
Diet-Regulating Microbiota and Host Immune System in Liver Disease

 ,
,  , , , , ,
, , , , ,  and
and
Abstract
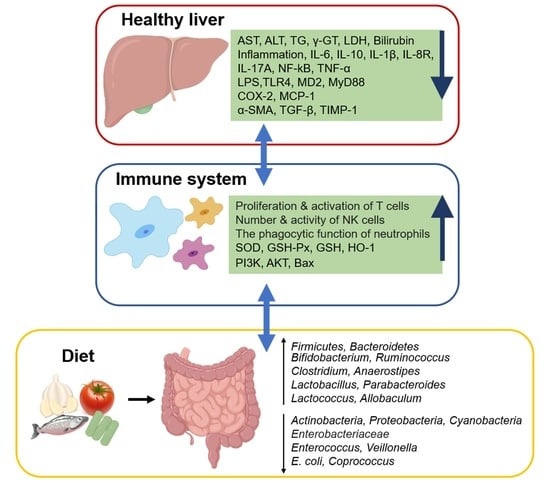
1. Introduction
2. Gut-Liver Axis
3. Diet, Microbiota, and Immune Responses in Chronic Liver Disease
3.1. Nonalcoholic Liver Disease
3.2. Alcoholic Liver Disease
3.3. Liver Cirrhosis
3.4. Hepatocellular Carcinoma
3.5. Diet, the Immune System and Liver Disease
4. Perspective
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Son, S.W.; Song, D.S.; Chang, U.I.; Yang, J.M. Definition of Sarcopenia in Chronic Liver Disease. Life 2021, 11, 349. [Google Scholar] [CrossRef] [PubMed]
- Moon, A.M.; Singal, A.G.; Tapper, E.B. Contemporary Epidemiology of Chronic Liver Disease and Cirrhosis. Clin. Gastroenterol. Hepatol. 2020, 18, 2650–2666. [Google Scholar] [CrossRef]
- Jiang, W.; Wu, N.; Wang, X.; Chi, Y.; Zhang, Y.; Qiu, X.; Hu, Y.; Li, J.; Liu, Y. Dysbiosis gut microbiota associated with inflammation and impaired mucosal immune function in intestine of humans with non-alcoholic fatty liver disease. Sci. Rep. 2015, 5, 8096. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Ye, L.; Li, J.; Jin, L.; Wang, W.; Li, S.; Bao, M.; Wu, S.; Li, L.; Geng, B.; et al. Metagenomic and metabolomic analyses unveil dysbiosis of gut microbiota in chronic heart failure patients. Sci. Rep. 2018, 8, 635. [Google Scholar] [CrossRef] [PubMed]
- Kundu, P.; Blacher, E.; Elinav, E.; Pettersson, S. Our Gut Microbiome: The Evolving Inner Self. Cell 2017, 171, 1481–1493. [Google Scholar] [CrossRef]
- Lindheim, L.; Bashir, M.; Munzker, J.; Trummer, C.; Zachhuber, V.; Leber, B.; Horvath, A.; Pieber, T.R.; Gorkiewicz, G.; Stadlbauer, V.; et al. Alterations in Gut Microbiome Composition and Barrier Function Are Associated with Reproductive and Metabolic Defects in Women with Polycystic Ovary Syndrome (PCOS): A Pilot Study. PLoS ONE 2017, 12, e0168390. [Google Scholar] [CrossRef]
- Alexander, M.; Turnbaugh, P.J. Deconstructing Mechanisms of Diet-Microbiome-Immune Interactions. Immunity 2020, 53, 264–276. [Google Scholar] [CrossRef]
- Honda, K.; Littman, D.R. The microbiota in adaptive immune homeostasis and disease. Nature 2016, 535, 75–84. [Google Scholar] [CrossRef]
- Acharya, C.; Sahingur, S.E.; Bajaj, J.S. Microbiota, cirrhosis, and the emerging oral-gut-liver axis. JCI Insight 2017, 2. [Google Scholar] [CrossRef]
- Tap, J.; Mondot, S.; Levenez, F.; Pelletier, E.; Caron, C.; Furet, J.P.; Ugarte, E.; Munoz-Tamayo, R.; Paslier, D.L.; Nalin, R.; et al. Towards the human intestinal microbiota phylogenetic core. Environ. Microbiol. 2009, 11, 2574–2584. [Google Scholar] [CrossRef]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Nageshwar Reddy, D. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787–8803. [Google Scholar] [CrossRef]
- Milosevic, I.; Vujovic, A.; Barac, A.; Djelic, M.; Korac, M.; Radovanovic Spurnic, A.; Gmizic, I.; Stevanovic, O.; Djordjevic, V.; Lekic, N.; et al. Gut-Liver Axis, Gut Microbiota, and Its Modulation in the Management of Liver Diseases: A Review of the Literature. Int. J. Mol. Sci. 2019, 20, 395. [Google Scholar] [CrossRef] [PubMed]
- Fukui, H. Role of Gut Dysbiosis in Liver Diseases: What Have We Learned So Far? Diseases 2019, 7, 58. [Google Scholar] [CrossRef] [PubMed]
- Arab, J.P.; Martin-Mateos, R.M.; Shah, V.H. Gut-liver axis, cirrhosis and portal hypertension: The chicken and the egg. Hepatol. Int. 2018, 12, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Fukui, H. Gut-liver axis in liver cirrhosis: How to manage leaky gut and endotoxemia. World J. Hepatol. 2015, 7, 425–442. [Google Scholar] [CrossRef]
- Miura, K.; Ohnishi, H. Role of gut microbiota and Toll-like receptors in nonalcoholic fatty liver disease. World J. Gastroenterol. 2014, 20, 7381–7391. [Google Scholar] [CrossRef]
- Fukui, H. Leaky Gut and Gut-Liver Axis in Liver Cirrhosis: Clinical Studies Update. Gut Liver 2020. [Google Scholar] [CrossRef]
- Chiang, J.Y.L.; Ferrell, J.M. Bile Acid Metabolism in Liver Pathobiology. Gene Expr. 2018, 18, 71–87. [Google Scholar] [CrossRef]
- Jennison, E.; Byrne, C.D. The role of the gut microbiome and diet in the pathogenesis of non-alcoholic fatty liver disease. Clin. Mol. Hepatol. 2021, 27, 22–43. [Google Scholar] [CrossRef]
