
Telomere Length in Chromosomally Normal and Abnormal Miscarriages and Ongoing Pregnancies and Its Association with 5-hydroxymethylcytosine Patterns

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
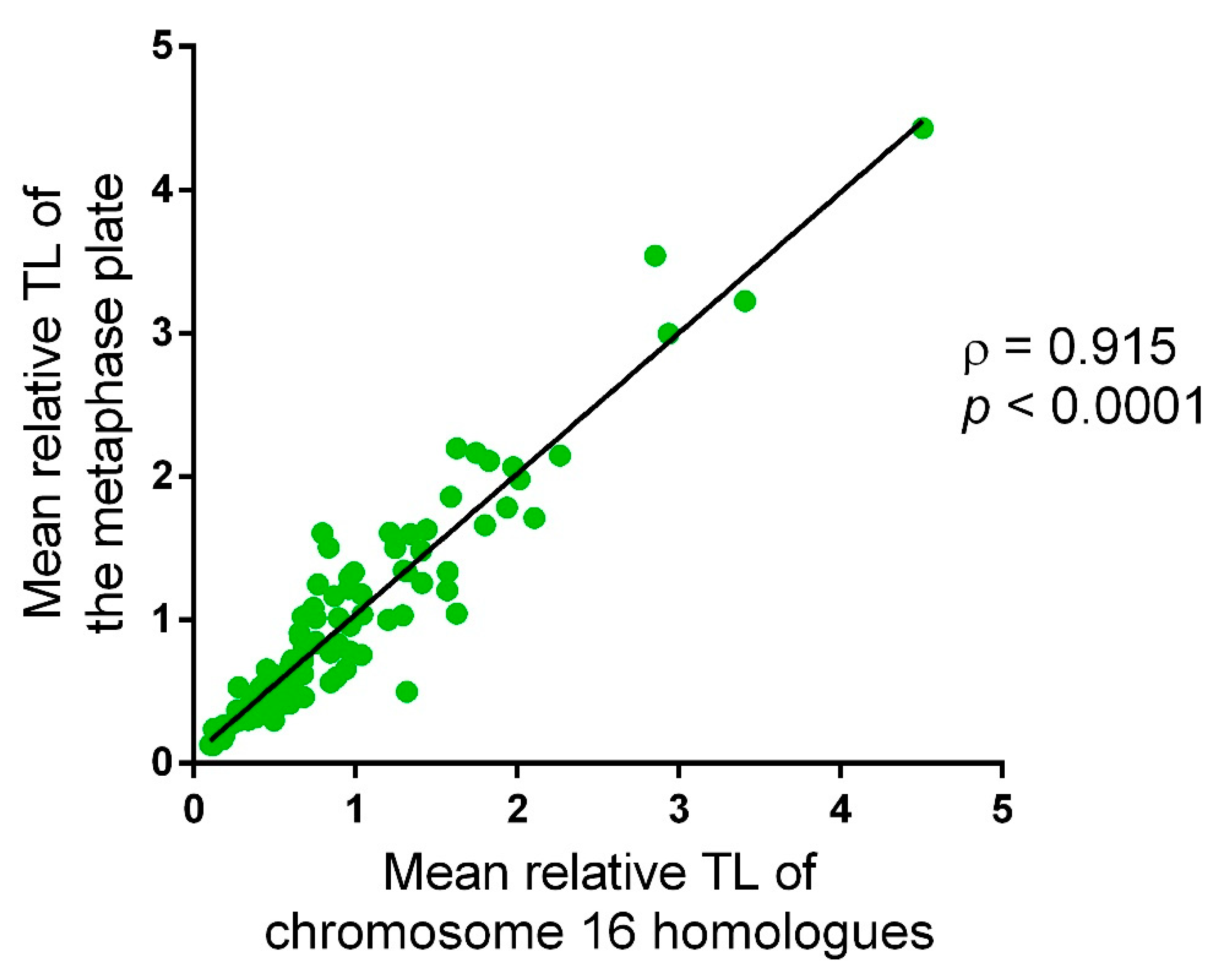
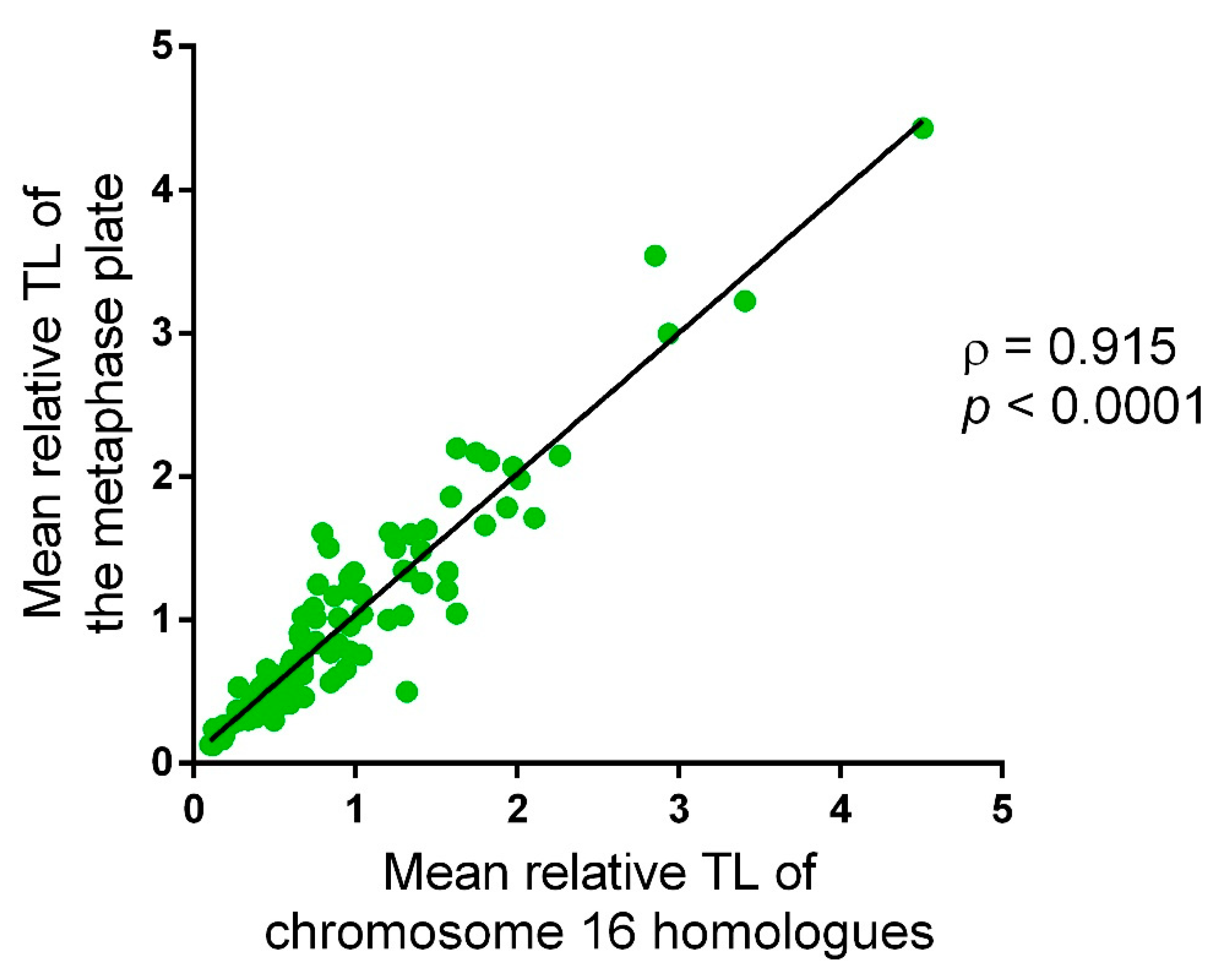
2.1. Approach Applicability to Telomere Length Analysis in Chorionic Cytotrophoblast Cells
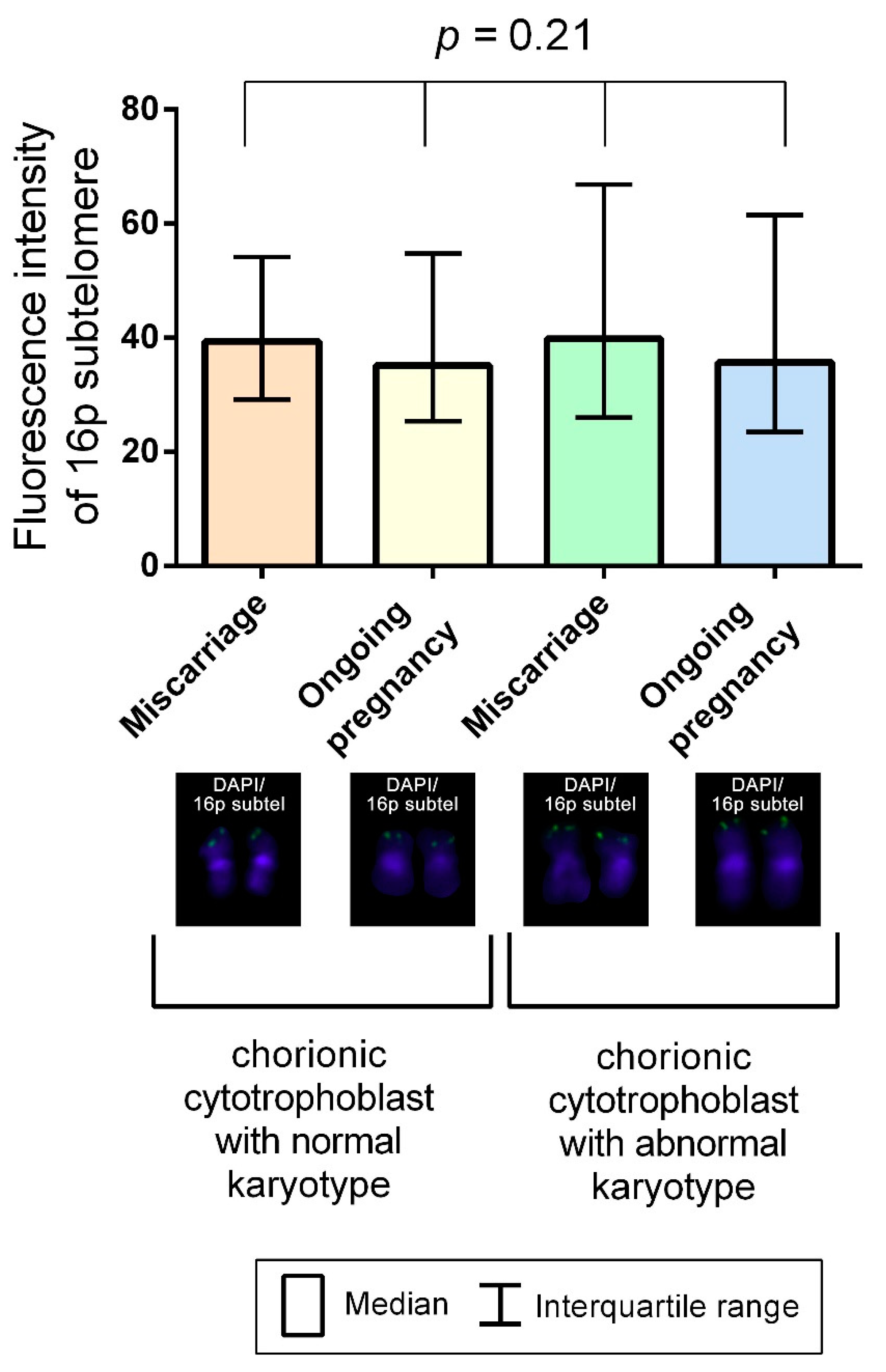
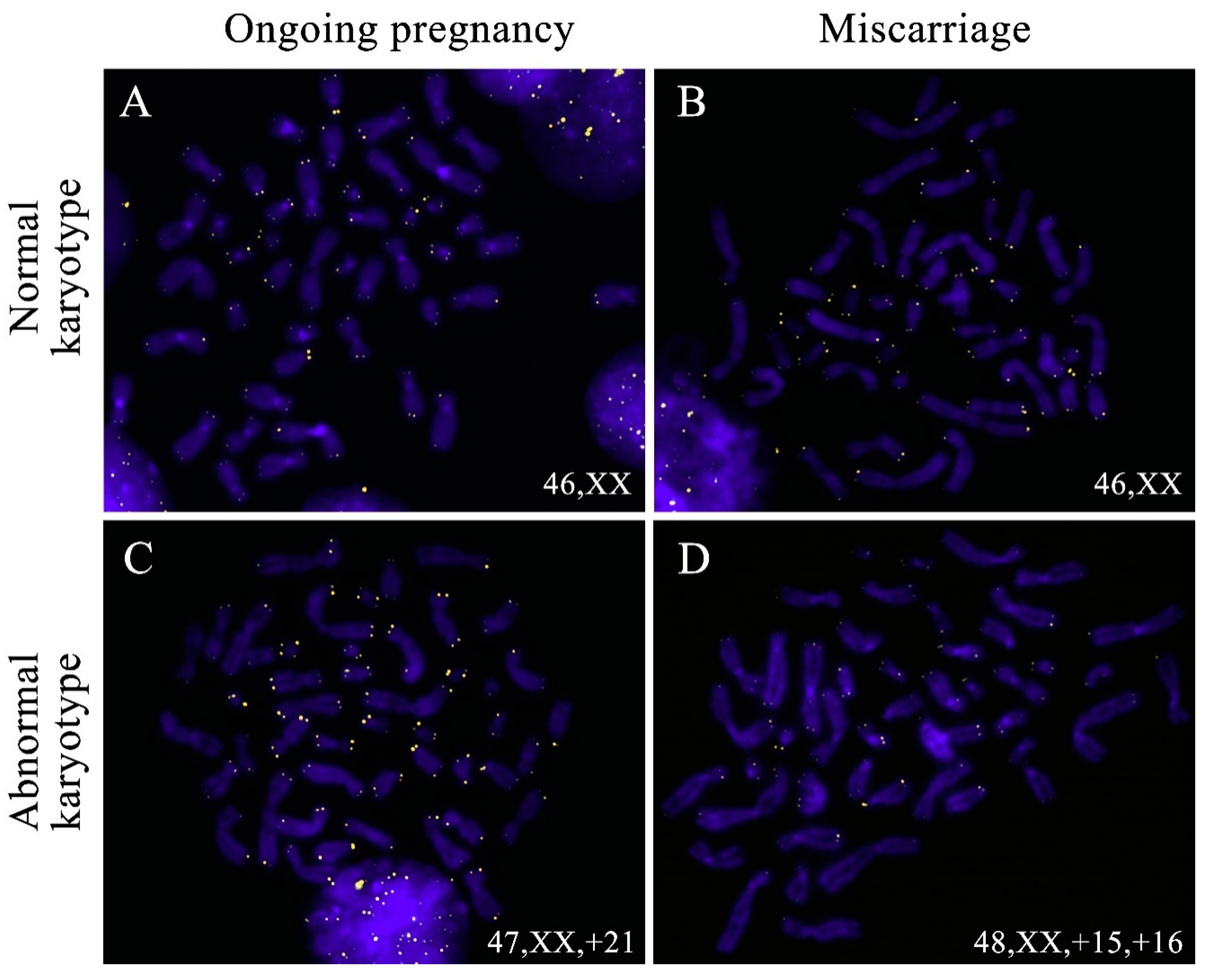
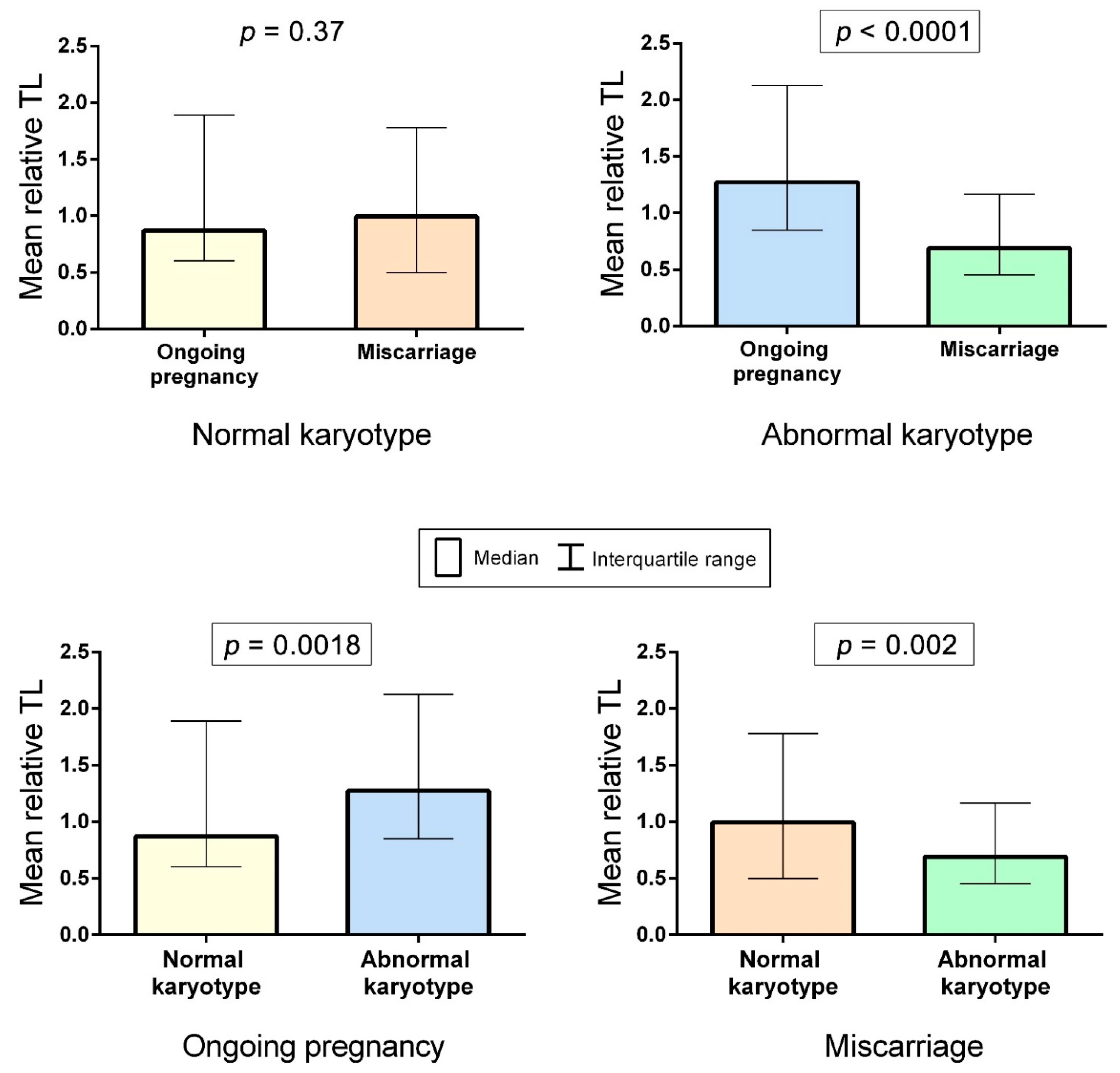
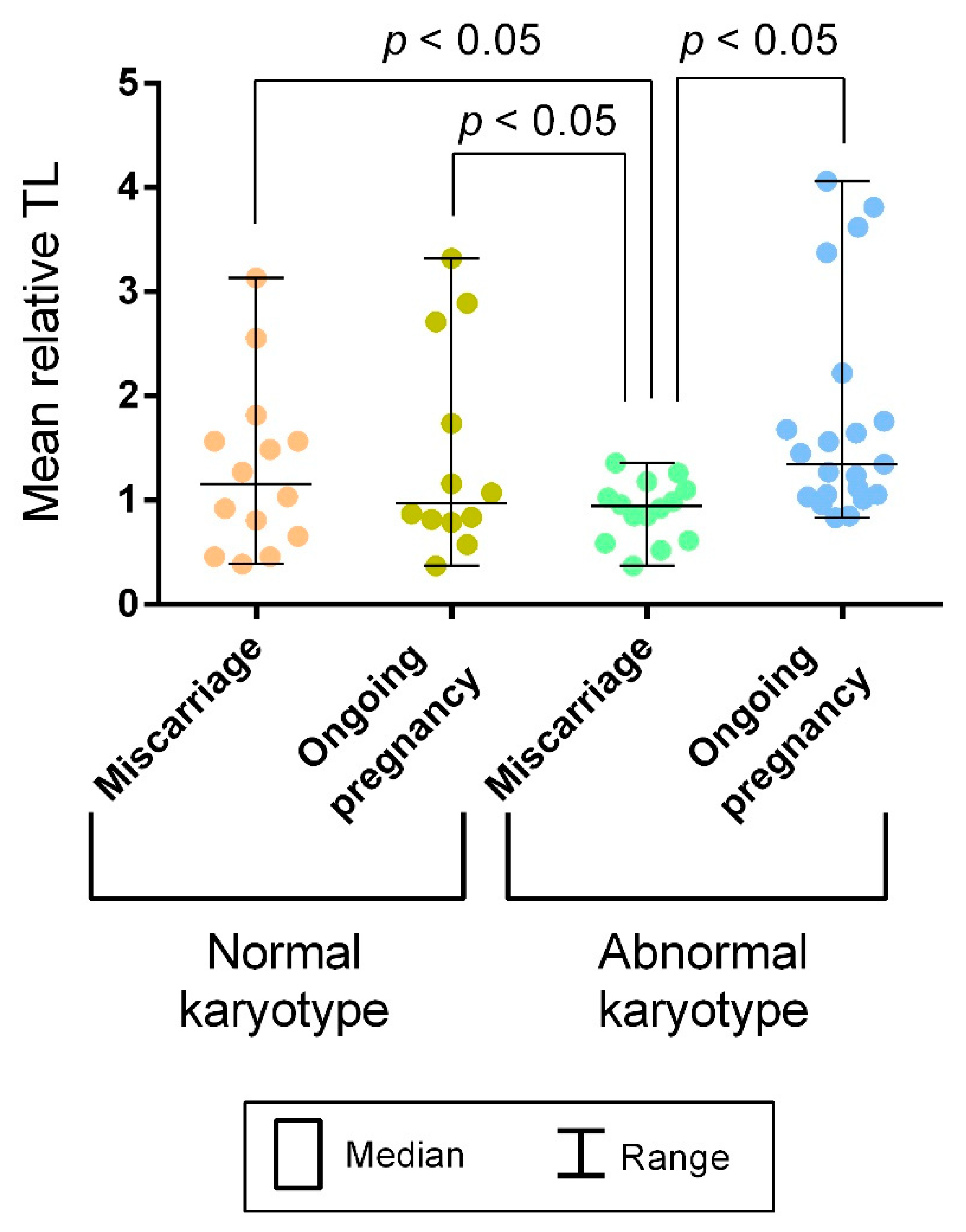
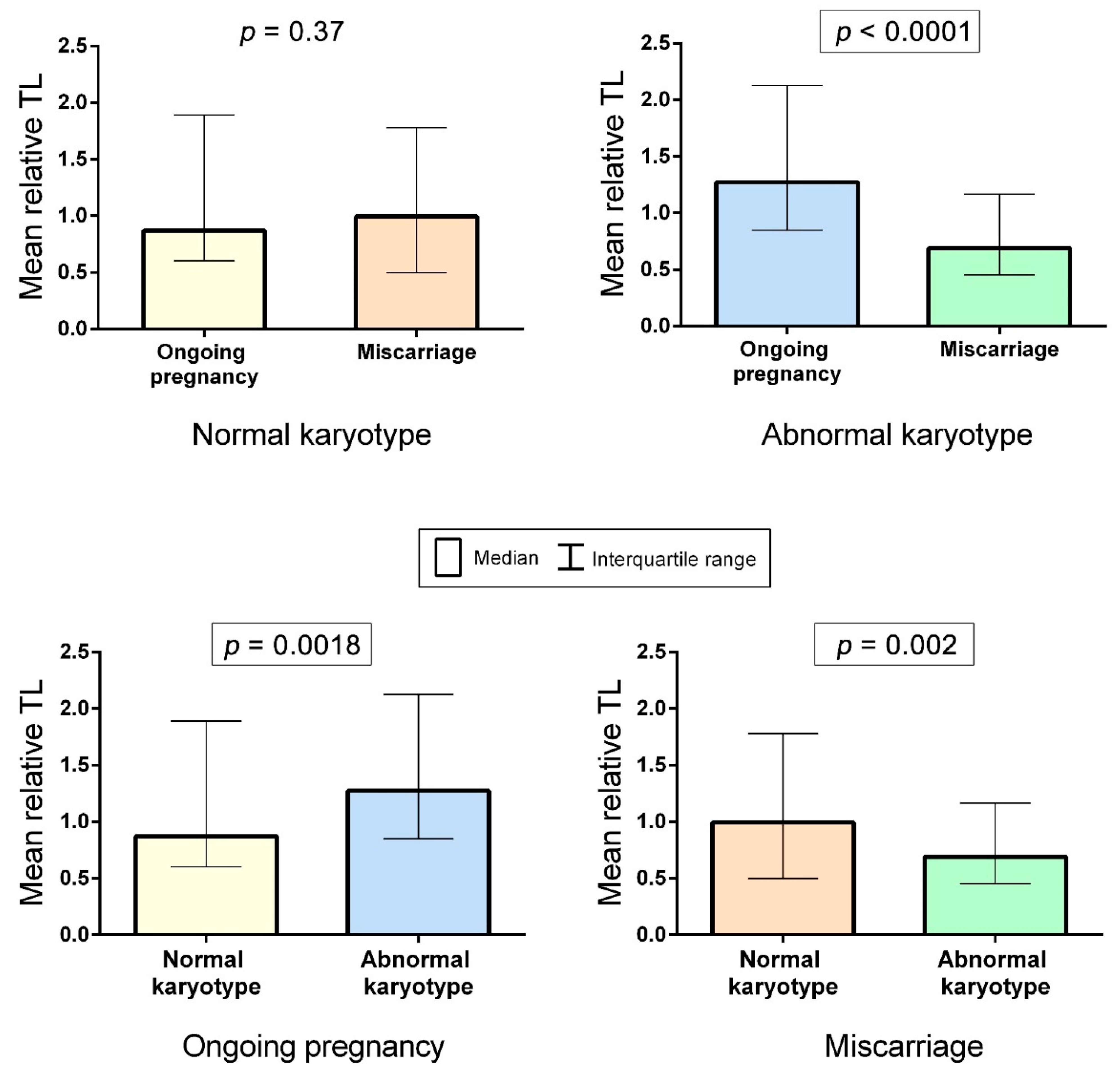
2.2. A Comparison of Telomere Lengths in Chorionic Cytotrophoblast from Karyotypically Normal and Abnormal Miscarriages and Ongoing Pregnancies
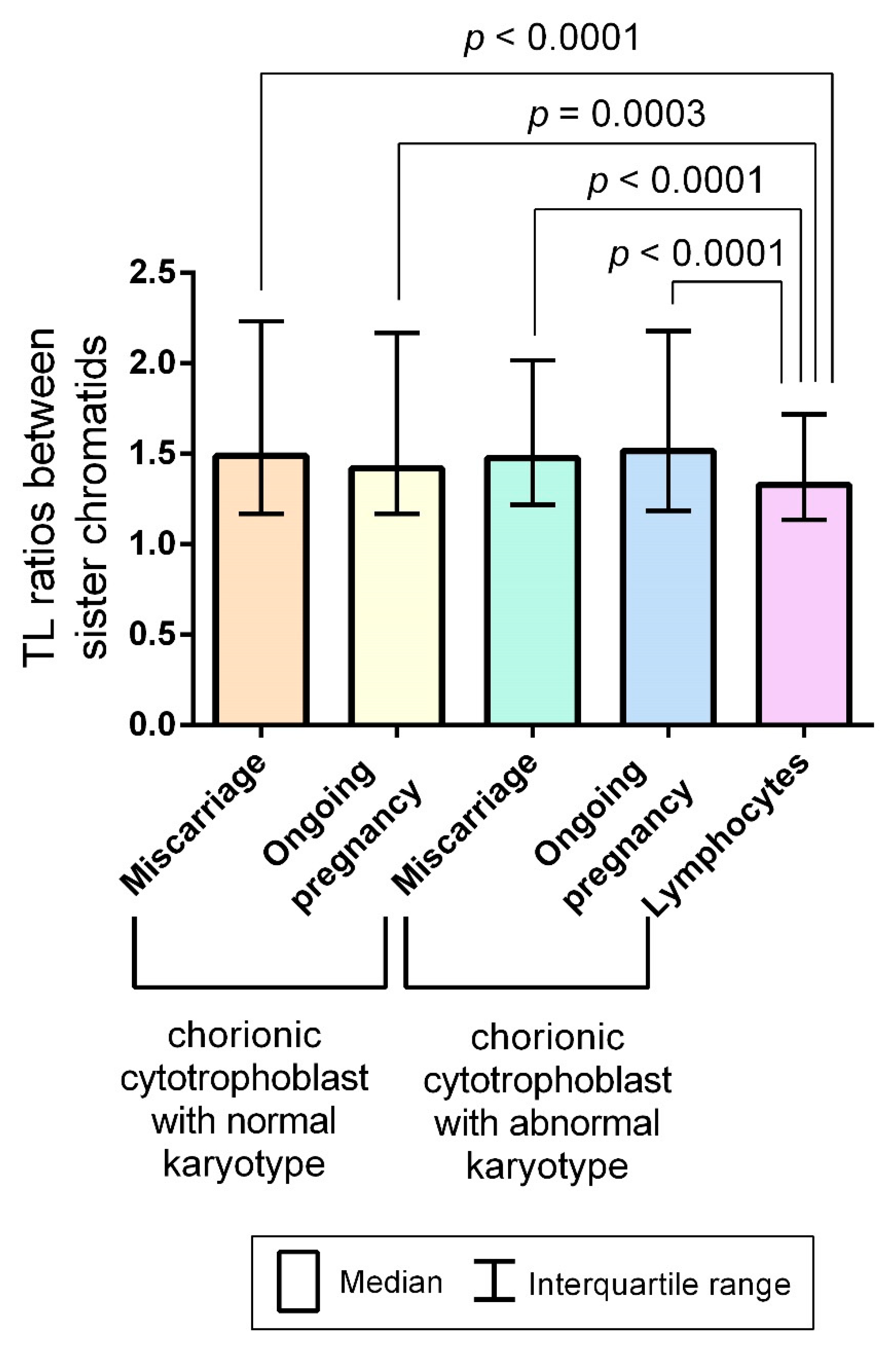
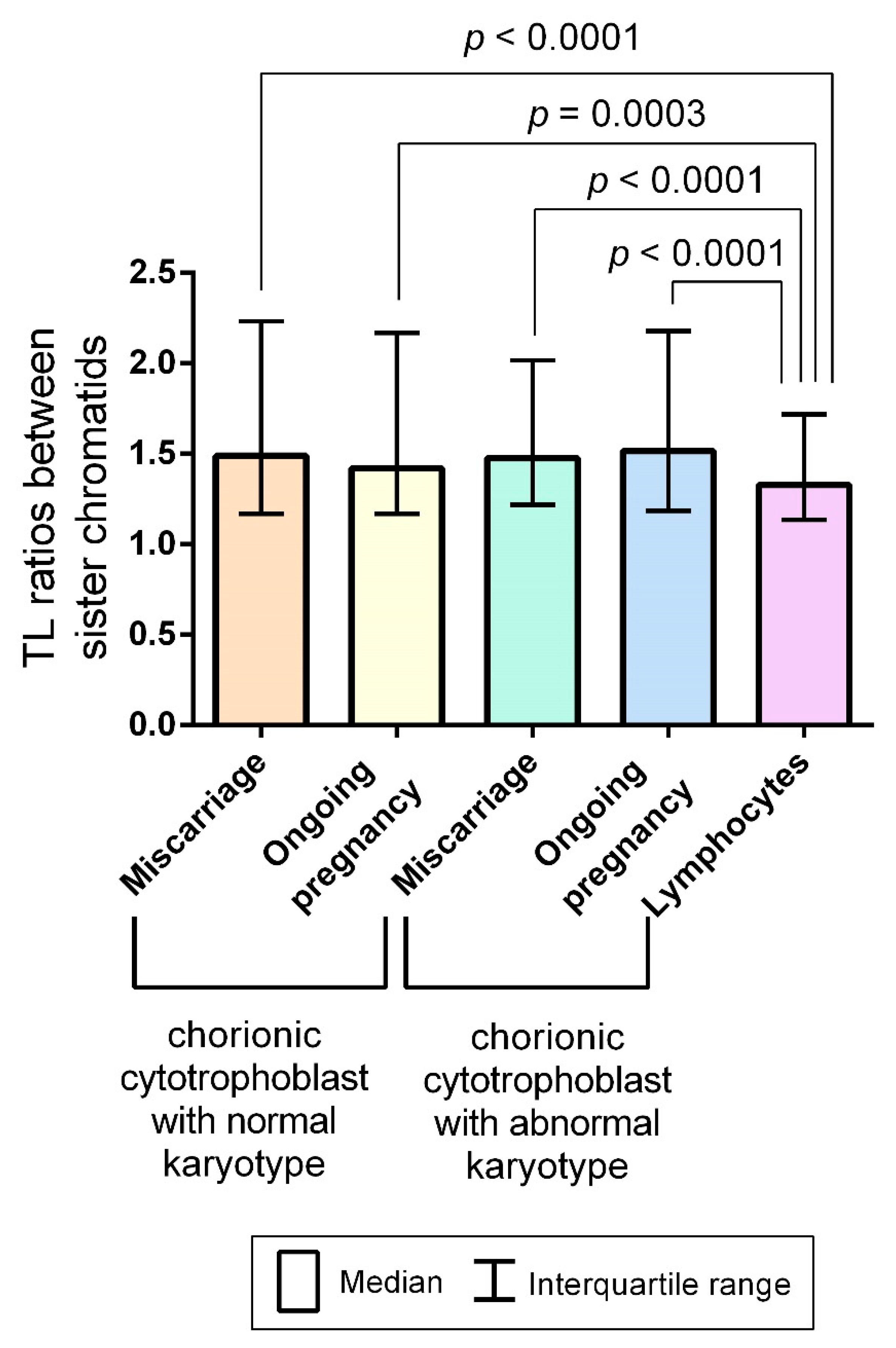
2.3. Interchromatid Telomere Length Asymmetry in Chorionic Cytotrophoblast and Adult Lymphocytes
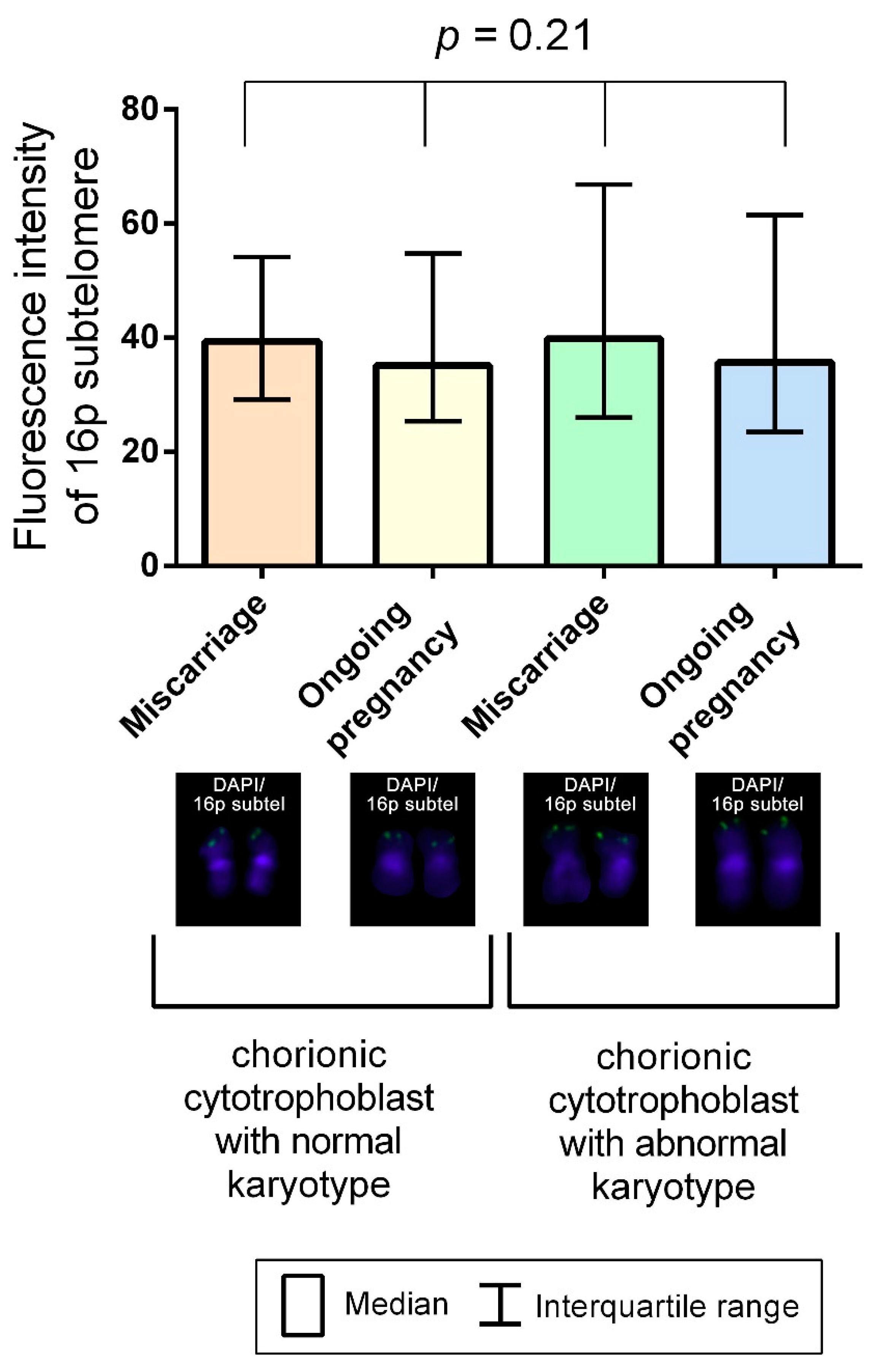
2.4. Interchromatid Telomere Length Asymmetry in Chorionic Cytotrophoblast Is Linked to DNA Hydroxymethylation Pattern
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Chromosome Preparation
4.3. Fluorescence In Situ Hybridisation (FISH)
4.4. Immunodetection of 5-Hydroxymethylcytosine (5hmC)
4.5. Image Acquisition and Evaluation of Telomeric and Subtelomeric FISH-Signal Intensity
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Moyzis, R.K.; Buckingham, J.M.; Cram, L.S.; Dani, M.; Deaven, L.L.; Jones, M.D.; Meyne, J.; Ratliff, R.L.; Wu, J.R. A highly conserved repetitive DNA sequence, (TTAGGG)n, present at the telomeres of human chromosomes. Proc. Natl. Acad. Sci. USA 1988, 85, 6622–6626. [Google Scholar] [CrossRef] [Green Version]
- De Lange, T. Shelterin: The protein complex that shapes and safeguards human telomeres. Genes Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azzalin, C.M.; Reichenbach, P.; Khoriauli, L.; Giulotto, E.; Lingner, J. Telomeric Repeat Containing RNA and RNA Surveillance Factors at Mammalian Chromosome Ends. Science 2007, 318, 798–801. [Google Scholar] [CrossRef]
- Sfeir, A.; De Lange, T. Removal of Shelterin Reveals the Telomere End-Protection Problem. Science 2012, 336, 593–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackburn, E.H.; Epel, E.S.; Lin, J. Human telomere biology: A contributory and interactive factor in aging, disease risks, and protection. Science 2015, 350, 1193–1198. [Google Scholar] [CrossRef] [Green Version]
