
Roles of Plant Glycine-Rich RNA-Binding Proteins in Development and Stress Responses



Abstract
1. Introduction
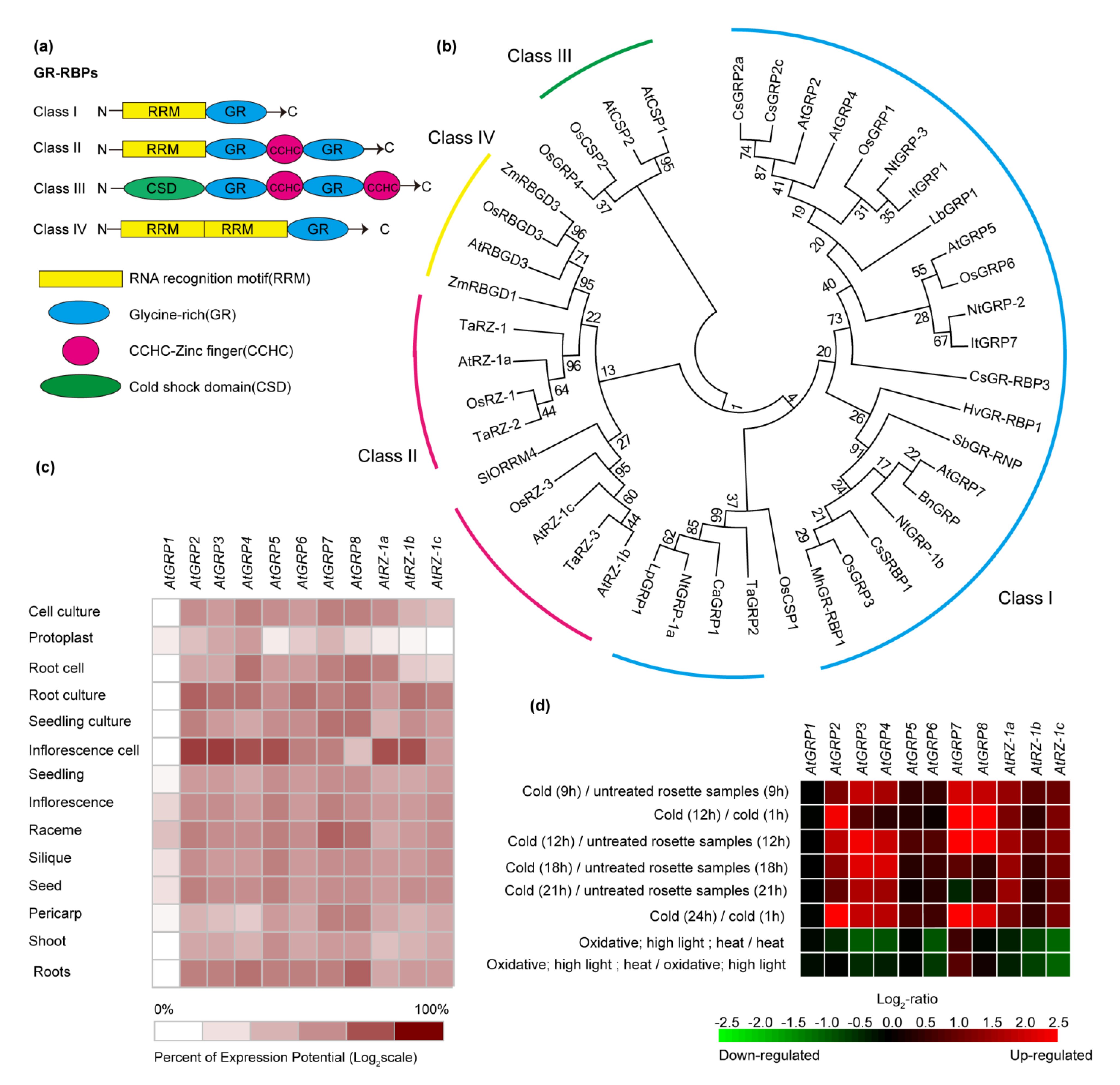
2. The GR-RBP Family
3. Roles of GR-RBPs in Plant Growth and Development
3.1. GR-RBPs in Stress-Related Seed Germination
3.2. GR-RBP Function during Vegetative Growth
3.3. GR-RBPs and Reproductive Growth
4. GR-RBPs and Stress Responses
4.1. Temperature Treatments
4.2. Salinity Stress
4.3. Drought Stress
4.4. Biotic Stress
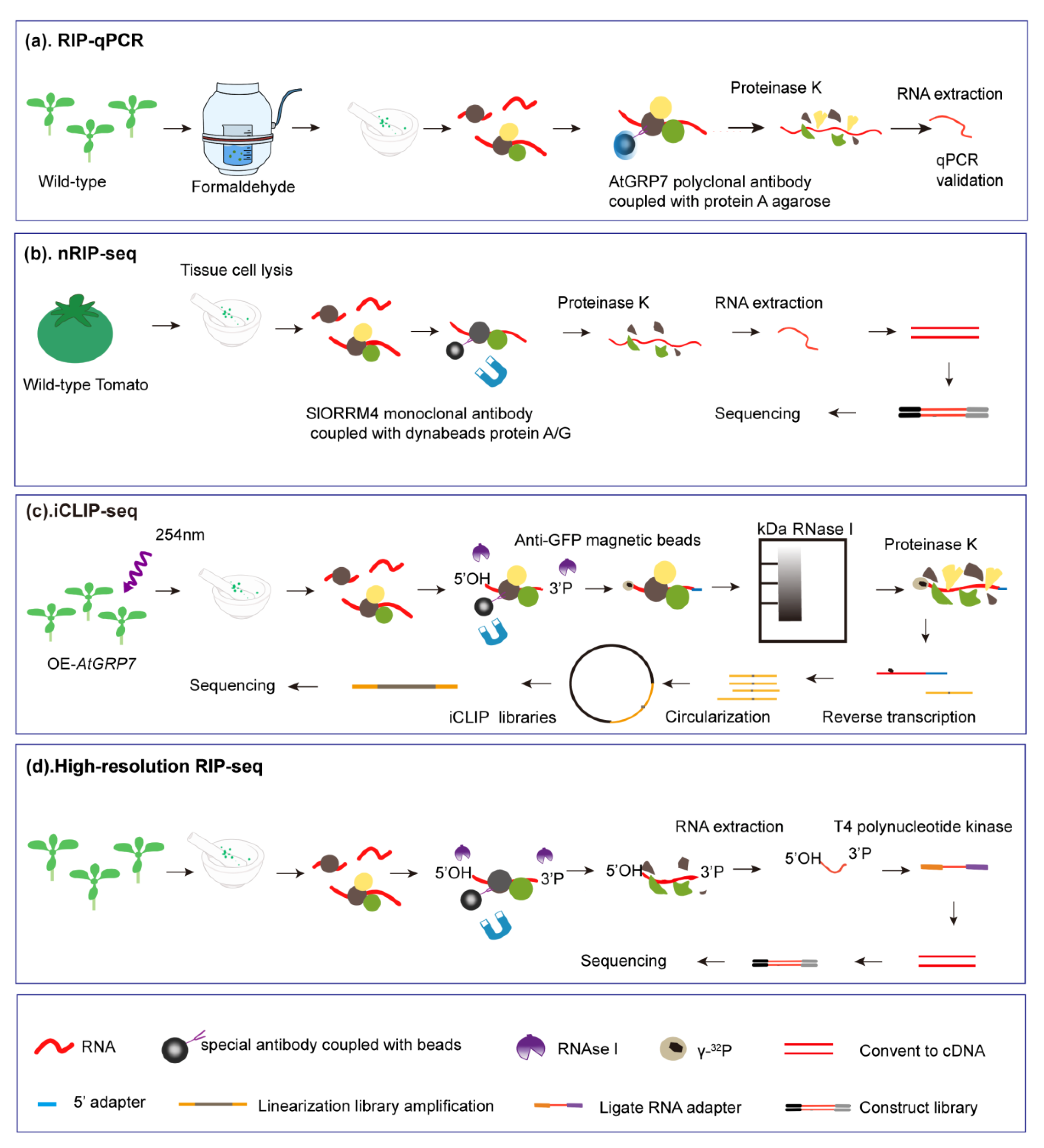
5. Technologies Used to Investigate RNA–GR-RBP Interactions
5.1. RIP-qPCR
5.2. ICLIP-Seq
5.3. High-Resolution RIP-Seq
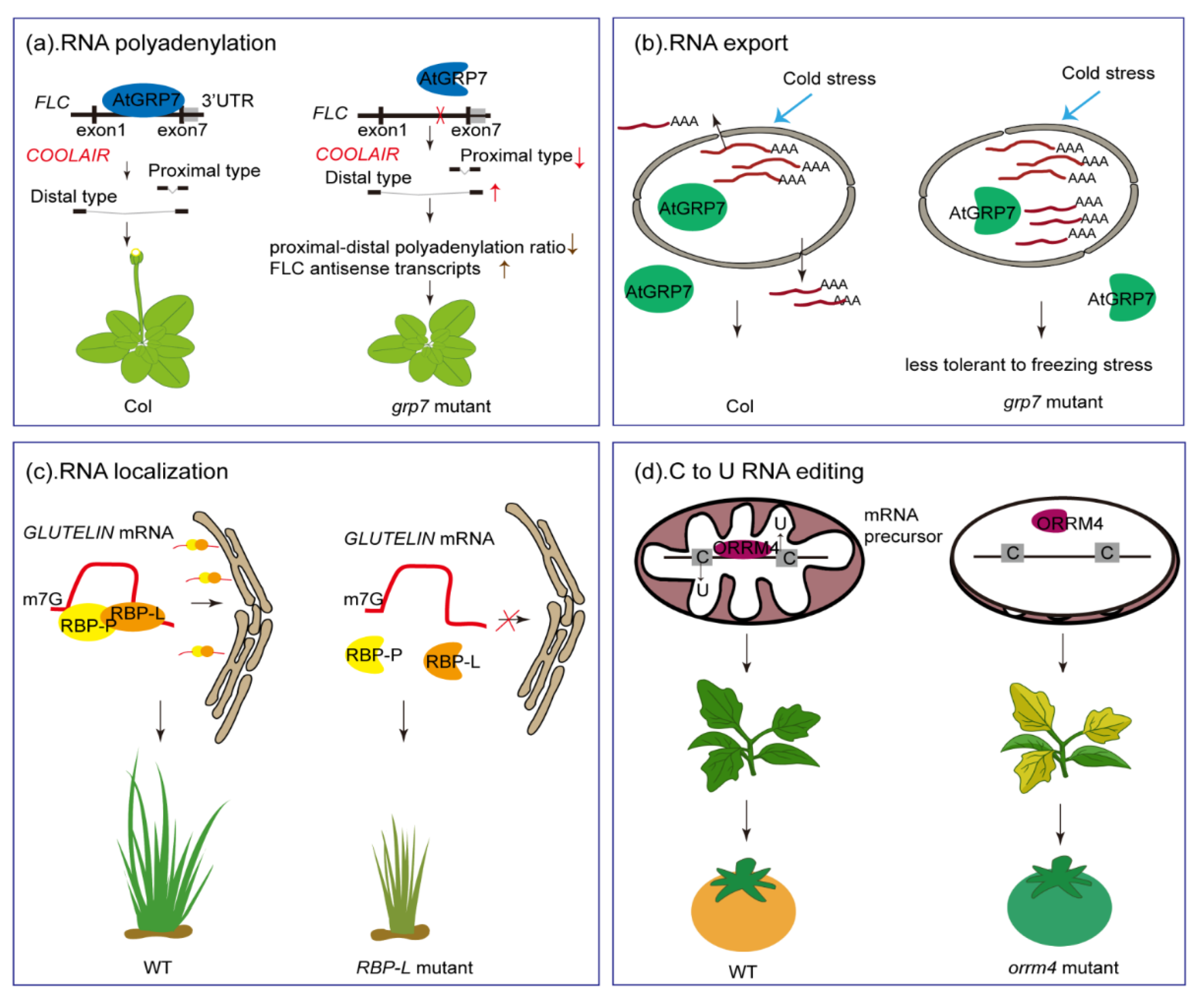
6. Post-Transcriptional RNA Regulation by GR-RBPs
6.1. GR-RBP Function in AS and Polyadenylation
6.2. The Role of GR-RBPs in RNA Export
6.3. RNA Localization
6.4. The Role of GR-RBPs in C-to-U RNA Editing
7. Conclusions and Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, E.G.; Schroeder, J.I. Genetic strategies for improving crop yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Sachetto-Martins, G.; Franco, L.O.; Oliveira, D.E.D. Plant glycine-rich proteins: A family or just proteins with a common motif? Biochimica. Biophys. Acta. 2000, 1492, 1–14. [Google Scholar] [CrossRef]
- Nocker, S.V.; Vierstra, R.D. Two cDNAs from Arabidopsis thaliana encode putative RNA binding proteins containing glycine-rich domains. Plant Mol. Biol. 1993, 21, 695–699. [Google Scholar] [CrossRef]
- Chen, X.; Zeng, Q.C.; Lu, X.P.; Yu, D.Q.; Li, W.Z. Characterization and Expression Analysis of Four Glycine-Rich RNA-Binding Proteins Involved in Osmotic Response in Tobacco (Nicotiana tabacum cv. Xanthi). Agric. Sci. China 2010, 9, 1577–1587. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, W.Y.; Kwak, K.J.; Oh, S.H.; Han, Y.S.; Kang, H. Glycine-rich RNA-binding proteins are functionally conserved in Arabidopsis thaliana and Oryza sativa during cold adaptation process. J. Exp. Bot. 2010, 61, 2317–2325. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, Y.; Xiao, H.; Zheng, Y.; Yue, B. Genome-wide identification, evolution, and expression analysis of RNA-binding glycine-rich protein family in maize. J. Integr. Plant Biol. 2014, 56, 1020–1031. [Google Scholar] [CrossRef]
- Lu, Y.; Sun, J.; Yang, Z.; Zhao, C.; Xu, T. Genome-wide identification and expression analysis of glycine-rich RNA-binding protein family in sweet potato wild relative Ipomoea trifida. Gene 2018, 686, 177–186. [Google Scholar] [CrossRef]
- Kwak, K.J.; Kang, H.; Han, K.H.; Ahn, S.J. Molecular cloning, characterization, and stress-responsive expression of genes encoding glycine-rich RNA-binding proteins in Camelina sativa L. Plant Physiol. Biochem. 2013, 68, 44–51. [Google Scholar] [CrossRef]
- Köster, T.; Reichel, M.; Staiger, D. CLIP and RNA interactome studies to unravel genome-wide RNA-protein interactions in vivo in Arabidopsis thaliana. Methods 2020, 178, 63–71. [Google Scholar] [CrossRef]
- Huppertz, I.; Attig, J.; D’Ambrogio, A.; Easton, L.E.; Sibley, C.R.; Sugimoto, Y.; Tajnik, M.; Kônig, J.; Ule, J. iCLIP: Protein–RNA interactions at nucleotide resolution. Methods 2014, 65, 274–287. [Google Scholar] [CrossRef] [PubMed]
