
Mirna Expression in Glaucomatous and TGFβ2 Treated Lamina Cribrosa Cells


Abstract
1. Introduction
2. Results
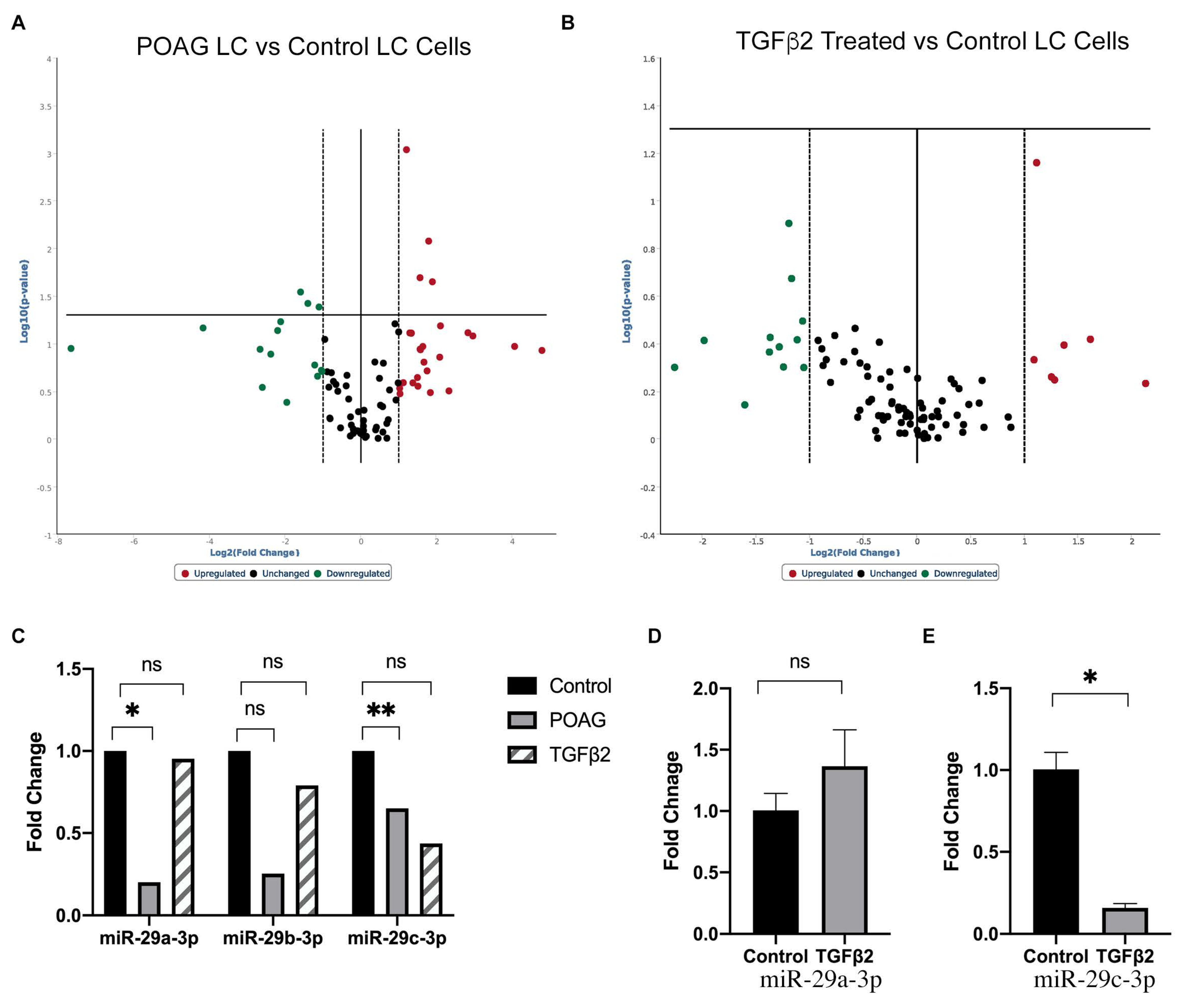
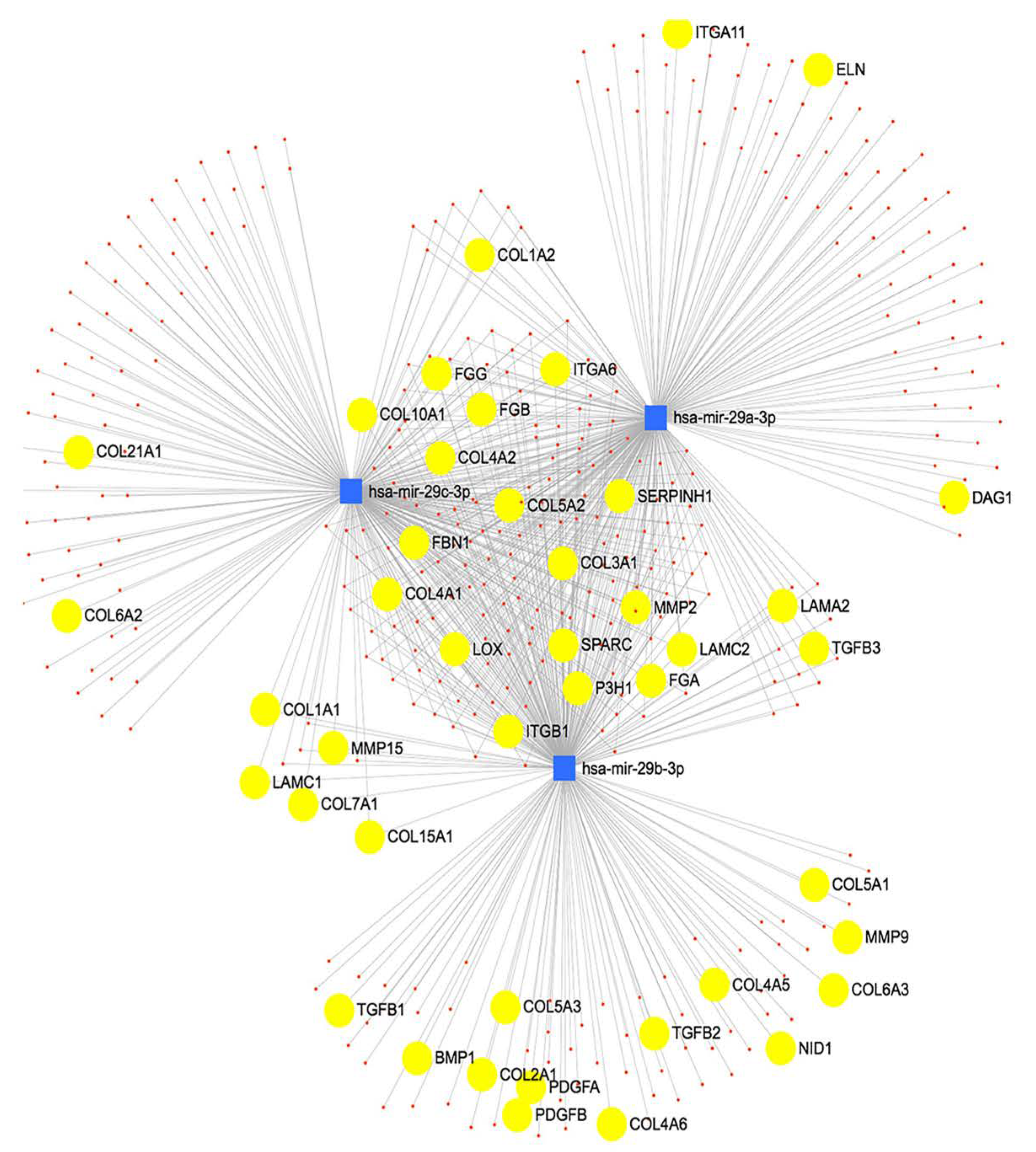
2.1. Differentially Expressed miRNAs in POAG and TGFβ2 Treated LC Cells Compared to Non-Glaucomatous Control LC Cells
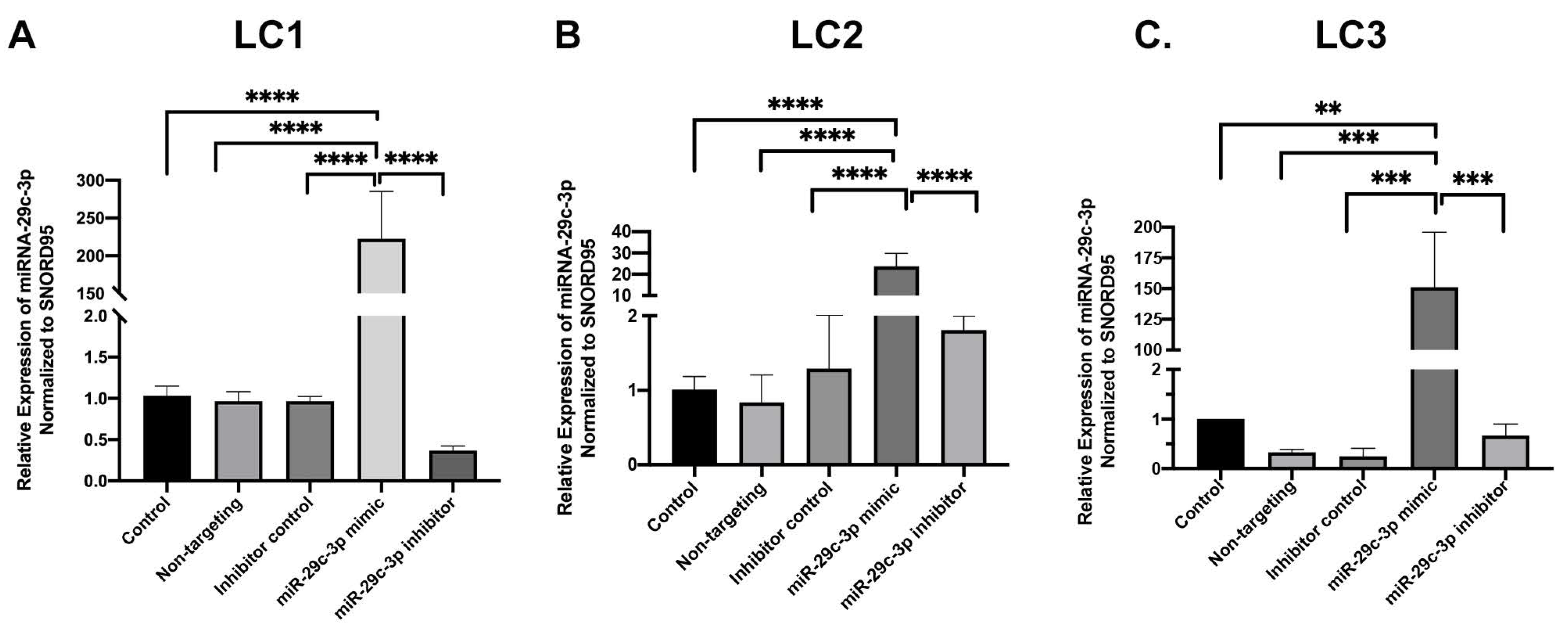
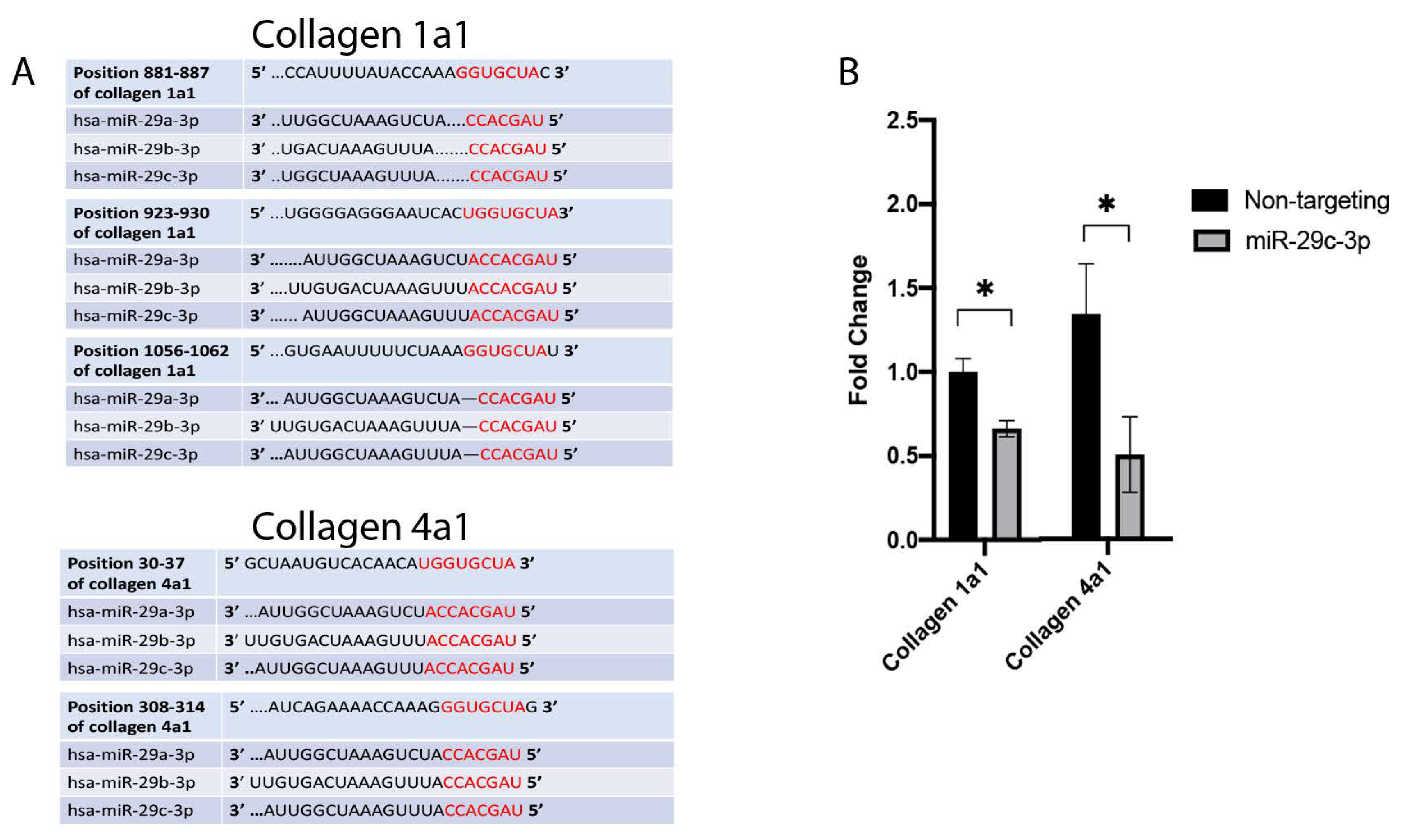
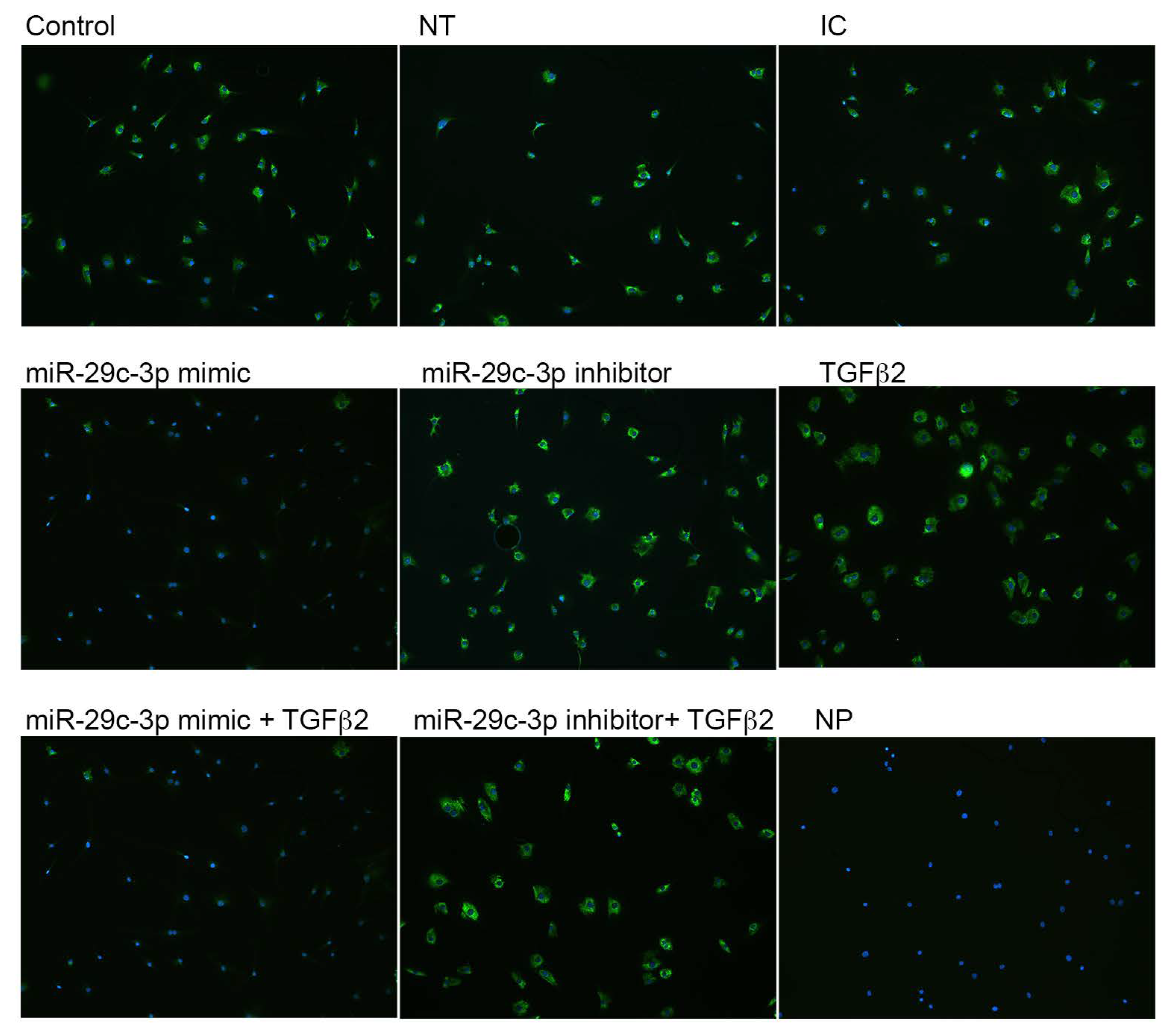
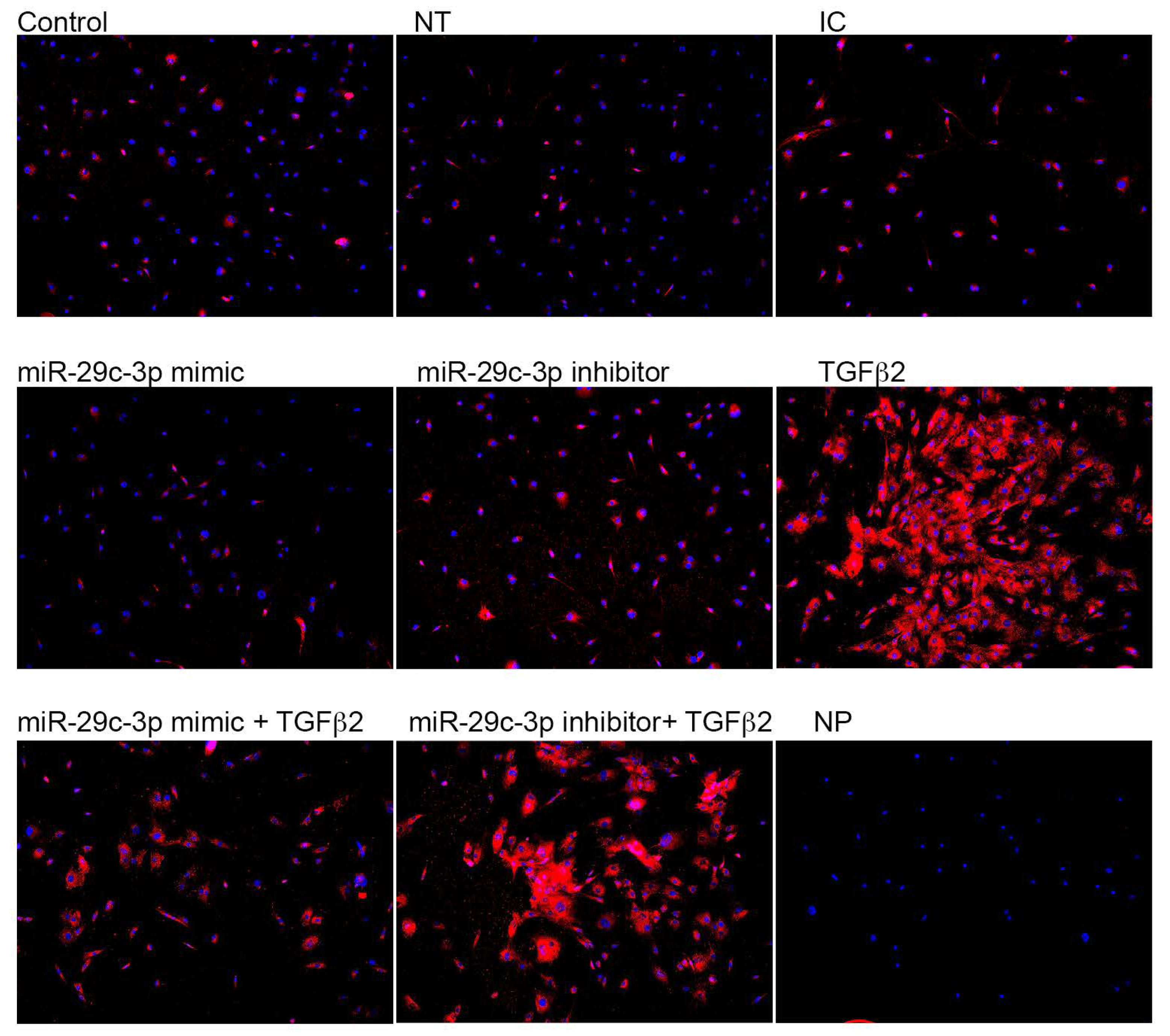
2.2. Effects of miR-29c-3p on the ECM
2.3. Effects of miR-29c on TGFβ2 Induced-ECM Proteins
3. Discussion
4. Materials and Methods
4.1. Cell Culture and TGFβ2 Treatment
4.2. miRNA PCR Array
4.3. In Vitro Transfection of LC Cells Using miRNA Mimics and Inhibitors
4.4. RNA Isolation and qPCR
4.5. Immunocytochemistry
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Armaly, M.F.; Krueger, D.E.; Maunder, L.; Becker, B.; Hetherington, J., Jr.; Kolker, A.E.; Levene, R.Z.; Maumenee, A.E.; Pollack, I.P.; Shaffer, R.N. Biostatistical Analysis of the Collaborative Glaucoma Study: I. Summary Report of the Risk Factors for Glaucomatous Visual-Field Defects. Arch. Ophthalmol. 1980, 98, 2163–2171. [Google Scholar] [CrossRef] [PubMed]
