
Histone Acetylation Dynamics during In Vivo and In Vitro Oocyte Aging in Common Carp Cyprinus carpio

 ,
,  ,
, 

Abstract
1. Introduction
2. Results
2.1. Egg Quality Indices
2.2. Image Analysis of 2D AUT × SDS PAGE
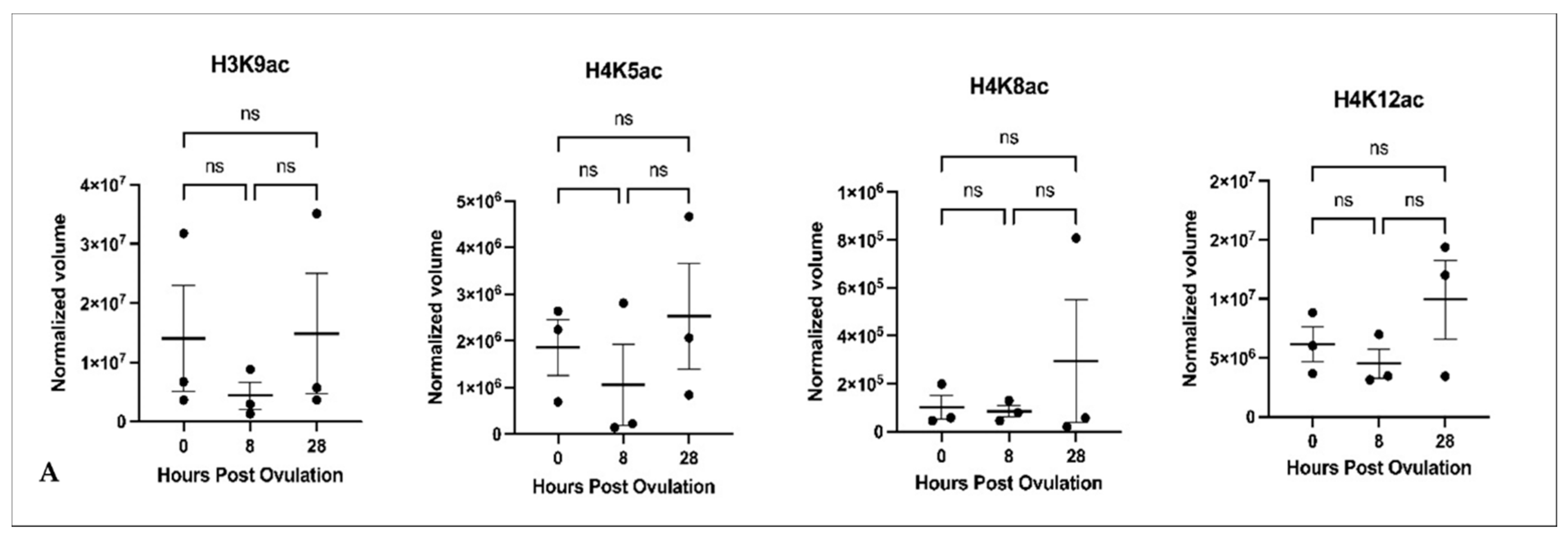
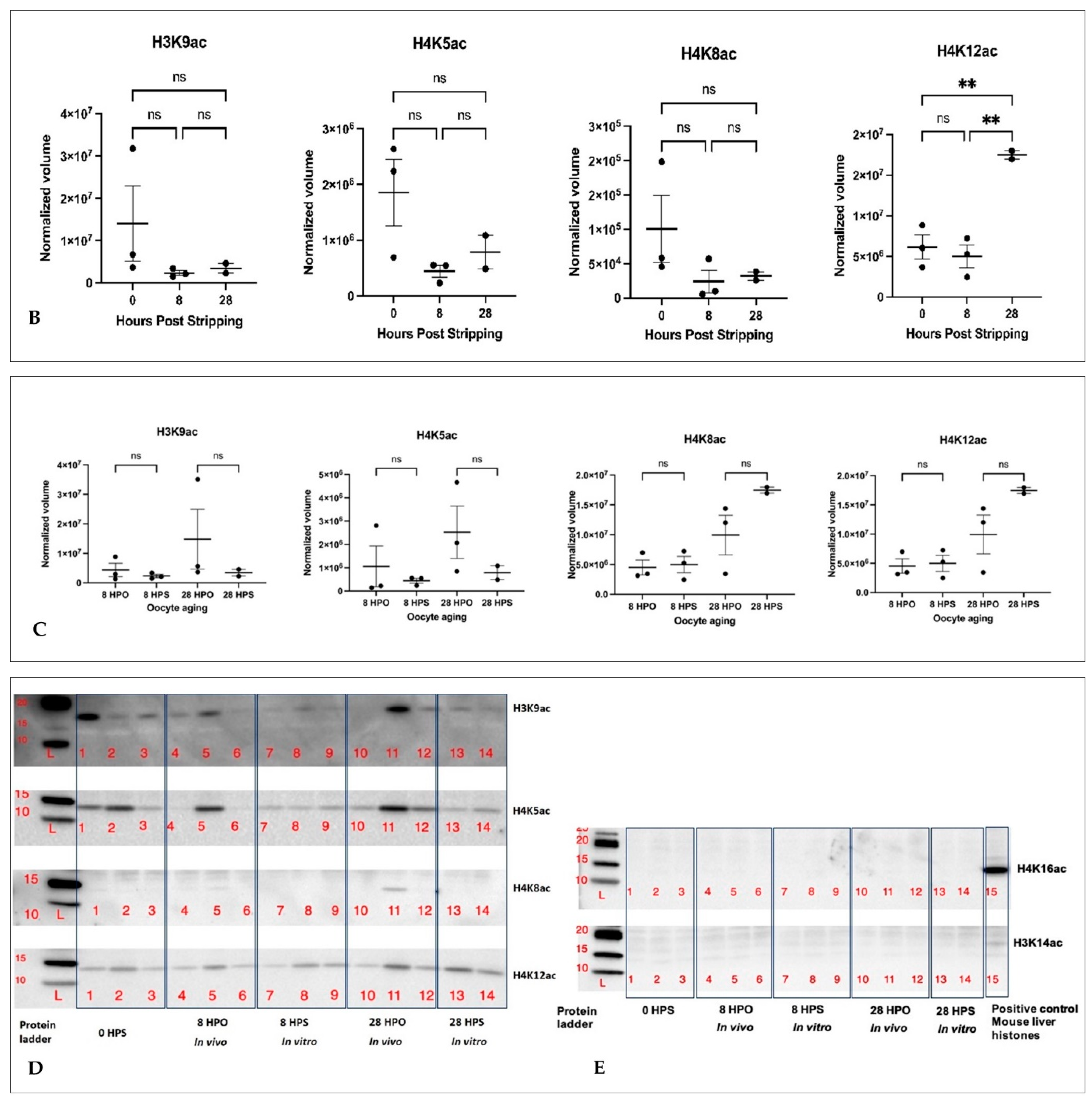
2.3. Histone Acetylation during In Vivo and In Vitro Oocyte Aging
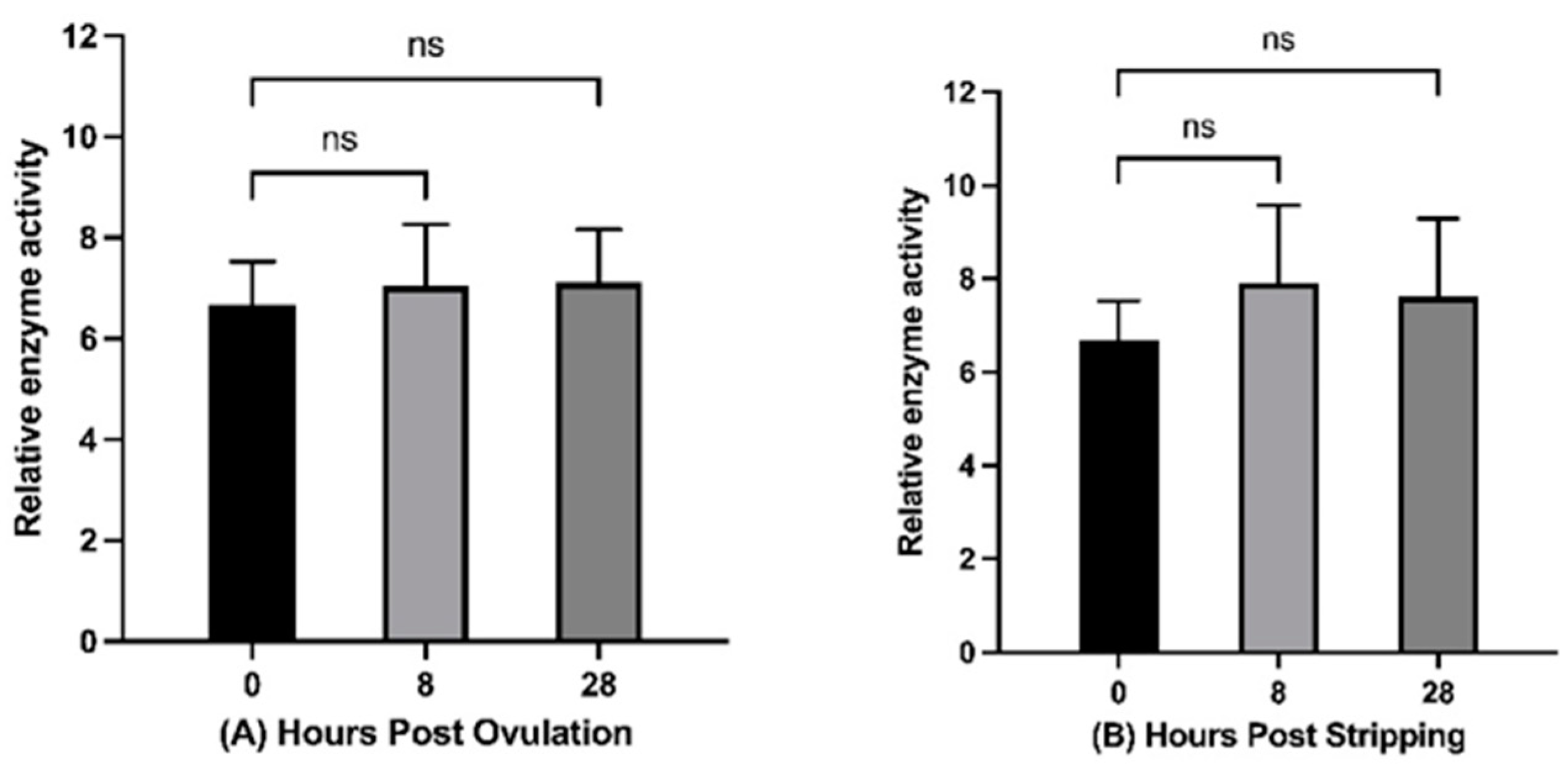
2.4. Histone Acetyltransferase Activity during In Vivo and In Vitro Oocyte Aging
3. Discussion
4. Materials and Methods
4.1. Egg Storage and Quality Assessment
4.1.1. Experimental Fish Preparation
4.1.2. In Vivo and In Vitro Oocyte Aging
4.1.3. Artificial Fertilisation and Egg Developmental Success
4.2. Histone Modifications
4.2.1. Sample Collection for Histone Modification Analysis
4.2.2. Histone Isolation
4.2.3. Separation of Histone Modifications
4.2.4. Immunodetection of Specific Histone Acetylation
4.3. Histone Acetyltransferase Activity Assay
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Petri, T.; Dankert, D.; Demond, H.; Wennemuth, G.; Horsthemke, B.; Grummer, R. In vitro postovulatory oocyte aging affects H3K9 trimethylation in two-cell embryos after IVF. Ann. Anat. 2020, 227, 151424. [Google Scholar] [CrossRef]
- Reading, B.; Andersen, L.; Ryu, Y.-W.; Mushirobira, Y.; Todo, T.; Hiramatsu, N. Oogenesis and Egg Quality in Finfish: Yolk Formation and Other Factors Influencing Female Fertility. Fishes 2018, 3, 45. [Google Scholar] [CrossRef]
- Aegerter, S.; Jalabert, B. Effects of post-ovulatory oocyte ageing and temperature on egg quality and on the occurrence of triploid fry in rainbow trout, Oncorhynchus mykiss. Aquaculture 2004, 231, 59–71. [Google Scholar] [CrossRef]
- Samarin, A.M.; Policar, T.; Lahnsteiner, F. Fish Oocyte Ageing and its Effect on Egg Quality. Rev. Fish. Sci. Aquac. 2015, 23, 302–314. [Google Scholar] [CrossRef]