- Mangeon, A.; Junqueira, R.M.; Sachetto-Martins, G. Functional diversity of the plant glycine-rich proteins superfamily. Plant Signal. Behav. 2010, 5, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, P.; Kim, J.A.; Jeong, M.J.; Kang, C.H.; Lee, S.I. Defining the RNA-binding glycine-rich (RBG) gene superfamily: New insights into nomenclature, phylogeny, and evolutionary trends obtained by genome-wide comparative analysis of Arabidopsis, Chinese cabbage, rice and maize genomes. Mol. Genet. Genom. 2015, 290, 2279–2295. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, W.Y.; Kwak, K.J.; Oh, S.H.; Kang, H. Zinc finger-containing glycine-rich RNA-binding protein in Oryza sativa has an RNA chaperone activity under cold stress conditions. Plant Cell Environ. 2010, 33, 759–768. [Google Scholar] [PubMed]
- Fusaro, A.F.; Bocca, S.N.; Ramos, R.L.B.; Barrôco, R.M.; Magioli, C.; Jorge, V.C.; Coutinho, T.C.; Rangel-Lima, C.M.; De Rycke, R.; Inzé, D.; et al. AtGRP2, a cold-induced nucleo-cytoplasmic RNA-binding protein, has a role in flower and seed development. Planta 2007, 225, 1339–1351. [Google Scholar] [CrossRef] [PubMed]
- Juntawong, P.; Sorenson, R.; Bailey-Serres, J. Cold shock protein 1 chaperones mRNAs during translation in Arabidopsis thaliana. Plant J. 2013, 74, 1016–1028. [Google Scholar] [CrossRef]
- Karlson, D.; Imai, R. Conservation of the cold shock domain protein family in plants. Plant Physiol. 2003, 131, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Karlson, D.T. Overexpression of AtCSP4 affects late stages of embryo development in Arabidopsis. J. Exp. Bot. 2011, 62, 2079–2091. [Google Scholar] [CrossRef]
- Kim, J.Y.; Park, S.J.; Jang, B.; Jung, C.H.; Ahn, S.J.; Goh, C.H.; Cho, K.; Han, O.; Kang, H. Functional characterization of a glycine-rich RNA-binding protein 2 in Arabidopsis thaliana under abiotic stress conditions. Plant J. 2007, 50, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Kwak, K.J.; Kim, Y.O.; Kang, H. Characterization of transgenic Arabidopsis plants overexpressing GR-RBP4 under high salinity, dehydration, or cold stress. J. Exp. Bot. 2005, 56, 3007–3016. [Google Scholar] [CrossRef]
- Kim, Y.O.; Kim, J.S.; Kang, H. Cold-inducible zinc finger-containing glycine-rich RNA-binding protein contributes to the enhancement of freezing tolerance in Arabidopsis thaliana. Plant J. 2005, 42, 890–900. [Google Scholar] [CrossRef]
- Kim, Y.O.; Pan, S.; Jung, C.H.; Kang, H. A zinc finger-containing glycine-rich RNA-binding protein, atRZ-1a, has a negative impact on seed germination and seedling growth of Arabidopsis thaliana under salt or drought stress conditions. Plant Cell Physiol. 2007, 48, 1170–1181. [Google Scholar] [CrossRef]
- Kim, W.Y.; Kim, J.Y.; Jung, H.J.; Oh, S.H.; Han, Y.S.; Kang, H. Comparative analysis of Arabidopsis zinc finger-containing glycine-rich RNA-binding proteins during cold adaptation. Plant Physiol. Biochem. 2010, 48, 866–872. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Gu, L.; Choi, M.J.; Kim, R.J.; Suh, M.C.; Kang, H. Comparative functional analysis of wheat (Triticum aestivum). zinc finger-containing glycine-rich RNA-binding proteins in response to abiotic stresses. PLoS ONE 2014, 9, e96877. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Qin, Y.; Li, Y.; Li, M.; Ma, F. Overexpression of MpGR-RBP1, a glycine-rich RNA-binding protein gene from Malus prunifolia (Willd.) Borkh., confers salt stress tolerance and protects against oxidative stress in Arabidopsis. Plant Cell Tissue Organ Cult. 2014, 119, 635–646. [Google Scholar] [CrossRef]
