Abstract
Recently, much attention has been paid to the COVID-19 pandemic. Yet bacterial resistance to antibiotics remains a serious and unresolved public health problem that kills hundreds of thousands of people annually, being an insidious and silent pandemic. To contain the spreading of the SARS-CoV-2 virus, populations confined and tightened hygiene measures. We performed this study with computer simulations and by using mobility data of mobile phones from Google in the region of Lisbon, Portugal, comprising 3.7 million people during two different lockdown periods, scenarios of 40 and 60% mobility reduction. In the simulations, we assumed that the network of physical contact between people is that of a small world and computed the antibiotic resistance in human microbiomes after 180 days in the simulation. Our simulations show that reducing human contacts drives a reduction in the diversity of antibiotic resistance genes in human microbiomes. Kruskal–Wallis and Dunn’s pairwise tests show very strong evidence (p < 0.000, adjusted using the Bonferroni correction) of a difference between the four confinement regimes. The proportion of variability in the ranked dependent variable accounted for by the confinement variable was η2 = 0.148, indicating a large effect of confinement on the diversity of antibiotic resistance. We have shown that confinement and hygienic measures, in addition to reducing the spread of pathogenic bacteria in a human network, also reduce resistance and the need to use antibiotics.
1. Introduction
Antibiotic resistance is today one of the biggest threats in human and public health and has been considered a pandemic for at least two decades [1]. Antimicrobial-resistant pathogens already cause 25,000 deaths per year in Europe, 700,000 worldwide. It is estimated that it could increase to 10 million by 2050, in the most alarming scenario, if no action is taken [2]. This also implies higher treatment costs due to prolonged recovery time in biosafety facilities and the need to resort to more expensive alternative antibiotics [3].
Antibiotic misuse and over-prescription have long been documented as one of the most common errors in human health and one of the main factors that drive development of antibiotic resistance in hospitals. Misuse of antibiotics in humans can include unnecessary prescriptions or treatments that are not streamlined when microbiological culture data become available [4,5].
Hygienic measures, such as hand washing, have long been recommended to break the transmission of pathogens, especially in hospitals and health care facilities. However, recently, with the emergence of the new pandemic of COVID-19, new and stricter recommendations have been made, such as avoiding interpersonal contacts, disinfecting hands when touching physical surfaces in anthropogenic environments, and following strict rules of respiratory etiquette, quarantine, and confinement. Therefore, it is expected that, under such drastic hygienic conditions and interruption of the transmission of pathogenic microorganisms between people, antibiotic resistance in human microbiomes will also be reduced.
Human metagenomes with higher diversity of virulence genes tend to be precisely those with higher diversity of antibiotic-resistance genes [6], possibly facilitating the emergence of superbugs. However, the diversity of both gene types is dynamic over time, changing according to antibiotics usage and microbial transmission between people [7]. Bacterial transmission occurs between people that establish a network of contacts among them. What would happen to the diversity of antibiotic resistance genes if most contacts between people were abolished?
This study aims to simulate the effect of human contact reduction (person-to-person transmission in a network of contacts) on the accumulation of antibiotic resistance genes due to the sharing of bacteria belonging to the human microbiota. In the same way, we intended to find out if it is to be expected that the decrease in human contacts could, by itself, explain a decrease in antibiotic intake in humans.
This study is based on data from a densely populated region of Portugal, the Lisbon and Tagus Valley region. Portugal was one of the European countries that most quickly managed to control its first wave of the COVID-19 pandemic. Learning from the experience of other countries such as Spain or Italy, where SARS-CoV-2 caused many human losses, the Portuguese population massively followed the measures of isolation and social distancing enacted by their government to combat the pandemic.
The Lisbon and Tagus Valley region comprises a metropolitan area and the surrounding suburbs with about 3.7 million persons over 11.741 Km2 [8], with high daily traffic to and from Lisbon and an overloaded transport network. Therefore, for the development of this study, we used real data on the change in population mobility in this geographical region, immediately after the peak of the first wave, on 1 April, and extending until 30 September 2020.
2. Results
2.1. Lockdown Breaks Many Human Contacts on the Social Network
In Portugal, the first pandemic wave of COVID-19 occurred between March and August 2020. From 16 March, the Portuguese authorities decreed a state of emergency and a global lockdown. The Lisbon and Tagus Valley area in Portugal is a predominantly urban area that showed greater variations in circulation, and therefore, in human networks, before and during lockdown due to the COVID-19 pandemic.