- Liu, L.; Li, P.; Liu, Y.; Zhang, Y. Efficacy of Probiotics and Synbiotics in Patients with Nonalcoholic Fatty Liver Disease: A Meta-Analysis. Dig. Dis. Sci. 2019, 64, 3402–3412. [Google Scholar] [CrossRef]
- Mokhtari, Z.; Gibson, D.L.; Hekmatdoost, A. Nonalcoholic Fatty Liver Disease, the Gut Microbiome, and Diet. Adv. Nutr. 2017, 8, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Ding, Z.; Ishaq, M.; Bacha, A.S.; Khan, I.; Hanif, A.; Li, W.; Guo, X. Understanding the Effects of Gut Microbiota Dysbiosis on Nonalcoholic Fatty Liver Disease and the Possible Probiotics Role: Recent Updates. Int. J. Biol. Sci. 2021, 17, 818–833. [Google Scholar] [CrossRef] [PubMed]
- Koopman, N.; Molinaro, A.; Nieuwdorp, M.; Holleboom, A.G. Review article: Can bugs be drugs? The potential of probiotics and prebiotics as treatment for non-alcoholic fatty liver disease. Aliment. Pharmacol. Ther. 2019, 50, 628–639. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.E.; Parnell, J.A.; Eksteen, B.; Raman, M.; Bomhof, M.R.; Rioux, K.P.; Madsen, K.L.; Reimer, R.A. Gut microbiota manipulation with prebiotics in patients with non-alcoholic fatty liver disease: A randomized controlled trial protocol. BMC Gastroenterol. 2015, 15, 169. [Google Scholar] [CrossRef] [PubMed]
- Mofidi, F.; Poustchi, H.; Yari, Z.; Nourinayyer, B.; Merat, S.; Sharafkhah, M.; Malekzadeh, R.; Hekmatdoost, A. Synbiotic supplementation in lean patients with non-alcoholic fatty liver disease: A pilot, randomised, double-blind, placebo-controlled, clinical trial. Br. J. Nutr. 2017, 117, 662–668. [Google Scholar] [CrossRef]
- Younossi, Z.; Tacke, F.; Arrese, M.; Chander Sharma, B.; Mostafa, I.; Bugianesi, E.; Wai-Sun Wong, V.; Yilmaz, Y.; George, J.; Fan, J.; et al. Global Perspectives on Nonalcoholic Fatty Liver Disease and Nonalcoholic Steatohepatitis. Hepatology 2019, 69, 2672–2682. [Google Scholar] [CrossRef]
- Brunt, E.M.; Wong, V.W.; Nobili, V.; Day, C.P.; Sookoian, S.; Maher, J.J.; Bugianesi, E.; Sirlin, C.B.; Neuschwander-Tetri, B.A.; Rinella, M.E. Nonalcoholic fatty liver disease. Nat. Rev. Dis. Primers 2015, 1, 15080. [Google Scholar] [CrossRef]
- Finelli, C.; Tarantino, G. Non-alcoholic fatty liver disease, diet and gut microbiota. EXCLI J. 2014, 13, 461–490. [Google Scholar]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef]
- Eslamparast, T.; Eghtesad, S.; Hekmatdoost, A.; Poustchi, H. Probiotics and Nonalcoholic Fatty liver Disease. Middle East J. Dig. Dis. 2013, 5, 129–136. [Google Scholar]
- Meroni, M.; Longo, M.; Dongiovanni, P. The role of probiotics in nonalcoholic fatty liver disease: A new insight into therapeutic strategies. Nutrients 2019, 11, 2642. [Google Scholar] [CrossRef]
- Kopec, K.L.; Burns, D. Nonalcoholic fatty liver disease: A review of the spectrum of disease, diagnosis, and therapy. Nutr. Clin. Pract. 2011, 26, 565–576. [Google Scholar] [CrossRef]
- Xue, L.; He, J.; Gao, N.; Lu, X.; Li, M.; Wu, X.; Liu, Z.; Jin, Y.; Liu, J.; Xu, J. Probiotics may delay the progression of nonalcoholic fatty liver disease by restoring the gut microbiota structure and improving intestinal endotoxemia. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Ritze, Y.; Bardos, G.; Claus, A.; Ehrmann, V.; Bergheim, I.; Schwiertz, A.; Bischoff, S.C. Lactobacillus rhamnosus GG protects against non-alcoholic fatty liver disease in mice. PLoS ONE 2014, 9, e80169. [Google Scholar] [CrossRef]
- Zhao, Z.; Chen, L.; Zhao, Y.; Wang, C.; Duan, C.; Yang, G.; Niu, C.; Li, S. Lactobacillus plantarum NA136 ameliorates nonalcoholic fatty liver disease by modulating gut microbiota, improving intestinal barrier integrity, and attenuating inflammation. Appl. Microbiol. Biotechnol. 2020, 104, 5273–5282. [Google Scholar] [CrossRef]
- Liang, Y.; Liang, S.; Zhang, Y.; Deng, Y.; He, Y.; Chen, Y.; Liu, C.; Lin, C.; Yang, Q. Oral Administration of Compound Probiotics Ameliorates HFD-Induced Gut Microbe Dysbiosis and Chronic Metabolic Inflammation via the G Protein-Coupled Receptor 43 in Non-alcoholic Fatty Liver Disease Rats. Probiotics Antimicrob. Proteins 2019, 11, 175–185. [Google Scholar] [CrossRef]
- Wang, W.; Xu, A.L.; Li, Z.C.; Li, Y.; Xu, S.F.; Sang, H.C.; Zhi, F. Combination of Probiotics and Salvia miltiorrhiza Polysaccharide Alleviates Hepatic Steatosis via Gut Microbiota Modulation and Insulin Resistance Improvement in High Fat-Induced NAFLD Mice. Diabetes Metab. J. 2020, 44, 336–348. [Google Scholar] [CrossRef]
