
Non-Coding RNAs in Kidney Diseases: The Long and Short of Them

 ,
, 
 ,
, 
 and
and 
Abstract
1. Introduction
2. Role of lncRNAs in Renal Diseases
2.1. Acute Kidney Injury
2.1.1. Sepsis-Induced AKI
2.1.2. Ischemia-Reperfusion

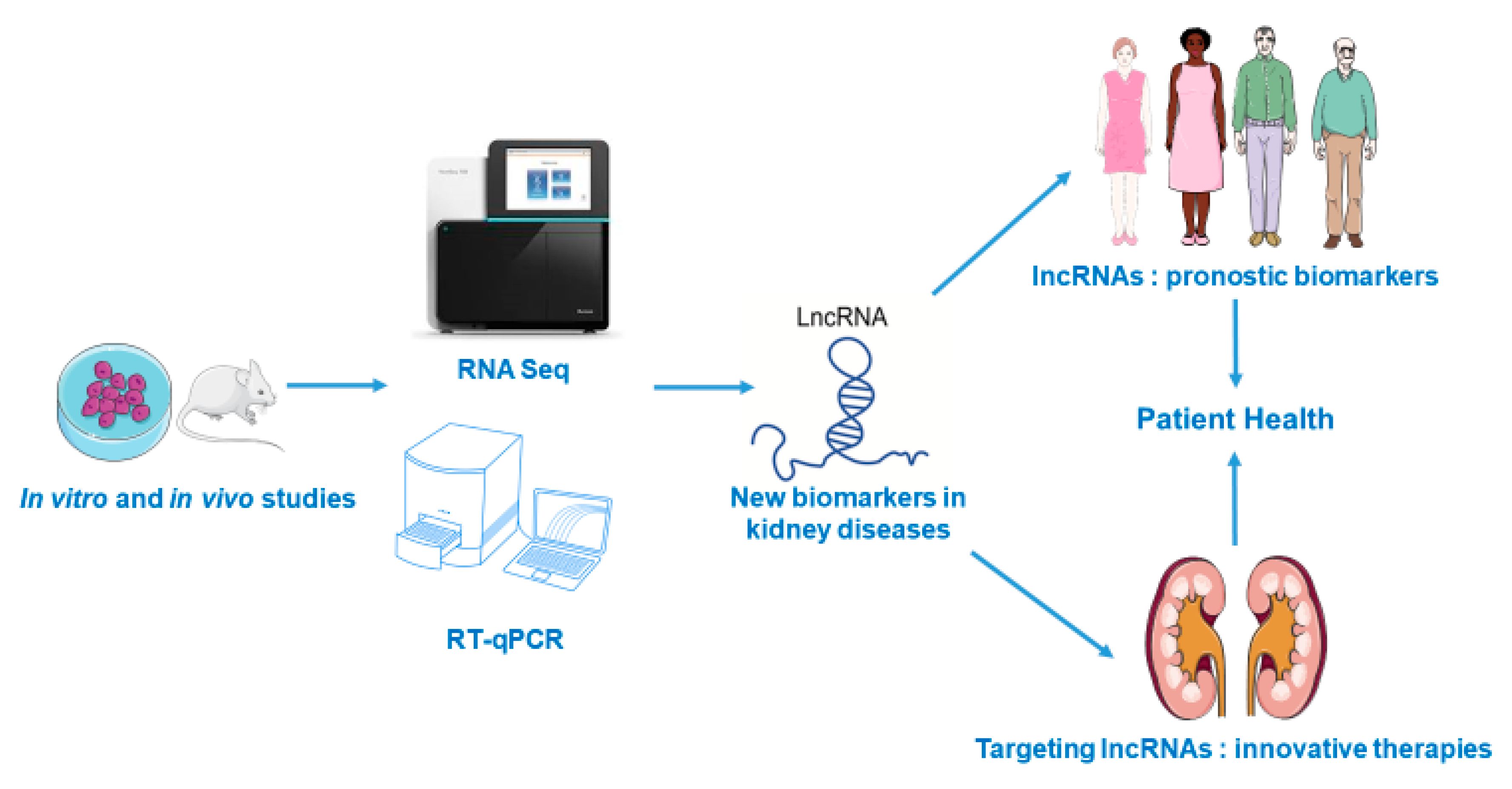{kind=link}
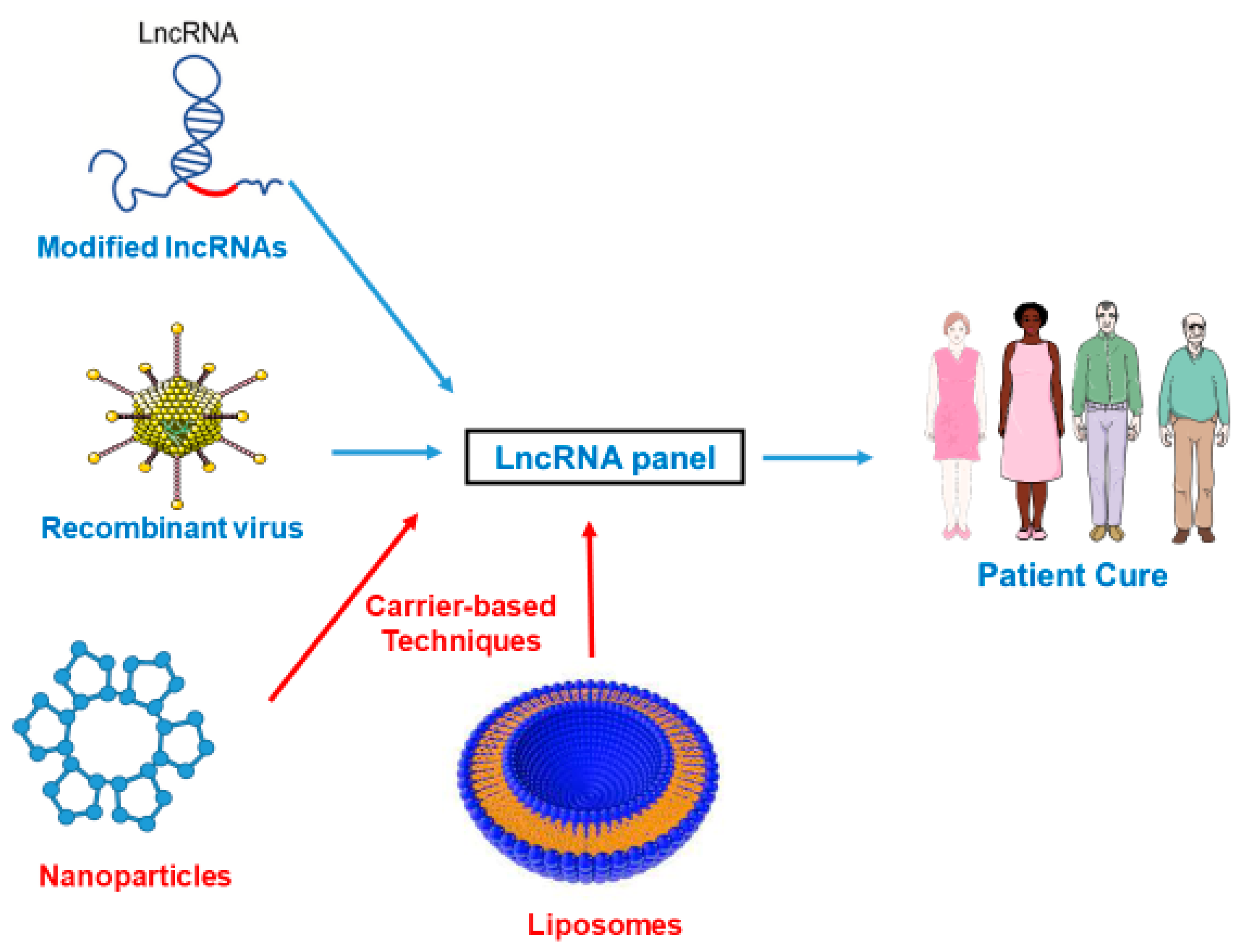{kind=link}
| AKI Model | LncRNA | Targeted miRNA | miRNA Beneficial Effects | References |
|---|---|---|---|---|
| Sepsis | lncRNA 6406 | miR-687 | Reduces inflammation, oxidative stress and cell death. | [13] |
| TCONS_00016233 | miR-22-3p | Inhibits apoptosis and TLR4-mediated inflammatory response. | [15] | |
| MALAT1 | miR-146a | Reduces cell death and NF-κB mediated inflammation. | [16,17,18] | |
| NEAT1 | miR-204 | Decreases oxidative stress, apoptosis and NF-κB associated inflammation. | [19] | |
| miR-22-3p | Reduces oxidative stress, cell death, autophagy and inflammation | [20] | ||
| miR-125a-5p | Decreases inflammation by promoting macrophage M2 polarization. | [22] | ||
| miR-27a-3p | Alleviates oxidative stress, apoptosis and reduces kidney injury. | [23] | ||
| TUG1 | miR-223 | Reduces oxidative stress and cell death by activating Nrf2 pathway. | [24,25,26] | |
| HOXA-AS2 | miR-106b-5p | Inhibits NF-κB/Wnt/β-catenin pathways and reduces renal damage. | [27] | |
| CASC2 | miR-155 | Suppresses NF- κB mediated inflammatory cytokines expression. | [28] | |
| TapSAKI | miR-22 | Inhibits cell apoptosis and inflammation. | [29,30] | |
| HOTAIR | miR-22 | Decreases cell death and improves renal function. | [31] | |
| miR-34a | Improves renal function, reduces cell death and inflammation. | [32] | ||
| MEG3 | miR-21 | Reduces cell apoptosis. | [33] | |
| DANCR | miR-214 | Suppresses cell death and inflammatory cytokines production. | [34] | |
| CRNDE | miR-181a-5p | Induces cell proliferation and decreases cell death and inflammation. | [35] | |
| miR-146a | Reduces cell death and inflammatory TLR4/NF-κB signaling pathway. | [37] | ||
| SIKIAT1 | miR-96 | Decreases cell apoptosis. | [38] | |
| MIAT | miR-29a | Reduces cell death by inhibiting caspase 8. | [39] | |
| LINC00261 | miR-654-5p | Suppresses cell apoptosis and inflammation via. NF-κB inhibition. | [41] | |
| DLX6-AS1 | miR-223-3p | Decreases NLRP3 expression and pyroptosis. | [42] | |
| SNHG5 | miR-374a-3p | Reduces NF-κB/TLR4 mediated inflammation and cell death. | [43] | |
| SNHG14 | miR-93 | Ameliorates oxidative stress, cell death and inflammation by activating IRAK4/NF-κB signaling. | [44] | |
| I/R | XIST | miR-124-3p | Improves renal function and decreases inflammation. | [49] |
| miR142-5p | Reduces renal injury and inflammation. | [50] | ||
| H19 | miR-30a-5p | Decreases cell death and inflammation. | [51] | |
| MALAT1 | miR-146a | Improves renal injury, inflammation, and fibrosis. | [52,53] | |
| NEAT1 | miR27a-3p | Ameliorates hypoxia-mediated cell death. | [54] | |
| GAS5 | miR-21 | Reduces renal injury and cell death. | [60] | |
| LINC00520 | miR-27b-3p | Improves renal injury and cell death. | [56] | |
| SNHG14 | miR-124-3p | Decreases inflammation and oxidative stress. | [57] | |
| MEG3 | miR-129-5p | Reduces tubuloepithelial cell death. | [59] | |
| Cisplatin | XLOC-032768 | Decreases apoptosis and inflammation. | [61] | |
| LRNA9884 | Reduces tubular cell death and inflammation. | [62] | ||
| UUO | TCONS_00088786 | Reduces TGF-β/Smad mediated fibrosis. | [63] | |
| TCONS_01496394 | [63] | |||
| Gm10251 | Decreases fibrosis in tubular cells. | [64] | ||
| Fam120aos | [64] | |||
| Gm16076 | [64] | |||
| lncRNA 74.1 | Reduces fibrosis by inhibiting Nrf2-Keap1 pathway. | [65,66] | ||
| MIAT | Targets α-SMAD and reduces fibrosis. | [67,68] | ||
| H19 | [69] | |||
| HOTAIR | miR-124 | Reduces fibrosis by inhibiting Notch1. | [70,71] |
2.1.3. Nephrotoxic Agents
2.1.4. Obstructive AKI
2.2. Chronic Kidney Disease
2.3. Diabetic Nephropathy
2.4. Glomerular Diseases
2.4.1. Lupus Nephritis
2.4.2. Focal Segmental Glomerulosclerosis
2.4.3. IgA Nephropathy
2.4.4. Membranous Nephropathy
2.5. Renal Cancers
3. Perspectives in Therapy
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cech, T.R. The RNA worlds in context. Cold Spring Harb. Perspect. Biol. 2012, 4, a006742. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Froberg, J.E.; Lee, J.T. Long noncoding RNAs: Fresh perspectives into the RNA world. Trends Biochem. Sci. 2014, 39, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Crick, F.H. On protein synthesis. Symp. Soc. Exp. Biol. 1958, 12, 138–163. [Google Scholar]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in control of gene expression: An overview of nuclear functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef]