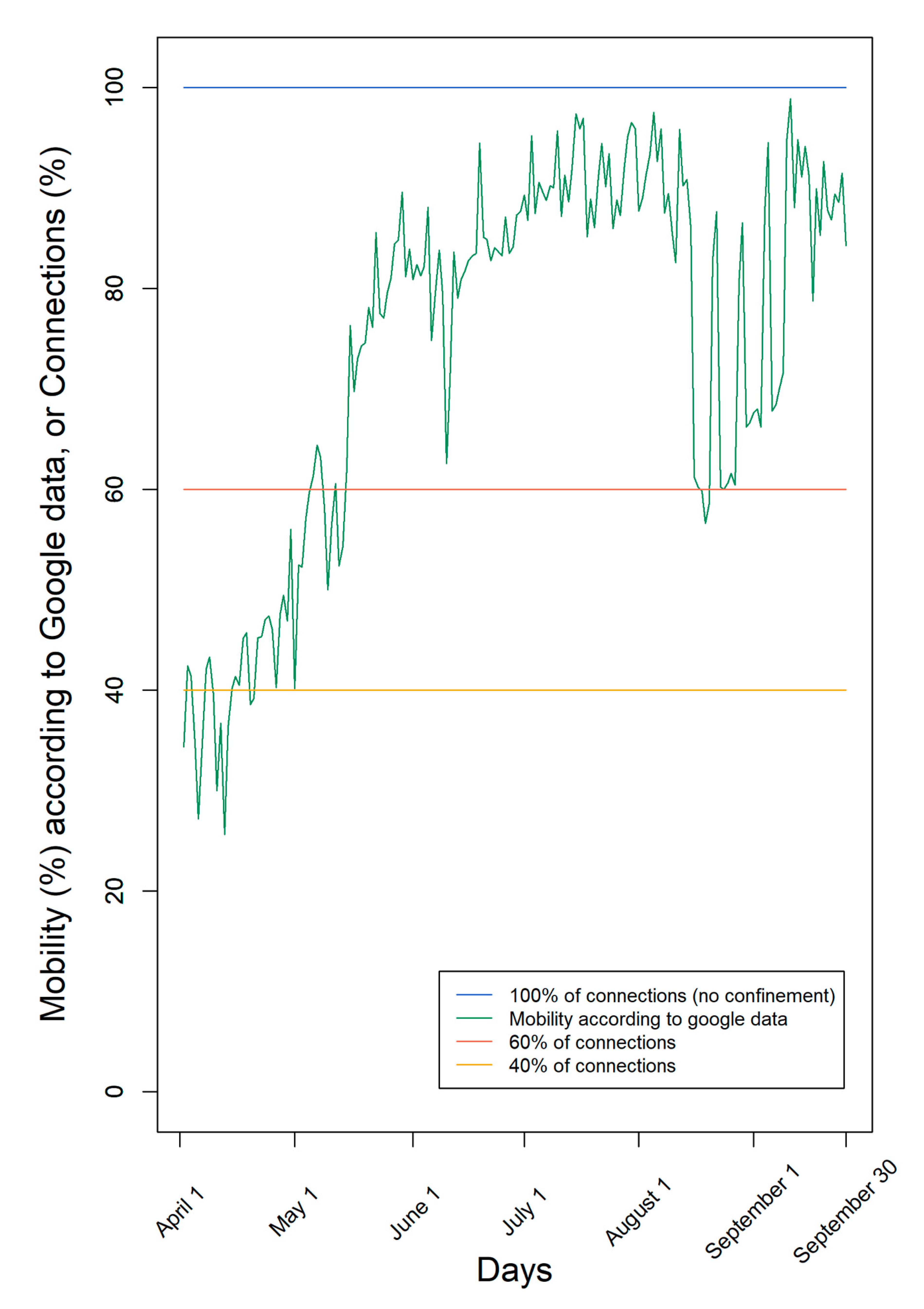
According to Google data [9], mobility was very low at the beginning of April and increased mainly since May with many oscillations (Figure 1). The lower mobility registered through Google data indicates the decrease in the number of contacts between individuals. However, it is only an estimate of the human social contacts, since it contemplates only individuals who move with connected mobile data and does not distinguish between individuals who move and contact other people from another household, from those who move alone, complying with the social distancing rules.
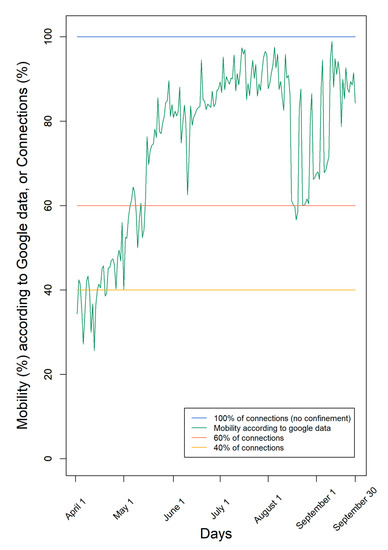
Figure 1.
Levels of confinement used in the simulations. We used either a fixed number of connections (100%, 60%, or 40%) or mobility according to Google data from the region of Lisbon and Tagus Valley in Portugal in the period 1 April–30 September.