- Shi, X.W.; Bentolila, S.; Hanson, M.R. Organelle RNA recognition motif-containing (ORRM) proteins are plastid and mitochondrial editing factors in Arabidopsis. Plant Signal. Behav. 2016, 11, e1167299. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.W.; Castandet, B.; Germain, A.; Hanson, M.R.; Bentolil, B. ORRM5, an RNA recognition motif-containing protein, has a unique effect on mitochondrial RNA editing. J. Exp. Bot. 2017, 11, 2833–2847. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Amaro, M.A.; Rodríguez-Hernández, A.A.; Rodríguez-Kessler, M.; Hernández-Lucero, E.; Rosales-Mendoza, S.; Ibáñez-Salazar, A.; Delagado-Sánchez, P.; Jiménez-Bremont, J.F. Overexpression of AtGRDP2, a novel glycine-rich domain protein, accelerates plant growth and improves stress tolerance. Front. Plant Sci. 2015, 5, 782. [Google Scholar] [CrossRef]
- Yang, D.H.; Kwak, K.J.; Kim, M.K.; Park, S.J.; Yang, K.Y.; Kang, H. Expression of Arabidopsis glycine-rich RNA-binding protein AtGRP2 or AtGRP7 improves grain yield of rice (Oryza sativa) under drought stress conditions. Plant Sci. Int. J. Exp. Plant Biol. 2014, 214, 106–112. [Google Scholar] [CrossRef]
- Yan, Y.; Ham, B.K.; Chong, Y.H.; Yeh, S.D.; Lucas, W.J. A Plant SMALL RNA-BINDING PROTEIN 1 Family Mediates Cell-to-Cell Trafficking of RNAi Signals. Mol. Plant 2019, 13, 321–335. [Google Scholar] [CrossRef]
- Xiao, J.; Li, C.H.; Xu, S.J.; Xing, L.J.; Chong, K. Jacalin-Lectin Like1 Regulates the Nuclear Accumulation of Glycine-Rich RNA-Binding Protein7, influencing the RNA Processing of Flowering Locus C Antisense Transcripts and Flowering Time in Arabidopsis. Plant Physiol. 2015, 69, 2102–2117. [Google Scholar]
- Kim, J.S.; Park, S.J.; Kwak, J.K.; Kim, Y.O.; Kim, J.Y.; Song, J.Y.; Jang, B.; Jung, C.H.; Kang, H. Cold shock domain proteins and glycine-rich RNA-binding proteins from Arabidopsis thaliana can promote the cold adaptation process in Escherichia coli. Nucleic Acids Res. 2007, 35, 506–516. [Google Scholar] [CrossRef]
- Park, S.J.; Kwak, K.J.; Oh, T.R.; Kim, Y.O.; Kang, H. Cold shock domain proteins affect seed germination and growth of Arabidopsis thaliana under abiotic stress conditions. Plant Cell Physiol. 2009, 50, 869–878. [Google Scholar] [CrossRef]
- Cao, S.Q.; Jiang, L.; Song, S.Y.; Jing, R.; Xu, G.S. AtGRP7 is involved in the regulation of abscisic acid and stress responses in Arabidopsis. Cell. Mol. Biol. Lett. 2006, 11, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.; KoSter, T.; Nolte, C.; Weinholdt, C.; Lewinski, M.; Grosse, I.; Staiger, D. Adaptation of iCLIP to plants determines the binding landscape of the clock-regulated RNA-binding protein AtGRP7. Genome Biol. 2017, 18, 204. [Google Scholar] [CrossRef] [PubMed]
- Staiger, D.; Zecca, L.; Wieczorek-Kirk, D.A.; Apel, K.; Eckstein, L. The circadian clock regulated RNA-binding protein AtGRP7 autoregulates its expression by infuencing alternative splicing of its own pre-mRNA. Plant J. 2003, 33, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.W.; Germain, A.; Hanso, M.R.; Bentolil, B. RNA Recognition Motif-Containing Protein ORRM4 Broadly Affects Mitochondrial RNA Editing and Impacts Plant Development and Flowering. Plant Physiol. 2015, 17, 294–309. [Google Scholar] [CrossRef]