- Muller, H.J. The remaking of chromosomes. Collect. Nat. 1938, 13, 181–198. [Google Scholar]
- McClintock, B. The Behavior in Successive Nuclear Divisions of a Chromosome Broken at Meiosis. Proc. Natl. Acad. Sci. USA 1939, 25, 405–416. [Google Scholar] [CrossRef] [Green Version]
- Hastie, N.D.; Dempster, M.; Dunlop, M.G.; Thompson, A.M.; Green, D.K.; Allshire, R.C. Telomere reduction in human colorectal carcinoma and with ageing. Nat. Cell Biol. 1990, 346, 866–868. [Google Scholar] [CrossRef]
- Olovnikov, A.M. A theory of marginotomy: The incomplete copying of template margin in enzymic synthesis of polynucleotides and biological significance of the phenomenon. J. Theor. Biol. 1973, 41, 181–190. [Google Scholar] [CrossRef]
- Oikawa, S.; Kawanishi, S. Site-specific DNA damage at GGG sequence by oxidative stress may accelerate telomere shortening. FEBS Lett. 1999, 453, 365–368. [Google Scholar] [CrossRef] [Green Version]
- Di Fagagna, F.D.; Reaper, P.M.; Clay-Farrace, L.; Fiegler, H.; Carr, P.; von Zglinicki, T.; Saretzki, G.; Carter, N.P.; Jackson, S.P. A DNA damage checkpoint response in telomere-initiated senescence. Nat. Cell Biol. 2003, 426, 194–198. [Google Scholar] [CrossRef]
- Kawanishi, S.; Oikawa, S. Mechanism of Telomere Shortening by Oxidative Stress. Ann. N. Y. Acad. Sci. 2004, 1019, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Greider, C.W.; Blackburn, E.H. Identification of a specific telomere terminal transferase activity in tetrahymena extracts. Cell 1985, 43, 405–413. [Google Scholar] [CrossRef]
- Bryan, T.M.; Englezou, A.; Dalla-Pozza, L.; Dunham, M.A.; Reddel, R.R. Evidence for an alternative mechanism for maintaining telomere length in human tumors and tumor-derived cell lines. Nat. Med. 1997, 3, 1271–1274. [Google Scholar] [CrossRef] [PubMed]
- Dunham, M.A.; Neumann, A.A.; Fasching, C.L.; Reddel, R.R. Telomere maintenance by recombination in human cells. Nat. Genet. 2000, 26, 447–450. [Google Scholar] [CrossRef]
- Bailey, S.M.; Brenneman, M.A.; Goodwin, E.H. Frequent recombination in telomeric DNA may extend the proliferative life of telomerase-negative cells. Nucleic Acids Res. 2004, 32, 3743–3751. [Google Scholar] [CrossRef]