- Metzinger-Le Meuth, V.; Fourdinier, O.; Charnaux, N.; Massy, Z.A.; Metzinger, L. The expanding roles of microRNAs in kidney pathophysiology. Nephrol. Dial. Transpl. 2019, 34, 7–15. [Google Scholar] [CrossRef]
- Mattick, J.S. The state of long non-coding RNA biology. Noncoding RNA 2018, 4, 17. [Google Scholar] [CrossRef]
- Bhat, S.A.; Ahmad, S.M.; Mumtaz, P.T.; Malik, A.A.; Dar, M.A.; Urwat, U.; Shah, R.A.; Ganai, N.A. Long non-coding RNAs: Mechanism of action and functional utility. Noncoding RNA Res. 2016, 1, 43–50. [Google Scholar] [CrossRef]
- Guttman, M.; Donaghey, J.; Carey, B.W.; Garber, M.; Grenier, J.K.; Munson, G.; Young, G.; Lucas, A.B.; Ach, R.; Bruhn, L.; et al. lincRNAs act in the circuitry controlling pluripotency and differentiation. Nature 2011, 477, 295–300. [Google Scholar] [CrossRef]
- Song, Z.; Lin, L.; Li, Z.; Huang, C. The nuclear functions of long noncoding RNAs come into focus. Noncoding RNA Res. 2021, 6, 70–79. [Google Scholar] [CrossRef]
- Geisler, S.; Coller, J. RNA in unexpected places: Long non-coding RNA functions in diverse cellular contexts. Nat. Rev. Mol. Cell Biol. 2013, 14, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Poston, J.T.; Koyner, J.L. Sepsis associated acute kidney injury. BMJ 2019, 364, k4891. [Google Scholar] [CrossRef]
- Chun-Mei, H.; Qin-Min, G.; Shu-Ming, P.; Xiang-Yang, Z. Expression profiling and ontology analysis of circulating long non-coding RNAs in septic acute kidney injury patients. Clin. Chem. Lab. Med. 2016, 54, e395–e399. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhu, N.; Zhang, B.; Xu, S.B. Long noncoding RNA TCONS_00016406 attenuates Lipopolysaccharide-induced acute kidney injury by regulating the miR-687/PTEN pathway. Front. Physiol. 2020, 11, 622. [Google Scholar] [CrossRef]
- Zhang, P.; Yi, L.; Qu, S.; Dai, J.; Li, X.; Liu, B.; Li, H.; Ai, K.; Zheng, P.; Qiu, S.; et al. The biomarker TCONS_00016233 drives septic AKI by targeting the miR-22-3p/AIFM1 signaling axis. Mol. Ther. Nucleic Acids 2020, 19, 1027–1042. [Google Scholar] [CrossRef]
- Xu, L.; Hu, G.; Xing, P.; Zhou, M.; Wang, D. Paclitaxel alleviates the sepsis-induced acute kidney injury via lnc-MALAT1/miR-370-3p/HMGB1 axis. Life Sci. 2020, 262, 118505. [Google Scholar] [CrossRef]
- Ding, Y.; Guo, F.; Zhu, T.; Li, J.; Gu, D.; Jiang, W.; Lu, Y.; Zhou, D. Mechanism of long non-coding RNA MALAT1 in lipopolysaccharide-induced acute kidney injury is mediated by the miR-146a/NF-κB signaling pathway. Int. J. Mol. Med. 2018, 41, 446–454. [Google Scholar] [CrossRef]
- Zhu, S.; Lu, Y. Dexmedetomidine suppressed the biological behavior of HK-2 cells treated with LPS by down-regulating ALKBH5. Inflammation 2020, 43, 2256–2263. [Google Scholar] [CrossRef]
- Chen, Y.; Qiu, J.; Chen, B.; Lin, Y.; Chen, Y.; Xie, G.; Qiu, J.; Tong, H.; Jiang, D. Long non-coding RNA NEAT1 plays an important role in sepsis-induced acute kidney injury by targeting miR-204 and modulating the NF-κB pathway. Int. Immunopharmacol. 2018, 59, 252–260. [Google Scholar] [CrossRef]
- Feng, Y.; Liu, J.; Wu, R.; Yang, P.; Ye, Z.; Song, F. NEAT1 aggravates sepsis-induced acute kidney injury by sponging miR-22-3p. Open Med. 2020, 15, 333–342. [Google Scholar] [CrossRef]
- Gao, C.; Zou, X.; Chen, H.; Shang, R.; Wang, B. Long non-coding RNA nuclear paraspeckle assembly transcript 1 (NEAT1)Relieves sepsis-induced kidney injury and Lipopolysaccharide (LPS)-induced inflammation in HK-2 cells. Med. Sci. Monit. 2020, 26, e921906. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Guo, Z.-H. Downregulation of lncRNA NEAT1 ameliorates LPS-induced inflammatory responses by promoting macrophage M2 polarization via miR-125a-5p/TRAF6/TAK1 axis. Inflammation 2020, 43, 1548–1560. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Y.; Tang, Z.; Hu, D.; Yao, C.; Yang, L. LncRNA NEAT1 regulated inflammation and apoptosis in a rat model of sepsis-induced acute kidney injury via MiR-27a-3p/TAB3 axis. Biosci. Biotechnol. Biochem. 2020, 84, 2215–2227. [Google Scholar] [CrossRef]
- M’baya-Moutoula, E.; Louvet, L.; Molinié, R.; Guerrera, I.C.; Cerutti, C.; Fourdinier, O.; Nourry, V.; Gutierrez, L.; Morlière, P.; Mesnard, F.; et al. A multi-omics analysis of the regulatory changes induced by miR-223 in a monocyte/macrophage cell line. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 2664–2678. [Google Scholar] [CrossRef] [PubMed]