- Cano, P.G.; Santacruz, A.; Trejo, F.M.; Sanz, Y. Bifidobacterium CECT 7765 improves metabolic and immunological alterations associated with obesity in high-fat diet-fed mice. Obesity 2013, 21, 2310–2321. [Google Scholar] [CrossRef] [PubMed]
- Baeck, C.; Wehr, A.; Karlmark, K.R.; Heymann, F.; Vucur, M.; Gassler, N.; Huss, S.; Klussmann, S.; Eulberg, D.; Luedde, T.; et al. Pharmacological inhibition of the chemokine CCL2 (MCP-1) diminishes liver macrophage infiltration and steatohepatitis in chronic hepatic injury. Gut 2012, 61, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Raso, G.M.; Simeoli, R.; Iacono, A.; Santoro, A.; Amero, P.; Paciello, O.; Russo, R.; D’Agostino, G.; Di Costanzo, M.; Canani, R.B.; et al. Effects of a Lactobacillus paracasei B21060 based synbiotic on steatosis, insulin signaling and toll-like receptor expression in rats fed a high-fat diet. J. Nutr. Biochem. 2014, 25, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.B.; Jun, D.W.; Kang, B.K.; Lim, J.H.; Lim, S.; Chung, M.J. Randomized, Double-blind, Placebo-controlled Study of a Multispecies Probiotic Mixture in Nonalcoholic Fatty Liver Disease. Sci. Rep. 2019, 9, 5688. [Google Scholar] [CrossRef]
- Bomhof, M.R.; Parnell, J.A.; Ramay, H.R.; Crotty, P.; Rioux, K.P.; Probert, C.S.; Jayakumar, S.; Raman, M.; Reimer, R.A. Histological improvement of non-alcoholic steatohepatitis with a prebiotic: A pilot clinical trial. Eur. J. Nutr. 2019, 58, 1735–1745. [Google Scholar] [CrossRef]
- Gao, B.; Bataller, R. Alcoholic liver disease: Pathogenesis and new therapeutic targets. Gastroenterology 2011, 141, 1572–1585. [Google Scholar] [CrossRef]
- Szabo, G. Gut-liver axis in alcoholic liver disease. Gastroenterology 2015, 148, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Lin, Z.; Zeng, Y.; Lin, X.; Zhang, Y. Probiotic and glutamine treatments attenuate alcoholic liver disease in a rat model. Exp. Ther. Med. 2019, 18, 4733–4739. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Zhong, W. Targeting the gut barrier for the treatment of alcoholic liver disease. Liver Res. 2017, 1, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Sarin, S.K.; Pande, A.; Schnabl, B. Microbiome as a therapeutic target in alcohol-related liver disease. J. Hepatol. 2019, 70, 260–272. [Google Scholar] [CrossRef]
- Albillos, A.; de Gottardi, A.; Rescigno, M. The gut-liver axis in liver disease: Pathophysiological basis for therapy. J. Hepatol. 2020, 72, 558–577. [Google Scholar] [CrossRef]
- Meroni, M.; Longo, M.; Dongiovanni, P. Alcohol or Gut Microbiota: Who Is the Guilty? Int. J. Mol. Sci. 2019, 20, 4568. [Google Scholar] [CrossRef]
- Ceni, E.; Mello, T.; Galli, A. Pathogenesis of alcoholic liver disease: Role of oxidative metabolism. World J. Gastroenterol. 2014, 20, 17756–17772. [Google Scholar] [CrossRef]
- Ghosh Dastidar, S.; Warner, J.B.; Warner, D.R.; McClain, C.J.; Kirpich, I.A. Rodent Models of Alcoholic Liver Disease: Role of Binge Ethanol Administration. Biomolecules 2018, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.; Kim, S.W.; Han, S.H.; Kim, D.J.; Suk, K.T.; Kim, Y.S.; Kim, M.J.; Kim, M.Y.; Baik, S.K.; Ham, Y.L. Probiotics (Lactobacillus rhamnosus R0011 and acidophilus R0052) reduce the expression of toll-like receptor 4 in mice with alcoholic liver disease. PLoS ONE 2015, 10, e0117451. [Google Scholar] [CrossRef] [PubMed]
- Kawaratani, H.; Moriya, K.; Namisaki, T.; Uejima, M.; Kitade, M.; Takeda, K.; Okura, Y.; Kaji, K.; Takaya, H.; Nishimura, N.; et al. Therapeutic strategies for alcoholic liver disease: Focusing on inflammation and fibrosis (Review). Int. J. Mol. Med. 2017, 40, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Shoaib, M.; Shehzad, A.; Omar, M.; Rakha, A.; Raza, H.; Sharif, H.R.; Shakeel, A.; Ansari, A.; Niazi, S. Inulin: Properties, health benefits and food applications. Carbohydr. Polym. 2016, 147, 444–454. [Google Scholar] [CrossRef]
- Yang, X.; He, F.; Zhang, Y.; Xue, J.; Li, K.; Zhang, X.; Zhu, L.; Wang, Z.; Wang, H.; Yang, S. Inulin Ameliorates Alcoholic Liver Disease via Suppressing LPS-TLR4-Mpsi Axis and Modulating Gut Microbiota in Mice. Alcohol. Clin. Exp. Res. 2019, 43, 411–424. [Google Scholar] [CrossRef]
- Chiu, W.C.; Huang, Y.L.; Chen, Y.L.; Peng, H.C.; Liao, W.H.; Chuang, H.L.; Chen, J.R.; Yang, S.C. Synbiotics reduce ethanol-induced hepatic steatosis and inflammation by improving intestinal permeability and microbiota in rats. Food Funct. 2015, 6, 1692–1700. [Google Scholar] [CrossRef]