- Sommer, A. Intraocular Pressure and Glaucoma. Am. J. Ophthalmol. 1989, 107, 186–188. [Google Scholar] [CrossRef]
- Drance, S.M.; Schulzer, M.; Thomas, B.; Douglas, G.R. Multivariate Analysis in Glaucoma: Use of Discriminant Analysis in Predicting Glaucomatous Visual Field Damage. Arch. Ophthalmol. 1981, 99, 1019–1022. [Google Scholar] [CrossRef]
- Quigley, H.A.; Hohman, R.M.; Addicks, E.M.; Massof, R.W.; Green, W.R. Morphologic changes in the lamina cribrosa correlated with neural loss in open-angle glaucoma. Am. J. Ophthalmol. 1983, 95, 673–691. [Google Scholar] [CrossRef]
- Hernandez, M.R.; Luo, X.X.; Igoe, F.; Neufeld, A.H. Extracellular matrix of the human lamina cribrosa. Am. J. Ophthalmol. 1987, 104, 567–576. [Google Scholar] [CrossRef]
- Caparas, V.L.; Cintron, C.; Hernandez-Neufeld, M.R. Immunohistochemistry of proteoglycans in human lamina cribrosa. Am. J. Ophthalmol. 1991, 112, 489–495. [Google Scholar] [CrossRef]
- Sawaguchi, S.; Yue, B.Y.; Fukuchi, T.; Iwata, K.; Kaiya, T. Sulfated proteoglycans in the human lamina cribrosa. Investig. Ophthalmol. Vis. Sci. 1992, 33, 2388–2398. [Google Scholar]