- Li, S.Y.; Susztak, K. The long noncoding RNA Tug1 connects metabolic changes with kidney disease in podocytes. J. Clin. Investig. 2016, 126, 4072–4075. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, S.; Yang, F.; Xie, J.; Chen, J.; Li, Z. Diosmetin alleviates acute kidney injury by promoting the TUG1/Nrf2/HO-1 pathway in sepsis rats. Int. Immunopharmacol. 2020, 88, 106965. [Google Scholar] [CrossRef]
- Wu, H.; Wang, J.; Ma, Z. Long noncoding RNA HOXA-AS2 mediates microRNA-106b-5p to repress sepsis-engendered acute kidney injury. J. Biochem. Mol. Toxicol. 2020, 34, e22453. [Google Scholar] [CrossRef]
- Wang, M.; Wei, J.; Shang, F.; Zang, K.; Ji, T. Long non-coding RNA CASC2 ameliorates sepsis-induced acute kidney injury by regulating the miR-155 and NF-κB pathway. Int. J. Mol. Med. 2020, 45, 1554–1562. [Google Scholar] [CrossRef]
- Shen, J.; Liu, L.; Zhang, F.; Gu, J.; Pan, G. LncRNA TapSAKI promotes inflammation injury in HK-2 cells and urine derived sepsis-induced kidney injury. J. Pharm. Pharmacol. 2019, 71, 839–848. [Google Scholar] [CrossRef]
- Huang, W.; Lan, X.; Li, X.; Wang, D.; Sun, Y.; Wang, Q.; Gao, H.; Yu, K. Long non-coding RNA PVT1 promote LPS-induced septic acute kidney injury by regulating TNFα and JNK/NF-κB pathways in HK-2 cells. Int. Immunopharmacol. 2017, 47, 134–140. [Google Scholar] [CrossRef]
- Shen, J.; Zhang, J.; Jiang, X.; Wang, H.; Pan, G. LncRNA HOX transcript antisense RNA accelerated kidney injury induced by urine-derived sepsis through the miR-22/high mobility group box 1 pathway. Life Sci. 2018, 210, 185–191. [Google Scholar] [CrossRef]
- Jiang, Z.-J.; Zhang, M.-Y.; Fan, Z.-W.; Sun, W.-L.; Tang, Y. Influence of lncRNA HOTAIR on acute kidney injury in sepsis rats through regulating miR-34a/Bcl-2 pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 3512–3519. [Google Scholar]
- Yang, R.; Liu, S.; Wen, J.; Xue, L.; Zhang, Y.; Yan, D.; Wang, G.; Liu, Z. Inhibition of maternally expressed gene 3 attenuated lipopolysaccharide-induced apoptosis through sponging miR-21 in renal tubular epithelial cells. J. Cell. Biochem. 2018, 119, 7800–7806. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, B.; Li, Z.; Wang, B.; Li, L. Long noncoding RNA DANCR suppressed Lipopolysaccharide-induced septic acute kidney injury by regulating miR-214 in HK-2 cells. Med. Sci. Monit. 2020, 26, e921822. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Song, J.; Li, Y.; Shao, J.; Xie, Z.; Sun, K. Down-regulation of LncRNA CRNDE aggravates kidney injury via increasing MiR-181a-5p in sepsis. Int. Immunopharmacol. 2020, 79, 105933. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.-Q.; Sui, Y.-D.; Huang, H.; Zou, X.-B.; Chen, S.-C.; Yu, Z.-K. Effect of lncRNA CRNDE on sepsis-related kidney injury through the TLR3/NF-κB pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 10489–10497. [Google Scholar] [PubMed]
- Wu, S.; Qiu, H.; Wang, Q.; Cao, Z.; Wang, J. Effects and mechanism of lncRNA CRNDE on sepsis-induced acute kidney injury. anal cell pathol. Anal. Cell Pathol. 2020, 2020, 8576234. [Google Scholar] [CrossRef]
- Lu, S.; Wu, H.; Xu, J.; He, Z.; Li, H.; Ning, C. SIKIAT1/miR-96/FOXA1 axis regulates sepsis-induced kidney injury through induction of apoptosis. Inflamm. Res. 2020, 69, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Y.-Y.; Xia, F.; Yang, A.-X.; Qian, J.-X.; Zhao, H.; Tao, W.-Y. Effect of lncRNA-MIAT on kidney injury in sepsis rats via regulating miR-29a expression. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 10942–10949. [Google Scholar] [PubMed]
- Xie, Z.; Guo, Z.; Liu, J. Whey acidic protein/four-disulfide core domain 21 regulate sepsis pathogenesis in a mouse model and a macrophage cell line via the Stat3/Toll-like receptor 4 (TLR4) signaling pathway. Med. Sci. Monit. 2018, 24, 4054–4063. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, J.; Lu, P.; Li, M. LINC00261 relieves the progression of sepsis-induced acute kidney injury by inhibiting NF-κB activation through targeting the miR-654-5p/SOCS3 axis. J. Bioenerg. Biomembr. 2021, 53, 129–137. [Google Scholar] [CrossRef]
