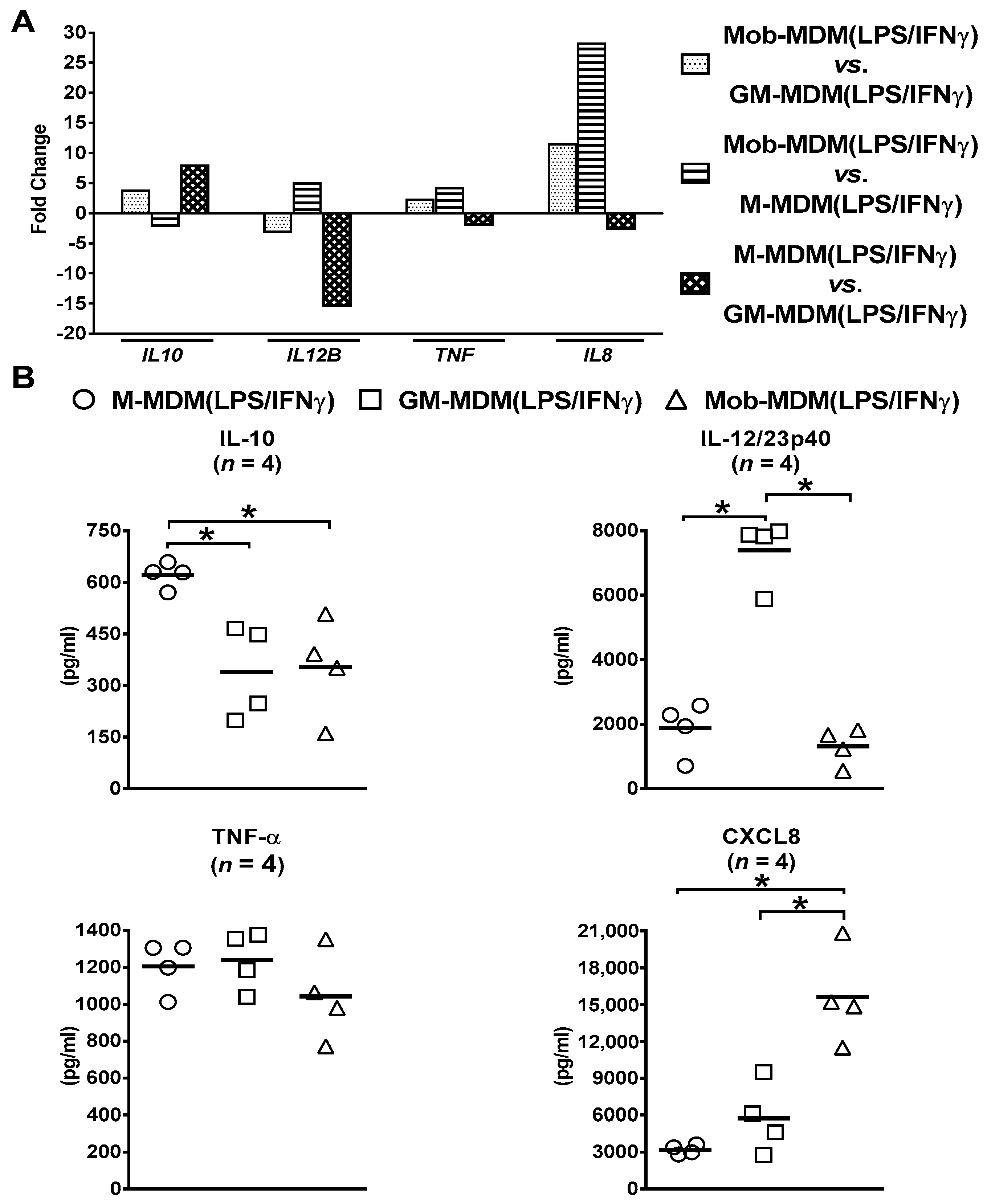
2.2. Comparative Whole-Transcriptome Analysis of M-MDM(LPS/IFNγ), GM-MDM(LPS/IFNγ), and Mob-MDM(LPS/IFNγ)
3. Discussion
4. Materials and Methods
4.1. Blood Collection
4.2. Monocyte Isolation and Generation of MDMs
4.3. Activation of MDMs
4.4. Measurement of Cytokine and Chemokine Levels in MDM Culture Supernatants
4.5. RNA Extraction from MDM
4.6. Library Preparation and RNA-Seq
4.7. RNA-Seq Data Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage activation and polarization: Nomenclature and experimental guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef]
- Italiani, P.; Boraschi, D. From monocytes to M1/M2 macrophages: Phenotypical vs. functional differentiation. Front. Immunol. 2014, 5, 514. [Google Scholar] [CrossRef] [PubMed]
- Orecch, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Corrigendum: Macrophage polarization: Different gene signatures in M1(LPS+) Vs. classically and M2(LPS-) vs. alternatively activated macrophages. Front. Immunol. 2020, 11, 234. [Google Scholar] [CrossRef]
- Lacey, D.C.; Achuthan, A.; Fleetwood, A.J.; Dinh, H.; Roiniotis, J.; Scholz, G.M.; Chang, M.W.; Beckman, S.K.; Cook, A.D.; Hamilton, J.A. Defining GM-CSF- and macrophage-CSF-dependent macrophage responses by in vitro models. J. Immunol. 2012, 188, 5752–5765. [Google Scholar] [CrossRef] [PubMed]
- Buchacher, T.; Ohradanova-Repic, A.; Stockinger, H.; Fischer, M.B.; Weber, V. M2 polarization of human macrophages favors survival of the intracellular pathogen chlamydia pneumoniae. PLoS ONE 2015, 10, e0143593. [Google Scholar] [CrossRef] [PubMed]
- Trus, E.; Basta, S.; Gee, K. Who’s in charge here? Macrophage colony stimulating factor and granulocyte macrophage colony stimulating factor: Competing factors in macrophage polarization. Cytokine 2020, 127, 154939. [Google Scholar] [CrossRef]
- Rey-Giraud, F.; Hafner, M.; Ries, C.H. In vitro generation of monocyte-derived macrophages under serum-free conditions improves their tumor promoting functions. PLoS ONE 2012, 7, e42656. [Google Scholar] [CrossRef] [PubMed]
- Vogel, D.Y.; Glim, J.E.; Stavenuiter, A.W.; Breur, M.; Heijnen, P.; Amor, S.; Dijkstra, C.D.; Beelen, R.H. Human macrophage polarization in vitro: Maturation and activation methods compared. Immunobiology 2014, 219, 695–703. [Google Scholar] [CrossRef]
- Rook, G.A.; Adams, V.; Hunt, J.; Palmer, R.; Martinelli, R.; Brunet, L.R. Mycobacteria and other environmental organisms as immunomodulators for immunoregulatory disorders. Springer Semin. Immunopathol. 2004, 25, 237–255. [Google Scholar] [CrossRef]
- Kleinnijenhuis, J.; Oosting, M.; Joosten, L.A.; Netea, M.G.; Van Crevel, R. Innate immune recognition of mycobacterium tuberculosis. Clin. Dev. Immunol. 2011, 2011, 405310. [Google Scholar] [CrossRef]