- Koltsova, A.S.; Pendina, A.A.; Efimova, O.A.; Chiryaeva, O.G.; Kuznetzova, T.V.; Baranov, V.S. On the Complexity of Mechanisms and Consequences of Chromothripsis: An Update. Front. Genet. 2019, 10, 393. [Google Scholar] [CrossRef]
- Shorokhova, M.; Nikolsky, N.; Grinchuk, T. Chromothripsis—Explosion in Genetic Science. Cells 2021, 10, 1102. [Google Scholar] [CrossRef]
- Wu, X.; Amos, C.I.; Zhu, Y.; Zhao, H.; Grossman, B.H.; Shay, J.W.; Luo, S.; Hong, W.K.; Spitz, M.R. Telomere Dysfunction: A Potential Cancer Predisposition Factor. J. Natl. Cancer Inst. 2003, 95, 1211–1218. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.S.; Choi, Y.Y.; Lee, W.K.; Choi, J.E.; Cha, S.I.; Kim, Y.J.; Kim, C.H.; Kam, S.; Jung, T.H.; Park, J.Y. Telomere length and the risk of lung cancer. Cancer Sci. 2008, 99, 1385–1389. [Google Scholar] [CrossRef]
- Shen, J.; Gammon, M.D.; Terry, M.B.; Wang, Q.; Bradshaw, P.; Teitelbaum, S.L.; Neugut, A.I.; Santella, R.M. Telomere length, oxidative damage, antioxidants and breast cancer risk. Int. J. Cancer 2009, 124, 1637–1643. [Google Scholar] [CrossRef] [Green Version]
- Rampazzo, E.; Cecchin, E.; Del Bianco, P.; Menin, C.; Spolverato, G.; Giunco, S.; Lonardi, S.; Malacrida, S.; De Paoli, A.; Toffoli, G.; et al. Genetic Variants of the TERT Gene, Telomere Length, and Circulating TERT as Prognostic Markers in Rectal Cancer Patients. Cancers 2020, 12, 3115. [Google Scholar] [CrossRef]
- Samani, N.J.; Boultby, R.; Butler, R.; Thompson, J.R.; Goodall, A.H. Telomere shortening in atherosclerosis. Lancet 2001, 358, 472–473. [Google Scholar] [CrossRef]
- Fitzpatrick, A.L.; Kronmal, R.A.; Gardner, J.P.; Psaty, B.M.; Jenny, N.S.; Tracy, R.P.; Walston, J.; Kimura, M.; Aviv, A. Leukocyte Telomere Length and Cardiovascular Disease in the Cardiovascular Health Study. Am. J. Epidemiol. 2006, 165, 14–21. [Google Scholar] [CrossRef]
- Guo, N.; Parry, E.M.; Li, L.-S.; Kembou, F.; Lauder, N.; Hussain, M.A.; Berggren, P.-O.; Armanios, M. Short Telomeres Compromise β-Cell Signaling and Survival. PLoS ONE 2011, 6, e17858. [Google Scholar] [CrossRef] [Green Version]
- Kota, L.N.; Purushottam, M.; Moily, N.S.; Jain, S. Shortened telomere in unremitted schizophrenia. Psychiatry Clin. Neurosci. 2015, 69, 292–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Huo, Y.R.; Wang, J.; Wang, C.; Liu, S.; Liu, S.; Wang, J.; Ji, Y. Telomere Shortening in Alzheimer’s Disease Patients. Ann. Clin. Lab. Sci. 2016, 46, 260–265. [Google Scholar]