- Tan, J.; Fan, J.; He, J.; Zhao, L.; Tang, H. Knockdown of LncRNA DLX6-AS1 inhibits HK-2 cell pyroptosis via regulating miR-223-3p/NLRP3 pathway in lipopolysaccharide-induced acute kidney injury. J. Bioenerg. Biomembr. 2020, 52, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wei, J.; Shang, F.; Zang, K.; Zhang, P. Down-regulation of lncRNA SNHG5 relieves sepsis-induced acute kidney injury by regulating the miR-374a-3p/TLR4/NF-κB pathway. J. Biochem. 2021. [Google Scholar] [CrossRef]
- Shi, C.; Zhao, Y.; Li, Q.; Li, J. lncRNA SNHG14 Plays a Role in Sepsis-Induced Acute Kidney Injury by Regulating miR-93. Mediat. Inflamm. 2021, 2021, 5318369. [Google Scholar] [CrossRef]
- Fang, Y.; Hu, J.-F.; Wang, Z.-H.; Zhang, S.-G.; Zhang, R.-F.; Sun, L.-M.; Cui, H.-W.; Yang, F. GAS5 promotes podocyte injury in sepsis by inhibiting PTEN expression. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8423–8430. [Google Scholar]
- Arslan, F.; Keogh, B.; McGuirk, P.; Parker, A.E. TLR2 and TLR4 in ischemia reperfusion injury. Mediat. Inflamm. 2010, 2010, 704202. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lv, X.; Fan, Q.; Wang, X.; Xu, L.; Lu, X.; Chen, T. Analysis of circulating lncRNA expression profiles in patients with diabetes mellitus and diabetic nephropathy: Differential expression profile of circulating lncRNA. Clin. Nephrol. 2019, 92, 25–35. [Google Scholar] [CrossRef]
- Tao, Q.; Tianyu, W.; Jiangqiao, Z.; Zhongbao, C.; Xiaoxiong, M.; Long, Z.; Jilin, Z. Expression analysis of long non-coding RNAs in a renal ischemia-reperfusion injury model. Acta Cir. Bras. 2019, 34, e201900403. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Hu, Y.; Xie, Y.; Zhao, Z.; Ma, L.; Li, Z.; Tan, W. Total glucosides of paeony alleviate cell apoptosis and inflammation by targeting the long noncoding RNA XIST/MicroRNA-124-3p/ITGB1 axis in renal ischemia/reperfusion injury. Mediat. Inflamm. 2020, 2020, 8869511. [Google Scholar] [CrossRef]
- Tang, B.; Li, W.; Ji, T.; Li, X.; Qu, X.; Feng, L.; Zhu, Y.; Qi, Y.; Zhu, C.; Bai, S. Downregulation of XIST ameliorates acute kidney injury by sponging miR-142-5p and targeting PDCD4. J. Cell Physiol. 2020, 235, 8852–8863. [Google Scholar] [CrossRef]
- Haddad, G.; Kölling, M.; Wegmann, U.A.; Dettling, A.; Seeger, H.; Schmitt, R.; Soerensen-Zender, I.; Haller, H.; Kistler, A.D.; Dueck, A.; et al. Renal AAV2-mediated overexpression of long non-coding RNA H19 attenuates ischemic acute kidney injury through sponging of microRNA-30a-5p. J. Am. Soc. Nephrol. 2021, 32, 323–341. [Google Scholar] [CrossRef] [PubMed]
- Kölling, M.; Genschel, C.; Kaucsar, T.; Hübner, A.; Rong, S.; Schmitt, R.; Soerensen-Zender, I.; Haddad, G.; Kistler, A.D.; Seeger, H.; et al. Hypoxia-induced long non-coding RNA Malat1 is dispensable for renal ischemia/reperfusion-injury. Sci. Rep. 2018, 8, 3438. [Google Scholar] [CrossRef]
- Tian, H.; Wu, M.; Zhou, P.; Huang, C.; Ye, C.; Wang, L. The long non-coding RNA MALAT1 is increased in renal ischemia-reperfusion injury and inhibits hypoxia-induced inflammation. Ren. Fail. 2018, 40, 527–533. [Google Scholar] [CrossRef]
- Jiang, X.; Li, D.; Shen, W.; Shen, X.; Liu, Y. LncRNA NEAT1 promotes hypoxia-induced renal tubular epithelial apoptosis through downregulating miR-27a-3p. J. Cell Biochem. 2019, 120, 16273–16282. [Google Scholar] [CrossRef]
- Wang, I.-K.; Palanisamy, K.; Sun, K.-T.; Yu, S.-H.; Yu, T.-M.; Li, C.-H.; Li, F.-Y.; Chou, A.-K.; Wang, G.-J.; Chen, K.-B.; et al. The functional interplay of lncRNA EGOT and HuR regulates hypoxia-induced autophagy in renal tubular cells. J. Cell Biochem. 2020, 121, 4522–4534. [Google Scholar] [CrossRef]
- Tian, X.; Ji, Y.; Liang, Y.; Zhang, J.; Guan, L.; Wang, C. LINC00520 targeting miR-27b-3p regulates OSMR expression level to promote acute kidney injury development through the PI3K/AKT signaling pathway. J. Cell Physiol. 2019, 234, 14221–14233. [Google Scholar] [CrossRef] [PubMed]
- Xue, Q.; Yang, L.; Wang, H.; Han, S. Silence of long noncoding RNA SNHG14 alleviates ischemia/reperfusion-induced acute kidney injury by regulating miR-124-3p/MMP2 axis. Biomed Res. Int. 2021, 2021, 8884438. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.-M.; Palanisamy, K.; Sun, K.-T.; Day, Y.-J.; Shu, K.-H.; Wang, I.-K.; Shyu, W.-C.; Chen, P.; Chen, Y.-L.; Li, C.-Y. RANTES mediates kidney ischemia reperfusion injury through a possible role of HIF-1α and LncRNA PRINS. Sci. Rep. 2016, 6, 18424. [Google Scholar] [CrossRef]
- Mao, H.; Huang, Q.; Liu, Y. MEG3 aggravates hypoxia/reoxygenation induced apoptosis of renal tubular epithelial cells via the miR-129-5p/HMGB1 axis. J. Biochem. Mol. Toxicol. 2020, 35, e22649. [Google Scholar] [CrossRef]