- Yang, Y.F.; Zhu, G.N.; Li, R.; Yan, S.J.; Fu, D.Q.; Zhu, B.Z.; Tian, H.Q.; Luo, Y.B.; Zhu, H.L. The RNA editing factor SlORRM4 is required for normal fruit ripening in tomato. Plant Physiol. 2017, 175, 1690–1702. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.F.; Liu, X.Y.; Wang, K.R.; Li, J.Y.; Zhu, G.N.; Ren, S.; Deng, Z.P.; Zhu, B.Z.; Fu, D.Q.; Luo, Y.B.; et al. Molecular and functional diversity of organelle RNA editing mediated by RNA recognition motif-containing protein ORRM4 in tomato. New Phytol. 2020, 228, 570–585. [Google Scholar] [CrossRef]
- Wu, Z.; Zhu, D.L.; Lin, X.Y.; Miao, J.; Gu, L.F.; Deng, X.; Yang, Q.; Sun, K.T.; Zhu, D.M.; Cao, X.F.; et al. RNA Binding Proteins RZ-1B and RZ-1C play critical roles in regulating pre-mRNA splicing and gene expression during development in Arabidopsis. Plant Cell 2016, 28, 55–73. [Google Scholar] [CrossRef]
- Wang, F.R.; Bai, M.Y.; Deng, Z.P.; Oses-Prieto, J.A.; Burlingame, A.L.; Lu, T.L.; Chong, K.; Wang, Z.Y. Proteomic Study Identifies Proteins Involved in Brassinosteroid Regulation of Rice Growth. J. Integr. Plant Biol. 2010, 12, 1075–1085. [Google Scholar] [CrossRef]
- Kim, K.M.; Jung, H.J.; Kim, D.H.; Kang, H. Characterization of glycine-rich RNA-binding proteins in Brassica napus under stress conditions. Physiol. Plant 2012, 146, 297–307. [Google Scholar] [CrossRef]
- Wang, B.; Wang, G.; Shen, F.; Zhu, S.J. A Glycine-Rich RNA-Binding Protein, CsGR-RBP3, Is Involved in Defense Responses Against Cold Stress in Harvested Cucumber (Cucumis sativus L.) Fruit. Front. Plant Sci. 2018, 9, 540. [Google Scholar] [CrossRef]
- Shinozuka, H.; Hisano, H.; Yoneyama, S.; Shimamoto, Y.; Jones, E.S.; Forster, J.W.; Yamada, T.; Kanazawa, A. Gene expression and genetic mapping analyses of a perennial ryegrass glycine-rich RNA-binding protein gene suggest a role in cold adaptation. Mol. Genet. Genet. 2006, 275, 399–408. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, D.W.; Wang, Y.C.; Zheng, L.; Yang, C.P. A glycine-rich RNA-binding protein can mediate physiological responses in transgenic plants under salt stress. Mol. Biol. Rep. 2012, 39, 1047–1053. [Google Scholar] [CrossRef]
- Aneeta; Sanan-Mishra, N.; Tuteja, N.; Sopory, S.K. Salinity- and ABA-induced up-regulation and light-mediated modulation of mRNA encoding glycine-rich RNA-binding protein from Sorghum bicolor. Biochem. Biophys. Res. Commun. 2002, 296, 1063–1068. [Google Scholar] [CrossRef]
- Teng, K.; Tan, P.; Xiao, G.; Han, L.; Chang, Z.; Chao, Y. Heterologous expression of a novel Zoysia japonica salt-induced glycine-rich RNA-binding protein gene, ZjGRP, caused salt sensitivity in Arabidopsis. Plant Cell Rep. 2016, 36, 179–191. [Google Scholar] [CrossRef]
- Wang, S.C.; Wang, R.C.; Liang, D.; Ma, F.W.; Shu, H.R. Molecular characterization and expression analysis of a glycine-rich rna-binding protein gene from malus hupehensis rehd. Mol. Biol. Rep. 2012, 39, 4145–4153. [Google Scholar] [CrossRef]
- Xiao, J.; Xu, S.J.; Li, C.H.; Xu, Y.Y.; Chong, K. O-GlcNAc-mediated interaction between VER2 and TaGRP2 elicits TaVRN1 mRNA accumulation during vernalization in winter wheat. Nat. Commun. 2014, 5, 4572. [Google Scholar] [CrossRef] [PubMed]