- Kaviarasan, S.; Sundarapandiyan, R.; Anuradha, C.V. Epigallocatechin gallate, a green tea phytochemical, attenuates alcohol-induced hepatic protein and lipid damage. Toxicol. Mech. Methods 2008, 18, 645–652. [Google Scholar] [CrossRef]
- Rishi, P.; Arora, S.; Kaur, U.J.; Chopra, K.; Kaur, I.P. Better Management of Alcohol Liver Disease Using a ’Microstructured Synbox’ System Comprising L. plantarum and EGCG. PLoS ONE 2017, 12, e0168459. [Google Scholar] [CrossRef]
- Kirpich, I.A.; Solovieva, N.V.; Leikhter, S.N.; Shidakova, N.A.; Lebedeva, O.V.; Sidorov, P.I.; Bazhukova, T.A.; Soloviev, A.G.; Barve, S.S.; McClain, C.J.; et al. Probiotics restore bowel flora and improve liver enzymes in human alcohol-induced liver injury: A pilot study. Alcohol 2008, 42, 675–682. [Google Scholar] [CrossRef]
- Grabherr, F.; Grander, C.; Effenberger, M.; Adolph, T.E.; Tilg, H. Gut Dysfunction and Non-alcoholic Fatty Liver Disease. Front. Endocrinol. 2019, 10, 611. [Google Scholar] [CrossRef]
- Han, S.H.; Suk, K.T.; Kim, D.J.; Kim, M.Y.; Baik, S.K.; Kim, Y.D.; Cheon, G.J.; Choi, D.H.; Ham, Y.L.; Shin, D.H.; et al. Effects of probiotics (cultured Lactobacillus subtilis/Streptococcus faecium) in the treatment of alcoholic hepatitis: Randomized-controlled multicenter study. Eur. J. Gastroenterol. Hepatol. 2015, 27, 1300–1306. [Google Scholar] [CrossRef] [PubMed]
- Schuppan, D.; Afdhal, N.H. Liver Cirrhosis. Lancet 2008, 371, 838–851. [Google Scholar] [CrossRef]
- Natarajan, S.K.; Thomas, S.; Ramamoorthy, P.; Basivireddy, J.; Pulimood, A.B.; Ramachandran, A.; Balasubramanian, K.A. Oxidative stress in the development of liver cirrhosis: A comparison of two different experimental models. J. Gastroenterol. Hepatol. 2006, 21, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Masarone, M.; Rosato, V.; Dallio, M.; Gravina, A.G.; Aglitti, A.; Loguercio, C.; Federico, A.; Persico, M. Role of Oxidative Stress in Pathophysiology of Nonalcoholic Fatty Liver Disease. Oxid. Med. Cell Longev. 2018, 2018, 9547613. [Google Scholar] [CrossRef]
- Li, S.; Zheng, X.; Zhang, X.; Yu, H.; Han, B.; Lv, Y.; Liu, Y.; Wang, X.; Zhang, Z. Exploring the liver fibrosis induced by deltamethrin exposure in quails and elucidating the protective mechanism of resveratrol. Ecotoxicol. Environ. Saf. 2021, 207, 111501. [Google Scholar] [CrossRef]
- Tsuchida, T.; Friedman, S.L. Mechanisms of hepatic stellate cell activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef]
- Arriazu, E.; Ruiz de Galarreta, M.; Cubero, F.J.; Varela-Rey, M.; Perez de Obanos, M.P.; Leung, T.M.; Lopategi, A.; Benedicto, A.; Abraham-Enachescu, I.; Nieto, N. Extracellular matrix and liver disease. Antioxid. Redox. Signal 2014, 21, 1078–1097. [Google Scholar] [CrossRef]
- Roehlen, N.; Crouchet, E.; Baumert, T.F. Liver Fibrosis: Mechanistic Concepts and Therapeutic Perspectives. Cells 2020, 9, 875. [Google Scholar] [CrossRef]
- Iwaisako, K.; Jiang, C.; Zhang, M.; Cong, M.; Moore-Morris, T.J.; Park, T.J.; Liu, X.; Xu, J.; Wang, P.; Paik, Y.H.; et al. Origin of myofibroblasts in the fibrotic liver in mice. Proc. Natl. Acad. Sci. USA 2014, 111, E3297–E3305. [Google Scholar] [CrossRef]
- Cho1, I.; Blaser, M.J. The Human Microbiome: At the interface of health and disease. Nat. Rev. Genet. 2012, 13, 260–270. [Google Scholar] [CrossRef]
- Mogler, C.; Wieland, M.; Konig, C.; Hu, J.; Runge, A.; Korn, C.; Besemfelder, E.; Breitkopf-Heinlein, K.; Komljenovic, D.; Dooley, S.; et al. Hepatic stellate cell-expressed endosialin balances fibrogenesis and hepatocyte proliferation during liver damage. EMBO Mol. Med. 2015, 7, 332–338. [Google Scholar] [CrossRef]