- Pena, J.D.; Netland, P.A.; Vidal, I.; Dorr, D.A.; Rasky, A.; Hernandez, M.R. Elastosis of the lamina cribrosa in glaucomatous optic neuropathy. Exp. Eye Res. 1998, 67, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Quigley, H.A.; Brown, A.; Dorman-Pease, M.E. Alterations in elastin of the optic nerve head in human and experimental glaucoma. Br. J. Ophthalmol. 1991, 75, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, M.R. Ultrastructural immunocytochemical analysis of elastin in the human lamina cribrosa. Changes in elastic fibers in primary open-angle glaucoma. Invest. Ophthalmol. Vis. Sci. 1992, 33, 2891–2903. [Google Scholar] [PubMed]
- Hernandez, M.R.; Andrzejewska, W.M.; Neufeld, A.H. Changes in the extracellular matrix of the human optic nerve head in primary open-angle glaucoma. Am. J. Ophthalmol. 1990, 109, 180–188. [Google Scholar] [CrossRef]
- Hernandez, M.R.; Ye, H. Glaucoma: Changes in extracellular matrix in the optic nerve head. Ann. Med. 1993, 25, 309–315. [Google Scholar] [CrossRef]
- Morrison, J.C.; Dorman-Pease, M.E.; Dunkelberger, G.R.; Quigley, H.A. Optic nerve head extracellular matrix in primary optic atrophy and experimental glaucoma. Arch. Ophthalmol. 1990, 108, 1020–1024. [Google Scholar] [CrossRef]