- Alptekin, B.; Mangel, D.; Pauli, D.; Blake, T.; Lachowiec, J.; Hoogland, T.; Fischer, A.; Sherman, J. Combined effects of a glycine-rich RNA-binding protein and a NAC transcription factor extend grain fill duration and improve malt barley agronomic performance. Theor. Appl. Genet. 2021, 134, 351–366. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Kim, N.H.; Hwang, B.K. Glycine-rich RNA-binding protein1 interacts with receptor-like cytoplasmic protein kinase1 and suppresses cell death and defense responses in pepper (Capsicum annuum). New Phytol. 2015, 205, 786–800. [Google Scholar] [CrossRef]
- Tian, L.; Chou, H.L.; Zhang, L.; Hwang, S.K.; Starkenburg, S.R.; Doroshenk, K.A.; Kumamaru, T.; Okita, T.W. RNA-binding protein RBP-P is required for glutelin and prolamine mRNA localization in rice endosperm cells. Plant Cell 2018, 30, 2529–2552. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Chou, H.L.; Zhang, L.; Okita, T.W. Targeted endoplasmic reticulum localization of storage protein mRNAs requires the RNA-binding protein RBP-L. Plant Physiol. 2019, 179, 1111–1131. [Google Scholar] [CrossRef] [PubMed]
- Staszak, A.; Pawlowski, T. Proteomic Analysis of Embryogenesis and the Acquisition of Seed Dormancy in Norway Maple (Acer platanoides L.). Int. J. Mol. Sci. 2014, 15, 10868–10891. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Germain, A.; Giloteaux, L.; Hammani, K.; Barkan, A.; Hanson, M.R.; Bentolila, S. An RNA recognition motif-containing protein is required for plastid RNA editing in Arabidopsis and maize. Proc. Natl. Acad. Sci. USA 2013, 110, E1169–E1178. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Virdi, A.S.; Jaswal, R.; Chawla, M.; Kapoor, S.; Mohapatra, S.B.; Manoj, N.; Pareek, A.; Kumar, S.; Singh, P. A temperature-responsive gene in sorghum encodes a glycine-rich protein that interacts with calmodulin. Biochimie 2017, 137, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Boyer, J.S. Plant productivity and environment. Science 1982, 218, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chan, Z.L.; Gao, J.H.; Xing, L.; Cao, M.J.; Yu, C.M.; Hu, Y.L.; You, J.; Shi, H.T.; Zhu, Y.F.; et al. BA receptor PYL9 promotes drought resistance and leaf senescence. Proc. Natl. Acad. Sci. USA 2016, 113, 1949–1954. [Google Scholar] [CrossRef]
- Zargar, S.M.; Nagar, P.; Deshmukh, R.; Nazir, M.; Wani, A.A.; Masoodi, K.Z.; Agrawal, G.K.; Rakwal, R. Aquaporins as potential drought tolerance inducing proteins: Towards instigating stress tolerance. J. Proteom. 2017, 169, 233–238. [Google Scholar] [CrossRef]
- Zhao, L.; Feng, C.; Wu, K.; Chen, W.; Wu, Y. Advances and prospects in biogenic substances against plant virus: A review. Pestic. Biochem. Physiol. 2015, 135, 15–26. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, X.; Zhou, G.; Zhang, T. Engineering plant virus resistance: From RNA silencing to genome editing strategies. Plant Biotechnol. J. 2020, 18, 328–336. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, J.S.; Yoo, S.J.; Kang, E.Y.; Han, S.H. Different roles of glycine-rich RNA-binding protein7 in plant defense against Pectobacterium carotovorum, Botrytis cinerea, and tobacco mosaic viruses. Plant Physiol. Biochem. 2012, 60, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Yu, R.W.; Li, R.; Geng, L.; Jing, X.; Zhu, C.; Liu, H. Identification and characterisation of a glycine-rich RNA-binding protein as an endogenous suppressor of RNA silencing from Nicotiana glutinosa. Planta 2019, 249, 1811–1822. [Google Scholar] [CrossRef] [PubMed]