- Roman, E.; Nieto, J.C.; Gely, C.; Vidal, S.; Pozuelo, M.; Poca, M.; Juarez, C.; Guarner, C.; Manichanh, C.; Soriano, G. Effect of a Multistrain Probiotic on Cognitive Function and Risk of Falls in Patients With Cirrhosis: A Randomized Trial. Hepatol. Commun. 2019, 3, 632–645. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, Q.; Ye, G.; Khan, A.; Liu, J.; Gan, F.; Zhang, X.; Kumbhar, S.; Huang, K. Protective effects of Selenium-enriched probiotics on carbon tetrachloride-induced liver fibrosis in rats. J. Agric. Food Chem. 2015, 63, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Lighthouse, J.; Naito, Y.; Helmy, A.; Hotten, P.; Fuji, H.; Min, C.H.; Yoshioka, M.; Marotta, F. Endotoxinemia and benzodiazepine-like substances in compensated cirrhotic patients: A randomized study comparing the effect of rifaximine alone and in association with a symbiotic preparation. Hepatol. Res. 2004, 28, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Pan, D.D.; Zhou, J.; Jiang, Y.Z. Protective effect of selenium-enriched Lactobacillus on CCl4-induced liver injury in mice and its possible mechanisms. World J. Gastroenterol. 2005, 11, 5795–5800. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Lv, L.; Fang, D.; Wu, W.; Hu, C.; Xu, L.; Chen, Y.; Guo, J.; Hu, X.; Li, A.; et al. Administration of Lactobacillus salivarius LI01 or Pediococcus pentosaceus LI05 prevents CCl4-induced liver cirrhosis by protecting the intestinal barrier in rats. Sci. Rep. 2017, 7, 6927. [Google Scholar] [CrossRef] [PubMed]
- Tandon, P.; Moncrief, K.; Madsen, K.; Arrieta, M.C.; Owen, R.J.; Bain, V.G.; Wong, W.W.; Ma, M.M. Effects of probiotic therapy on portal pressure in patients with cirrhosis: A pilot study. Liver Int. 2009, 29, 1110–1115. [Google Scholar] [CrossRef] [PubMed]
- McGee, R.G.; Bakens, A.; Wiley, K.; Riordan, S.M.; Webster, A.C. Probiotics for patients with hepatic encephalopathy. Cochrane Database Syst Rev. 2011, 74, 22071855. [Google Scholar] [CrossRef]
- Dhiman, R.K.; Rana, B.; Agrawal, S.; Garg, A.; Chopra, M.; Thumburu, K.K.; Khattri, A.; Malhotra, S.; Duseja, A.; Chawla, Y.K. Probiotic VSL#3 reduces liver disease severity and hospitalization in patients with cirrhosis: A randomized, controlled trial. Gastroenterology 2014, 147, 1327–1337. [Google Scholar] [CrossRef] [PubMed]
- Fouts, D.E.; Torralba, M.; Nelson, K.E.; Brenner, D.A.; Schnabl, B. Bacterial translocation and changes in the intestinal microbiome in mouse models of liver disease. J. Hepatol. 2012, 56, 1283–1292. [Google Scholar] [CrossRef]
- Bajaj, J.S.; Heuman, D.M.; Hylemon, P.B.; Sanyal, A.J.; Puri, P.; Sterling, R.K.; Luketic, V.; Stravitz, R.T.; Siddiqui, M.S.; Fuchs, M.; et al. Randomised clinical trial: Lactobacillus GG modulates gut microbiome, metabolome and endotoxemia in patients with cirrhosis. Aliment. Pharmacol. Ther. 2014, 39, 1113–1125. [Google Scholar] [CrossRef]
- Xia, X.; Chen, J.; Xia, J.; Wang, B.; Liu, H.; Yang, L.; Wang, Y.; Ling, Z. Role of probiotics in the treatment of minimal hepatic encephalopathy in patients with HBV-induced liver cirrhosis. J. Int. Med. Res. 2018, 46, 3596–3604. [Google Scholar] [CrossRef]
- Garcia-Tsao, G.; Wiest, R. Gut microflora in the pathogenesis of the complications of cirrhosis. Best Pract. Res. Clin. Gastroenterol. 2004, 18, 353–372. [Google Scholar] [CrossRef] [PubMed]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.W.; Talati, C.; Kim, R. Hepatocellular carcinoma (HCC): Beyond sorafenib-chemotherapy. J. Gastrointest. Oncol. 2017, 8, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Luedde, T.; Schwabe, R.F. NF-kappaB in the liver--linking injury, fibrosis and hepatocellular carcinoma. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Fattovich, G.; Stroffolini, T.; Zagni, I.; Donato, F. Hepatocellular carcinoma in cirrhosis: Incidence and risk factors. Gastroenterology 2004, 127, S35–S50. [Google Scholar] [CrossRef]
- Sapisochin, G.; Sapisochin, G. Liver transplantation for hepatocellular carcinoma: Outcomes and novel surgical approaches. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 203–217. [Google Scholar] [CrossRef]
- Ren, Z.; Li, A.; Jiang, J.; Zhou, L.; Yu, Z.; Lu, H.; Xie, H.; Chen, X.; Shao, L.; Zhang, R.; et al. Gut microbiome analysis as a tool towards targeted non-invasive biomarkers for early hepatocellular carcinoma. Gut 2019, 68, 1014–1023. [Google Scholar] [CrossRef]
- Brandi, G.; De Lorenzo, S.; Candela, M.; Pantaleo, M.A.; Bellentani, S.; Tovoli, F.; Saccoccio, G.; Biasco, G. Microbiota, NASH, HCC and the potential role of probiotics. Carcinogenesis 2017, 38, 231–240. [Google Scholar] [CrossRef]
- Chassaing, B.; Etienne-Mesmin, L.; Gewirtz, A.T. Microbiota-liver axis in hepatic disease. Hepatology 2014, 59, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Wang, N.; Qin, W. Gut Microbiota and Hepatocellular Carcinoma. Gastrointest. Tumors 2015, 2, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sung, C.Y.; Lee, N.; Ni, Y.; Pihlajamaki, J.; Panagiotou, G.; El-Nezami, H. Probiotics modulated gut microbiota suppresses hepatocellular carcinoma growth in mice. Proc. Natl. Acad. Sci. USA 2016, 113, E1306–E1315. [Google Scholar] [CrossRef]
