
Single Copy Oligonucleotide Fluorescence In Situ Hybridization Probe Design Platforms: Development, Application and Evaluation


Abstract
1. Introduction
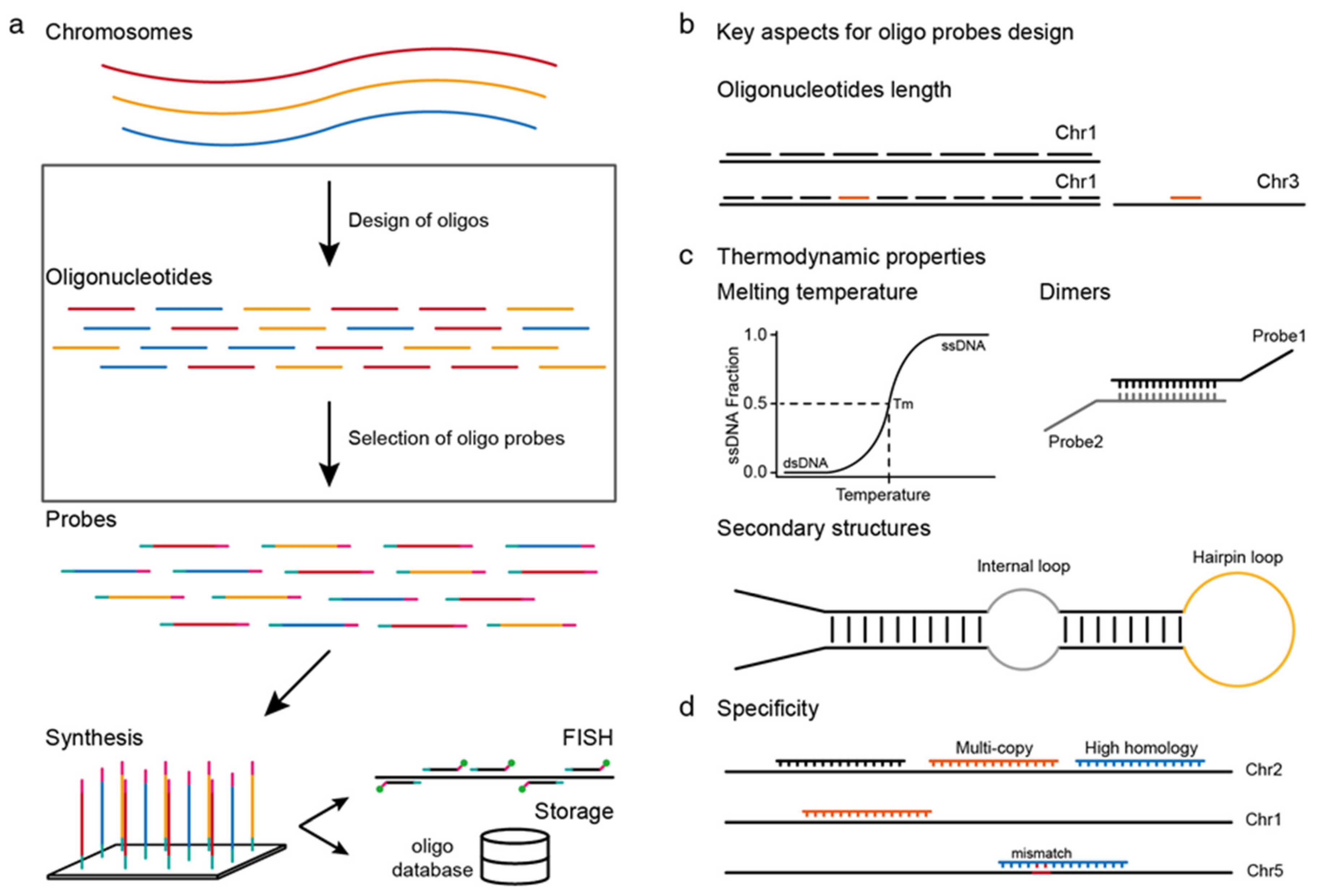
2. Key Aspects of Oligo-FISH Probe Design
2.1. Oligonucleotide Length
2.2. Thermodynamic Properties
2.3. Specificity
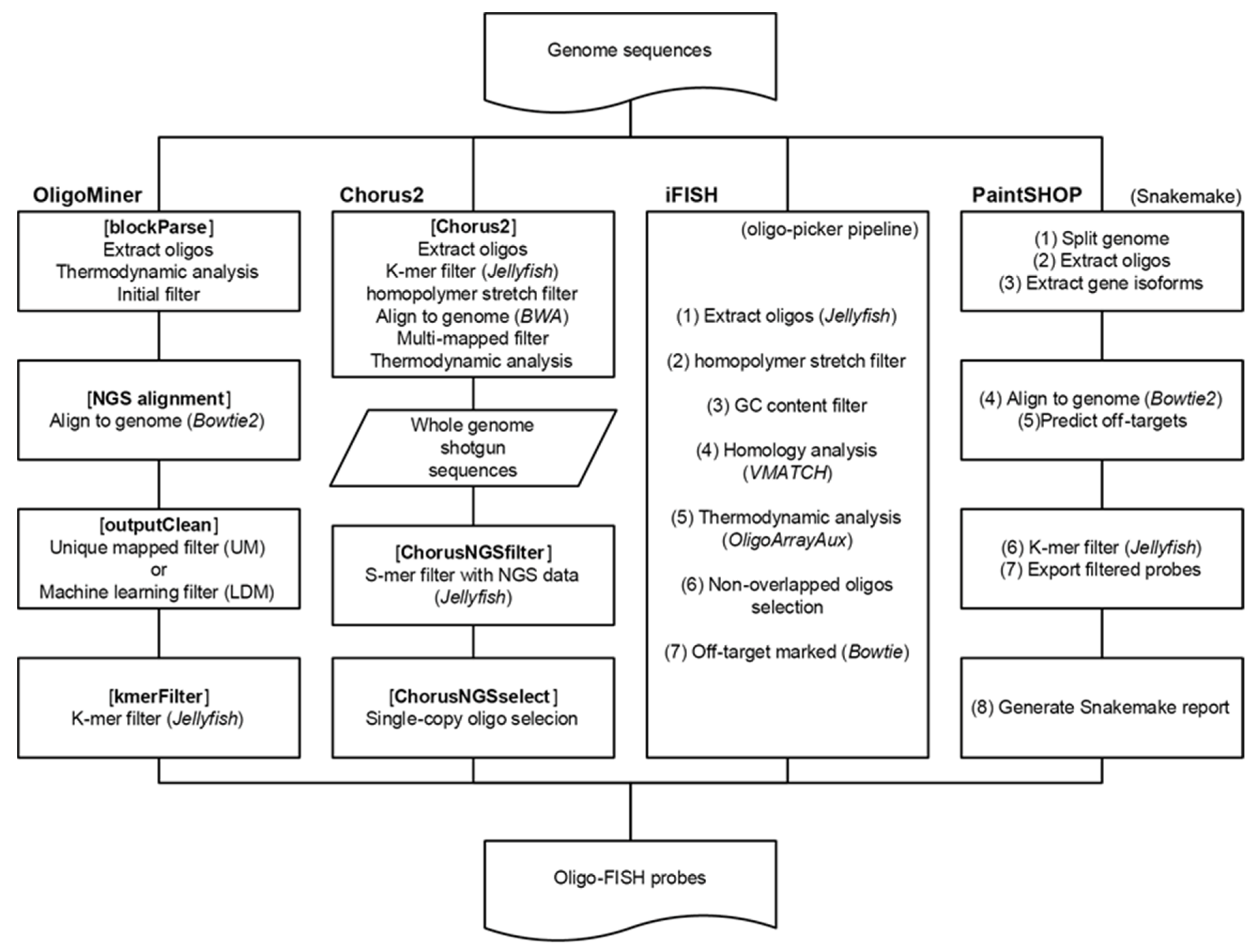
3. Development and Application of Oligo-FISH Probe Design Tools
4. Evaluation of State-of-Art Tools for Oligo-FISH Probe Design
4.1. OligoMiner
4.2. iFISH
4.3. Chorus2
4.4. PaintSHOP
4.5. Different Pipelines of Oligo Probe Design among Different Tools
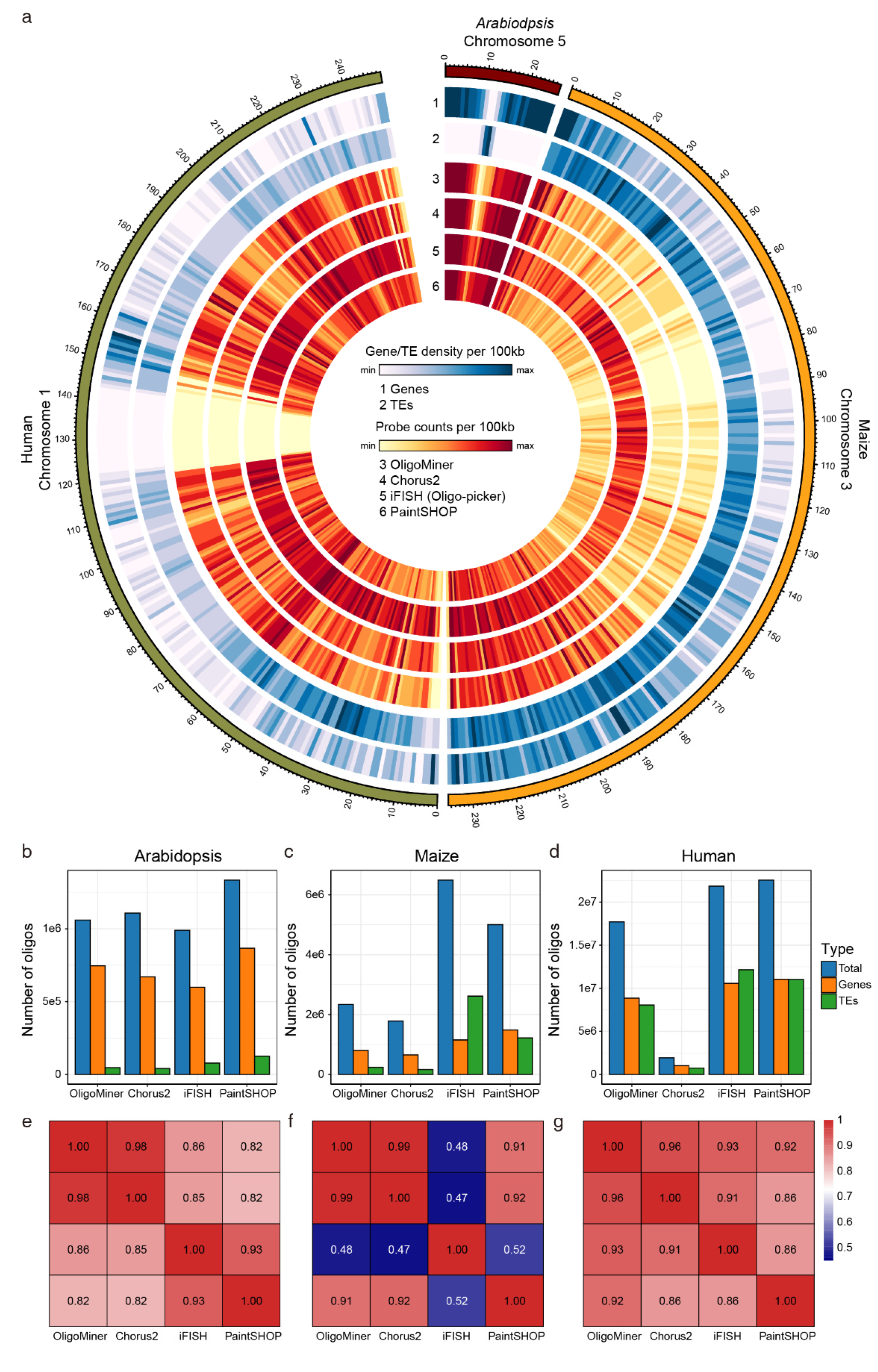
4.6. Performance of Probe Design Pipelines among Different Tools
5. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bauman, J.G.; Wiegant, J.; Borst, P.; van Duijn, P. A new method for fluorescence microscopical localization of specific DNA sequences by in situ hybridization of fluorochrome-labelled RNA. Exp. Cell Res. 1980, 128, 485–490. [Google Scholar] [CrossRef]
- Pardue, M.L.; Gall, J.G. Molecular hybridization of radioactive DNA to the DNA of cytological preparations. Proc. Natl. Acad. Sci. USA 1969, 64, 600–604. [Google Scholar] [CrossRef]
- Rudkin, G.T.; Stollar, B.D. High resolution detection of DNA–RNA hybrids in situ by indirect immunofluorescence. Nature 1977, 265, 472–473. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J. Fluorescence in situ hybridization in plants: Recent developments and future applications. Chromosome Res. 2019, 27, 153–165. [Google Scholar] [CrossRef]
- Jiang, J.; Gill, B.S. Current status and the future of fluorescence in situ hybridization (FISH) in plant genome research. Genome 2006, 49, 1057–1068. [Google Scholar] [CrossRef]
- Braz, G.T.; do Vale Martins, L.; Zhang, T.; Albert, P.S.; Birchler, J.A.; Jiang, J. A universal chromosome identification system for maize and wild Zea species. Chromosome Res. 2020, 28, 183–194. [Google Scholar] [CrossRef]
- do Vale Martins, L.; Yu, F.; Zhao, H.; Dennison, T.; Lauter, N.; Wang, H.; Deng, Z.; Thompson, A.; Semrau, K.; Rouillard, J.M.; et al. Meiotic crossovers characterized by haplotype-specific chromosome painting in maize. Nat. Commun. 2019, 10, 4604. [Google Scholar] [CrossRef]
- Han, Y.; Zhang, T.; Thammapichai, P.; Weng, Y.; Jiang, J. Chromosome-Specific Painting in Cucumis Species Using Bulked Oligonucleotides. Genetics 2015, 200, 771–779. [Google Scholar] [CrossRef]
- Badaeva, E.D.; Friebe, B.; Gill, B.S. Genome differentiation in Aegilops. 2. Physical mapping of 5S and 18S–26S ribosomal RNA gene families in diploid species. Genome 1996, 39, 1150–1158. [Google Scholar] [CrossRef] [PubMed]
- Fukui, K.; Ohmido, N.; Khush, G.S. Variability in rDNA loci in the genus Oryza detected through fluorescence in situ hybridization. Theor. Appl. Genet. 1994, 87, 893–899. [Google Scholar] [CrossRef]