During the lockdown, intimate human connections should remain preserved at home. However, we expect a reduction in the transmission of bacteria and pathogens between individuals in the community and, therefore, less bacterial enrichment of human microbiomes. Consequently, we can predict a reduced flow of antibiotic-resistant bacterial genes between people’s microbiomes during confinement [10].
Likewise, as a measure of the number of times an individual leaves home, mobility is not the only factor that influences the exchange of microorganisms in a social network of contacts. The use of a mask and other behaviors like social distancing also reduce the spread of pathogens, particularly in airborne disease agents.
2.2. Confinement Leads to Lower Diversity of Antibiotic Resistance Genes in Human Microbiomes
In order to test the effect of the confinement in reducing the transmission of antibiotic resistance genes (ARGs) in the human microbiomes of the population, we simulated four different scenarios: (i) no confinement, hence no connections eliminated; (ii) using the real mobility data from Google that comprises all the period of the first pandemic wave in Portugal assuming that a certain decrease of mobility corresponds to the same decrease of the number of connections; and assuming constant reductions of connection between individuals of (iii) 40% and (iv) 60%.
According to Google data, mobility fluctuated over time, ending with mobility of the individuals nearly 90%, consistent with the end of the first pandemic COVID-19 wave and summer holidays (Figure 1). Moreover, according to Google, mobility was, on average, 75% of the maximum during this period.
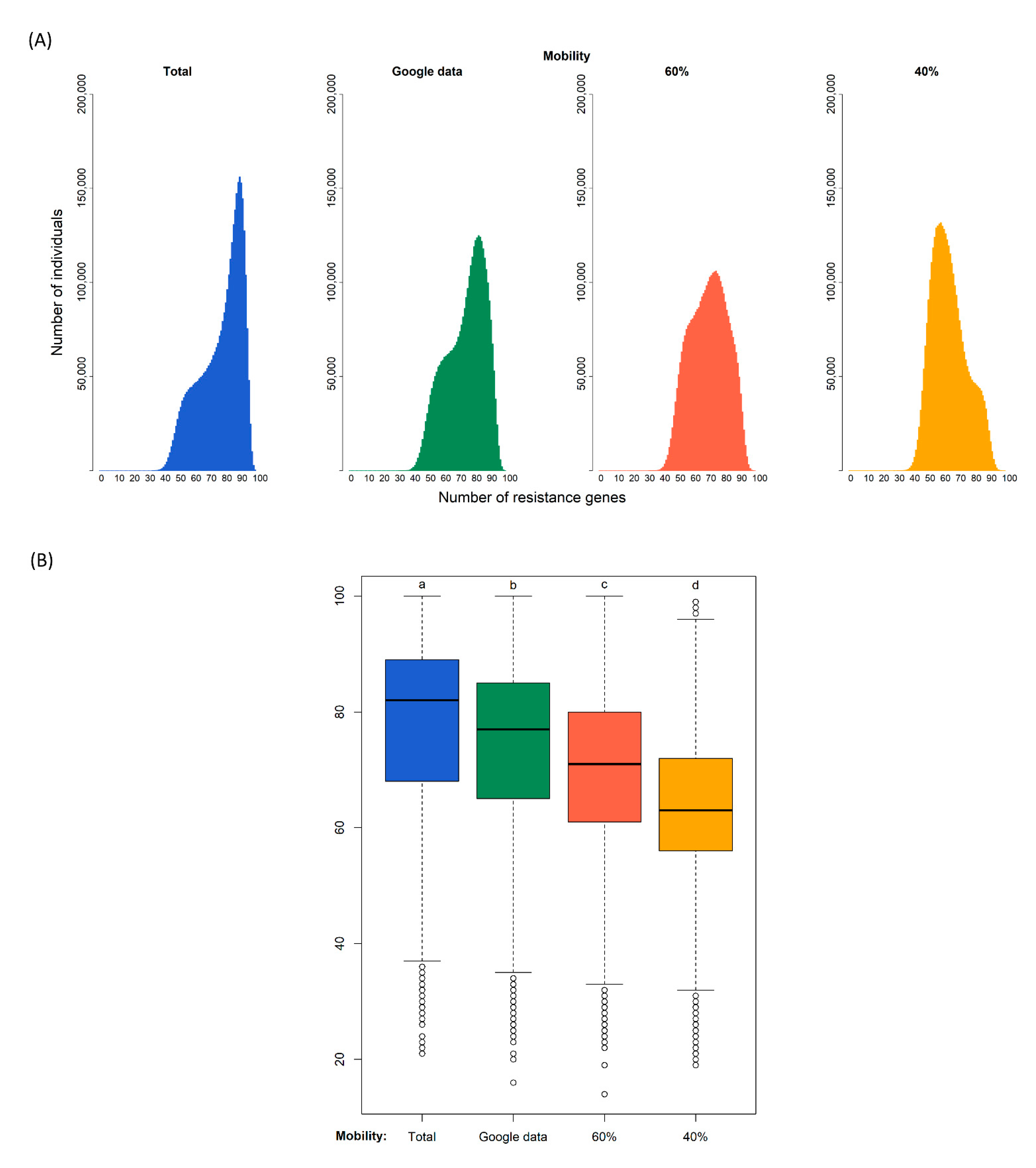
Our simulations of 180 days generated the distributions of the diversity of resistance genes in the population shown in Figure 2. The modes of the four distributions are 90 gene types for the case of no restrictions to connections, 83 assuming that google data corresponds to connections relative to the normal situation, 75 if the population only has 60% of contacts, and 59 if the population only has 40% of contacts.