- Belmares, R.; Raychaudhuri, U.; Maansson, S.; Clark, A.F. Histological investigation of human glaucomatous eyes: Extracellular fibrotic changes and galectin 3 expression in the trabecular meshwork and optic nerve head. Clin. Anat. 2018, 31, 1031–1049. [Google Scholar] [CrossRef] [PubMed]
- Kirwan, R.P.; Fenerty, C.H.; Crean, J.; Wordinger, R.J.; Clark, A.F.; O’Brien, C.J. Influence of cyclical mechanical strain on extracellular matrix gene expression in human lamina cribrosa cells in vitro. Mol. Vis. 2005, 11, 798–810. [Google Scholar]
- Lopez, N.N.; Clark, A.F.; Tovar-Vidales, T. Isolation and characterization of human optic nerve head astrocytes and lamina cribrosa cells. Exp. Eye Res. 2020, 197, 108103. [Google Scholar] [CrossRef] [PubMed]
- Tovar-Vidales, T.; Wordinger, R.J.; Clark, A.F. Identification and localization of lamina cribrosa cells in the human optic nerve head. Exp. Eye Res. 2016, 147, 94–97. [Google Scholar] [CrossRef]
- Zode, G.S.; Sethi, A.; Brun-Zinkernagel, A.M.; Chang, I.F.; Clark, A.F.; Wordinger, R.J. Transforming growth factor-beta2 increases extracellular matrix proteins in optic nerve head cells via activation of the Smad signaling pathway. Mol. Vis. 2011, 17, 1745–1758. [Google Scholar]
