
Homology-Based Interactions between Small RNAs and Their Targets Control Dominance Hierarchy of Male Determinant Alleles of Self-Incompatibility in Arabidopsis lyrata


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
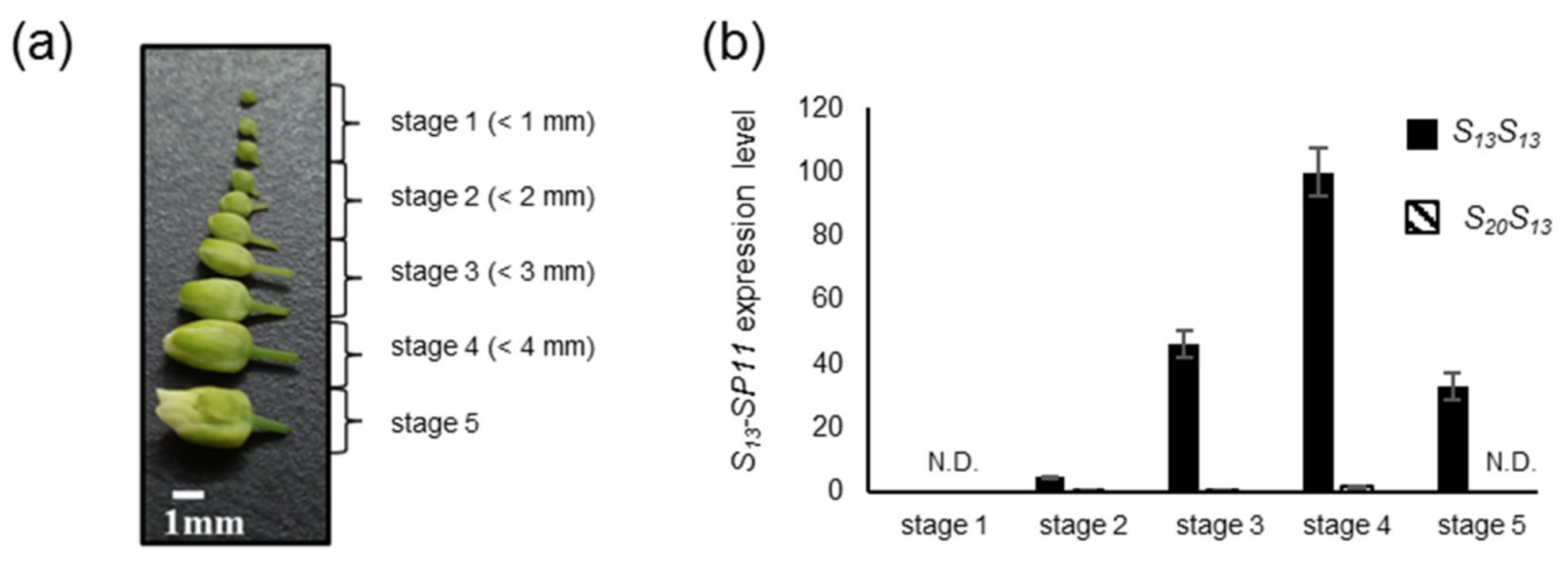
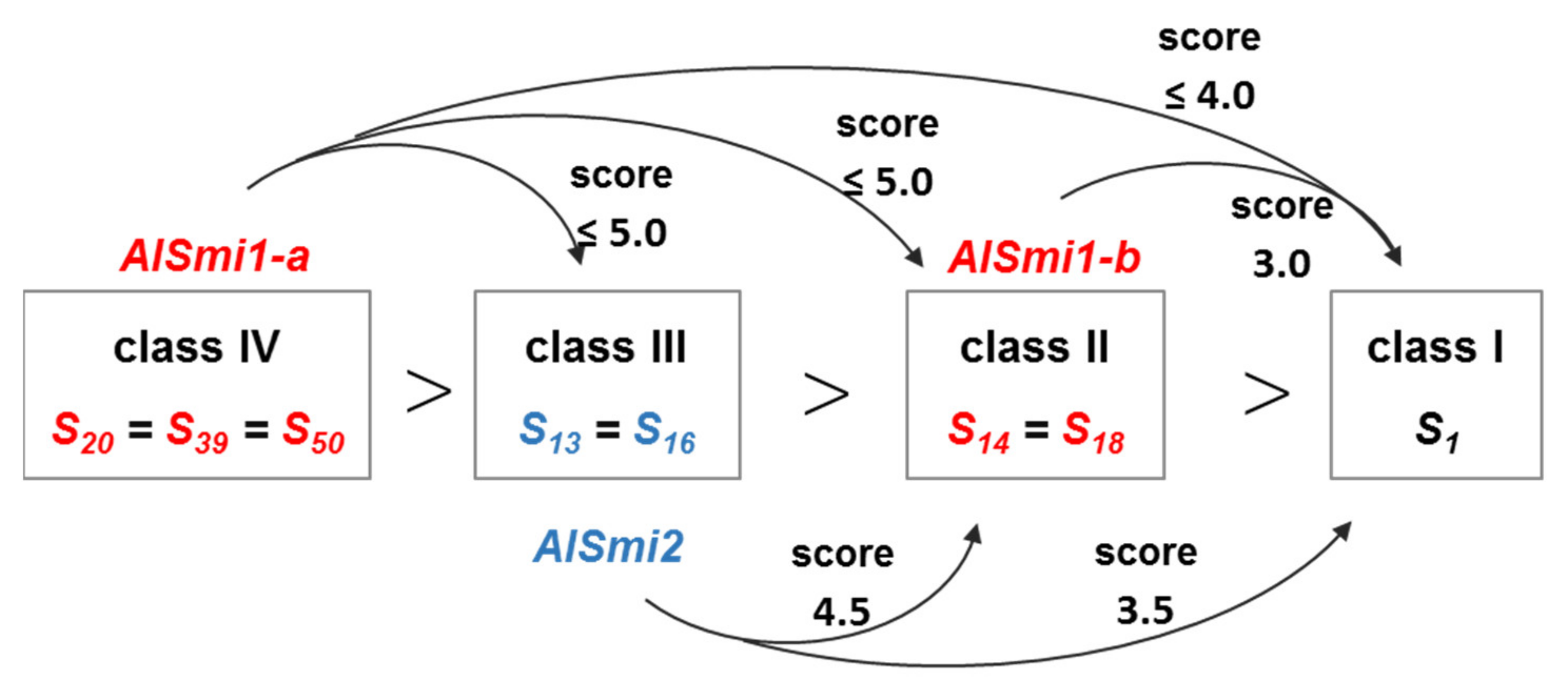
2.1. Analysis of the Dominance–Recessivity Hierarchy of the SP11 Alleles
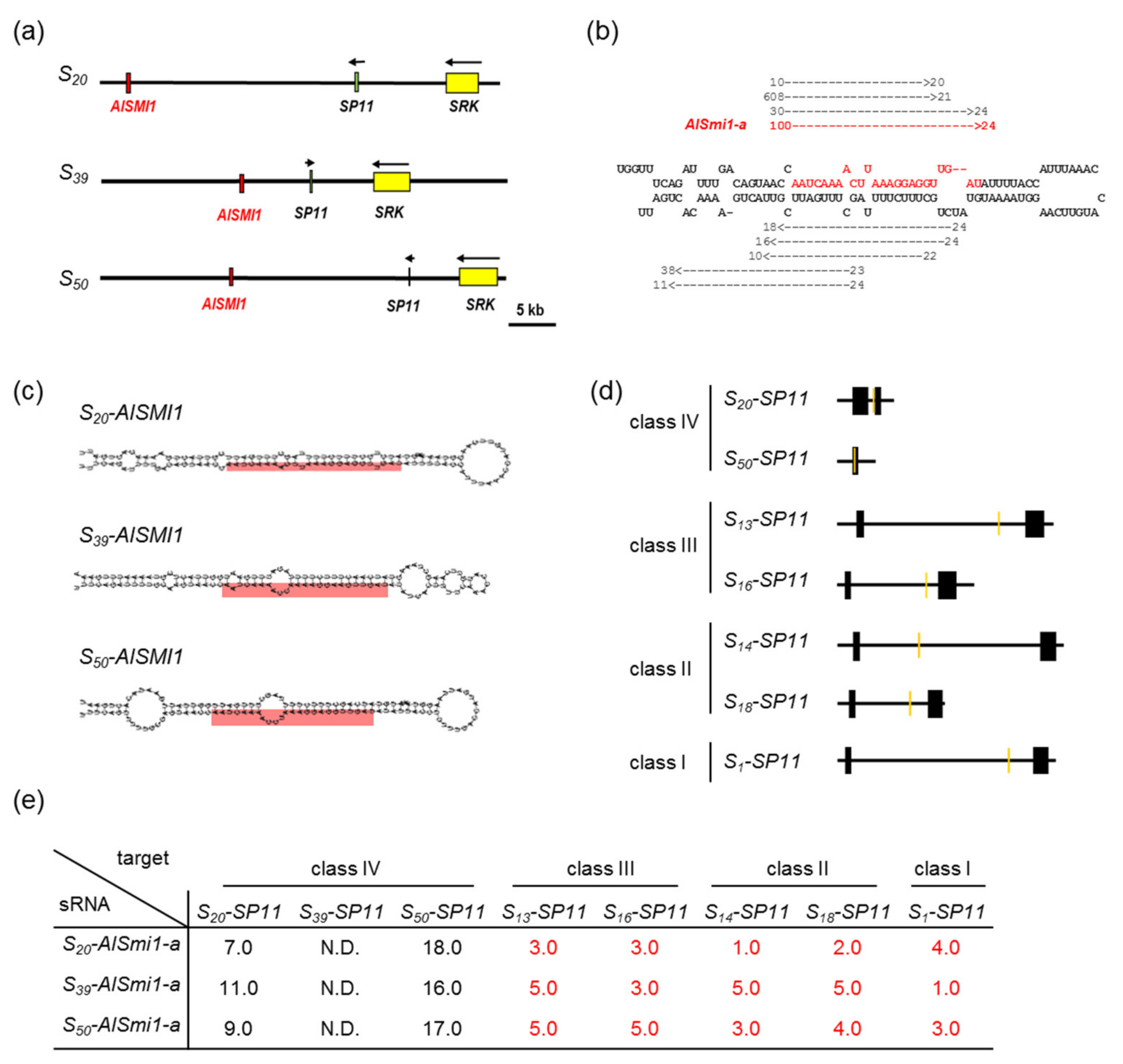
2.2. Identification of sRNA and Its Precursor Genes in Class-IV S Haplotypes
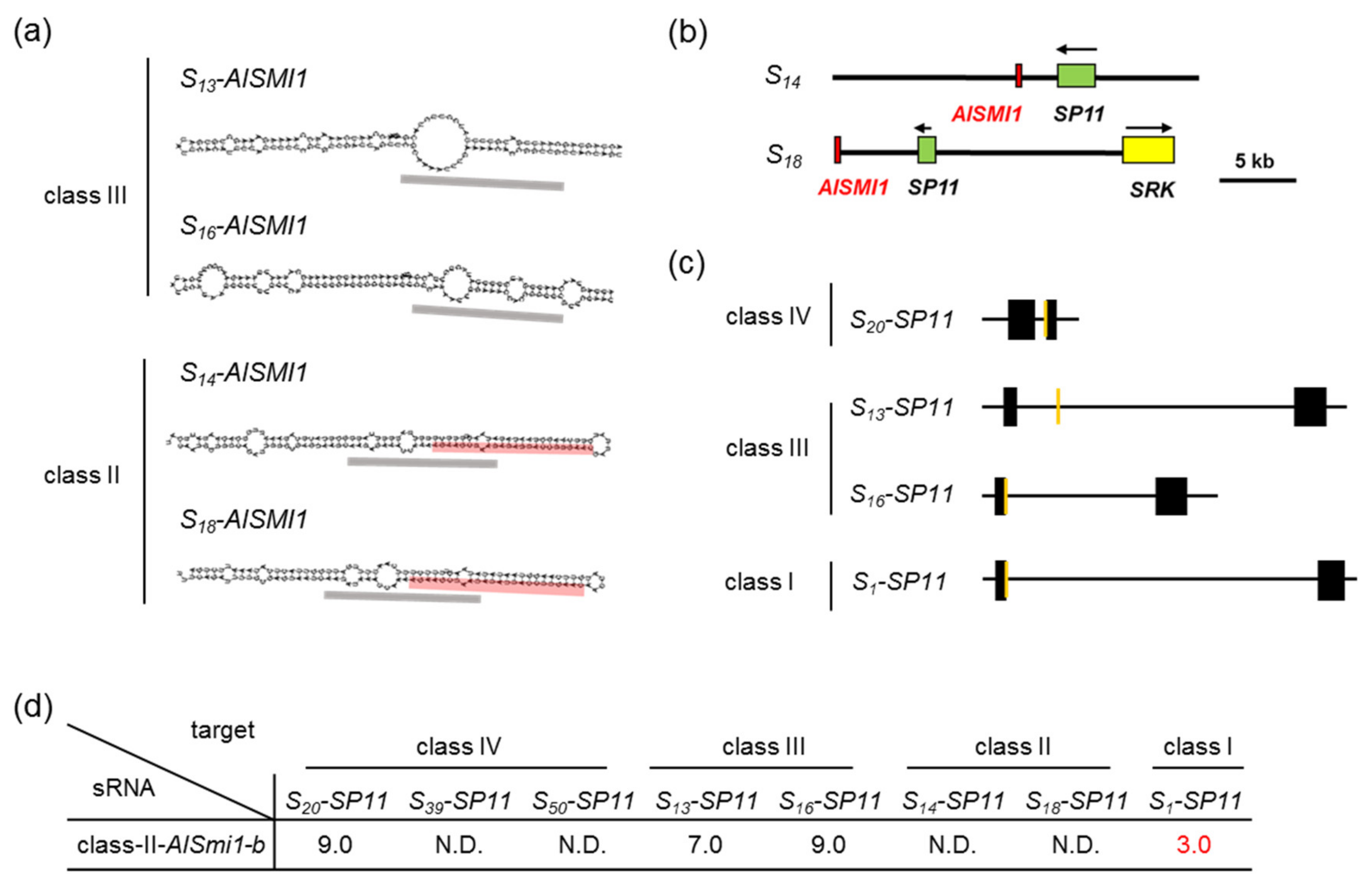
2.3. Identification of sRNA and Its Precursor Genes in Class-II S Haplotypes
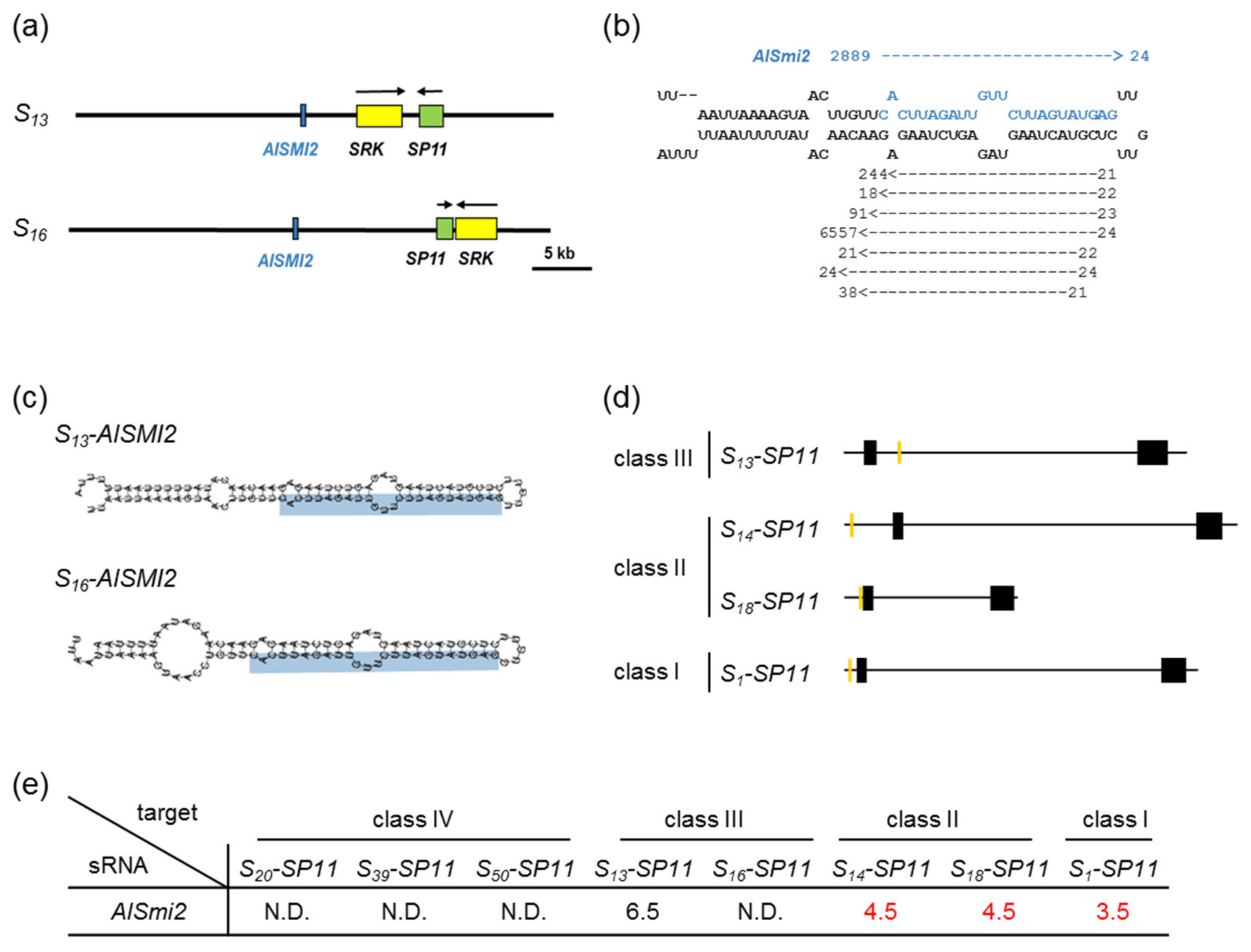
2.4. Identification of sRNA and Its Precursor Genes in Class-III S Haplotypes
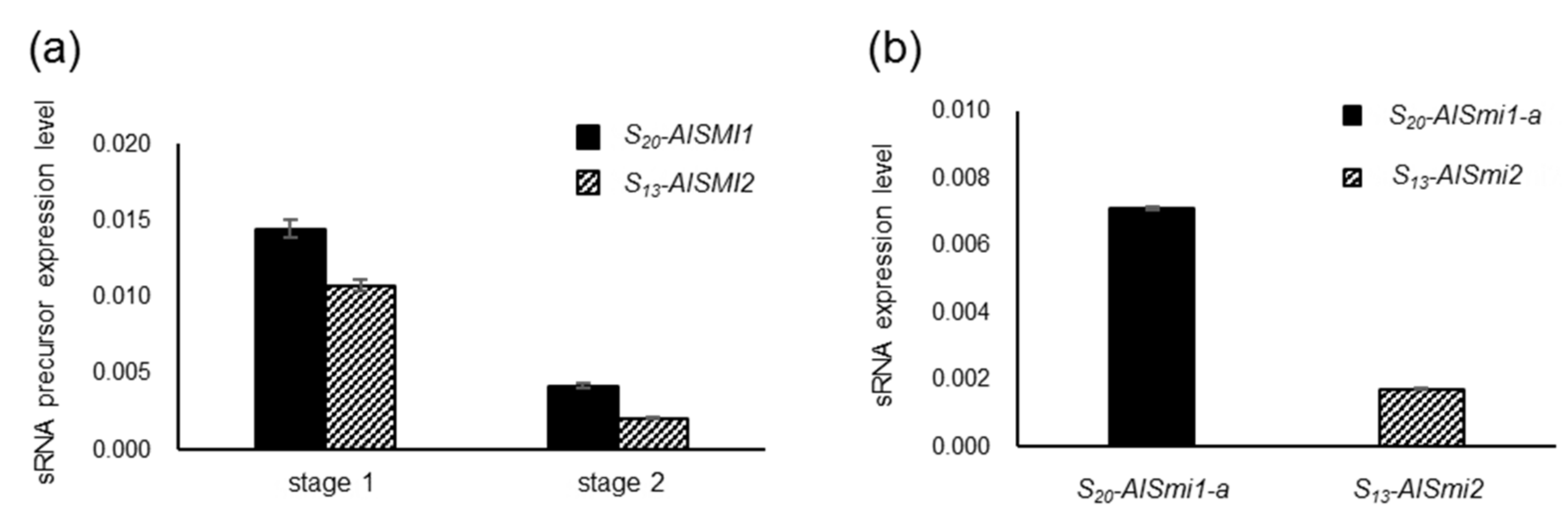
2.5. Quantification of a 24-nt sRNA and Its Precursor
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. RT-qPCR of SP11
4.3. Prediction of Inverted Repeat Sequence Regions
4.4. Small RNA Sequencing and Mapping
4.5. RT-qPCR of sRNA Precursor
4.6. Stem-Loop RT-PCR