- Thilagavathi, J.; Mishra, S.S.; Kumar, M.; Vemprala, K.; Deka, D.; Dhadwal, V.; Dada, R. Analysis of telomere length in couples experiencing idiopathic recurrent pregnancy loss. J. Assist. Reprod. Genet. 2013, 30, 793–798. [Google Scholar] [CrossRef] [Green Version]
- Thilagavathi, J.; Kumar, M.; Mishra, S.S.; Venkatesh, S.; Kumar, R.; Dada, R. Analysis of sperm telomere length in men with idiopathic infertility. Arch. Gynecol. Obstet. 2013, 287, 803–807. [Google Scholar] [CrossRef]
- Treff, N.R.; Su, J.; Taylor, D.; Scott, R.T. Telomere DNA Deficiency Is Associated with Development of Human Embryonic Aneuploidy. PLoS Genet. 2011, 7, e1002161. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Feingold, E.; Chakraborty, S.; Dey, S.K. Telomere length is associated with types of chromosome 21 nondisjunction: A new insight into the maternal age effect on Down syndrome birth. Hum. Genet. 2010, 127, 403–409. [Google Scholar] [CrossRef]
- Davy, P.; Nagata, M.; Bullard, P.; Fogelson, N.; Allsopp, R. Fetal Growth Restriction is Associated with Accelerated Telomere Shortening and Increased Expression of Cell Senescence Markers in the Placenta. Placenta 2009, 30, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biron-Shental, T.; Sukenik-Halevy, R.; Sharon, Y.; Goldberg-Bittman, L.; Kidron, D.; Fejgin, M.D.; Amiel, A. Short telomeres may play a role in placental dysfunction in preeclampsia and intrauterine growth restriction. Am. J. Obstet. Gynecol. 2010, 202, 381.e1–381.e7. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, F.; Facchinetti, F.; Saade, G.; Menon, R. Placental telomere shortening in stillbirth: A sign of premature senescence? J. Matern. Fetal. Neonatal Med. 2015, 29, 1283–1288. [Google Scholar] [CrossRef]
- Gonzalo, S.; Jaco, I.; Fraga, M.F.; Chen, T.; Li, E.; Esteller, M.; Blasco, M.A. DNA methyltransferases control telomere length and telomere recombination in mammalian cells. Nat. Cell Biol. 2006, 8, 416–424. [Google Scholar] [CrossRef]