- Pena, J.D.; Taylor, A.W.; Ricard, C.S.; Vidal, I.; Hernandez, M.R. Transforming growth factor beta isoforms in human optic nerve heads. Br. J. Ophthalmol. 1999, 83, 209–218. [Google Scholar] [CrossRef]
- Rogers, R.; Dharsee, M.; Ackloo, S.; Flanagan, J.G. Proteomics analyses of activated human optic nerve head lamina cribrosa cells following biomechanical strain. Invest. Ophthalmol. Vis. Sci. 2012, 53, 3806–3816. [Google Scholar] [CrossRef]
- Luna, C.; Li, G.; Qiu, J.; Epstein, D.L.; Gonzalez, P. MicroRNA-24 regulates the processing of latent TGFbeta1 during cyclic mechanical stress in human trabecular meshwork cells through direct targeting of FURIN. J. Cell. Physiol. 2011, 226, 1407–1414. [Google Scholar] [CrossRef] [PubMed]
- Luna, C.; Li, G.; Qiu, J.; Epstein, D.L.; Gonzalez, P. Role of miR-29b on the regulation of the extracellular matrix in human trabecular meshwork cells under chronic oxidative stress. Mol. Vis. 2009, 15, 2488–2497. [Google Scholar] [PubMed]
- Davis, B.N.; Hata, A. Regulation of MicroRNA Biogenesis: A miRiad of mechanisms. Cell Commun. Signal. 2009, 7, 18. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, L.; Lu, L.; Jiang, P.; Sun, H.; Wang, H. Inhibition of miR-29 by TGF-beta-Smad3 signaling through dual mechanisms promotes transdifferentiation of mouse myoblasts into myofibroblasts. PLoS ONE 2012, 7, e33766. [Google Scholar] [CrossRef] [PubMed]