- Flanagan, J.M.; Popendikyte, V.; Pozdniakovaite, N.; Sobolev, M.; Assadzadeh, A.; Schumacher, A.; Zangeneh, M.; Lau, L.; Virtanen, C.; Wang, S.C.; et al. Intra- and interindividual epigenetic variation in human germ cells. Am. J. Hum. Genet. 2006, 79, 67–84. [Google Scholar] [CrossRef]
- Goldberg, A.D.; Allis, C.D.; Bernstein, E. Epigenetics: A landscape takes shape. Cell 2007, 128, 635–638. [Google Scholar] [CrossRef]
- Bobe, J.; Labbe, C. Egg and sperm quality in fish. Gen. Comp. Endocrinol. 2010, 165, 535–548. [Google Scholar] [CrossRef]
- Gonzalo, S. Epigenetic alterations in aging. J. Appl. Physiol. 2010, 109, 586–597. [Google Scholar] [CrossRef]
- Aegerter, S.; Jalabert, B.; Bobe, J. Large scale real-time PCR analysis of mRNA abundance in rainbow trout eggs in relationship with egg quality and post-ovulatory ageing. Mol. Reprod. Dev. 2005, 72, 377–385. [Google Scholar] [CrossRef]
- Bonnet, E.; Fostier, A.; Bobe, J. Microarray-based analysis of fish egg quality after natural or controlled ovulation. BMC Genom. 2007, 8, 55. [Google Scholar] [CrossRef]
- Mommens, M.; Fernandes, J.M.; Bizuayehu, T.T.; Bolla, S.L.; Johnston, I.A.; Babiak, I. Maternal gene expression in Atlantic halibut (Hippoglossus hippoglossus L.) and its relation to egg quality. BMC Res. Notes 2010, 3, 138. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Weber, G.M.; Hostuttler, M.A.; Wei, H.; Wang, L.; Yao, J. MicroRNA expression profiles from eggs of different qualities associated with post-ovulatory ageing in rainbow trout (Oncorhynchus mykiss). BMC Genom. 2015, 16, 201. [Google Scholar] [CrossRef]
- Bizuayehu, T.T.; Mommens, M.; Sundaram, A.Y.M.; Dhanasiri, A.K.S.; Babiak, I. Postovulatory maternal transcriptome in Atlantic salmon and its relation to developmental potential of embryos. BMC Genom. 2019, 20, 315. [Google Scholar] [CrossRef]
- Samarin, A.M.; Samarin, A.M.; Ostbye, T.K.; Ruyter, B.; Sampels, S.; Burkina, V.; Blecha, M.; Policar, T. The possible involvement of oxidative stress in the oocyte ageing process in goldfish Carassius auratus (Linnaeus, 1758). Sci. Rep. 2019, 9, 10469. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Ma, J.; Schatten, H.; Sun, Q. Epigenetic changes associated with oocyte aging. Sci. China Life Sci. 2012, 55, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Ge, Z.J.; Schatten, H.; Zhang, C.L.; Sun, Q.Y. Oocyte ageing and epigenetics. Reproduction 2015, 149, R103–R114. [Google Scholar] [CrossRef]