- Tolba, R.; Kraus, T.; Liedtke, C.; Schwarz, M.; Weiskirchen, R. Diethylnitrosamine (DEN)-induced carcinogenic liver injury in mice. Lab. Anim. 2015, 49, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.L.; Yu, L.X.; Yang, W.; Tang, L.; Lin, Y.; Wu, H.; Zhai, B.; Tan, Y.X.; Shan, L.; Liu, Q.; et al. Profound impact of gut homeostasis on chemically-induced pro-tumorigenic inflammation and hepatocarcinogenesis in rats. J. Hepatol. 2012, 57, 803–812. [Google Scholar] [CrossRef]
- Bindels, L.B.; Porporato, P.; Dewulf, E.M.; Verrax, J.; Neyrinck, A.M.; Martin, J.C.; Scott, K.P.; Buc Calderon, P.; Feron, O.; Muccioli, G.G.; et al. Gut microbiota-derived propionate reduces cancer cell proliferation in the liver. Br. J. Cancer 2012, 107, 1337–1344. [Google Scholar] [CrossRef]
- Huang, J.; Li, W.; Liao, W.; Hao, Q.; Tang, D.; Wang, D.; Wang, Y.; Ge, G. Green tea polyphenol epigallocatechin-3-gallate alleviates nonalcoholic fatty liver disease and ameliorates intestinal immunity in mice fed a high-fat diet. Food Funct. 2020, 11, 9924–9935. [Google Scholar] [CrossRef]
- Valinas, M.A.; Lanteri, M.L.; Ten Have, A.; Andreu, A.B. Chlorogenic Acid Biosynthesis Appears Linked with Suberin Production in Potato Tuber (Solanum tuberosum). J. Agric. Food Chem. 2015, 63, 4902–4913. [Google Scholar] [CrossRef]
- Chiang, W.D.; Huang, C.Y.; Paul, C.R.; Lee, Z.Y.; Lin, W.T. Lipolysis stimulating peptides of potato protein hydrolysate effectively suppresses high-fat-diet-induced hepatocyte apoptosis and fibrosis in aging rats. Food Nutr. Res. 2016, 60, 31417. [Google Scholar] [CrossRef]
- Negri, R.; Trinchese, G.; Carbone, F.; Caprio, M.G.; Stanzione, G.; di Scala, C.; Micillo, T.; Perna, F.; Tarotto, L.; Gelzo, M.; et al. Randomised Clinical Trial: Calorie Restriction Regimen with Tomato Juice Supplementation Ameliorates Oxidative Stress and Preserves a Proper Immune Surveillance Modulating Mitochondrial Bioenergetics of T-Lymphocytes in Obese Children Affected by Non-Alcoholic Fatty Liver Disease (NAFLD). J. Clin. Med. 2020, 9, 141. [Google Scholar] [CrossRef]
- Metcalf, R.; James, M.; Mantzioris, E.; Cleland, L. A practical approach to increasing intakes of n-3 polyunsaturated fatty acids: Use of novel foods enriched with n-3 fats. Eur. J. Clin. Nutr. 2003, 57, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Roberts, B.J.; Song, B.J.; Soh, Y.; Park, S.S.; Shoaf, S.E. Ethanol induces CYP2E1 by protein stabilization. Role of ubiquitin conjugation in the rapid degradation of CYP2E1. J. Biol. Chem. 1995, 270, 29632–29635. [Google Scholar] [CrossRef]
- Song, B.J.; Moon, K.H.; Olsson, N.U.; Salem, N., Jr. Prevention of alcoholic fatty liver and mitochondrial dysfunction in the rat by long-chain polyunsaturated fatty acids. J. Hepatol. 2008, 49, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.-L.; Wan, J.-B.; Wang, B.; He, C.-W.; Ma, H.; Li, T.-W.; Kang, J.X. Suppression of acute ethanol-induced hepatic steatosis by docosahexaenoic acid is associated with downregulation of stearoyl-CoA desaturase 1 and inflammatory cytokines. Prostaglandins Leukot. Essent. Fatty Acids 2013, 88, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, M.; Dobrzyn, A.; Sampath, H.; Lee, S.H.; Man, W.C.; Chu, K.; Peters, J.M.; Gonzalez, F.J.; Ntambi, J.M. Reduced adiposity and liver steatosis by stearoyl-CoA desaturase deficiency are independent of peroxisome proliferator-activated receptor-alpha. J. Biol. Chem. 2004, 279, 35017–35024. [Google Scholar] [CrossRef] [PubMed]
- Slebos, D.J.; Ryter, S.W.; Choi, A.M. Heme oxygenase-1 and carbon monoxide in pulmonary medicine. Respir Res. 2003, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, X.-J.; Yan, C.; He, C.; Li, P.; Chen, M.; Su, H.; Wan, J.-B. Preventive effect of α-linolenic acid-rich flaxseed oil against ethanol-induced liver injury is associated with ameliorating gut-derived endotoxin-mediated inflammation in mice. J. Funct. Foods 2016, 23, 532–541. [Google Scholar] [CrossRef]
- Rigotti, P.; Peters, J.C.; Tranberg, K.G.; Fischer, J.E. Effects of amino acid infusions on liver regeneration after partial hepatectomy in the rat. JPEN J. Parenter. Enteral. Nutr. 1986, 10, 17–20. [Google Scholar] [CrossRef]