- Geng, X.; Song, N.; Zhao, S.; Xu, J.; Liu, Y.; Fang, Y.; Liang, M.; Xu, X.; Ding, X. LncRNA GAS5 promotes apoptosis as a competing endogenous RNA for miR-21 via thrombospondin 1 in ischemic AKI. Cell Death Discov. 2020, 6, 19. [Google Scholar] [CrossRef]
- Zhou, X.; Jiang, K.; Luo, H.; Wu, C.; Yu, W.; Cheng, F. Novel lncRNA XLOC_032768 alleviates cisplatin-induced apoptosis and inflammatory response of renal tubular epithelial cells through TNF-α. Int. Immunopharmacol. 2020, 83, 106472. [Google Scholar] [CrossRef]
- Zhang, Y.; Tang, P.M.-K.; Niu, Y.; García Córdoba, C.A.; Huang, X.-R.; Yu, C.; Lan, H.-Y. Long non-coding RNA LRNA9884 promotes acute kidney injury via regulating NF-kB-mediated transcriptional activation of MIF. Front. Physiol. 2020, 11, 590027. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, S.; Shi, B.; Zheng, D.; Shi, J. Transcriptome identified lncRNAs associated with renal fibrosis in UUO rat model. Front. Physiol. 2017, 8, 658. [Google Scholar] [CrossRef]
- Xiao, H.; Liao, Y.; Tang, C.; Xiao, Z.; Luo, H.; Li, J.; Liu, H.; Sun, L.; Zeng, D.; Li, Y. RNA-Seq analysis of potential lncRNAs and genes for the anti-renal fibrotic effect of norcantharidin. J. Cell Biochem. 2019, 120, 17354–17367. [Google Scholar] [CrossRef]
- Lu, M.; Wang, P.; Qiao, Y.; Jiang, C.; Ge, Y.; Flickinger, B.; Malhotra, D.K.; Dworkin, L.D.; Liu, Z.; Gong, R. GSK3β-mediated Keap1-independent regulation of Nrf2 antioxidant response: A molecular rheostat of acute kidney injury to chronic kidney disease transition. Redox Biol. 2019, 26, 101275. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Yuan, Q.; Chen, Y.; Huang, Z.; Fang, X.; Zhang, H.; Peng, L.; Xiao, P. LncRNA ENST00000453774.1 contributes to oxidative stress defense dependent on autophagy mediation to reduce extracellular matrix and alleviate renal fibrosis. J. Cell Physiol. 2019, 234, 9130–9143. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, B.; Chen, Z.; He, Y.; Ru, F.; Liu, P.; Chen, X. The long noncoding RNA myocardial infarction-associated transcript modulates the epithelial-mesenchymal transition in renal interstitial fibrosis. Life Sci. 2020, 241, 117187. [Google Scholar] [CrossRef] [PubMed]
- Bijkerk, R.; Au, Y.W.; Stam, W.; Duijs, J.M.G.J.; Koudijs, A.; Lievers, E.; Rabelink, T.J.; van Zonneveld, A.J. Long non-coding RNAs rian and miat mediate myofibroblast formation in kidney fibrosis. Front. Pharmacol. 2019, 10, 215. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Xue, J.-D.; Chao, F.; Jin, Y.-F.; Fu, Q. Long non-coding RNA-H19 antagonism protects against renal fibrosis. Oncotarget 2016, 7, 51473–51481. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Qiu, Z.-Z.; Yu, Z.-H.; Gao, L.; He, J.-M.; Zhang, Z.-W.; Zheng, J. Paeonol reverses promoting effect of the HOTAIR/miR-124/Notch1 axis on renal interstitial fibrosis in a rat model. J. Cell Physiol. 2019, 234, 14351–14363. [Google Scholar] [CrossRef]
- Matwijiw, I.; Faiman, C. Control of gonadotropin secretion in the ovine fetus. II. A sex difference in pulsatile luteinizing hormone secretion after castration. Endocrinology 1989, 124, 1352–1358. [Google Scholar] [CrossRef]
- Arany, I.; Safirstein, R.L. Cisplatin nephrotoxicity. Semin. Nephrol. 2003, 23, 460–464. [Google Scholar] [CrossRef]
- Yousef, M.I.; Hussien, H.M. Cisplatin-induced renal toxicity via tumor necrosis factor-α, interleukin 6, tumor suppressor P53, DNA damage, xanthine oxidase, histological changes, oxidative stress and nitric oxide in rats: Protective effect of ginseng. Food Chem. Toxicol. 2015, 78, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Iñiguez, J.S.; Navarro-Gallardo, G.J.; Medina-González, R.; Alcantar-Vallin, L.; García-García, G. Acute kidney injury caused by obstructive nephropathy. Int. J. Nephrol. 2020, 2020, 8846622. [Google Scholar] [CrossRef]
- Ucero, A.C.; Benito-Martin, A.; Izquierdo, M.C.; Sanchez-Niño, M.D.; Sanz, A.B.; Ramos, A.M.; Berzal, S.; Ruiz-Ortega, M.; Egido, J.; Ortiz, A. Unilateral ureteral obstruction: Beyond obstruction. Int. Urol. Nephrol. 2014, 46, 765–776. [Google Scholar] [CrossRef] [PubMed]
- Docherty, N.G.; O’Sullivan, O.E.; Healy, D.A.; Fitzpatrick, J.M.; Watson, R.W.G. Evidence that inhibition of tubular cell apoptosis protects against renal damage and development of fibrosis following ureteric obstruction. Am. J. Physiol. Ren. Physiol. 2006, 290, F4–F13. [Google Scholar] [CrossRef]