- Kelley, R.Y.; Williams, W.P.; Mylroie, J.E.; Boykin, D.L.; Harper, J.W.; Windham, G.L.; Ankala, A.; Shan, X.Y. Identification of Maize Genes Associated with Host Plant Resistance or Susceptibility to Aspergillus flavus Infection and Aflatoxin Accumulation. PLoS ONE 2012, 7, e36892. [Google Scholar] [CrossRef] [PubMed]
- Gagliardi, M.; Matarazzo, M.R. RIP: RNA Immunoprecipitation. Methods Mol. Boil. 2016, 1480, 73–86. [Google Scholar]
- Zhao, J.C. nRIP-seq: A technique to identify RNA targets of an RNA binding protein on a genome-wide scale. Methods Mol. Med. 2015, 1206, 97–106. [Google Scholar]
- Mcdermott, J.J.; Watkins, K.P.; Williams-Carrier, R.; Barkan, A. Ribonucleoprotein capture by in vivo expression of a designer pentatricopeptide repeat protein in Arabidopsis. Plant Cell 2019, 31, 1723–1733. [Google Scholar] [CrossRef]
- Streitner, C.; Köster, T.; Simpson, C.G.; Shaw, P.; Danisman, S.; Brown, J.W.S.; Staiger, D. An hnRNP-like RNA-binding protein affects alternative splicing by in vivo interaction with transcripts in Arabidopsis thaliana. Nucleic Acids Res. 2012, 40, 11240–11255. [Google Scholar] [CrossRef]
- Qi, H.D.; Lin, Y.; Ren, Q.P.; Wang, Y.Y.; Xiong, F.; Wang, X.L. RNA Splicing of FLC Modulates the Transition to Flowering. Front. Plant Sci. 2019, 10, 1625. [Google Scholar] [CrossRef]
- Kim, J.S.; Jung, H.J.; Lee, H.J.; Kim, K.A.; Goh, C.H.; Woo, Y.; Oh, S.H.; Han, Y.S.; Kang, H. Glycine-rich RNA-binding protein7 affects abiotic stress responses by regulating stomata opening and closing in Arabidopsis thaliana. Plant J. 2008, 55, 455–466. [Google Scholar] [CrossRef]
- He, P.; Xiao, G.H.; Liu, H.; Zhang, L.H.; Zhao, L.; Tang, M.J.; Huang, S.; An, Y.J.; Yu, J.N. Two pivotal RNA editing sites in the mitochondrial atp1mRNA are required for ATP synthase to produce sufficient ATP for cotton fiber cell elongation. New Phytol. 2018, 218, 167–182. [Google Scholar] [CrossRef]
- Czolpinska, M.; Rurek, M. Plant Glycine-Rich Proteins in Stress Response: An Emerging, Still Prospective Story. Front. Plant Sci. 2018, 9, 302. [Google Scholar] [CrossRef]
- Dresios, J.; Aschrafi, A.; Owens, G.C.; Vanderklish, P.W.; Edelman, G.M.; Mauro, V.P. Cold stress-induced protein Rbm3 binds 60S ribosomal subunits, alters microRNA levels, and enhances global protein synthesis. Proc. Natl. Acad. Sci. USA 2005, 102, 1865–1870. [Google Scholar] [CrossRef]
- Lu, C.K.; Lai, Y.C.; Chen, H.R.; Chiang, M.K. Rbms3, an RNA-Binding Protein, Mediates the Expression of Ptf1a by Binding to Its 3′UTR during Mouse Pancreas Development. DNA Cell Biol. 2012, 31, 1245–1251. [Google Scholar] [CrossRef]
- Hennig, S.; Kong, G.; Mannen, T.; Sadowska, A.; Kobelke, S.; Blythe, A.; Knott, G.J.; Lyer, K.S.; Ho, D.W.; Newcombe, E.A.; et al. Prion-like domains in RNA binding proteins are essential for building subnuclear paraspeckles. J. Cell Biol. 2015, 210, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Maharana, S.; Wang, J.; Papadopoulos, D.K.; Richter, D.; Pozniakovsky, A.; Poser, I.; Poser, I.; Bickle, M.; Rizk, S.; Guillen-Boixét, J.; et al. RNA buffers the phase separation behavior of prion-like RNA binding proteins. Science 2018, 370, 918–921. [Google Scholar] [CrossRef] [PubMed]
- Banani, S.F.; Lee, H.O.; Hyman, A.A.; Rosen, M.K. Biomolecular condensates: Organizers of cellular biochemistry. Nat. Rev. Mol. Cell Biol. 2017, 18, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Emenecker, R.J.; Holehouse, A.S.; Strader, L.C. Emerging roles for phase separation in plants. Dev. Cell 2020, 55, 69–83. [Google Scholar] [CrossRef]