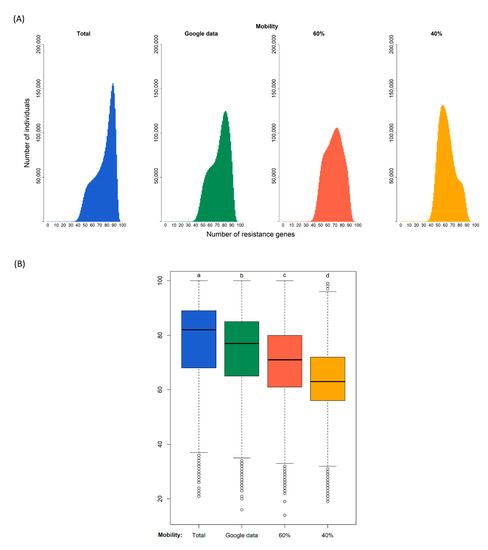
Figure 2.
Diversity of resistance genes in four different scenarios of confinement after 180 days. Simulation (i) without confinement (100% of connections); (ii) using the real mobility data from Google assuming that a certain reduction of mobility corresponds to a similar reduction of connections; and with a fixed proportion of connections of (iii) 60% and (iv) 40%. (A): Distribution of the predicted number of antibiotic resistance gene types among individuals at the end of the simulation. (B): Each box plot represents the distribution of values: the bottom and top of the box are the first and third quartiles, the horizontal line is the median, the vertical lines are the 1.5 interquartile range. Black circles represent outliers. After a Kruskal–Wallis analysis (see main text), post hoc comparisons using the Dunn test indicated that the effect of confinement is significantly different in the four confinement types as indicated by the four different letters a, b, c, and d (p-value < 0.000).
A Kruskal–Wallis test provided evidence of a difference (p < 0.000) between the mean ranks of at least one pair of groups. Dunn’s pairwise tests were carried out for the six pairs of groups. There was very strong evidence (p < 0.000, adjusted using the Bonferroni correction) of a difference between the four confinement regimes. The proportion of variability in the ranked dependent variable accounted for by the confinement variable was η2 = 0.148, indicating a large effect of confinement on the diversity of antibiotic resistance genes.
2.3. Less Contact between Humans Reduces the Need for Antibiotic Use
According to our algorithm, the use of antibiotics is triggered by exposure to bacterial pathogens during social interactions. When running the simulation, we assume that when an individual is more exposed to bacteria from other individuals, as part of a larger network of contacts, there is an increased likelihood of being infected by a pathogen and taking antibiotics. Thus, we counted the number of antibiotic administrations that occurred during the simulation. Compared with the unconfined condition, the number of antibiotic administrations fell 7, 11, and 16% in the other three conditions: Google data (assuming that mobility loss corresponds to the same connectivity loss), 60% of connections, and 40% of connections. Therefore, the daily use of antibiotics seems to follow a similar trend pattern of confinement. This trend may explain, at least partially, why, according to data from Infarmed (a Portuguese drug agency), the number of antibiotics used in Portugal in the period between January and September 2020 was lower than the corresponding months of 2019 [11]. In community pharmacies, the dispensing of these drugs decreased by 20%. However, the reduction in prescription and use of antibiotics may reflect a lower need for these drugs and a decrease in the number of medical appointments and access to primary health care due to the pandemic.
3. Discussion
Antibiotics are used as medicines to fight and eliminate bacterial infections, kill or inhibit their growth, and thus help the immune system resolve the disease [12,13,14]. In humans, antibiotic stewardship is dictated by bacterial infection, and, as such, the presence of bacterial pathogens can prompt an enrichment of human microbiomes with antibiotic resistance genes [7] whose expression enables bacteria to survive in the presence of these chemicals. Several examples in the literature on microbiome microbial dynamics are driven by human actions [15,16,17].
This work has shown that reducing contacts between people leads to a generally lower diversity of antibiotic-resistance genes among individuals’ metagenomes.
In our simulations, we assume that interpersonal contact enriches the human microbiome with genes, namely those that confer resistance to antibiotics and leverages the risk acquisition of pathogens which, in turn, triggers the taking of antibiotics. Thus, it seems easy to understand that the reduction in mobility that occurred during a period of confinement, in our simulations, led to a decrease in bacterial infections in humans. It also decreased the prescription of antibiotics and a decrease in the accumulation of antibiotic resistance genes in the microbiome. This result is concordant with the 20% drop in the prescription of antibiotics in the first three quarters of 2020 reported by the Portuguese National Health Authorities [11]. This conclusion reinforces the general recommendations to improve hygiene in disease control and antibiotic resistance in human populations.