- Heinzmann, J.; Mattern, F.; Aldag, P.; Bernal-Ulloa, S.M.; Schneider, T.; Haaf, T.; Niemann, H. Extended in vitro maturation affects gene expression and DNA methylation in bovine oocytes. Mol. Hum. Reprod. 2015, 21, 770–782. [Google Scholar] [CrossRef]
- Cui, M.S.; Wang, X.L.; Tang, D.W.; Zhang, J.; Liu, Y.; Zeng, S.M. Acetylation of H4K12 in porcine oocytes during in vitro aging: Potential role of ooplasmic reactive oxygen species. Theriogenology 2011, 75, 638–646. [Google Scholar] [CrossRef]
- Huang, J.C.; Yan, L.Y.; Lei, Z.L.; Miao, Y.L.; Shi, L.H.; Yang, J.W.; Wang, Q.; Ouyang, Y.C.; Sun, Q.Y.; Chen, D.Y. Changes in histone acetylation during postovulatory aging of mouse oocyte. Biol. Reprod. 2007, 77, 666–670. [Google Scholar] [CrossRef]
- Labbé, C.; Robles, V.; Herraez, M.P. Epigenetics in fish gametes and early embryo. Aquaculture 2017, 472, 93–106. [Google Scholar] [CrossRef]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Luger, K.; Mader, A.W.; Richmond, R.K.; Sargent, D.F.; Richmond, T.J. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 1997, 389, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Rothbart, S.B.; Strahl, B.D. Interpreting the language of histone and DNA modifications. Biochim. Biophys. Acta 2014, 1839, 627–643. [Google Scholar] [CrossRef] [PubMed]
- Gavazzo, P.; Vergani, L.; Mascetti, G.C.; Nicolini, C. Effects of histone acetylation on chromatin structure. J. Cell. Biochem. 1997, 64, 466–475. [Google Scholar] [CrossRef]
- Turner, B.M. Histone acetylation and an epigenetic code. Bioessays 2000, 22, 836–845. [Google Scholar] [CrossRef]
- Kurdistani, S.K.; Grunstein, M. Histone acetylation and deacetylation in yeast. Nat. Rev. Mol. Cell Biol. 2003, 4, 276–284. [Google Scholar] [CrossRef]