- Kato, A.; Lamb, J.C.; Birchler, J.A. Chromosome painting using repetitive DNA sequences as probes for somatic chromosome identification in maize. Proc. Natl. Acad. Sci. USA 2004, 101, 13554–13559. [Google Scholar] [CrossRef] [PubMed]
- Janda, J.; Safar, J.; Kubaláková, M.; Bartoš, J.; Kovářová, P.; Suchánková, P.; Pateyron, S.; Číhalíková, J.; Sourdille, P.; Šimková, H.; et al. Advanced resources for plant genomics: A BAC library specific for the short arm of wheat chromosome 1B. Plant J. 2006, 47, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, G.; Ogaki, Y.; Hokimoto, N.; Xiao, L.; Kikuchi-Taura, A.; Harada, C.; Okayama, R.; Tsuru, A.; Onishi, M.; Saito, N.; et al. Random BAC FISH of monocot plants reveals differential distribution of repetitive DNA elements in small and large chromosome species. Plant Cell Rep. 2012, 31, 621–628. [Google Scholar] [CrossRef]
- Cuadrado, A.; Golczyk, H.; Jouve, N. A novel, simple and rapid nondenaturing FISH (ND-FISH) technique for the detection of plant telomeres. Potential used and possible target structures detected. Chromosome Res. 2009, 17, 755–762. [Google Scholar] [CrossRef]
- Cuadrado, A.; Jouve, N. Chromosomal detection of simple sequence repeats (SSRs) using nondenaturing FISH (ND-FISH). Chromosoma 2010, 119, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.L.; Chen, L.; Wang, Y.Y.; Li, M.; Yang, Z.J.; Qiu, L.; Yan, B.J.; Ren, Z.L.; Tang, Z.X. Oligonucleotide Probes for ND-FISH Analysis to Identify Rye and Wheat Chromosomes. Sci. Rep. 2015, 5, 10552. [Google Scholar] [CrossRef]
- Jiang, W.X.; Jiang, C.Z.; Yuan, W.G.; Zhang, M.J.; Fang, Z.J.; Li, Y.; Li, G.R.; Jia, J.Q.; Yang, Z.J. A universal karyotypic system for hexaploid and diploid Avena species brings oat cytogenetics into the genomics era. BMC Plant Biol. 2021, 21, 213. [Google Scholar] [CrossRef] [PubMed]
- Braz, G.T.; He, L.; Zhao, H.; Zhang, T.; Semrau, K.; Rouillard, J.M.; Torres, G.A.; Jiang, J. Comparative Oligo-FISH Mapping: An Efficient and Powerful Methodology To Reveal Karyotypic and Chromosomal Evolution. Genetics 2018, 208, 513–523. [Google Scholar] [CrossRef]
- Nir, G.; Farabella, I.; Perez Estrada, C.; Ebeling, C.G.; Beliveau, B.J.; Sasaki, H.M.; Lee, S.D.; Nguyen, S.C.; McCole, R.B.; Chattoraj, S.; et al. Walking along chromosomes with super-resolution imaging, contact maps, and integrative modeling. PLoS Genet. 2018, 14, e1007872. [Google Scholar] [CrossRef]
- Šimoníková, D.; Němečková, A.; Karafiátová, M.; Uwimana, B.; Swennen, R.; Doležel, J.; Hřibová, E. Chromosome Painting Facilitates Anchoring Reference Genome Sequence to Chromosomes In Situ and Integrated Karyotyping in Banana (Musa spp.). Front. Plant Sci. 2019, 10, 1503. [Google Scholar] [CrossRef]
- Beliveau, B.J.; Kishi, J.Y.; Nir, G.; Sasaki, H.M.; Saka, S.K.; Nguyen, S.C.; Wu, C.T.; Yin, P. OligoMiner provides a rapid, flexible environment for the design of genome-scale oligonucleotide in situ hybridization probes. Proc. Natl. Acad. Sci. USA 2018, 115, E2183–E2192. [Google Scholar] [CrossRef]