- Xie, D.Q.; Chen, M.; Niu, J.R.; Wang, L.; Li, Y.; Fang, X.F.; Li, P.L.; Qi, Y. Phase separation of SERRATE drives dicing body assembly and promotes miRNA processing in Arabidopsis. Nat. Cell Biol. 2020, 23, 32–39. [Google Scholar] [CrossRef]
- Huang, X.Z.; Chen, S.D.; Li, W.P.; Tang, L.L.; Xu, C. ROS regulated reversible protein phase separation synchronizes plant flowering. Nat. Chem. Biol. 2021, 17, 549–557. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Plant | Subclass I | Subclass II | Subclass III | Subclass IV | Total |
|---|---|---|---|---|---|
| Arabidopsis thaliana | 8 | 3 | 2 | 5 | 18 |
| Oryza sativa | 6 | 3 | 2 | 4 | 15 |
| Zea mays | 6 | 6 | 2 | 9 | 23 |
| Theobroma cacao | 6 | 0 | 0 | 9 | 15 |
| Brassica rapan L. ss P. pekinensis | 15 | 4 | 5 | 6 | 30 |
| Gossypium raimondii | 11 | 6 | 6 | 9 | 32 |
| Gossypium arboreum | 14 | 4 | 7 | 12 | 37 |
| Gene | Subclass | Gene Source | Growth Phenotype | Roles | References |
|---|---|---|---|---|---|
| AtGRP2 ORRM5 | I | Arabidopsis thaliana | Under cold stress accelerate seed germination and seedling growth, under high salt and dehydration stress conditions affect plants growth and stress tolerance, slow growth and late flowering | Overexpressing enhances freezing tolerance of Arabidopsis plants, AtGRP2 in inhibition of the early stages of ZYMV infection, mitochondrial RNA editing | [14,25,26,27,28,29] |
| AtGRP4 | I | Arabidopsis thaliana | Overexpressing seeds delayed germination during high salt or dehydration stress | The transcripts increase under cold stress, and downregulated by high salinity and dehydration stress | [18,19] |
| AtGRP7 | I | Arabidopsis thaliana | Affect the growth and stress tolerance of A. thaliana plants, influence flowering time, overexpressing in rice shows higher recovery rates and grain yields | The transcripts increase significantly under cold stress, influence AS or polyadenylation, influence mRNA export from the nucleus to the cytoplasm under cold stress conditions, | [18,27,28,30,31,32,33,34,35] |
| involvement in plant defenses, such as Pseudomonas syringae and TMV | |||||
| ORRM4 GR-RBP5 | I | Arabidopsis thaliana | Slow growth and late flowering | Mitochondrial RNA editing | [19,36] |
| SlORRM4 | I | Solanum lycopersicum | Delayed tomato fruit ripening | Mitochondrial RNA editing | [37,38] |
| AtRZ-1a | III | Arabidopsis thaliana | Enhances tolerance to cold stress in A. thaliana | Overexpressing in salt and cold stress retards seed germination | [21,22] |
| AtRZ-1b | III | Arabidopsis thaliana | Enhances tolerance to cold stress, AtRZ-1b and AtRZ-1c knockout mutants delayed seed germination, reduced stature and serrated leaves | Promote efficient splicing of FLC introns and repress FLC transcription | [22,39] |
| AtRZ-1c | III | Arabidopsis thaliana | AtRZ-1b and AtRZ-1c knockout mutants delayed seed germination, reduced stature and serrated leaves | Promote efficient splicing of FLC introns and repress FLC transcription | [22,39] |
| TaRZ-2 | III | Triticum aestivum | Overexpressing retards seed germination under dehydration stress condition | Not determined | [23] |
| TaRZ-3 | III | Triticum aestivum | Overexpressing retards seed germination under dehydration stress condition | Not determined | [23] |