- Killick, K.E.; Ni Cheallaigh, C.; O’Farrelly, C.; Hokamp, K.; MacHugh, D.E.; Harris, J. Receptor-mediated recognition of mycobacterial pathogens. Cell. Microbiol. 2013, 15, 1484–1495. [Google Scholar] [CrossRef] [PubMed]
- Stebbing, J.; Dalgleish, A.; Gifford-Moore, A.; Martin, A.; Gleeson, C.; Wilson, G.; Brunet, L.R.; Grange, J.; Mudan, S. An intra-patient placebo-controlled phase I trial to evaluate the safety and tolerability of intradermal IMM-101 in melanoma. Ann. Oncol. 2012, 23, 1314–1319. [Google Scholar] [CrossRef] [PubMed]
- Dalgleish, A.G.; Stebbing, J.; Adamson, D.J.; Arif, S.S.; Bidoli, P.; Chang, D.; Cheeseman, S.; Diaz-Beveridge, R.; Fernandez-Martos, C.; Glynne-Jones, R.; et al. Randomised, open-label, phase II study of gemcitabine with and without IMM-101 for advanced pancreatic cancer. Br. J. Cancer 2016, 115, 789–796. [Google Scholar] [CrossRef]
- Dalgleish, A.G.; Mudan, S.; Fusi, A. Enhanced effect of checkpoint inhibitors when given after or together with IMM-101: Significant responses in four advanced melanoma patients with no additional major toxicity. J. Transl. Med. 2018, 16, 227–228. [Google Scholar] [CrossRef] [PubMed]
- Fowler, D.; Copier, J.; Wilson, N.; Dalgleish, A.G.; Bodman-Smith, M.D. Mycobacteria activate gammadelta T-cell anti-tumour responses via cytokines from type 1 myeloid dendritic cells: A mechanism of action for cancer immunotherapy. Cancer Immunol. Immunother. 2012, 61, 535–547. [Google Scholar] [CrossRef]
- Bazzi, S.; Modjtahedi, H.; Mudan, S.; Akle, C.; Bahr, G.M. Analysis of the immunomodulatory properties of two heat-killed mycobacterial preparations in a human whole blood model. Immunobiology 2015, 220, 1293–1304. [Google Scholar] [CrossRef] [PubMed]
- Bazzi, S.; Modjtahedi, H.; Mudan, S.; Achkar, M.; Akle, C.; Bahr, G.M. Immunomodulatory effects of heat-killed mycobacterium obuense on human blood dendritic cells. Innate Immun. 2017, 1753425917727838. [Google Scholar] [CrossRef]
- Bazzi, S.; El-Darzi, E.; McDowell, T.; Modjtahedi, H.; Mudan, S.; Achkar, M.; Akle, C.; Kadara, H.; Bahr, G.M. Defining genome-wide expression and phenotypic contextual cues in macrophages generated by granulocyte/macrophage colony-stimulating factor, macrophage colony-stimulating factor, and heat-killed mycobacteria. Front. Immunol. 2017, 8, 1253. [Google Scholar] [CrossRef]
- Verreck, F.A.; de Boer, T.; Langenberg, D.M.; Hoeve, M.A.; Kramer, M.; Vaisberg, E.; Kastelein, R.; Kolk, A.; de Waal-Malefyt, R.; Ottenhoff, T.H. Human IL-23-producing type 1 macrophages promote but IL-10-producing type 2 macrophages subvert immunity to (myco)bacteria. Proc. Natl. Acad. Sci. USA 2004, 101, 4560–4565. [Google Scholar] [CrossRef]