- Chevalier, R.L.; Forbes, M.S.; Thornhill, B.A. Ureteral obstruction as a model of renal interstitial fibrosis and obstructive nephropathy. Kidney Int. 2009, 75, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Li, G.; Gao, L.; Cheng, X.; Wang, L.; Qin, Y.; Zhang, D. Exaggerated renal fibrosis in lncRNA Gas5-deficient mice after unilateral ureteric obstruction. Life Sci. 2021, 264, 118656. [Google Scholar] [CrossRef]
- Parikh, N.I.; Hwang, S.-J.; Larson, M.G.; Meigs, J.B.; Levy, D.; Fox, C.S. Cardiovascular disease risk factors in chronic kidney disease: Overall burden and rates of treatment and control. Arch. Intern. Med. 2006, 166, 1884. [Google Scholar] [CrossRef]
- Hamza, E.; Metzinger, L.; Meuth, V.M.-L. Uremic toxins affect erythropoiesis during the course of chronic kidney disease: A review. Cells 2020, 9, 2039. [Google Scholar] [CrossRef]
- Fourdinier, O.; Schepers, E.; Meuth, V.M.-L.; Glorieux, G.; Liabeuf, S.; Verbeke, F.; Vanholder, R.; Brigant, B.; Pletinck, A.; Diouf, M.; et al. Serum levels of miR-126 and miR-223 and outcomes in chronic kidney disease patients. Sci. Rep. 2019, 9, 4477. [Google Scholar] [CrossRef]
- Ishida, M.; Selaru, F.M. miRNA-based therapeutic strategies. Curr. Pathobiol. Rep. 2013, 1, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Khurana, R.; Ranches, G.; Schafferer, S.; Lukasser, M.; Rudnicki, M.; Mayer, G.; Hüttenhofer, A. Identification of urinary exosomal noncoding RNAs as novel biomarkers in chronic kidney disease. RNA 2017, 23, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Cui, Y.; Yin, M.; Liu, F. Screening potential prognostic biomarkers of long non-coding RNAs for predicting the risk of chronic kidney disease. Braz. J. Med. Biol. Res. 2019, 52, e8333. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.-F.; Chen, Y.-T.; Gu, J.; Nerbonne, J.M.; Lin, C.-H.; Yang, K.-C. Circulating long noncoding RNA DKFZP434I0714 predicts adverse cardiovascular outcomes in patients with end-stage renal disease. Int. J. Cardiol. 2019, 277, 212–219. [Google Scholar] [CrossRef]
- Santer, L.; López, B.; Ravassa, S.; Baer, C.; Riedel, I.; Chatterjee, S.; Moreno, M.U.; González, A.; Querejeta, R.; Pinet, F.; et al. Circulating long noncoding RNA LIPCAR predicts heart failure outcomes in patients without chronic kidney disease. Hypertension 2019, 73, 820–828. [Google Scholar] [CrossRef] [PubMed]
- Arbiol-Roca, A.; Padró-Miquel, A.; Hueso, M.; Navarro, E.; Alía-Ramos, P.; González-Álvarez, M.; Rama, I.; Torras, J.; Grinyó, J.; Cruzado, J.; et al. Association of ANRIL gene polymorphisms with major adverse cardiovascular events in hemodialysis patients. Clin. Chim. Acta 2017, 466, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.; Guo, Y.; Diao, Z.; Guo, W.; Liu, W. Genome-wide identification of lncRNAs and mRNAs differentially expressed in human vascular smooth muscle cells stimulated by high phosphorus. Ren. Fail. 2020, 42, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Lai, C.-F.; Chang-Panesso, M.; Humphreys, B.D. Proximal tubule translational profiling during kidney fibrosis reveals proinflammatory and long noncoding RNA expression patterns with sexual dimorphism. JASN 2020, 31, 23–38. [Google Scholar] [CrossRef]
- Meng, X.; Nikolic-Paterson, D.J.; Lan, H.Y. TGF-β: The master regulator of fibrosis. Nat. Rev. Nephrol. 2016, 12, 325–338. [Google Scholar] [CrossRef]
- Meng, X.M.; Huang, X.R.; Chung, A.C.K.; Qin, W.; Shao, X.; Igarashi, P.; Ju, W.; Bottinger, E.P.; Lan, H.Y. Smad2 protects against TGF-β/Smad3-mediated renal fibrosis. J. Am. Soc. Nephrol. 2010, 21, 1477–1487. [Google Scholar] [CrossRef]
- Zhou, Q.; Chung, A.C.K.; Huang, X.R.; Dong, Y.; Yu, X.; Lan, H.Y. Identification of novel long noncoding RNAs associated with TGF-β/Smad3-mediated renal inflammation and fibrosis by RNA sequencing. Am. J. Pathol. 2014, 184, 409–417. [Google Scholar] [CrossRef]
- Pu, Y.; Zhao, H.; Wu, X.; Mei, M.; Shen, B. The long noncoding RNA Ptprd-IR is a novel molecular target for TGF-β1-mediated nephritis. Int. J. Biochem. Cell Biol. 2020, 122, 105742. [Google Scholar] [CrossRef]
- Long, J.; Badal, S.S.; Ye, Z.; Wang, Y.; Ayanga, B.A.; Galvan, D.L.; Green, N.H.; Chang, B.H.; Overbeek, P.A.; Danesh, F.R. Long noncoding RNA Tug1 regulates mitochondrial bioenergetics in diabetic nephropathy. J. Clin. Investig. 2016, 126, 4205–4218. [Google Scholar] [CrossRef]
- Allison, S.J. A lncRNA and miRNA megacluster in diabetic nephropathy. Nat. Rev. Nephrol. 2016, 12, 713. [Google Scholar] [CrossRef]