For the parameterization of the model developed in this work, we have used data from a mixed origin: close-to-real data of the population of a geographical area, data that was estimated from the bibliography, for example, the network of contacts, or theoretical data, such as the probability of an individual taking antibiotics. However, during the development of this work, we only varied one of the parameters, namely the changes in the human contact network as an indicator of the spread of pathogens among different individuals. Thus, we consider that the breakdown of human contacts leads to a decrease in the enrichment of the human microbiome in antibiotic genes is significant and is also supported by reality.
As a conclusion, the key points of this study are: (i) The resistance of bacterial pathogens to antibiotics remains a silent and life-threatening human pandemic; (ii) Our simulations show that a reduction in human contacts decreases bacterial transmission and drives a reduction in the diversity of antibiotic resistance genes in human microbiomes; (iii) Our simulations also predict that a reduction in the spreading of pathogenic bacteria in a human network also reduces the need to use antibiotics; (iv) Hygienic measures are critical in reducing human infections and in antibiotic stewardship.
4. Methods
The model used in this article is based on our previously developed model [7].
4.1. Data
In this work, we used mobility data from Google (https://www.google.com/covid19/mobility?hl=pt-PT, accessed on 30 October 2020) [9]. This data is made available to provide statistics on what has changed in response to policies aimed at combating COVID-19. The data are divided by region and discriminated into the following categories: grocery and pharmacy, parks, transit stations, workplaces, and residential. For this study, we used data from Portugal, namely from the Lisbon and Tagus Valley region, with approximately 3,658,623 individuals. Data were collected to correspond to the period between 1 April and 30 September 2020, which includes a full lockdown period, between 1 April and 4 May, followed by a lighter one.
4.2. Building the Human Network
We simulated a network where each node represents a person or, more precisely, a person’s metagenome. The edges represent possible transmission avenues of microorganisms.
We built the social contact network following the Watts and Strogatz method [18]. At the beginning of each simulation, we construct a regular network in which each individual is connected to the n nearest nodes. Subsequently, we allow each connection to change with a probability (p) of 0.5. If it changes, the node will be connected to another node chosen at random. In this way, we obtain a small-world network that remains unchanged in all cycles.
4.3. Number of Contacts
The distribution of the number of contacts per individual per day is not known in Portugal. We, therefore, used the data from the Polymod project, based on diaries of individual daily contacts in eight European countries [19]. In this project, participants were chosen to register all the individuals they had contact during the day. This provided an idea of the average number of daily contacts for each country analyzed in the project.
In order to estimate the average number of daily contacts in Portugal, we assumed that mobility is correlated with the number of contacts. By comparing the mobility data of google from the different countries that participated in this project with the mobility data from Portugal, we found that the country’s most similar to Portugal are Belgium and Poland. In these countries, there are an average number of 12 and 16 daily contacts, respectively. Following this line of reasoning, in our simulations, we define that the average number of connections—which may or may not become a contact in a given day—between two individuals in the network (n) is 14.
4.4. Contacts per Cycle
As previously mentioned, we initially established a small-world contact network. From the beginning of the simulation, each individual has an assigned set of different individuals to whom he or she will be connected during all the cycles. However, in each cycle, not all contacts occur as there may be a reduction in mobility (in percentage). This reduction is reflected in an equal percentage decrease in the number of contacts that each individual has. For example, suppose with a 100% mobility (base line), an individual is connected to 10 other individuals, with 60% mobility in one cycle. In that case, the individual will be connected in that cycle to only six other individuals. We assume that the connections occurring preferentially are the ones with the closest individuals in the network (representing, for example, the household). For instance, if the connections of individual 1 defined initially are with individuals 2, 4, 5, 9, 18, 22, 27, 35, 48, 87 (ten connections), if there is 60% mobility (a reduction of 4 connections), the connections that do not occur will be with individuals 27, 35, 48 and 87.
In the simulations, we considered three mobility scenarios: (i) using the real mobility data from Google from the period between 1 April and 30 September; (ii) with fixed mobility of 60%; and (iii) with fixed mobility of 40%.
4.5. The Metagenome, Pathogenic Bacteria, and Antibiotic Administration
The model considers the transmission of bacterial pathogens (capable of causing infections) and antibiotic resistance genes between people. These genes are present in the metagenome. We focused on the presence or absence of genes encoding different functions, irrespective of their copy-number in the metagenome. In the simulations, each gene represents a gene family (with similar functions). We divided resistance genes into groups, each group having the same number of families. Each group represents genes associated with resistance to an antibiotic. Of note, we did not consider resistance to multiple drugs in our simulations. Therefore, there will be as many groups as there are antibiotics accounted for in the simulations. We define the diversity of a specific gene kind as the number of genes of that type present in a human metagenome.