- Nakamura, I.; Ochiai, K.; Imawari, M. Phagocytic function of neutrophils of patients with decompensated liver cirrhosis is restored by oral supplementation of branched-chain amino acids. Hepatol. Res. 2004, 29, 207–211. [Google Scholar] [CrossRef]
- Lam, V.W.; Poon, R.T. Role of branched-chain amino acids in management of cirrhosis and hepatocellular carcinoma. Hepatol. Res. 2008, 38 (Suppl. 1), S107–S115. [Google Scholar] [CrossRef]
- Percival, S.S. Aged Garlic Extract Modifies Human Immunity. J. Nutr. 2016, 146, 433S–436S. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Saeki, T.; Otani, T.; Suzuki, T.; Shimozuma, K.; Nishino, H.; Fukuda, S.; Morimoto, K. Aged garlic extract prevents a decline of NK cell number and activity in patients with advanced cancer. J. Nutr. 2006, 136, 816S–820S. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Conditions | Treatment | Main Results | Ref | |
|---|---|---|---|---|
| Animal | HSHF diet | B. infantis, L. acidopilus, B. cereus | (↓) E. coli, Enterococcus, TNF-α, IL-18, Serum LPS, TLR4 (↑) Lactobacillus, Bifidobacteria, Bacteroides | [33] |
| High-fructose diet | LGG | (↓): TNF-α, IL-1β, IL-8R (↑): Firmicutes, Bacteroidetes | [34] | |
| HFD/F diet | L. plantarum | (↓): Serum LPS, TNF-α, IL-6, IL-1β, NF-κB P65 (↑): Barnesiella, Parabacteroides, Bifidobacterium, IκB | [35] | |
| HFD diet | Lactobacillus, Bifidobacterium | (↓): Akkermansia, Bacteroides, Prevotella, Veillonella, Coprococcus, Roseburia, LPS, IL-18, IL-1β (↑): Bifidobacterium, Ruminococcus, Clostridium, Anaerostipes | [36] | |
| L. plantarum X, B. bifidum V Polysaccharide (Salvia miltiorrhiza) | (↓): Cyanobacteria, Oscillospira, Alistipes, LPS, TNF-α, IL-6, IL-1β (↑): Bacteroidetes, Firmicutes, Lactobacillus, Parabacteroides | [37] | ||
| B. pseudocatenulatum CECT 7765 | (↓): Enterobacteriaceae, IL-6, MCP-1, IL-10 (↑): Bifidobacterium spp., IL-4, Induction of T-lymphocyte proliferation | [38] | ||
| L. paracasei B21060, Fructo-oligosaccharides, Arabinogalactan | (↓): Gram-negative bacteria, ALT, AST, TNF-a, IL-6, p50 NF-κB, TLR4, CD14 (↑): IκBα | [40] | ||
| Human | NASH patients | Probiotics | (↓): Total cholesterol, TG, TNF-α (↑): Agathobaculum, Dorea (OTU 527923), Dorea (OTU 195044), Blautia, Ruminococcus, Dorea (OTU 470168) | [41] |
| Oligofructose | (↓): Clostridium cluster XI, Clostridium cluster I, LPS, IL-6, TNF-α (↑): Bifidobacterium spp., C. leptum, F. prausnitzii | [42] | ||
| Conditions | Treatment | Main Results | Ref | |
|---|---|---|---|---|
| Animal | EtOH-induced liver injury | Glutamine, Golden Bifido (probiotic mixture containing live L. bulgaricus, Bifidobacterium, Streptococcus thermophilus), Medilac S® (probiotic mixture containing live B. subtilis and E. faecium) | (↓): Proteobacteria, Actinobacteria AST, ALT, TG, TNF-α, IL-6 (↑): Firmicutes | [45] |
| High fat diet+ EtOH + LPS | L. rhamnosus R0011, L. acidophilus R0052, KRG (Korea red ginseng), urushiol (Rhus verniciflua Stokes) | (↓): AST, ALT, TNF-α, IL-1β, TLR4 (↑): IL-10 | [52] | |
| EtOH-containing modified Lieber-DeCarli liquid diets | Inulin | (↓): Parasutterella, Plasma LPS Levels, TNF-α, IL-6, IL-17A, IL-10, Macrophages, TLR4 (↑): Allobaculum, Lactobacillus, Lactococcus | [55] | |
| Ethanol by oral gavage | Synbox (L. plantarum + EGCG a phenolic compound) | (↓): NF-kB, TNF-α, IL-12 / p 40, TLR4 MD2, CD14, MyD88, COX-2(↑): L. acidophilus, SOD | [58] | |
| Ethanol liquid diet | Symbiotic (L. acidophilus, L. bulgaricus, B. bifidum, B. longum, Inulin, Vitamin B1, Niacinamide.) | (↓): TNF-α, IL-1β, IL-6 (↑): The numbers of lactobacilli and bifidobacterial, IL-10 | [56] | |
| Human | Adult alcoholics | B. bifidum, L. plantarum 8PA3 | (↓): AST, ALT, GGT, LDH, Bilirubin (↑): Restoration of bowel flora | [59] |
| Alcoholic hepatitis (AH) | L. subtilis, S. faecium or placebo | (↓): E. coli, AST, ALT, γ-GT, ALP, LPS, TNF-α, IL-1β | [61] | |
| Condition | Treatment | Main Results | Ref | |
|---|---|---|---|---|
| Animal | CCl4 | Selenium-enriched lactobacillus | (↓): ALT, AST, Ca2+, Phagocytic index, TNF-α (↑): GSH-Px, SOD | [75] |
| L. acidophilus, S. cerevisiae | (↓): ALT, AST, α-SMA, TGF-β1, TIMP-1, Collagen-TNF-α, IL-6, MCP-1, Bcl-2, Bcl-Xl, GSH-Px (↑): Bax, SOD, GSH | [73] | ||
| P. pentosaceus LI05 | (↓): ALT, AST, Endotoxin, Collagen α1, TIMP-1, iNOS-2, TNF-α, IL-6, IL-17A (↑): ZO-1 | [76] | ||