- Davis, B.N.; Hilyard, A.C.; Lagna, G.; Hata, A. SMAD proteins control DROSHA-mediated microRNA maturation. Nature 2008, 454, 56–61. [Google Scholar] [CrossRef]
- Sethi, A.; Mao, W.; Wordinger, R.J.; Clark, A.F. Transforming growth factor-beta induces extracellular matrix protein cross-linking lysyl oxidase (LOX) genes in human trabecular meshwork cells. Invest. Ophthalmol. Vis. Sci. 2011, 52, 5240–5250. [Google Scholar] [CrossRef]
- Tovar-Vidales, T.; Fitzgerald, A.M.; Clark, A.F.; Wordinger, R.J. Transforming growth factor-beta2 induces expression of biologically active bone morphogenetic protein-1 in human trabecular meshwork cells. Invest. Ophthalmol. Vis. Sci. 2013, 54, 4741–4748. [Google Scholar] [CrossRef]
- Zhang, Y.; Ghazwani, M.; Li, J.; Sun, M.; Stolz, D.B.; He, F.; Fan, J.; Xie, W.; Li, S. MiR-29b inhibits collagen maturation in hepatic stellate cells through down-regulating the expression of HSP47 and lysyl oxidase. Biochem. Biophys. Res. Commun. 2014, 446, 940–944. [Google Scholar] [CrossRef]