We inserted some different bacterial pathogenic species into random individuals per cycle to simulate the migration of bacteria from individuals outside the network or the contamination from sources such as food or contaminated water. In this model, the only difference that we consider between species is the antibiotic to which they are susceptible, as explained below.
Individuals infected by pathogenic bacteria feel sick and take an antibiotic. The antibiotic administered is specific for the bacteria that caused the infectious disease. The antibiotic selects cells carrying resistance genes by eliminating the remaining susceptible bacteria. We assume that all families of resistance genes are present in all metagenomes, but in two different possible states: in some metagenomes, they are present in low copy number, so they are not likely to be transmissible to other individuals in the network; in other metagenomes, the copy number of resistance genes is high due to the selective pressure of antibiotics to which they were previously submitted. In the latter case, resistance genes are likely to be transmissible from person to person.
Moreover, upon antibiotic consumption, the following events can occur (i) elimination of susceptible pathogenic bacterial species; (ii) selection of resistance genes belonging to the group of resistance to that antibiotic (which means their copy number gets so high that they become transferable); (iii) loss of resistance genes associated with other antibiotics with a given probability (becoming non-transferable but still present in minute copy number).
Several processes lead to gene loss. First, genes are lost because of the selective pressure by antibiotics and because we assume that resistance determinants impose a fitness cost (in the absence of antibiotics). To include this cost in the simulations, we consider that each metagenome may lose specific resistance genes according to a “loss rate” (with this process, these genes become non-transferable).
4.6. Algorithm of the Program
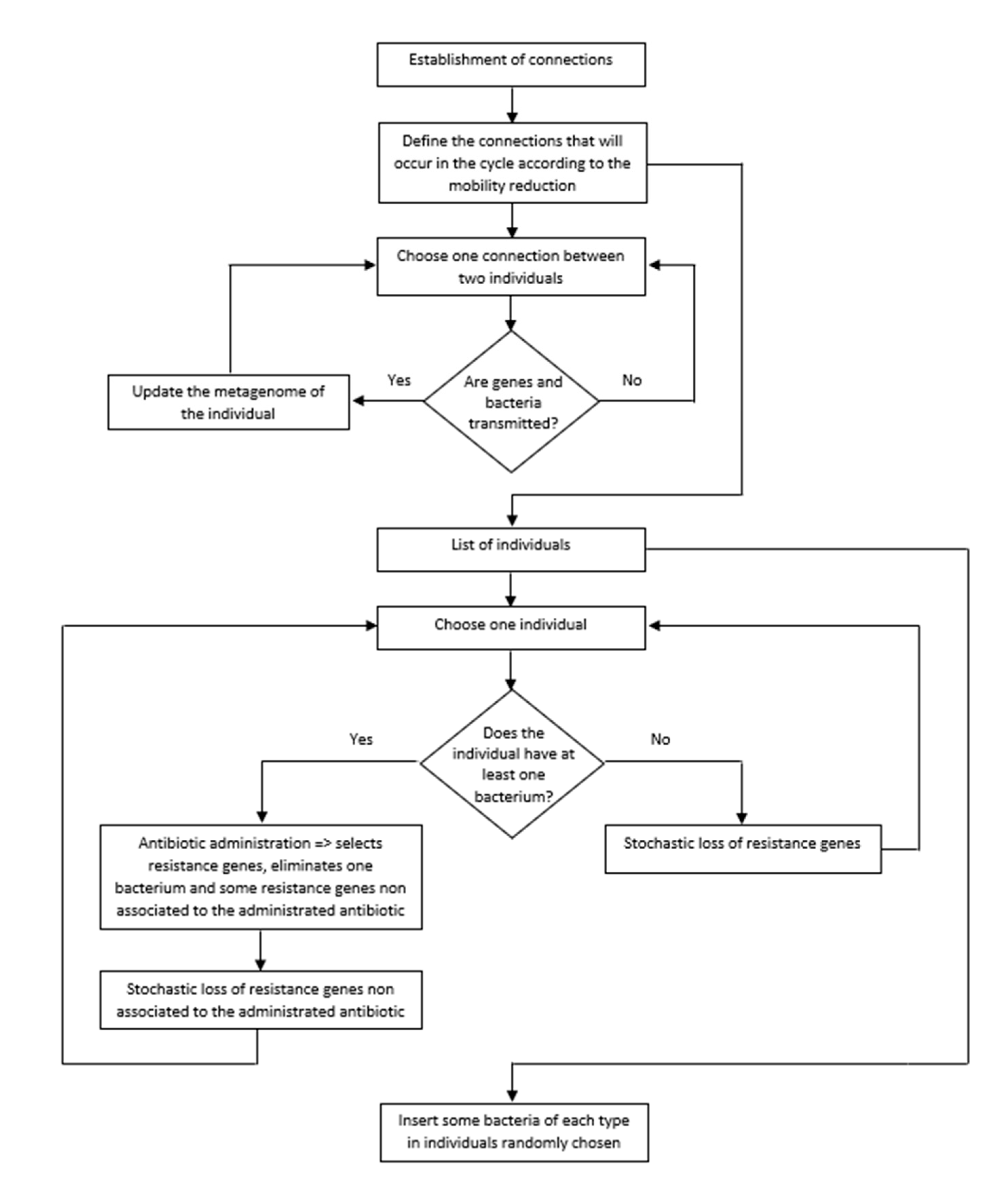
Each simulation is composed of several cycles, where each one corresponds to a day. In each cycle, we considered all procedures described in the pseudocode (Figure 3). In addition, we used as default the parameterized values of our model [7]. The main steps of the program in each cycle are:
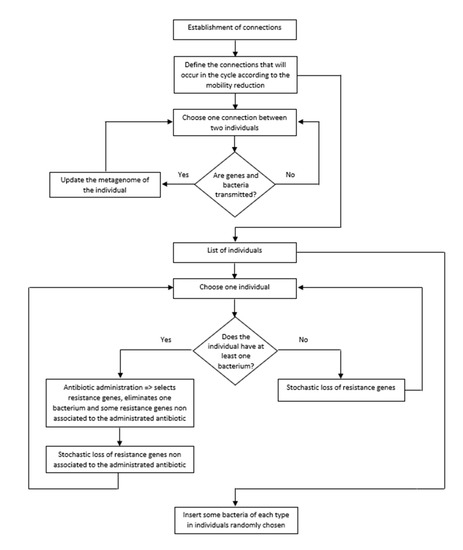
Figure 3.
Flowchart of the algorithm of the program.
- (i)
- Choose the connections that will occur in that cycle according to the percentage of mobility in that cycle (as explained above).
- (ii)
- Transfer of pathogenic bacteria and resistance genes between people (i.e., between linked nodes in this cycle).
- (iii)