- Benetti, R.; Gonzalo, S.; Jaco, I.; Muñoz, P.; Gonzalez, S.; Schoeftner, S.; Murchison, E.; Andl, T.; Chen, T.; Klatt, P.; et al. A mammalian microRNA cluster controls DNA methylation and telomere recombination via Rbl2-dependent regulation of DNA methyltransferases. Nat. Struct. Mol. Biol. 2008, 15, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Dan, J.; Rousseau, P.; Hardikar, S.; Veland, N.; Wong, J.; Autexier, C.; Chen, T. Zscan4 Inhibits Maintenance DNA Methylation to Facilitate Telomere Elongation in Mouse Embryonic Stem Cells. Cell Rep. 2017, 20, 1936–1949. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, A.K.; Sharma, S.; Sengupta, S.; Saha, D.; Kumar, P.; Hussain, T.; Srivastava, V.; Roy, S.D.; Shay, J.W.; Chowdhury, S. Telomere length-dependent transcription and epigenetic modifications in promoters remote from telomere ends. PLoS Genet. 2018, 14, e1007782. [Google Scholar] [CrossRef]
- Szulwach, K.E.; Li, X.; Li, Y.; Song, C.-X.; Han, J.W.; Kim, S.; Namburi, S.; Hermetz, K.; Kim, J.J.; Rudd, M.K.; et al. Integrating 5-Hydroxymethylcytosine into the Epigenomic Landscape of Human Embryonic Stem Cells. PLoS Genet. 2011, 7, e1002154. [Google Scholar] [CrossRef] [Green Version]
- Kubiura, M.; Okano, M.; Kimura, H.; Kawamura, F.; Tada, M. Chromosome-wide regulation of euchromatin-specific 5mC to 5hmC conversion in mouse ES cells and female human somatic cells. Chromosom. Res. 2012, 20, 837–848. [Google Scholar] [CrossRef] [Green Version]
- Bogomazova, A.N.; Lagarkova, M.A.; Panova, A.V.; Nekrasov, E.D.; Kiselev, S.L. Reactivation of X chromosome upon reprogramming leads to changes in the replication pattern and 5hmC accumulation. Chromosoma 2013, 123, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Efimova, O.A.; Pendina, A.A.; Krapivin, M.I.; Kopat, V.V.; Tikhonov, A.V.; Petrovskaia-Kaminskaia, A.V.; Navodnikova, P.M.; Talantova, O.E.; Glotov, O.S.; Baranov, V.S. Inter-Cell and Inter-Chromosome Variability of 5-Hydroxymethylcytosine Patterns in Noncultured Human Embryonic and Extraembryonic Cells. Cytogenet. Genome Res. 2018, 156, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Kyo, S.; Takakura, M.; Tanaka, M.; Kanaya, T.; Sagawa, T.; Kohama, T.; Ishikawa, H.; Nakano, T.; Shimoya, K.; Inoue, M. Expression of Telomerase Activity in Human Chorion. Biochem. Biophys. Res. Commun. 1997, 241, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.-J.; Chu, C.-T.; Huang, S.-C.; Chow, S.-N.; Hsieh, C.-Y. Telomerase activity in gestational trophoblastic disease and placental tissue from early and late human pregnancies. Hum. Reprod. 2002, 17, 463–468. [Google Scholar] [CrossRef] [Green Version]