| OsGRP1 | I | Oryza sativa | Overexpressing suppresses the dwarf phenotype of Arabidopsis bri1-5 mutant, under low temperatures promotes seed germination and seedling growth | Promotes cell expansion and elongation, enhances freezing tolerance | [5,40] |
| OsGRP4 | I | Oryza sativa | Under low temperatures promotes seed germination and seedling growth | Enhances tolerance to cold stress | [5] |
| OsGRP6 | I | Oryza sativa | Not determined | Enhances tolerance to cold stress | [5] |
| OsRZ-2 | III | Oryza sativa | Under low temperatures rescues grp7- knockout plants | Not determined | [13] |
| NtGRP-1a | I | Nicotiana tabacum | Not determined | Upregulation of abundance during heat and drought stress | [4] |
| NtGRP-3 | I | Nicotiana tabacum | Not determined | Upregulation of abundance during heat and drought stress | [4] |
| BnGRP1 | I | Brassica napus | Under cold stress accelerates seed germination | Enhances tolerance to cold stress | [41] |
| CsGRP2 | I | Camelina sativa | Complement cold-sensitive mutants at low temperatures | Upregulation of cold stress | [8] |
| CsGRP3 | I | Cucumis sativa | Overexpressing contributes to cold and freezing stress tolerance | Not determined | [42] |
| LpGRP1 | I | Lolium perenne | Not determined | Upregulation of cold stress | [43] |
| ItGRP1 ItGRP5 ItGRP7 | I | Ipomoea trifida | Not determined | Upregulation of heat stress | [7] |
| LbGRP1 | I | Limonium bicolor | Not determined | Improves tolerance to salt stress | [44] |
| SbGR-RNP | I | Sorghum bicolor | Not determined | Upregulation of salt stress | [45] |
| ZjGRP | I | Zoysia japonica | Overexpressing increases salt sensitivity in A. thaliana | Not determined | [46] |
| MhGR-RBP1 | I | Malus hupehensis | Not determined | Upregulation of salt stress | [47] |
| CsSRBP1 | I | Cucumis sativus | Not determined | Inhibits the initial stages of Zucchini yellow mosaic virus (ZYMV) infection | [29] |
| TaGRP2 | I | Triticum aestivum | Not determined | Inhibits transcript accumulation of TaVRN1 | [48] |
| HvGR-RBP1 | I | Hordeum vulgare | Not determined | Involvement in the timing of anthesis, senescence and levels of grain protein | [49] |
| CaGRP1 | I | Capsicum annuum | Not determined | Resistance to Xanthomonas campestris pv vesicatoria (Xcv) infection | [50] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, L.; Cheng, K.; Li, J.; Deng, Z.; Zhang, C.; Zhu, H. Roles of Plant Glycine-Rich RNA-Binding Proteins in Development and Stress Responses. Int. J. Mol. Sci. 2021, 22, 5849. https://doi.org/10.3390/ijms22115849
Ma L, Cheng K, Li J, Deng Z, Zhang C, Zhu H. Roles of Plant Glycine-Rich RNA-Binding Proteins in Development and Stress Responses. International Journal of Molecular Sciences. 2021; 22(11):5849. https://doi.org/10.3390/ijms22115849
Chicago/Turabian StyleMa, Liqun, Ke Cheng, Jinyan Li, Zhiqi Deng, Chunjiao Zhang, and Hongliang Zhu. 2021. "Roles of Plant Glycine-Rich RNA-Binding Proteins in Development and Stress Responses" International Journal of Molecular Sciences 22, no. 11: 5849. https://doi.org/10.3390/ijms22115849
APA StyleMa, L., Cheng, K., Li, J., Deng, Z., Zhang, C., & Zhu, H. (2021). Roles of Plant Glycine-Rich RNA-Binding Proteins in Development and Stress Responses. International Journal of Molecular Sciences, 22(11), 5849. https://doi.org/10.3390/ijms22115849