- Gelali, E.; Girelli, G.; Matsumoto, M.; Wernersson, E.; Custodio, J.; Mota, A.; Schweitzer, M.; Ferenc, K.; Li, X.; Mirzazadeh, R.; et al. iFISH is a publically available resource enabling versatile DNA FISH to study genome architecture. Nat. Commun. 2019, 10, 1636. [Google Scholar] [CrossRef]
- Yilmaz, L.S.; Parnerkar, S.; Noguera, D.R. mathFISH, a Web Tool That Uses Thermodynamics-Based Mathematical Models for In Silico Evaluation of Oligonucleotide Probes for Fluorescence In Situ Hybridization. Appl. Environ. Microbiol. 2011, 77, 1118–1122. [Google Scholar] [CrossRef]
- Dieffenbach, C.W.; Lowe, T.M.; Dveksler, G.S. General concepts for PCR primer design. Genome Res. 1993, 3, S30–S37. [Google Scholar] [CrossRef] [PubMed]
- Chavali, S.; Mahajan, A.; Tabassum, R.; Maiti, S.; Bharadwaj, D. Oligonucleotide properties determination and primer designing: A critical examination of predictions. Bioinformatics 2005, 21, 3918–3925. [Google Scholar] [CrossRef] [PubMed]
- SantaLucia, J., Jr. A unified view of polymer, dumbbell, and oligonucleotide DNA nearest-neighbor thermodynamics. Proc. Natl. Acad. Sci. USA 1998, 95, 1460–1465. [Google Scholar] [CrossRef]
- Smith, S.; Vigilant, L.; Morin, P.A. The effects of sequence length and oligonucleotide mismatches on 5′ exonuclease assay efficiency. Nucleic Acids Res. 2002, 30, e111. [Google Scholar] [CrossRef]
- Sawh, A.N.; Mango, S.E. Multiplexed Sequential DNA FISH in Caenorhabditis elegans Embryos. STAR Protoc. 2020, 1, 100107. [Google Scholar] [CrossRef]
- Steger, G. Thermal denaturation of double-stranded nucleic acids: Prediction of temperatures critical for gradient gel electrophoresis and polymerase chain reaction. Nucleic Acids Res. 1994, 22, 2760–2768. [Google Scholar] [CrossRef] [PubMed]
- Fontenete, S.; Guimarães, N.; Wengel, J.; Azevedo, N.F. Prediction of melting temperatures in fluorescence in situ hybridization (FISH) procedures using thermodynamic models. Crit. Rev. Biotechnol. 2016, 36, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Marmur, J.; Doty, P. Determination of the base composition of deoxyribonucleic acid from its thermal denaturation temperature. J. Mol. Biol. 1962, 5, 109–118. [Google Scholar] [CrossRef]
- Wallace, R.B.; Shaffer, J.; Murphy, R.F.; Bonner, J.; Hirose, T.; Itakura, K. Hybridization of synthetic oligodeoxyribonucleotides to phi chi 174 DNA: The effect of single base pair mismatch. Nucleic Acids Res. 1979, 6, 3543–3557. [Google Scholar] [CrossRef]
- Howley, P.M.; Israel, M.A.; Law, M.F.; Martin, M.A. A rapid method for detecting and mapping homology between heterologous DNAs. Evaluation of polyomavirus genomes. J. Biol. Chem. 1979, 254, 4876–4883. [Google Scholar] [CrossRef]
- Breslauer, K.J.; Frank, R.; Blocker, H.; Marky, L.A. Predicting DNA duplex stability from the base sequence. Proc. Natl. Acad. Sci. USA 1986, 83, 3746–3750. [Google Scholar] [CrossRef]