- Pahl, J.H.; Kwappenberg, K.M.; Varypataki, E.M.; Santos, S.J.; Kuijjer, M.L.; Mohamed, S.; Wijnen, J.T.; van Tol, M.J.; Cleton-Jansen, A.M.; Egeler, R.M.; et al. Macrophages inhibit human osteosarcoma cell growth after activation with the bacterial cell wall derivative liposomal muramyl tripeptide in combination with interferon-gamma. J. Exp. Clin. Cancer Res. 2014, 33, 27. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Xue, J.; Schmidt, S.V.; Sander, J.; Draffehn, A.; Krebs, W.; Quester, I.; De Nardo, D.; Gohel, T.D.; Emde, M.; Schmidleithner, L.; et al. Transcriptome-based network analysis reveals a spectrum model of human macrophage activation. Immunity 2014, 40, 274–288. [Google Scholar] [CrossRef]
- Hamilton, T.A.; Zhao, C.; Pavicic, P.G., Jr.; Datta, S. Myeloid colony-stimulating factors as regulators of macrophage polarization. Front. Immunol. 2014, 5, 554. [Google Scholar] [CrossRef]
- Cananzi, F.C.; Mudan, S.; Dunne, M.; Belonwu, N.; Dalgleish, A.G. Long-term survival and outcome of patients originally given mycobacterium vaccae for metastatic malignant melanoma. Hum. Vaccin Immunother. 2013, 9, 2427–2433. [Google Scholar] [CrossRef][Green Version]
- von Reyn, C.F.; Mtei, L.; Arbeit, R.D.; Waddell, R.; Cole, B.; Mackenzie, T.; Matee, M.; Bakari, M.; Tvaroha, S.; Adams, L.V.; et al. Prevention of tuberculosis in Bacille Calmette-Guerin-primed, HIV-infected adults boosted with an inactivated whole-cell mycobacterial vaccine. AIDS 2010, 24, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Decaestecker, K.; Oosterlinck, W. Managing the adverse events of intravesical bacillus Calmette-Guerin therapy. Res. Rep. Urol. 2015, 7, 157–163. [Google Scholar]
- Ehrt, S.; Schnappinger, D.; Bekiranov, S.; Drenkow, J.; Shi, S.; Gingeras, T.R.; Gaasterland, T.; Schoolnik, G.; Nathan, C. Reprogramming of the macrophage transcriptome in response to interferon-gamma and Mycobacterium tuberculosis: Signaling roles of nitric oxide synthase-2 and phagocyte oxidase. J. Exp. Med. 2001, 194, 1123–1140. [Google Scholar] [CrossRef] [PubMed]
- Thiriot, J.D.; Martinez-Martinez, Y.B.; Endsley, J.J.; Torres, A.G. Hacking the host: Exploitation of macrophage polarization by intracellular bacterial pathogens. Pathog. Dis. 2020, 78. [Google Scholar] [CrossRef] [PubMed]
- Verreck, F.A.; de Boer, T.; Langenberg, D.M.; van der Zanden, L.; Ottenhoff, T.H. Phenotypic and functional profiling of human proinflammatory type-1 and anti-inflammatory type-2 macrophages in response to microbial antigens and IFN-gamma- and CD40L-mediated costimulation. J. Leukoc. Biol. 2006, 79, 285–293. [Google Scholar] [CrossRef]