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Suzuki, G.; Kai, N.; Hirose, T.; Fukui, K.; Nishio, T.; Takayama, S.; Isogai, A.; Watanabe, M.; Hinata, K. Genomic organization of the S locus: Identification and characterization of genes in SLG/SRK region of S9 haplotype of Brassica campestris (syn. rapa). Genetics 1999, 153, 391–400. [Google Scholar] [CrossRef]
- Schopfer, C.R.; Nasrallah, M.E.; Nasrallah, J.B. The male determinant of self-incompatibility in Brassica. Science 1999, 286, 1697–1700. [Google Scholar] [CrossRef]
- Takayama, S.; Shiba, H.; Iwano, M.; Shimosato, H.; Che, F.S.; Kai, N.; Watanabe, M.; Suzuki, G.; Hinata, K.; Isogai, A. The pollen determinant of self-incompatibility in Brassica campestris. Proc. Natl. Acad. Sci. USA 2000, 97, 1920–1925. [Google Scholar] [CrossRef]
- Stein, J.C.; Howlett, B.; Boyes, D.C.; Nasrallah, M.E.; Nasrallah, J.B. Molecular cloning of a putative receptor protein kinase gene encoded at the self-incompatibility locus of Brassica oleracea. Proc. Natl. Acad. Sci. USA 1991, 88, 8816–8820. [Google Scholar] [CrossRef]
- Takasaki, T.; Hatakeyama, K.; Suzuki, G.; Watanabe, M.; Isogai, A.; Hinata, K. The S receptor kinase determines self-incompatibility in Brassica stigma. Nature 2000, 403, 913–916. [Google Scholar] [CrossRef]