- Fuchshofer, R. The pathogenic role of transforming growth factor-beta2 in glaucomatous damage to the optic nerve head. Exp. Eye Res. 2011, 93, 165–169. [Google Scholar] [CrossRef]
- Suzuki, H.I. MicroRNA Control of TGF-β Signaling. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Han, J.; Yeom, K.H.; Jin, H.; Kim, V.N. Drosha in primary microRNA processing. Cold Spring Harb. Symp. Quant. Biol. 2006, 71, 51–57. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Jiang, J.; Liu, Q.; Yang, L. A high-throughput method to monitor the expression of microRNA precursors. Nucleic Acids Res. 2004, 32, e43. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Lee, E.J.; Jiang, J.; Sarkar, A.; Yang, L.; Elton, T.S.; Chen, C. Real-time PCR quantification of precursor and mature microRNA. Methods 2008, 44, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Sigal, I.A.; Flanagan, J.G.; Tertinegg, I.; Ethier, C.R. Predicted extension, compression and shearing of optic nerve head tissues. Exp. Eye Res. 2007, 85, 312–322. [Google Scholar] [CrossRef]
- Rogers, R.S.; Dharsee, M.; Ackloo, S.; Sivak, J.M.; Flanagan, J.G. Proteomics analyses of human optic nerve head astrocytes following biomechanical strain. Mol. Cell. Proteom. 2012, 11. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Source | Dilution |