- Freier, S.M.; Kierzek, R.; Jaeger, J.A.; Sugimoto, N.; Caruthers, M.H.; Neilson, T.; Turner, D.H. Improved free-energy parameters for predictions of RNA duplex stability. Proc. Natl. Acad. Sci. USA 1986, 83, 9373–9377. [Google Scholar] [CrossRef]
- SantaLucia, J., Jr.; Allawi, H.T.; Seneviratne, P.A. Improved Nearest-Neighbor Parameters for Predicting DNA Duplex Stability. Biochemistry 1996, 35, 3555–3562. [Google Scholar] [CrossRef]
- Sugimoto, N.; Nakano, S.; Yoneyama, M.; Honda, K. Improved Thermodynamic Parameters and Helix Initiation Factor to Predict Stability of DNA Duplexes. Nucleic Acids Res. 1996, 24, 4501–4505. [Google Scholar] [CrossRef]
- Xia, T.; SantaLucia, J., Jr.; Burkard, M.E.; Kierzek, R.; Schroeder, S.J.; Jiao, X.; Cox, C.; Turner, D.H. Thermodynamic parameters for an expanded nearest-neighbor model for formation of RNA duplexes with Watson-Crick base pairs. Biochemistry 1998, 37, 14719–14735. [Google Scholar] [CrossRef]
- SantaLucia, J., Jr.; Hicks, D. The Thermodynamics of DNA Structural Motifs. Annu. Rev. Biophys. Biomol. Struct. 2004, 33, 415–440. [Google Scholar] [CrossRef]
- Stull, R.A.; Taylor, L.A.; Szoka, F.C., Jr. Predicting antisense oligonucleotide inhibitory efficacy: A computational approach using histograms and thermodynamic indices. Nucleic Acids Res. 1992, 20, 3501–3508. [Google Scholar] [CrossRef]
- Nazarenko, I.; Pires, R.; Lowe, B.; Obaidy, M.; Rashtchian, A. Effect of primary and secondary structure of oligodeoxyribonucleotides on the fluorescent properties of conjugated dyes. Nucleic Acids Res. 2002, 30, 2089–2195. [Google Scholar] [CrossRef]
- Allawi, H.T.; SantaLucia, J., Jr. Thermodynamics and NMR of Internal G·T Mismatches in DNA. Biochemistry 1997, 36, 10581–10594. [Google Scholar] [CrossRef] [PubMed]
- Peyret, N.; Seneviratne, P.A.; Allawi, H.T.; SantaLucia, J., Jr. Nearest-Neighbor Thermodynamics and NMR of DNA Sequences with Internal A·A, C·C, G·G, and T·T Mismatches. Biochemistry 1999, 38, 3468–3477. [Google Scholar] [CrossRef] [PubMed]
- Bommarito, S.; Peyret, N.; SantaLucia, J., Jr. Thermodynamic parameters for DNA sequences with dangling ends. Nucleic Acids Res. 2000, 28, 1929–1934. [Google Scholar] [CrossRef] [PubMed]
- SantaLucia, J., Jr. Physical Principles and Visual-OMP Software for Optimal PCR Design. Methods Mol. Biol. 2007, 402, 3–33. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Novák, P.; Neumann, P.; Pech, J.; Steinhaisl, J.; Macas, J. RepeatExplorer: A Galaxy-based web server for genome-wide characterization of eukaryotic repetitive elements from next-generation sequence reads. Bioinformatics 2013, 29, 792–793. [Google Scholar] [CrossRef]
- Beier, S.; Ulpinnis, C.; Schwalbe, M.; Münch, T.; Hoffie, R.; Koeppel, I.; Hertig, C.; Budhagatapalli, N.; Hiekel, S.; Pathi, K.M.; et al. Kmasker plants—A tool for assessing complex sequence space in plant species. Plant J. 2020, 102, 631–642. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, G.; Zhao, H.; Braz, G.T.; Jiang, J. Chorus2: Design of genome-scale oligonucleotide-based probes for fluorescence in situ hybridization. Plant Biotechnol. J. 2021. [Google Scholar] [CrossRef] [PubMed]