- Quero, L.; Hanser, E.; Manigold, T.; Tiaden, A.N.; Kyburz, D. TLR2 stimulation impairs anti-inflammatory activity of M2-like macrophages, generating a chimeric M1/M2 phenotype. Arthritis Res. Ther. 2017, 19, 1–13. [Google Scholar] [CrossRef]
- Jaguin, M.; Houlbert, N.; Fardel, O.; Lecureur, V. Polarization profiles of human M-CSF-generated macrophages and comparison of M1-markers in classically activated macrophages from GM-CSF and M-CSF origin. Cell. Immunol. 2013, 281, 51–61. [Google Scholar] [CrossRef]
- Bayer, C.; Varani, S.; Wang, L.; Walther, P.; Zhou, S.; Straschewski, S.; Bachem, M.; Soderberg-Naucler, C.; Mertens, T.; Frascaroli, G. Human cytomegalovirus infection of M1 and M2 macrophages triggers inflammation and autologous T-cell proliferation. J. Virol. 2013, 87, 67–79. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Locati, M. Macrophage polarization comes of age. Immunity 2005, 23, 344–346. [Google Scholar] [CrossRef]
- Roszer, T. Understanding the mysterious M2 macrophage through activation markers and effector mechanisms. Mediat. Inflamm. 2015, 2015, 816460. [Google Scholar] [CrossRef]
- Mia, S.; Warnecke, A.; Zhang, X.M.; Malmstrom, V.; Harris, R.A. An optimized protocol for human M2 macrophages using M-CSF and IL-4/IL-10/TGF-beta yields a dominant immunosuppressive phenotype. Scand. J. Immunol. 2014, 79, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Yuksel, Z.S.; Buber, E.; Kocagoz, T.; Alp, A.; Saribas, Z.; Acan, N.L. Mycobacterial strains that stimulate the immune system most efficiently as candidates for the treatment of bladder cancer. J. Mol. Microbiol. Biotechnol. 2011, 20, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Raman, D.; Sobolik-Delmaire, T.; Richmond, A. Chemokines in health and disease. Exp. Cell Res. 2011, 317, 575–589. [Google Scholar] [CrossRef]
- Groom, J.R.; Luster, A.D. CXCR3 ligands: Redundant, collaborative and antagonistic functions. Immunol. Cell Biol. 2011, 89, 207–215. [Google Scholar] [CrossRef]
- Metzemaekers, M.; Vanheule, V.; Janssens, R.; Struyf, S.; Proost, P. Overview of the mechanisms that may contribute to the non-redundant activities of interferon-inducible CXC chemokine receptor 3 ligands. Front. Immunol. 2018, 8, 1970. [Google Scholar] [CrossRef]
- Bachmann, M.; Scheiermann, P.; Hardle, L.; Pfeilschifter, J.; Muhl, H. IL-36gamma/IL-1F9, an innate T-bet target in myeloid cells. J. Biol. Chem. 2012, 287, 41684–41696. [Google Scholar] [CrossRef] [PubMed]