- Kachroo, A.; Schopfer, C.R.; Nasrallah, M.E.; Nasrallah, J.B. Allele-specific receptor-ligand interactions in Brassica self-incompatibility. Science 2001, 293, 1824–1826. [Google Scholar] [CrossRef]
- Takayama, S.; Shimosato, H.; Shiba, H.; Funato, M.; Che, F.S.; Watanabe, M.; Iwano, M.; Isogai, A. Direct ligand-receptor complex interaction controls Brassica self-incompatibility. Nature 2001, 413, 534–538. [Google Scholar] [CrossRef]
- Kusaba, M.; Dwyer, K.; Hendershot, J.; Vrebalov, J.; Nasrallah, J.B.; Nasrallah, M.E. Self-incompatibility in the genus Arabidopsis: Characterization of the S locus in the outcrossing A. lyrata and its autogamous relative A. thaliana. Plant Cell. 2001, 13, 627–644. [Google Scholar] [CrossRef]
- Durand, E.; Méheust, R.; Soucaze, M.; Goubet, P.M.; Gallina, S.; Poux, C.; Fobis-Loisy, I.; Guillon, E.; Gaude, T.; Sarazin, A.; et al. Dominance hierarchy arising from the evolution of a complex small RNA regulatory network. Science 2014, 346, 1200–1205. [Google Scholar] [CrossRef]
- Hatakeyama, K.; Watanabe, M.; Takasaki, T.; Ojima, K.; Hinata, K. Dominance relationships between S-alleles in self-incompatible Brassica campestris L. Heredity 1998, 80, 241–247. [Google Scholar] [CrossRef][Green Version]