|---|---|---|
| Rabbit anti-Collagen I | Abcam | 1:200 |
| Rabbit anti-Collagen IV | Abcam | 1:200 |
| Donkey anti-Rabbit 488 | Invitrogen | 1:500 |
| Donkey anti-Rabbit 594 | Invitrogen | 1:500 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopez, N.N.; Rangan, R.; Clark, A.F.; Tovar-Vidales, T. Mirna Expression in Glaucomatous and TGFβ2 Treated Lamina Cribrosa Cells. Int. J. Mol. Sci. 2021, 22, 6178. https://doi.org/10.3390/ijms22126178
Lopez NN, Rangan R, Clark AF, Tovar-Vidales T. Mirna Expression in Glaucomatous and TGFβ2 Treated Lamina Cribrosa Cells. International Journal of Molecular Sciences. 2021; 22(12):6178. https://doi.org/10.3390/ijms22126178
Chicago/Turabian StyleLopez, Navita N., Rajiv Rangan, Abbot F. Clark, and Tara Tovar-Vidales. 2021. "Mirna Expression in Glaucomatous and TGFβ2 Treated Lamina Cribrosa Cells" International Journal of Molecular Sciences 22, no. 12: 6178. https://doi.org/10.3390/ijms22126178
APA StyleLopez, N. N., Rangan, R., Clark, A. F., & Tovar-Vidales, T. (2021). Mirna Expression in Glaucomatous and TGFβ2 Treated Lamina Cribrosa Cells. International Journal of Molecular Sciences, 22(12), 6178. https://doi.org/10.3390/ijms22126178