- Shahbazian, M.D.; Grunstein, M. Functions of Site-Specific Histone Acetylation and Deacetylation. Annu. Rev. Biochem. 2007, 76, 75–100. [Google Scholar] [CrossRef]
- Zhang, G.M.; Gu, C.H.; Zhang, Y.L.; Sun, H.Y.; Qian, W.P.; Zhou, Z.R.; Wan, Y.J.; Jia, R.X.; Wang, L.Z.; Wang, F. Age-associated changes in gene expression of goat oocytes. Theriogenology 2013, 80, 328–336. [Google Scholar] [CrossRef]
- Linhart, O.; Kudo, S.; Billard, R.; Slechta, V.; Mikodina, E.V. Morphology, Composition and Fertilization of Carp Eggs—A Review. Aquaculture 1995, 129, 75–93. [Google Scholar] [CrossRef]
- Strahl, B.D.; Allis, C.D. The language of covalent histone modifications. Nature 2000, 403, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef]
- Bloushtain-Qimron, N.; Yao, J.; Shipitsin, M.; Maruyama, R.; Polyak, K. Epigenetic patterns of embryonic and adult stem cells. Cell Cycle 2009, 8, 809–817. [Google Scholar] [CrossRef]
- Wang, N.; Tilly, J.L. Epigenetic status determines germ cell meiotic commitment in embryonic and postnatal mammalian gonads. Cell Cycle 2010, 9, 339–349. [Google Scholar] [CrossRef]
- Gu, L.; Wang, Q.; Sun, Q.Y. Histone modifications during mammalian oocyte maturation: Dynamics, regulation and functions. Cell Cycle 2010, 9, 1942–1950. [Google Scholar] [CrossRef]
- Cunliffe, V.T. Histone modifications in zebrafish development. Methods Cell Biol. 2016, 135, 361–385. [Google Scholar] [CrossRef]
- Turner, B.M. Defining an epigenetic code. Nat. Cell. Biol. 2007, 9, 2–6. [Google Scholar] [CrossRef]
- Xing, X.; Zhang, J.; Wu, T.; Zhang, J.; Wang, Y.; Su, J.; Zhang, Y. SIRT1 reduces epigenetic and non-epigenetic changes to maintain the quality of postovulatory aged oocytes in mice. Exp. Cell Res. 2021, 399, 112421. [Google Scholar] [CrossRef]
- Liu, N.; Wu, Y.G.; Lan, G.C.; Sui, H.S.; Ge, L.; Wang, J.Z.; Liu, Y.; Qiao, T.W.; Tan, J.H. Pyruvate prevents aging of mouse oocytes. Reproduction 2009, 138, 223–234. [Google Scholar] [CrossRef]