| Human | HCV-related Child B liver cirrhosis | L.acidophilus L.helveticus Bifidobacteria | (↓): Endotoxin, Ammonia level | [74] |
| Cirrhosis and HVPG >10 mmHg | VSL#3 | (↓): Endotoxin, Plasma renin, Aldosterone (↑): TNF-α, IL-6, IL-8 | [77] | |
| Cirrhotic patients with MHE | LGG | (↓): Ammoniagenic amino acids, TNF-α, IL-13, (↑): Lachnospiraceae, SIP | [81] | |
| Cirrhosis with PHES less than-4 | Mixture of eight strains (DSM 24731, DSM 24732, DSM 24736, DSM 24737, DSM 24733, DSM 24735, DSM 24734, DSM 24730) | (↓): Walking problems, AST, ALT, GGT, Serum bilirubin, CRP, TNF-α, FABP-6 (↑): Mean arterial pressure (mm Hg), MFI | [72] | |
| Cirrhosis with HE | VSL#3 | (↓): CTP score, MELD score, Ammonia, Renin, Aldosterone, TNF-a, IL-1β, IL-6, Indole (↑): SBP | [79] | |
| Conditions | Treatment | Main Results | Ref | |
|---|---|---|---|---|
| Animal | Injection of mouse hepatoma cell line Hepa1-6 | Prohep (LGG, E. coli Nissle 1917, heat inactivated VSL#3 (S. thermophilus, B. breve, B. longum, B. infantis, L. acidophilus, L. plantarum, L. paracasei, L. delbrueckii subsp.)) | (↓): Firmicutes, Proteobacteria, Recruitment of Th17, IL-17 (↑): Bacteroidetes, IL-27, IL-13, IL-10 | [93] |
| Injection of DEN | Probiotic VSL#3 (four Lactobacilli, three Bifidobacteria, and one Streptococcus thermophilus subsp Salivarius) | (↓): E. coli, Atopobium cluster, B. fragilis group, Prevotella, ALT, Plasma LPS, IL-6, Nuclear NF-κB translocation (↑): IL-10 | [95] | |
| Injection of Bcr-Abl-transfected BaF3 cells | Prebiotics (ITF) | (↓): IL-4, IL-6, IL-8, IFN-γ, MCP-1 | [96] | |
| Conditions | Treatment | Main Results | Ref | |
|---|---|---|---|---|
| Animal | High- fat/Western diet | EGCG | (↓): ALT, AST, TNF-α, Genes involved in the differentiation and formation of IgA+ B cells, TLR4, BAFF | [97] |
| HFD diet | APPH | (↓): Number of apoptosis nuclei, Fat accumulation (↑): p-PI3K, p-Akt | [99] | |
| Ethanol feeding for 9 weeks | polyunsaturated fatty acids (PUFA) | (↓): TG, Cholesterol, CYP2E1, NOS, H2O2, 3-ketoacyl-CoA thiolase | [103] | |
| Ethanol every 12 h for 3 administrations | DHA | (↓): TG, SCD-1, IL-6, TNF-α (↑): HO-1 | [104] | |
| Binge liquid ethanol feeding | Flaxseed oil | (↓): AST, ALP, TG, MYD88, LPS, CD14, NF-κB p65 (↑): SOD, ZO-1 | [107] | |
| Protein-free, calorie-rich diet | 10% dextrose + 3% amino acids (35% BCAA) | (↓): Phenylalanine, Tyrosine (↑): 3H-thymidine | [108] | |
| Human | NAFLD patients | lycopene-rich tomato juice | (↓): ALT, AST, TC, LDL, TG, IL-4, MDA (↑): HDL, GSH, T cell activation | [100] |
| Cirrhosis patients | BCAA | (↑): Improve the phagocytic function of neutrophils, Natural killer activity of lymphocytes | [109] | |
| HCC patients Child B and C liver disease | BCAA | (↓): Morbidity rate (↑): Red Blood cell and Serum albumin levels Body weight and performance status | [110] | |
| HCC patients | AGE | (↑): Total cholesterol, HDL, Number and activity of NK cells, | [112] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eom, J.A.; Kwon, G.H.; Kim, N.Y.; Park, E.J.; Won, S.M.; Jeong, J.J.; Raja, G.; Gupta, H.; Asmelash Gebru, Y.; Sharma, S.; et al. Diet-Regulating Microbiota and Host Immune System in Liver Disease. Int. J. Mol. Sci. 2021, 22, 6326. https://doi.org/10.3390/ijms22126326
Eom JA, Kwon GH, Kim NY, Park EJ, Won SM, Jeong JJ, Raja G, Gupta H, Asmelash Gebru Y, Sharma S, et al. Diet-Regulating Microbiota and Host Immune System in Liver Disease. International Journal of Molecular Sciences. 2021; 22(12):6326. https://doi.org/10.3390/ijms22126326
Chicago/Turabian StyleEom, Jung A, Goo Hyun Kwon, Na Yeon Kim, Eun Ju Park, Sung Min Won, Jin Ju Jeong, Ganesan Raja, Haripriya Gupta, Yoseph Asmelash Gebru, Satyapriya Sharma, and et al. 2021. "Diet-Regulating Microbiota and Host Immune System in Liver Disease" International Journal of Molecular Sciences 22, no. 12: 6326. https://doi.org/10.3390/ijms22126326
APA StyleEom, J. A., Kwon, G. H., Kim, N. Y., Park, E. J., Won, S. M., Jeong, J. J., Raja, G., Gupta, H., Asmelash Gebru, Y., Sharma, S., Choi, Y. R., Kim, H. S., Yoon, S. J., Hyun, J. Y., Jeong, M. K., Park, H. J., Min, B. H., Choi, M. R., Kim, D. J., & Suk, K. T. (2021). Diet-Regulating Microbiota and Host Immune System in Liver Disease. International Journal of Molecular Sciences, 22(12), 6326. https://doi.org/10.3390/ijms22126326