- Vigne, S.; Palmer, G.; Martin, P.; Lamacchia, C.; Strebel, D.; Rodriguez, E.; Olleros, M.L.; Vesin, D.; Garcia, I.; Ronchi, F.; et al. IL-36 signaling amplifies Th1 responses by enhancing proliferation and Th1 polarization of naive CD4+ T cells. Blood 2012, 120, 3478–3487. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, X.; Feng, C.; Weinstein, A.; Xia, R.; Wen, W.; Lv, Q.; Zuo, S.; Tang, P.; Yang, X.; et al. IL-36gamma transforms the tumor microenvironment and promotes type 1 lymphocyte-mediated antitumor immune responses. Cancer Cell. 2015, 28, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Marisa, L.; de Reynies, A.; Duval, A.; Selves, J.; Gaub, M.P.; Vescovo, L.; Etienne-Grimaldi, M.C.; Schiappa, R.; Guenot, D.; Ayadi, M.; et al. Gene expression classification of colon cancer into molecular subtypes: Characterization, validation, and prognostic value. PLoS Med. 2013, 10, e1001453. [Google Scholar] [CrossRef]
- Weinstein, A.M.; Giraldo, N.A.; Petitprez, F.; Julie, C.; Lacroix, L.; Peschaud, F.; Emile, J.F.; Marisa, L.; Fridman, W.H.; Storkus, W.J.; et al. Association of IL-36gamma with tertiary lymphoid structures and inflammatory immune infiltrates in human colorectal cancer. Cancer Immunol. Immunother. 2019, 68, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Wein, A.N.; Dunbar, P.R.; McMaster, S.R.; Li, Z.T.; Denning, T.L.; Kohlmeier, J.E. IL-36gamma protects against severe influenza infection by promoting lung alveolar macrophage survival and limiting viral replication. J. Immunol. 2018, 201, 573–582. [Google Scholar] [CrossRef]
- Cesar-Razquin, A.; Snijder, B.; Frappier-Brinton, T.; Isserlin, R.; Gyimesi, G.; Bai, X.; Reithmeier, R.A.; Hepworth, D.; Hediger, M.A.; Edwards, A.M.; et al. A call for systematic research on solute carriers. Cell 2015, 162, 478–487. [Google Scholar] [CrossRef]
- Cai, Y.; Yang, Q.; Liao, M.; Wang, H.; Zhang, C.; Nambi, S.; Wang, W.; Zhang, M.; Wu, J.; Deng, G.; et al. xCT increases tuberculosis susceptibility by regulating antimicrobial function and inflammation. Oncotarget 2016, 7, 31001–31013. [Google Scholar] [CrossRef]
- Staiano, R.I.; Granata, F.; Secondo, A.; Petraroli, A.; Loffredo, S.; Annunziato, L.; Triggiani, M.; Marone, G. Human macrophages and monocytes express functional Na(+)/Ca (2+) exchangers 1 and 3. Adv. Exp. Med. Biol. 2013, 961, 317–326. [Google Scholar]
- Yin, S.S.; Gao, F.H. Molecular mechanism of tumor cell immune escape mediated by CD24/Siglec-10. Front. Immunol. 2020, 11, 1324. [Google Scholar] [CrossRef]
- Barkal, A.A.; Brewer, R.E.; Markovic, M.; Kowarsky, M.; Barkal, S.A.; Zaro, B.W.; Krishnan, V.; Hatakeyama, J.; Dorigo, O.; Barkal, L.J.; et al. CD24 signalling through macrophage Siglec-10 is a target for cancer immunotherapy. Nature 2019, 572, 392–396. [Google Scholar] [CrossRef]
- Wang, N.; Liang, H.; Zen, K. Molecular mechanisms that influence the macrophage M1–M2 polarization balance. Front. Immunol. 2014, 5, 614. [Google Scholar] [CrossRef]