- Trapphoff, T.; Heiligentag, M.; Dankert, D.; Demond, H.; Deutsch, D.; Frohlich, T.; Arnold, G.J.; Grummer, R.; Horsthemke, B.; Eichenlaub-Ritter, U. Postovulatory aging affects dynamics of mRNA, expression and localization of maternal effect proteins, spindle integrity and pericentromeric proteins in mouse oocytes. Hum. Reprod. 2016, 31, 133–149. [Google Scholar] [CrossRef]
- Samarin, A.M.; Samarin, A.M.; Ostbye, T.K.; Ruyter, B.; Sampels, S.; Burkina, V.; Blecha, M.; Gela, D.; Policar, T. Alteration of mRNA abundance, oxidation products and antioxidant enzyme activities during oocyte ageing in common carp Cyprinus carpio. PLoS ONE 2019, 14, e0212694. [Google Scholar] [CrossRef]
- Shang, W.-H.; Hori, T.; Westhorpe, F.G.; Godek, K.M.; Toyoda, A.; Misu, S.; Monma, N.; Ikeo, K.; Carroll, C.W.; Takami, Y.; et al. Acetylation of histone H4 lysine 5 and 12 is required for CENP-A deposition into centromeres. Nat. Commun. 2016, 7, 13465. [Google Scholar] [CrossRef] [PubMed]
- Régnier, V.; Vagnarelli, P.; Fukagawa, T.; Zerjal, T.; Burns, E.; Trouche, D.; Earnshaw, W.; Brown, W. CENP-A Is Required for Accurate Chromosome Segregation and Sustained Kinetochore Association of BubR1. Mol. Cell. Biol. 2005, 25, 3967–3981. [Google Scholar] [CrossRef] [PubMed]
- Ruan, K.; Yamamoto, T.G.; Asakawa, H.; Chikashige, Y.; Kimura, H.; Masukata, H.; Haraguchi, T.; Hiraoka, Y. Histone H4 acetylation required for chromatin decompaction during DNA replication. Sci. Rep. 2015, 5, 12720. [Google Scholar] [CrossRef]
- Demond, H.; Dankert, D.; Grümmer, R.; Horsthemke, B. Preovulatory oocyte aging in mice affects fertilization rate and embryonic genome activation. bioRxiv 2017. [Google Scholar] [CrossRef]
- Schulz, K.N.; Harrison, M.M. Mechanisms regulating zygotic genome activation. Nat. Rev. Genet. 2019, 20, 221–234. [Google Scholar] [CrossRef]
- Sato, Y.; Hilbert, L.; Oda, H.; Wan, Y.; Heddleston, J.M.; Chew, T.L.; Zaburdaev, V.; Keller, P.; Lionnet, T.; Vastenhouw, N.; et al. Histone H3K27 acetylation precedes active transcription during zebrafish zygotic genome activation as revealed by live-cell analysis. Development 2019, 146. [Google Scholar] [CrossRef]