- Tarutani, Y.; Shiba, H.; Iwano, M.; Kakizaki, T.; Suzuki, G.; Watanabe, M.; Isogai, A.; Takayama, S. Trans-acting small RNA determines dominance relationships in Brassica self-incompatibility. Nature 2010, 466, 983–986. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, S.; Wada, Y.; Kakizaki, T.; Tarutani, Y.; Miura-Uno, E.; Murase, K.; Fujii, S.; Hioki, T.; Shimoda, T.; Takada, Y.; et al. A complex dominance hierarchy is controlled by polymorphism of small RNAs and their targets. Nat. Plants 2016, 3, 16206. [Google Scholar] [CrossRef]
- Shiba, H.; Kakizaki, T.; Iwano, M.; Tarutani, Y.; Watanabe, M.; Isogai, A.; Takayama, S. Dominance relationships between self-incompatibility alleles controlled by DNA methylation. Nat. Genet. 2006, 38, 297–299. [Google Scholar] [CrossRef]
- Prigoda, N.L.; Nassuth, A.; Mable, B.K. Phenotypic and genotypic expression of self-incompatibility haplotypes in Arabidopsis lyrata suggests unique origin of alleles in different dominance classes. Mol. Biol. Evol. 2005, 22, 1609–1620. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.L.; Zhao, X.; Lanz, C.; Weigel, D. Evolution of the S-locus region in Arabidopsis relatives. Plant Physiol. 