- Pagliari, L.J.; Perlman, H.; Liu, H.; Pope, R.M. Macrophages require constitutive NF-kappaB activation to maintain A1 expression and mitochondrial homeostasis. Mol. Cell. Biol. 2000, 20, 8855–8865. [Google Scholar] [CrossRef]
- Tak, P.P.; Firestein, G.S. NF-kappaB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef]
- Connelly, L.; Barham, W.; Onishko, H.M.; Chen, L.; Sherrill, T.P.; Zabuawala, T.; Ostrowski, M.C.; Blackwell, T.S.; Yull, F.E. NF-kappaB activation within macrophages leads to an anti-tumor phenotype in a mammary tumor lung metastasis model. Breast Cancer Res. 2011, 13, R83. [Google Scholar] [CrossRef]
- Liu, X.; Li, J.; Peng, X.; Lv, B.; Wang, P.; Zhao, X.; Yu, B. Geraniin inhibits LPS-induced THP-1 macrophages switching to M1 phenotype via SOCS1/NF-kappaB pathway. Inflammation 2016, 39, 1421–1433. [Google Scholar] [CrossRef] [PubMed]
- Lo, T.H.; Tseng, K.Y.; Tsao, W.S.; Yang, C.Y.; Hsieh, S.L.; Chiu, A.W.; Takai, T.; Mak, T.W.; Tarng, D.C.; Chen, N.J. TREM-1 regulates macrophage polarization in ureteral obstruction. Kidney Int. 2014, 86, 1174–1186. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Syed, M.A.; Panchal, D.; Joo, M.; Colonna, M.; Brantly, M.; Sadikot, R.T. Triggering receptor expressed on myeloid cells 1 (TREM-1)-mediated Bcl-2 induction prolongs macrophage survival. J. Biol. Chem. 2014, 289, 15118–15129. [Google Scholar] [CrossRef] [PubMed]
- Bryson, B.D.; Rosebrock, T.R.; Tafesse, F.G.; Itoh, C.Y.; Nibasumba, A.; Babunovic, G.H.; Corleis, B.; Martin, C.; Keegan, C.; Andrade, P.; et al. Heterogeneous GM-CSF signaling in macrophages is associated with control of Mycobacterium tuberculosis. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Singh, V.K.; Actor, J.K.; Hunter, R.L.; Jagannath, C.; Subbian, S.; Khan, A. GM-CSF dependent differential control of Mycobacterium tuberculosis infection in human and mouse macrophages: Is macrophage source of GM-CSF critical to tuberculosis immunity? Front. Immunol. 2020, 11, 1599. [Google Scholar] [CrossRef]
- Anderson, N.R.; Minutolo, N.G.; Gill, S.; Klichinsky, M. Macrophage-based approaches for cancer immunotherapy. Cancer Res. 2020, 81, 1201–1208. [Google Scholar] [CrossRef] [PubMed]
- Klichinsky, M.; Ruella, M.; Shestova, O.; Lu, X.M.; Best, A.; Zeeman, M.; Schmierer, M.; Gabrusiewicz, K.; Anderson, N.R.; Petty, N.E.; et al. Human chimeric antigen receptor macrophages for cancer immunotherapy. Nat. Biotechnol. 2020, 38, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Xing, Y.; Yu, T.; Wu, Y.N.; Roy, M.; Kim, J.; Lee, C. An expectation-maximization algorithm for probabilistic reconstructions of full-length isoforms from splice graphs. Nucleic Acids Res. 2006, 34, 3150–3160. [Google Scholar] [CrossRef] [PubMed]