- Colatto, B.N.; De Souza, I.F.; Schinke, L.A.A.; Noda-Nicolau, N.M.; Da Silva, M.G.; Morceli, G.; Menon, R.; Polettini, J. Telomere Length and Telomerase Activity in Foetal Membranes from Term and Spontaneous Preterm Births. Reprod. Sci. 2020, 27, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Polettini, J.; Da Silva, M.G. Telomere-Related Disorders in Fetal Membranes Associated with Birth and Adverse Pregnancy Outcomes. Front. Physiol. 2020, 11, 561771. [Google Scholar] [CrossRef]
- Novakovic, B.; Napier, C.E.; Vryer, R.; Dimitriadis, E.; Manuelpillai, U.; Sharkey, A.; Craig, J.M.; Reddel, R.R.; Saffery, R. DNA methylation mediated up-regulation of TERRA non-coding RNA is coincident with elongated telomeres in the human placenta. Mol. Hum. Reprod. 2016, 22, 791–799. [Google Scholar] [CrossRef] [Green Version]
- Lu, F.; Liu, Y.; Jiang, L.; Yamaguchi, S.; Zhang, Y. Role of Tet proteins in enhancer activity and telomere elongation. Genes Dev. 2014, 28, 2103–2119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Guo, R.; Wang, H.; Ye, X.; Zhou, Z.; Dan, J.; Wang, H.; Gong, P.; Deng, W.; Yin, Y.; et al. Tet Enzymes Regulate Telomere Maintenance and Chromosomal Stability of Mouse ESCs. Cell Rep. 2016, 15, 1809–1821. [Google Scholar] [CrossRef] [Green Version]
- Buscarlet, M.; Tessier, A.; Provost, S.; Mollica, L.; Busque, L. Human blood cell levels of 5-hydroxymethylcytosine (5hmC) decline with age, partly related to acquired mutations in TET2. Exp. Hematol. 2016, 44, 1072–1084. [Google Scholar] [CrossRef]
- Yamada, S.; Misawa, K.; Mima, M.; Imai, A.; Mochizuki, D.; Yamada, T.; Shinmura, D.; Kita, J.; Ishikawa, R.; Yamaguchi, Y.; et al. Telomere shortening in head and neck cancer: Association between DNA demethylation and survival. J. Cancer 2021, 12, 2165–2172. [Google Scholar] [CrossRef]
- Efimova, O.A.; Koltsova, A.S.; Krapivin, M.I.; Tikhonov, A.V.; Pendina, A.A. Environmental Epigenetics and Genome Flexibility: Focus on 5-Hydroxymethylcytosine. Int. J. Mol. Sci. 2020, 21, 3223. [Google Scholar] [CrossRef]
- Burbano, M.S.J.; Gilson, E. The Power of Stress: The Telo-Hormesis Hypothesis. Cells 2021, 10, 1156. [Google Scholar] [CrossRef]
- Slagboom, P.E.; Droog, S.; Boomsma, D.I. Genetic determination of telomere size in humans: A twin study of three age groups. Am. J. Hum. Genet. 1994, 55, 876–882. [Google Scholar]