- Look for people infected by at least one pathogenic bacterial species. These people take antibiotics (chosen according to the pathogen). The antibiotic eliminates the pathogenic species and selects the resistance genes associated with the antibiotic used. The antibiotic also eliminates resistance genes unrelated to the administered antibiotic. Finally, the metagenome loses a few more resistance genes not associated with the antibiotic. The cause of this loss is the fitness cost of resistance genes.
- (iv)
- The metagenomes of people that did not take an antibiotic in this cycle lose resistance genes. This loss is a consequence of the fitness cost imposed by resistance genes on their hosts.
- (v)
- Insert five bacterial pathogenic species in individuals randomly chosen from the population.
4.7. Statistical Analysis
In each simulation, we counted the number of resistance genes of each individual. From these data, we constructed a graph that represents how many individuals (vertical axis) have each amount of resistance genes (horizontal axis) at the end of the 180 cycles. Next, a Kruskal–Wallis test was used to compare the effect of different confinement regimes, followed by a Dunn post hoc analysis. We performed both tests with R—version 3.5.1 [20] and used the FSA package [21] to perform Dunn’s test.
Author Contributions
Conceptualization: J.S.R., C.P.F.D., F.D. and T.N.; Formal Analysis: J.S.R., C.P.F.D., F.D., M.C.G. and T.N.; Investigation and Methodology: J.S.R., C.P.F.D., F.D. and T.N.; Project administration and Supervision: F.D. and T.N.; Software: J.S.R. and C.P.F.D.; Visualization: J.S.R., C.P.F.D., F.D. and T.N.; Writing—original draft: J.S.R., C.P.F.D., F.D. and T.N.; Writing—review & editing: J.S.R., C.P.F.D., M.C.G., F.D., A.B. and T.N. All authors have read and agreed to the published version of the manuscript.
Funding
This work received no specific grant from any funding agency. T.N. was supported by contract ALG-01-0145-FEDER-028824/PTDC/BIA-MIC/28824/2017. Fundação para a Ciência e a Tecnologia (FCT), I.P., supports cE3c through contract UIDP/00329/2020.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Ethical Approval
No ethics approval was required for this work, as we do not use any personal data. The public databases used for the parameterization of computer simulations have anonymous information on mobility and the average of human contacts.
Acknowledgments
We thank Margarida Duarte for her enthusiasm and motivation during the conceptualization of this work, Octávio Serra for the technical support, and Fernanda Simões for the access to the server at INIAV.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Anderson, R.M. The Pandemic of Antibiotic Resistance. Nat. Med. 1999, 5, 147–149. [Google Scholar] [CrossRef] [PubMed]
- WHO. No Time to Wait: Securing the Future from Drug-Resistant Infections. Available online: http://www.who.int/antimicrobial-resistance/interagency-coordination-group/final-report/en/ (accessed on 25 February 2021).