- Simon, R.; Lam, A.; Li, M.C.; Ngan, M.; Menenzes, S.; Zhao, Y. Analysis of gene expression data using BRB-ArrayTools. Cancer. Inform. 2007, 3, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Perou, C.M.; Sorlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef]
- Kadara, H.; Shen, L.; Fujimoto, J.; Saintigny, P.; Chow, C.W.; Lang, W.; Chu, Z.; Garcia, M.; Kabbout, M.; Fan, Y.H.; et al. Characterizing the molecular spatial and temporal field of injury in early-stage smoker non-small cell lung cancer patients after definitive surgery by expression profiling. Cancer. Prev. Res. 2013, 6, 8–17. [Google Scholar] [CrossRef]
- Kadara, H.; Fujimoto, J.; Yoo, S.Y.; Maki, Y.; Gower, A.C.; Kabbout, M.; Garcia, M.M.; Chow, C.W.; Chu, Z.; Mendoza, G.; et al. Transcriptomic architecture of the adjacent airway field cancerization in non-small cell lung cancer. J. Natl. Cancer Inst. 2014, 106, dju004. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| RefSeq Transcript ID | Gene Symbol | Entrez Gene Name | FC (vs. GM-MDM(LPS/IFNγ)) |
|---|---|---|---|
| NM_002933 | RNASE1 | Ribonuclease, RNase A family, 1 | 20.65 |
| NM_006274 | CCL19 | Chemokine (C-C motif) ligand 19 | 18.02 |
| NM_001400 | S1PR1 | sphingosine-1-phosphate receptor 1 | 11.92 |
| NM_000240 | MAOA | monoamine oxidase A | 10.01 |
| NM_000572 | IL10 | Interleukin 10 | 7.88 |
| NM_203416 | CD163 | CD163 molecule | 7.44 |
| NM_004244 | CD163 | CD163 molecule | 7.38 |
| NM_015136 | STAB1 | Stabilin 1 | 6.72 |
| NM_001270471 | SOCS2 | Suppressor of cytokine signaling 2 | 5.16 |
| NM_001270467 | SOCS2 | Suppressor of cytokine signaling 2 | 3.97 |
| NM_014479 | ADAMDEC1 | ADAM-like, decysin 1 | 3.64 |
| NM_001127443 | CD36 | CD36 molecule | 3.55 |
| NM_002987 | CCL17 | Chemokine (C-C motif) ligand 17 | 3.45 |
| NM_003877 | SOCS2 | Suppressor of cytokine signaling 2 | 3.22 |
| NM_004994 | MMP9 | Matrix metallopeptidase 9 | 2.82 |
| NM_001127444 | CD36 | CD36 molecule | 2.82 |
| NM_001001547 | CD36 | CD36 molecule | 2.63 |
| NM_001001548 | CD36 | CD36 molecule | 2.62 |
| NM_181054 | HIF1A | Hypoxia-inducible factor 1, alpha subunit (basic helix–loop–helix transcription factor) | 2.48 |
| NM_000072 | CD36 | CD36 molecule | 2.29 |
| NM_005410 | SEPP1 | Selenoprotein P, plasma, 1 | 2.13 |
| NM_016584 | IL23A | Interleukin 23 subunit alpha (p19) | −2.34 |
| NM_000584 | IL8 | Chemokine (C-X-C motif) ligand 8 | −2.46 |
| NM_000759 | CSF3 | Colony-stimulating factor 3 | −2.49 |
| NM_003775 | S1PR4 | Sphingosine-1-phosphate receptor 4 | −2.64 |
| NM_001764 | CD1B | CD1b molecule | −3.39 |
| NM_000575 | IL1A | Interleukin 1 alpha | −3.63 |
| NM_002438.1 | MRC1 | Mannose receptor, C type 1 (CD206) | −3.96 |