- Frickmann, H.; Zautner, A.E.; Moter, A.; Kikhney, J.; Hagen, R.M.; Stender, H.; Poppert, S. Fluorescence in situ hybridization (FISH) in the microbiological diagnostic routine laboratory: A review. Crit. Rev. Microbiol. 2017, 43, 263–293. [Google Scholar] [CrossRef] [PubMed]
- Prudent, E.; Raoult, D. Fluorescence in situ hybridization, a complementary molecular tool for the clinical diagnosis of infectious diseases by intracellular and fastidious bacteria. FEMS Microbiol. Rev. 2019, 43, 88–107. [Google Scholar] [CrossRef]
- Rouillard, J.M.; Zuker, M.; Gulari, E. OligoArray 2.0: Design of oligonucleotide probes for DNA microarrays using a thermodynamic approach. Nucleic Acids Res. 2003, 31, 3057–3062. [Google Scholar] [CrossRef]
- Navin, N.; Grubor, V.; Hicks, J.; Leibu, E.; Thomas, E.; Troge, J.; Riggs, M.; Lundin, P.; Månér, S.; Sebat, J.; et al. PROBER: Oligonucleotide FISH probe design software. Bioinformatics 2006, 22, 2437–2438. [Google Scholar] [CrossRef] [PubMed]
- Nedbal, J.; Hobson, P.S.; Fear, D.J.; Heintzmann, R.; Gould, H.J. Comprehensive FISH Probe Design Tool Applied to Imaging Human Immunoglobulin Class Switch Recombination. PLoS ONE 2012, 7, e51675. [Google Scholar] [CrossRef]
- Hendling, M.; Pabinger, S.; Peters, K.; Wolff, N.; Conzemius, R.; Barišić, I. Oli2go: An automated multiplex oligonucleotide design tool. Nucleic Acids Res. 2018, 46, W252–W256. [Google Scholar] [CrossRef]
- Passaro, M.; Martinovic, M.; Bevilacqua, V.; Hershberg, E.A.; Rossetti, G.; Beliveau, B.J.; Bonnal, R.J.P.; Pagani, M. OligoMinerApp: A web-server application for the design of genome-scale oligonucleotide in situ hybridization probes through the flexible OligoMiner environment. Nucleic Acids Res. 2020, 48, W332–W339. [Google Scholar] [CrossRef]
- Hu, M.; Yang, B.; Cheng, Y.; Radda, J.S.D.; Chen, Y.; Liu, M.; Wang, S. ProbeDealer is a convenient tool for designing probes for highly multiplexed fluorescence in situ hybridization. Sci. Rep. 2020, 10, 22031. [Google Scholar] [CrossRef]
- Hershberg, E.A.; Close, J.L.; Camplisson, C.K.; Attar, S.; Chern, R.; Liu, Y.; Akilesh, S.; Nicovich, P.R.; Beliveau, B.J. PaintSHOP enables the interactive design of transcriptomeand genome-scale oligonucleotide FISH experiments. bioRxiv 2021. [Google Scholar] [CrossRef]
- Rouillard, J.M.; Herbert, C.J.; Zuker, M. OligoArray: Genome-scale oligonucleotide design for microarrays. Bioinformatics 2002, 18, 486–487. [Google Scholar] [CrossRef]
- Beliveau, B.J.; Joyce, E.F.; Apostolopoulos, N.; Yilmaz, F.; Fonseka, C.Y.; McCole, R.B.; Chang, Y.; Li, J.B.; Senaratne, T.N.; Williams, B.R.; et al. Versatile design and synthesis platform for visualizing genomes with Oligopaint FISH probes. Proc. Natl. Acad. Sci. USA 2012, 109, 21301–21306. [Google Scholar] [CrossRef]