- Li, X.Y.; Harrison, M.M.; Villalta, J.E.; Kaplan, T.; Eisen, M.B. Establishment of regions of genomic activity during the Drosophila maternal to zygotic transition. eLife 2014, 3, e03737. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.J.; Wang, K.; Miao, D.Q.; Guo, L.; Hou, Y.; Schatten, H.; Sun, Q.Y. Protein profile changes during porcine oocyte aging and effects of caffeine on protein expression patterns. PLoS ONE 2011, 6, e28996. [Google Scholar] [CrossRef][Green Version]
- Zhang, T.; Zhou, Y.; Li, L.; Wang, H.H.; Ma, X.S.; Qian, W.P.; Shen, W.; Schatten, H.; Sun, Q.Y. SIRT1, 2, 3 protect mouse oocytes from postovulatory aging. Aging 2016, 8, 685–696. [Google Scholar] [CrossRef]
- Hamatani, T.; Falco, G.; Carter, M.G.; Akutsu, H.; Stagg, C.A.; Sharov, A.A.; Dudekula, D.B.; VanBuren, V.; Ko, M.S. Age-associated alteration of gene expression patterns in mouse oocytes. Hum. Mol. Genet. 2004, 13, 2263–2278. [Google Scholar] [CrossRef]
- Samarin, A.M.; Gela, D.; Bytyutskyy, D.; Policar, T. Determination of the best post-ovulatory stripping time for the common carp (Cyprinus carpio Linnaeus, 1758). J. Appl. Ichthyol. 2015, 31, 51–55. [Google Scholar] [CrossRef]
- Horváth, L.; Tamás, G.; Coche, A. Common Carp: Mass Production of Eggs and Early Fry; Food and Agriculture Organization of the United Nations: Rome, Italy, 1985; p. 44. [Google Scholar]
- Wu, N.; Yue, H.M.; Chen, B.; Gui, J.F. Histone H2A has a novel variant in fish oocytes. Biol Reprod 2009, 81, 275–283. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Shechter, D.; Dormann, H.L.; Allis, C.D.; Hake, S.B. Extraction, purification and analysis of histones. Nat. Protoc. 2007, 2, 1445–1457. [Google Scholar] [CrossRef]
- Green, G.R.; Do, D.P. Purification and analysis of variant and modified histones using 2D PAGE. Methods Mol. Biol. 2009, 464, 285–302. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| In Vivo Oocyte Aging | ||||||||||||||