- Smith, R.; Coast, J. The True Cost of Antimicrobial Resistance. BMJ 2013, 346, f1493. [Google Scholar] [CrossRef] [Green Version]
- Gyssens, I.C.; Van Den Broek, P.J.; Kullberg, B.-J.; Hekster, Y.A.; Van Der Meer, J.W.M. Optimizing Antimicrobial Therapy. A Method for Antimicrobial Drug Me Evaluation. J. Antimicrob. Chemother. 1992, 30, 724–727. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Yu, V.L. Rational Empiric Antibiotic Prescription in the ICU. Chest 2000, 117, 1496–1499. [Google Scholar] [CrossRef] [PubMed]
- Escudeiro, P.; Pothier, J.; Dionisio, F.; Nogueira, T. Antibiotic Resistance Gene Diversity and Virulence Gene Diversity Are Correlated in Human Gut and Environmental Microbiomes. mSphere 2019, 4, e00135-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domingues, C.P.F.; Rebelo, J.S.; Pothier, J.; Monteiro, F.; Nogueira, T.; Dionisio, F. The Perfect Condition for the Rising of Superbugs: Person-to-Person Contact and Antibiotic Use Are the Key Factors Responsible for the Positive Correlation between Antibiotic Resistance Gene Diversity and Virulence Gene Diversity in Human Metagenomes. Antibiotics 2021, 10, 605. [Google Scholar] [CrossRef] [PubMed]
- A Região. Available online: http://www.ccdr-lvt.pt/pt/a-regiao/10568.htm (accessed on 18 June 2021).
- COVID-19 Community Mobility Report. Available online: https://www.google.com/covid19/mobility?hl=pt-PT (accessed on 30 October 2020).
- Domingues, C.P.F.; Rebelo, J.S.; Dionisio, F.; Botelho, A.; Nogueira, T. The Social Distancing Imposed to Contain COVID-19 Can Affect Our Microbiome: A Double-Edged Sword in Human Health. mSphere 2020, 5, e00716-20. [Google Scholar] [CrossRef] [PubMed]
- Dia Europeu Do Antibiótico 2020—INSA. Available online: http://www.insa.min-saude.pt/dia-europeu-do-antibiotico-2020/ (accessed on 18 June 2021).
- David, P.H.C.; Sá-Pinto, X.; Nogueira, T. Using SimulATe to Model the Effects of Antibiotic Selective Pressure on the Dynamics of Pathogenic Bacterial Populations. Biol. Methods Protoc. 2019, 4, bpz004. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, T.; David, P.H.C.; Pothier, J. Antibiotics as Both Friends and Foes of the Human Gut Microbiome: The Microbial Community Approach. Drug Dev. Res. 2019, 80, 86–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, P.; Sá-Pinto, X.; Nogueira, T. Microbiomes Are Important Key Players in Human Health-Mutualistic Interactions with and between Our Microbes. JOJ Intern. Med. 2019, 1, 28–33. [Google Scholar]
- Jiang, T.; Wu, H.; Yang, X.; Li, Y.; Zhang, Z.; Chen, F.; Zhao, L.; Zhang, C. Lactobacillus Mucosae Strain Promoted by a High-Fiber Diet in Genetic Obese Child Alleviates Lipid Metabolism and Modifies Gut Microbiota in ApoE−/− Mice on a Western Diet. Microorganisms 2020, 8, 1225. [Google Scholar] [CrossRef] [PubMed]
- Lkhagva, E.; Chung, H.-J.; Hong, J.; Tang, W.H.W.; Lee, S.-I.; Hong, S.-T.; Lee, S. The Regional Diversity of Gut Microbiome along the GI Tract of Male C57BL/6 Mice. BMC Microbiol. 2021, 21, 44. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Rodriguez, E.; Egea-Zorrilla, A.; Plaza-Díaz, J.; Aragón-Vela, J.; Muñoz-Quezada, S.; Tercedor-Sánchez, L.; Abadia-Molina, F. The Gut Microbiota and Its Implication in the Development of Atherosclerosis and Related Cardiovascular Diseases. Nutrients 2020, 12, 605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, D.J.; Strogatz, S.H. Collective Dynamics of ‘Small-World’ Networks. Nature 1998, 393, 440–442. [Google Scholar] [CrossRef] [PubMed]
- Mossong, J.; Hens, N.; Jit, M.; Beutels, P.; Auranen, K.; Mikolajczyk, R.; Massari, M.; Salmaso, S.; Tomba, G.S.; Wallinga, J.; et al. Social Contacts and Mixing Patterns Relevant to the Spread of Infectious Diseases. PLoS Med. 2008, 5, e74. [Google Scholar] [CrossRef] [PubMed]
- R: The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 18 June 2021).
- Ogle, D.H.; Wheeler, P.; Dinno, A. FSA: Fisheries Stock Analysis. R Package Version 0.8.32. 2021. Available online: https://github.com/droglenc/FSA (accessed on 17 February 2021).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).