2011, 157, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Goubet, P.M.; Bergès, H.; Bellec, A.; Prat, E.; Helmstetter, N.; Mangenot, S.; Gallina, S.; Holl, A.C.; Fobis-Loisy, I.; Vekemans, X.; et al. Contrasted patterns of molecular evolution in dominant and recessive self-incompatibility haplotypes in Arabidopsis. PLoS Genet. 2012, 8, e1002495. [Google Scholar] [CrossRef] [PubMed]
- Schierup, M.H.; Mable, B.K.; Awadalla, P.; Charlesworth, D. Identification and characterization of a polymorphic receptor kinase gene linked to the self-incompatibility locus of Arabidopsis lyrata. Genetics 2001, 158, 387–399. [Google Scholar] [CrossRef]
- Mable, B.K.; Beland, J.; Di Berardo, C. Inheritance and dominance of self-incompatibility alleles in polyploid Arabidopsis lyrata. Heredity 2004, 93, 476–486. [Google Scholar] [CrossRef]
- Kusaba, M.; Tung, C.W.; Nasrallah, M.E.; Nasrallah, J.B. Monoallelic expression and dominance interactions in anthers of self-incompatible Arabidopsis lyrata. Plant Physiol. 2002, 128, 17–20. [Google Scholar] [CrossRef][Green Version]