- Graakjaer, J.; Bischoff, C.; Korsholm, L.; Holstebroe, S.; Vach, W.; Bohr, V.A.; Christensen, K.; Kølvraa, S. The pattern of chromosome-specific variations in telomere length in humans is determined by inherited, telomere-near factors and is maintained throughout life. Mech. Ageing Dev. 2003, 124, 629–640. [Google Scholar] [CrossRef]
- Graakjaer, J.; Pascoe, L.; Der-Sarkissian, H.; Thomas, G.; Kolvraa, S.; Christensen, K.; Londono-Vallejo, J.-A. The relative lengths of individual telomeres are defined in the zygote and strictly maintained during life. Aging Cell 2004, 3, 97–102. [Google Scholar] [CrossRef]
- Pendina, A.A.; Krapivin, M.I.; Efimova, O.A.; Tikhonov, A.V.; Mekina, I.D.; Komarova, E.M.; Koltsova, A.S.; Gzgzyan, A.M.; Kogan, I.Y.; Chiryaeva, O.G.; et al. Telomere Length in Metaphase Chromosomes of Human Triploid Zygotes. Int. J. Mol. Sci. 2021, 22, 5579. [Google Scholar] [CrossRef]
- Huleyuk, N.; Tkach, I.; Zastavna, D.; Tyrka, M. Can telomere shortening be the main indicator of non-viable fetus elimination? Mol. Cytogenet. 2018, 11, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaziri, H.; Schächter, F.; Uchida, I.; Wei, L.; Zhu, X.; Effros, R.; Cohen, D.; Harley, C.B. Loss of telomeric DNA during aging of normal and trisomy 21 human lymphocytes. Am. J. Hum. Genet. 1993, 52, 661–667. [Google Scholar] [PubMed]
- Bhaumik, P.; Bhattacharya, M.; Ghosh, P.; Ghosh, S.; Dey, S.K. Telomere length analysis in Down syndrome birth. Mech. Ageing Dev. 2017, 164, 20–26. [Google Scholar] [CrossRef]
- Pendina, A.; Efimova, O.A.; Chiryaeva, O.G.; Tikhonov, A.V.; Petrova, L.; Dudkina, V.; Sadik, N.A.; Fedorova, I.D.; Galembo, I.A.; Kuznetzova, T.V.; et al. A comparative cytogenetic study of miscarriages after IVF and natural conception in women aged under and over 35 years. J. Assist. Reprod. Genet. 2014, 31, 149–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pendina, A.A.; Efimova, O.A.; Tikhonov, A.V.; Chiryaeva, O.G.; Fedorova, I.D.; Koltsova, A.S.; Krapivin, M.I.; Parfenyev, S.E.; Kuznetzova, T.V.; Baranov, V.S. Immunofluorescence Staining for Cytosine Modifications Like 5-Methylcytosine and Its Oxidative Derivatives and FISH. In FlImmuorescenceity in Insiectu Hybridization (FISH); Liehr, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 337–346. [Google Scholar]