- Markham, N.R.; Zuker, M. UNAFold: Software for nucleic acid folding and hybridization. Methods Mol. Biol. 2008, 453, 3–31. [Google Scholar]
- Beliveau, B.J.; Boettiger, A.N.; Avendaño, M.S.; Jungmann, R.; McCole, R.B.; Joyce, E.F.; Kim-Kiselak, C.; Bantignies, F.; Fonseka, C.Y.; Erceg, J.; et al. Single-molecule super-resolution imaging of chromosomes and in situ haplotype visualization using Oligopaint FISH probes. Nat. Commun. 2015, 6, 7147. [Google Scholar] [CrossRef]
- Healy, J.; Thomas, E.E.; Schwartz, J.T.; Wigler, M. Annotating Large Genomes with Exact Word Matches. Genome Res. 2003, 13, 2306–2315. [Google Scholar] [CrossRef]
- Yamada, N.A.; Rector, L.S.; Tsang, P.; Carr, E.; Scheffer, A.; Sederberg, M.C.; Aston, M.E.; Ach, R.A.; Tsalenko, A.; Sampas, N.; et al. Visualization of Fine-Scale Genomic Structure by Oligonucleotide-Based High-Resolution FISH. Cytogenet. Genome Res. 2011, 132, 248–254. [Google Scholar] [CrossRef]
- Bienko, M.; Crosetto, N.; Teytelman, L.; Klemm, S.; Itzkovitz, S.; Van Oudenaarden, A. A versatile genome-scale PCR-based pipeline for high-definition DNA FISH. Nat. Methods 2013, 10, 122–124. [Google Scholar] [CrossRef]
- Dorman, S.N.; Shirley, B.C.; Knoll, J.H.; Rogan, P.K. Expanding probe repertoire and improving reproducibility in human genomic hybridization. Nucleic Acids Res. 2013, 41, e81. [Google Scholar] [CrossRef][Green Version]
- Kent, W.J. BLAT--the BLAST-like alignment tool. Genome Res. 2002, 12, 656–664. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Hoang, P.T.N.; Rouillard, J.M.; Macas, J.; Kubalová, I.; Schubert, V.; Schubert, I. Limitation of current probe design for oligo-cross-FISH, exemplified by chromosome evolution studies in duckweeds. Chromosoma 2021, 130, 15–25. [Google Scholar] [CrossRef]
- Xin, H.; Zhang, T.; Han, Y.; Wu, Y.; Shi, J.; Xi, M.; Jiang, J. Chromosome painting and comparative physical mapping of the sex chromosomes in Populus tomentosa and Populus deltoides. Chromosoma 2018, 127, 313–321. [Google Scholar] [CrossRef]
- Albert, P.S.; Zhang, T.; Semrau, K.; Rouillard, J.M.; Kao, Y.H.; Wang, C.R.; Danilova, T.V.; Jiang, J.; Birchler, J.A. Whole-chromosome paints in maize reveal rearrangements, nuclear domains, and chromosomal relationships. Proc. Natl. Acad. Sci. USA 2019, 116, 1679–1685. [Google Scholar] [CrossRef]
- Liu, X.Y.; Sun, S.; Wu, Y.; Zhou, Y.; Gu, S.W.; Yu, H.X.; Yi, C.D.; Gu, M.H.; Jiang, J.M.; Liu, B.; et al. Dual-color oligo-FISH can reveal chromosomal variations and evolution in Oryza species. Plant J. 2020, 101, 112–121. [Google Scholar] [CrossRef]
- Li, G.; Zhang, T.; Yu, Z.; Wang, H.; Yang, E.; Yang, Z. An efficient Oligo-FISH painting system for revealing chromosome rearrangements and polyploidization in Triticeae. Plant J. 2021, 105, 978–993. [Google Scholar] [CrossRef]
- Wu, D.; Ruban, A.; Fuchs, J.; Macas, J.; Novák, P.; Vaio, M.; Zhou, Y.; Houben, A. Nondisjunction and unequal spindle organization accompany the drive of Aegilops speltoides B chromosomes. New Phytol. 2019, 223, 1340–1352. [Google Scholar] [CrossRef]