- Yang, J.; Shen, Y.; Yang, X.; Long, Y.; Chen, S.; Lin, X.; Dong, R.; Yuan, J. Silencing of long noncoding RNA XIST protects against renal interstitial fibrosis in diabetic nephropathy via microRNA-93-5p-mediated inhibition of CDKN1A. Am. J. Physiol. Ren. Physiol. 2019, 317, F1350–F1358. [Google Scholar] [CrossRef]
- Jie, R.; Zhu, P.; Zhong, J.; Zhang, Y.; Wu, H. LncRNA KCNQ1OT1 affects cell proliferation, apoptosis and fibrosis through regulating miR-18b-5p/SORBS2 axis and NF-ĸB pathway in diabetic nephropathy. Diabetol Metab. Syndr. 2020, 12, 77. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.-F.; Chenier, I.; Lavoie, J.L.; Chan, J.S.; Hamet, P.; Tremblay, J.; Chen, X.M.; Wang, N.H.; Inagami, T. Development of hypertension and kidney hypertrophy in transgenic mice overexpressing ARAP1 gene in the kidney. Hypertension 2006, 48, 453–459. [Google Scholar] [CrossRef][Green Version]
- Ma, T.-K.; Xu, L.; Lu, L.-X.; Cao, X.; Li, X.; Li, L.-L.; Wang, X.; Fan, Q.-L. Ursolic acid treatment alleviates diabetic kidney injury by regulating the ARAP1/AT1R signaling pathway. Diabetes Metab. Syndr. Obes. 2019, 12, 2597–2608. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xu, L.; Wen, S.; Yang, Y.; Li, X.; Fan, Q. The effect of lncRNA-ARAP1-AS2/ARAP1 on high glucose-induced cytoskeleton rearrangement and epithelial–mesenchymal transition in human renal tubular epithelial cells. J. Cell Physiol. 2020, 235, 5787–5795. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Ye, Z.; Xue, Z.; Wu, L.; Ouyang, Y.; Yao, C.; Cui, C.; Xu, N.; Ma, J.; Hou, G.; et al. Identification of renal long non-coding RNA RP11-2B6.2 as a positive regulator of type i interferon signaling pathway in lupus nephritis. Front. Immunol. 2019, 10, 975. [Google Scholar] [CrossRef] [PubMed]
- Luan, J.; Jiao, C.; Kong, W.; Fu, J.; Qu, W.; Chen, Y.; Zu, X.; Zeng, Y.; Guo, G.; Qi, H.; et al. circHLA-C Plays an important role in lupus nephritis by sponging miR-150. Mol. Ther. Nucleic Acids 2018, 10, 245–253. [Google Scholar] [CrossRef]
- Wu, G.-C.; Li, J.; Leng, R.-X.; Li, X.-P.; Li, X.-M.; Wang, D.-G.; Pan, H.-F.; Ye, D.-Q. Identification of long non-coding RNAs GAS5, linc0597 and lnc-DC in plasma as novel biomarkers for systemic lupus erythematosus. Oncotarget 2017, 8, 23650–23663. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Q.; Huang, Q.; Jiang, Z.; Zhao, J.; Shi, G.-P.; Yang, M. Using plasma circRNA_002453 as a novel biomarker in the diagnosis of lupus nephritis. Mol. Immunol. 2018, 101, 531–538. [Google Scholar] [CrossRef]
- Han, R.; Hu, S.; Qin, W.; Shi, J.; Zeng, C.; Bao, H.; Liu, Z. Upregulated long noncoding RNA LOC105375913 induces tubulointerstitial fibrosis in focal segmental glomerulosclerosis. Sci. Rep. 2019, 9, 716. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Han, R.; Shi, J.; Zhu, X.; Qin, W.; Zeng, C.; Bao, H.; Liu, Z. The long noncoding RNA LOC105374325 causes podocyte injury in individuals with focal segmental glomerulosclerosis. J. Biol. Chem. 2018, 293, 20227–20239. [Google Scholar] [CrossRef] [PubMed]
- Sui, W.; Li, H.; Ou, M.; Tang, D.; Dai, Y. Altered long non-coding RNA expression profile in patients with IgA-negative mesangial proliferative glomerulonephritis. Int. J. Mol. Med. 2012, 30, 173–178. [Google Scholar]
- Zuo, N.; Li, Y.; Liu, N.; Wang, L. Differentially expressed long non-coding RNAs and mRNAs in patients with IgA nephropathy. Mol. Med. Rep. 2017, 16, 7724–7730. [Google Scholar] [CrossRef] [PubMed]
- Gholaminejad, A.; Gheisari, Y.; Jalali, S.; Roointan, A. Comprehensive analysis of IgA nephropathy expression profiles: Identification of potential biomarkers and therapeutic agents. BMC Nephrol. 2021, 22, 137. [Google Scholar] [CrossRef]
- Guo, N.; Zhou, Q.; Huang, X.; Yu, J.; Han, Q.; Nong, B.; Xiong, Y.; Liang, P.; Li, J.; Feng, M.; et al. Identification of differentially expressed circulating exosomal lncRNAs in IgA nephropathy patients. BMC Immunol. 2020, 21, 16. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-S.; Hsieh, H.-Y.; Shih, H.-M.; Sytwu, H.-K.; Wu, C.-C. Urinary Xist is a potential biomarker for membranous nephropathy. Biochem. Biophys. Res. Commun. 2014, 452, 415–421. [Google Scholar] [CrossRef]