| NM_001025194 | CES1 | Carboxylesterase 1 | −3.97 |
| NM_002990 | CCL22 | Chemokine (C-C motif) ligand 22 | −4.08 |
| NM_001025195 | CES1 | Carboxylesterase 1 | −4.22 |
| NM_001207019 | FCER2 | Fc fragment of IgE receptor II | −4.23 |
| NM_002438 | MRC1 | Mannose receptor, C type 1 (CD206) | −4.37 |
| NM_001266 | CES1 | carboxylesterase 1 | −4.38 |
| NM_000600 | IL6 | Interleukin 6 | −4.63 |
| NM_006770 | MARCO | Macrophage receptor with collagenous structure | −7.49 |
| NM_002981 | CCL1 | Chemokine (C-C motif) ligand 1 | −8.71 |
| NM_002187 | IL12B | Interleukin 12B (p40) | −15.28 |
| Activation Z-Score | ||
|---|---|---|
| Signaling Pathway | Mob-MDM(LPS/IFNγ) vs. GM-MDM(LPS/IFNγ) and M-MDM(LPS/IFNγ) | M-MDM(LPS/IFNγ) vs. GM-MDM(LPS/IFNγ) |
| Endothelin-1 Signaling | 2.65 | −0.38 |
| NF-κB Signaling | 2.65 | −0.82 |
| Estrogen-mediated S-phase Entry | 2.45 | 1.63 |
| Mitotic Roles of Polo-Like Kinase | 2.45 | 0.82 |
| Role of IL-17F in Allergic Inflammatory Airway Diseases | 2.24 | 0.45 |
| TREM1 Signaling | 2.24 | −0.45 |
| p53 Signaling | −2.65 | 1.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bazzi, S.; El-Darzi, E.; McDowell, T.; Modjtahedi, H.; Mudan, S.; Achkar, M.; Akle, C.; Kadara, H.; Bahr, G.M. Cytokine/Chemokine Release Patterns and Transcriptomic Profiles of LPS/IFNγ-Activated Human Macrophages Differentiated with Heat-Killed Mycobacterium obuense, M-CSF, or GM-CSF. Int. J. Mol. Sci. 2021, 22, 7214. https://doi.org/10.3390/ijms22137214
Bazzi S, El-Darzi E, McDowell T, Modjtahedi H, Mudan S, Achkar M, Akle C, Kadara H, Bahr GM. Cytokine/Chemokine Release Patterns and Transcriptomic Profiles of LPS/IFNγ-Activated Human Macrophages Differentiated with Heat-Killed Mycobacterium obuense, M-CSF, or GM-CSF. International Journal of Molecular Sciences. 2021; 22(13):7214. https://doi.org/10.3390/ijms22137214
Chicago/Turabian StyleBazzi, Samer, Emale El-Darzi, Tina McDowell, Helmout Modjtahedi, Satvinder Mudan, Marcel Achkar, Charles Akle, Humam Kadara, and Georges M. Bahr. 2021. "Cytokine/Chemokine Release Patterns and Transcriptomic Profiles of LPS/IFNγ-Activated Human Macrophages Differentiated with Heat-Killed Mycobacterium obuense, M-CSF, or GM-CSF" International Journal of Molecular Sciences 22, no. 13: 7214. https://doi.org/10.3390/ijms22137214
APA StyleBazzi, S., El-Darzi, E., McDowell, T., Modjtahedi, H., Mudan, S., Achkar, M., Akle, C., Kadara, H., & Bahr, G. M. (2021). Cytokine/Chemokine Release Patterns and Transcriptomic Profiles of LPS/IFNγ-Activated Human Macrophages Differentiated with Heat-Killed Mycobacterium obuense, M-CSF, or GM-CSF. International Journal of Molecular Sciences, 22(13), 7214. https://doi.org/10.3390/ijms22137214