- Marçais, G.; Kingsford, C. A fast, lock-free approach for efficient parallel counting of occurrences of k-mers. Bioinformatics 2011, 27, 764–770. [Google Scholar] [CrossRef]
- Grüning, B.; Dale, R.; Sjödin, A.; Chapman, B.A.; Rowe, J.; Tomkins-Tinch, C.H.; Valieris, R.; Köster, J.; The Bioconda Team. Bioconda: Sustainable and comprehensive software distribution for the life sciences. Nat. Methods 2018, 15, 475–476. [Google Scholar] [CrossRef]
- Koster, J.; Rahmann, S. Snakemake-a scalable bioinformatics workflow engine. Bioinformatics 2018, 34, 3600. [Google Scholar] [CrossRef]
- Boettiger, A.; Murphy, S. Advances in Chromatin Imaging at Kilobase-Scale Resolution. Trends Genet. 2020, 36, 273–287. [Google Scholar] [CrossRef]
- Weng, M.L.; Becker, C.; Hildebrandt, J.; Neumann, M.; Rutter, M.T.; Shaw, R.G.; Weigel, D.; Fenster, C.B. Fine-grained analysis of spontaneous mutation spectrum and frequency in Arabidopsis thaliana. Genetics 2019, 211, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Peluso, P.; Shi, J.; Liang, T.; Stitzer, M.C.; Wang, B.; Campbell, M.S.; Stein, J.C.; Wei, X.; Chin, C.S.; et al. Improved maize reference genome with single-MOLECULE technologies. Nature 2017, 546, 524–527. [Google Scholar] [CrossRef] [PubMed]
- Genomes Project, C.; Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Tools | Year | Platform | Language | Aligner | Features |
|---|---|---|---|---|---|
| OligoArray [54] | 2003 | Linux | Java | BLAST | Tm, secondary structure, specificity |
| PROBER [55] | 2006 | Windows, Web | C# | MerEngine | Tm, specificity, tiling oligo probes |
| mathFISH [23] | 2011 | Web | MATLAB | ClustalW | Thermodynamics, mismatch |
| webFISH [56] | 2012 | Web | MATLAB | Megablast | Specificity, homology, user-friendly |
| Chorus [8] | 2015 | Linux, MacOS | Python | BLAT | Genome-scale, specificity, homology, plants |
| Oli2go [57] | 2018 | Web | - | BLAST, BWA | Specificity, thermodynamics, user-friendly, non-human |
| OligoMiner [21] | 2018 | Linux, Windows, MacOS | Python | Bowtie2 | Genome-scale, specificity, thermodynamics, machine-learning, fast |
| iFISH [22] | 2019 | Linux, Web | Python, Perl | Vmatch, Bowtie | Genome-scale, pre-designed, selection, user-friendly, human |
| Kmasker plants [50] | 2020 | Linux, MacOS, Web | Perl, R, Python | BLAST | Specificity (WGS), plants |
| OligoMinerApp [58] | 2020 | Web | Python | Bowtie2 | Genome-scale, specificity, thermodynamics, machine-learning, user-friendly |
| ProbeDealer [59] | 2020 | Windows, MacOS | MATLAB | BLAST | Genome-scale, specificity, thermodynamics, user-friendly |
| Chorus2 [51] | 2021 | Linux, Windows, MacOS | Python | BWA | Genome-scale, specificity (WGS), homology, fast, comparative analysis, plants |
| PaintSHOP [60] | 2021 | Linux, MacOS, Web | Python, R | Bowtie2 | Genome-scale, specificity, thermodynamics, machine-learning, user-friendly |