- Efimova, O.A.; Pendina, A.; Tikhonov, A.V.; Fedorova, I.D.; Krapivin, M.I.; Chiryaeva, O.G.; Shilnikova, E.M.; Bogdanova, M.A.; Kogan, I.Y.; Kuznetzova, T.V.; et al. Chromosome hydroxymethylation patterns in human zygotes and cleavage-stage embryos. Reproduction 2015, 149, 223–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efimova, O.A.; Pendina, A.A.; Tikhonov, A.V.; Parfenyev, S.E.; Mekina, I.D.; Komarova, E.M.; Mazilina, M.A.; Daev, E.V.; Chiryaeva, O.G.; Galembo, I.A.; et al. Genome-wide 5-hydroxymethylcytosine patterns in human spermatogenesis are associated with semen quality. Oncotarget 2017, 8, 88294–88307. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krapivin, M.I.; Tikhonov, A.V.; Efimova, O.A.; Pendina, A.A.; Smirnova, A.A.; Chiryaeva, O.G.; Talantova, O.E.; Petrova, L.I.; Dudkina, V.S.; Baranov, V.S. Telomere Length in Chromosomally Normal and Abnormal Miscarriages and Ongoing Pregnancies and Its Association with 5-hydroxymethylcytosine Patterns. Int. J. Mol. Sci. 2021, 22, 6622. https://doi.org/10.3390/ijms22126622
Krapivin MI, Tikhonov AV, Efimova OA, Pendina AA, Smirnova AA, Chiryaeva OG, Talantova OE, Petrova LI, Dudkina VS, Baranov VS. Telomere Length in Chromosomally Normal and Abnormal Miscarriages and Ongoing Pregnancies and Its Association with 5-hydroxymethylcytosine Patterns. International Journal of Molecular Sciences. 2021; 22(12):6622. https://doi.org/10.3390/ijms22126622
Chicago/Turabian StyleKrapivin, Mikhail I., Andrei V. Tikhonov, Olga A. Efimova, Anna A. Pendina, Anna A. Smirnova, Olga G. Chiryaeva, Olga E. Talantova, Lubov’ I. Petrova, Vera S. Dudkina, and Vladislav S. Baranov. 2021. "Telomere Length in Chromosomally Normal and Abnormal Miscarriages and Ongoing Pregnancies and Its Association with 5-hydroxymethylcytosine Patterns" International Journal of Molecular Sciences 22, no. 12: 6622. https://doi.org/10.3390/ijms22126622
APA StyleKrapivin, M. I., Tikhonov, A. V., Efimova, O. A., Pendina, A. A., Smirnova, A. A., Chiryaeva, O. G., Talantova, O. E., Petrova, L. I., Dudkina, V. S., & Baranov, V. S. (2021). Telomere Length in Chromosomally Normal and Abnormal Miscarriages and Ongoing Pregnancies and Its Association with 5-hydroxymethylcytosine Patterns. International Journal of Molecular Sciences, 22(12), 6622. https://doi.org/10.3390/ijms22126622