- Lorenz, R.; Bernhart, S.H.; Höner zu Siederdissen, C.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. ViennaRNA package 2.0. Algorithms Mol. Biol. 2011, 6, 26–39. [Google Scholar] [CrossRef]
- Fahlgren, N.; Carrington, J.C. miRNA target prediction in plants. Methods Mol. Biol. 2010, 592, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Ridzon, D.A.; Broomer, A.J.; Zhou, Z.; Lee, D.H.; Nguyen, J.T.; Barbisin, M.; Xu, N.L.; Mahuvakar, V.R.; Andersen, M.R.; et al. Real-time quantification of microRNAs by stem–loop RT–PCR. Nucleic Acids Res. 2005, 33, e179. [Google Scholar] [CrossRef]
- Castric, V.; Bechsgaard, J.; Schierup, M.H.; Vekemans, X. Repeated Adaptive introgression at a gene under multiallelic balancing selection. PLoS Genet. 2008, 4, e1000168. [Google Scholar] [CrossRef]
- Kowyama, Y.; Takahasi, H.; Muraoka, K.; Tani, T.; Hara, K.; Shiotani, I. Number, frequency and dominance relationships of S-alleles in diploid Ipomoea trifida. Heredity 1994, 73, 275–283. [Google Scholar] [CrossRef]
- Brennan, A.C.; Tabah, D.A.; Harris, S.A.; Hiscock, S.J. Sporophytic self-incompatibility in Senecio squalidus (Asteraceae): S allele dominance interactions and modifiers of cross-compatibility and selfing rates. Heredity 2011, 106, 113–123. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Joron, M.; Papa, R.; Beltrán, M.; Chamberlain, N.; Mavárez, J.; Baxter, S.; Abanto, M.; Bermingham, E.; Humphray, S.J.; Rogers, J.; et al. A conserved supergene locus controls colour pattern diversity in Heliconius butterflies. PLoS Biol. 2006, 4, e303. [Google Scholar] [CrossRef]
- Guo, M.; Rupe, M.A.; Zinselmeier, C.; Habben, J.; Bowen, B.A.; Smith, O.S. Allelic variation of gene expression in maize hybrids. Plant Cell 2004, 16, 1707–1716. [Google Scholar] [CrossRef]
- Gimelbrant, A.; Hutchinson, J.N.; Thompson, B.R.; Chess, A. Widespread monoallelic expression on human autosomes. Science 2007, 318, 1136–1140. [Google Scholar] [CrossRef]
- Zhuang, Y.; Adams, K.L. Extensive allelic variation in gene expression in populus F1 hybrids. Genetics 2007, 177, 1987–1996. [Google Scholar] [CrossRef]
- Wang, J.; Valo, Z.; Smith, D.; Singer-Sam, J. Monoallelic expression of multiple genes in the CNS. PLoS ONE 2007, 2, e1293. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yasuda, S.; Kobayashi, R.; Ito, T.; Wada, Y.; Takayama, S. Homology-Based Interactions between Small RNAs and Their Targets Control Dominance Hierarchy of Male Determinant Alleles of Self-Incompatibility in Arabidopsis lyrata. Int. J. Mol. Sci. 2021, 22, 6990. https://doi.org/10.3390/ijms22136990
Yasuda S, Kobayashi R, Ito T, Wada Y, Takayama S. Homology-Based Interactions between Small RNAs and Their Targets Control Dominance Hierarchy of Male Determinant Alleles of Self-Incompatibility in Arabidopsis lyrata. International Journal of Molecular Sciences. 2021; 22(13):6990. https://doi.org/10.3390/ijms22136990
Chicago/Turabian StyleYasuda, Shinsuke, Risa Kobayashi, Toshiro Ito, Yuko Wada, and Seiji Takayama. 2021. "Homology-Based Interactions between Small RNAs and Their Targets Control Dominance Hierarchy of Male Determinant Alleles of Self-Incompatibility in Arabidopsis lyrata" International Journal of Molecular Sciences 22, no. 13: 6990. https://doi.org/10.3390/ijms22136990
APA StyleYasuda, S., Kobayashi, R., Ito, T., Wada, Y., & Takayama, S. (2021). Homology-Based Interactions between Small RNAs and Their Targets Control Dominance Hierarchy of Male Determinant Alleles of Self-Incompatibility in Arabidopsis lyrata. International Journal of Molecular Sciences, 22(13), 6990. https://doi.org/10.3390/ijms22136990