| OligoMiner | Chorus2 | iFISH | PaintSHOP | ||
|---|---|---|---|---|---|
| Number of oligo probes | Arabidopsis | 1,059,677 | 1,107,815 | 989,167 | 1,333,798 |
| Maize | 2,339,006 | 1,780,857 | 6,489,281 | 5,003,474 | |
| Human | 17,717,778 | 1,940,978 | 21,847,197 | 22,555,306 | |
| Running time (min) | Arabidopsis | 43.5 | 44.2 | - | 25.9 |
| Maize | 1038 | 381.7 | - | Failed on local server | |
| Human | 1311 | 243.3 | - | ||
| Max memory usage (GB) | Arabidopsis | 3.4 | 9.1 | - | 12.1 |
| Maize | 28.5 | 24.4 | - | Over 64 GB | |
| Human | 25.2 | 34.8 | - | ||
| Adjustable probe length | √ | √ | √ | √ | |
| Thermodynamics analysis | √ | √ | √ | √ | |
| GC content selection | √ | × | √ | √ | |
| Control distance between probes | √ | √ | √ | × | |
| De novoprobe design | √ | √ | Upon request | PaintSHOP_Snakemake | |
| Further probe selection | × | ChorusPBGUI | iFISH probe design | PaintSHOP | |
| Have GUI | OligoMinerApp | ChorusGUI | iFISH4U | PaintSHOP | |
| Characteristic | Machine-learning for temperature-specific probe design | Specialized for plants, probe design for closed related species and species without reference genome | Probe selection based on target size, centrality and homogeneity | One-step design and selection of oligo probes, enable appending primers and bridge sequences | |
| Species | Genome Window | OligoMiner | Chorus2 | iFISH | PaintSHOP |
|---|---|---|---|---|---|
| Arabidopsis | 1:4,000,000-6,000,000 | 10.706/kb | 10.923/kb | 9.056/kb | 12.205/kb |
| 5:20,000,000-22,000,000 | 10.014/kb | 10.640/kb | 8.389/kb | 11.375/kb | |
| Maize | 1:230,000,000-235,000,000 | 1.666/kb | 1.293/kb | 3.378/kb | 3.082/kb |
| 3:185,000,000-190,000,000 | 1.675/kb | 1.288/kb | 3.280/kb | 3.125/kb | |
| Human | chr1:40,000,000-45,000,000 | 6.550/kb | 0.743/kb | 8.228/kb | 7.736/kb |
| chr19:10,000,000-15,000,000 | 3.652/kb | 0.519/kb | 6.845/kb | 4.874/kb |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, G.; Zhang, T. Single Copy Oligonucleotide Fluorescence In Situ Hybridization Probe Design Platforms: Development, Application and Evaluation. Int. J. Mol. Sci. 2021, 22, 7124. https://doi.org/10.3390/ijms22137124
Liu G, Zhang T. Single Copy Oligonucleotide Fluorescence In Situ Hybridization Probe Design Platforms: Development, Application and Evaluation. International Journal of Molecular Sciences. 2021; 22(13):7124. https://doi.org/10.3390/ijms22137124
Chicago/Turabian StyleLiu, Guanqing, and Tao Zhang. 2021. "Single Copy Oligonucleotide Fluorescence In Situ Hybridization Probe Design Platforms: Development, Application and Evaluation" International Journal of Molecular Sciences 22, no. 13: 7124. https://doi.org/10.3390/ijms22137124
APA StyleLiu, G., & Zhang, T. (2021). Single Copy Oligonucleotide Fluorescence In Situ Hybridization Probe Design Platforms: Development, Application and Evaluation. International Journal of Molecular Sciences, 22(13), 7124. https://doi.org/10.3390/ijms22137124