| HPO | 0 | 2 | 4 | 6 | 8 | 10 | 12 | 28 | ||||||
| Eyeing % | 88 ± 9 a | 91 ± 5 a | 77 ± 16 ab | 66 ± 30 ab | 46 ± 25 bc | 15 ± 11 cd | 3 ± 4 d | 0 ± 0 d | ||||||
| Hatching % | 83 ± 16 a | 88 ± 10 a | 71 ± 21 a | 59 ± 31 ab | 31 ± 19 bc | 6 ± 5 c | 0.7 ± 1 c | 0 ± 0 c | ||||||
| In Vitro Oocyte Aging | ||||||||||||||
| HPO | 0 | 2 | 4 | 6 | 8 | 10 | 12 | 28 | ||||||
| Eyeing % | 95 ± 4 a | 94 ± 3 a | 92 ± 3 a | 84 ± 8 a | 62 ± 6 b | 40 ± 5 c | 0 ± 0 d | |||||||
| Hatching % | 94 ± 4 a | 92 ± 1 a | 88 ± 5 a | 68 ± 15 ab | 36 ± 22 bc | 21 ± 9 cd | 0 ± 0 d | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Waghmare, S.G.; Samarin, A.M.; Samarin, A.M.; Danielsen, M.; Møller, H.S.; Policar, T.; Linhart, O.; Dalsgaard, T.K. Histone Acetylation Dynamics during In Vivo and In Vitro Oocyte Aging in Common Carp Cyprinus carpio. Int. J. Mol. Sci. 2021, 22, 6036. https://doi.org/10.3390/ijms22116036
Waghmare SG, Samarin AM, Samarin AM, Danielsen M, Møller HS, Policar T, Linhart O, Dalsgaard TK. Histone Acetylation Dynamics during In Vivo and In Vitro Oocyte Aging in Common Carp Cyprinus carpio. International Journal of Molecular Sciences. 2021; 22(11):6036. https://doi.org/10.3390/ijms22116036
Chicago/Turabian StyleWaghmare, Swapnil Gorakh, Azin Mohagheghi Samarin, Azadeh Mohagheghi Samarin, Marianne Danielsen, Hanne Søndergård Møller, Tomáš Policar, Otomar Linhart, and Trine Kastrup Dalsgaard. 2021. "Histone Acetylation Dynamics during In Vivo and In Vitro Oocyte Aging in Common Carp Cyprinus carpio" International Journal of Molecular Sciences 22, no. 11: 6036. https://doi.org/10.3390/ijms22116036
APA StyleWaghmare, S. G., Samarin, A. M., Samarin, A. M., Danielsen, M., Møller, H. S., Policar, T., Linhart, O., & Dalsgaard, T. K. (2021). Histone Acetylation Dynamics during In Vivo and In Vitro Oocyte Aging in Common Carp Cyprinus carpio. International Journal of Molecular Sciences, 22(11), 6036. https://doi.org/10.3390/ijms22116036