- Jin, L.-W.; Pan, M.; Ye, H.-Y.; Zheng, Y.; Chen, Y.; Huang, W.-W.; Xu, X.-Y.; Zheng, S.-B. Down-regulation of the long non-coding RNA XIST ameliorates podocyte apoptosis in membranous nephropathy via the miR-217-TLR4 pathway. Exp. Physiol. 2019, 104, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Jia, Z.; Duan, R.; Yan, Z.; Jin, Z.; Yan, L.; Li, Q.; Yang, J. Long non-coding RNA XIST regulates miR-106b-5p/P21 axis to suppress tumor progression in renal cell carcinoma. Biochem. Biophys. Res. Commun. 2019, 510, 416–420. [Google Scholar] [CrossRef]
- Kong, Y.; Hsieh, C.-H.; Alonso, L.C. ANRIL: A lncRNA at the CDKN2A/B locus with roles in cancer and metabolic disease. Front. Endocrinol. (Lausanne) 2018, 9, 405. [Google Scholar] [CrossRef]
- Li, Q.; Tian, Y.; Hu, G.; Liang, Y.; Bai, W.; Li, H. Highly expressed antisense noncoding RNA in the INK4 locus promotes growth and invasion of renal clear carcinoma cells via the β-Catenin pathway. Oncol. Res. 2017, 25, 1373–1382. [Google Scholar] [CrossRef]
- Sun, Y.; Ma, L. New insights into long non-coding RNA MALAT1 in cancer and metastasis. Cancers 2019, 11, 216. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, W.; Gu, W.; Yan, Y.; Yao, X.; Zheng, J. MALAT1 accelerates the development and progression of renal cell carcinoma by decreasing the expression of miR-203 and promoting the expression of BIRC5. Cell Prolif. 2019, 52, e12640. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, Y.; Gu, L.; Li, X.; Gao, Y.; Lyu, X.; Chen, L.; Luo, G.; Wang, L.; Xie, Y.; et al. LncRNAs act as prognostic and diagnostic biomarkers in renal cell carcinoma: A systematic review and meta-analysis. Oncotarget 2016, 7, 74325–74336. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Duan, W.; Jiang, X.; Zhao, L.; Li, J.; Wang, R.; Yan, S.; Xie, Y.; Yan, K.; Wang, Q.; et al. Cell-free lncRNA expression signatures in urine serve as novel non-invasive biomarkers for diagnosis and recurrence prediction of bladder cancer. J. Cell Mol. Med. 2018, 22, 2838–2845. [Google Scholar] [CrossRef] [PubMed]
- von Siebenthal, M.; Besic, M.; Gheinani, A.H.; Akshay, A.; Lizun-Platoni, S.; Kunz, N.; Bukhard, F.C.; Monastyrskaya, K. Urinary miRNA profiles discriminate between obstruction-induced bladder dysfunction and healthy controls. Sci. Rep. 2021, 11, 10204. [Google Scholar] [CrossRef] [PubMed]
- Metzinger-Le Meuth, V.; Burtey, S.; Maitrias, P.; Massy, Z.A.; Metzinger, L. microRNAs in the pathophysiology of CKD-MBD: Biomarkers and innovative drugs. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Tabet, F.; Vickers, K.C.; Cuesta Torres, L.F.; Wiese, C.B.; Shoucri, B.M.; Lambert, G.; Catherinet, C.; Prado-Lourenco, L.; Levin, M.G.; Thacker, S.; et al. HDL-transferred microRNA-223 regulates ICAM-1 expression in endothelial cells. Nat. Commun. 2014, 5, 3292. [Google Scholar] [CrossRef] [PubMed]


| ncRNAs | Prediction/Biomarker | Reference |
|---|---|---|
| Urinary exosomal ncRNAs -miRNA-181a -9 antisense RNAs (EAF1-AS1, PCBP1-AS1, RP11-178F10.1, RP11-315I20.1, RP11-378E13.4, RP11-68I3.2, RP11-700F16.3, RP11-98D18.1 and RP11-1382.1) -trfVal, trfLeu | Diagnostic biomarker for early detection of CKD | [83] |
| LncRNA NOP14-AS1 and LncRNA HCP5 | Predicting CKD progression | [84] |
| LncRNA DKFZP434I0714 | Predicting adverse CV outcomes in ESRD patients | [85] |
| LncRNA LIPCAR | Predicting HF outcomes in patients without CKD | [86] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno, J.A.; Hamza, E.; Guerrero-Hue, M.; Rayego-Mateos, S.; García-Caballero, C.; Vallejo-Mudarra, M.; Metzinger, L.; Metzinger-Le Meuth, V. Non-Coding RNAs in Kidney Diseases: The Long and Short of Them. Int. J. Mol. Sci. 2021, 22, 6077. https://doi.org/10.3390/ijms22116077
Moreno JA, Hamza E, Guerrero-Hue M, Rayego-Mateos S, García-Caballero C, Vallejo-Mudarra M, Metzinger L, Metzinger-Le Meuth V. Non-Coding RNAs in Kidney Diseases: The Long and Short of Them. International Journal of Molecular Sciences. 2021; 22(11):6077. https://doi.org/10.3390/ijms22116077
Chicago/Turabian StyleMoreno, Juan Antonio, Eya Hamza, Melania Guerrero-Hue, Sandra Rayego-Mateos, Cristina García-Caballero, Mercedes Vallejo-Mudarra, Laurent Metzinger, and Valérie Metzinger-Le Meuth. 2021. "Non-Coding RNAs in Kidney Diseases: The Long and Short of Them" International Journal of Molecular Sciences 22, no. 11: 6077. https://doi.org/10.3390/ijms22116077
APA StyleMoreno, J. A., Hamza, E., Guerrero-Hue, M., Rayego-Mateos, S., García-Caballero, C., Vallejo-Mudarra, M., Metzinger, L., & Metzinger-Le Meuth, V. (2021). Non-Coding RNAs in Kidney Diseases: The Long and Short of Them. International Journal of Molecular Sciences, 22(11), 6077. https://doi.org/10.3390/ijms22116077