
Overexpression of the Apple (Malus × domestica) MdERF100 in Arabidopsis Increases Resistance to Powdery Mildew

 , ,
, ,  and
and 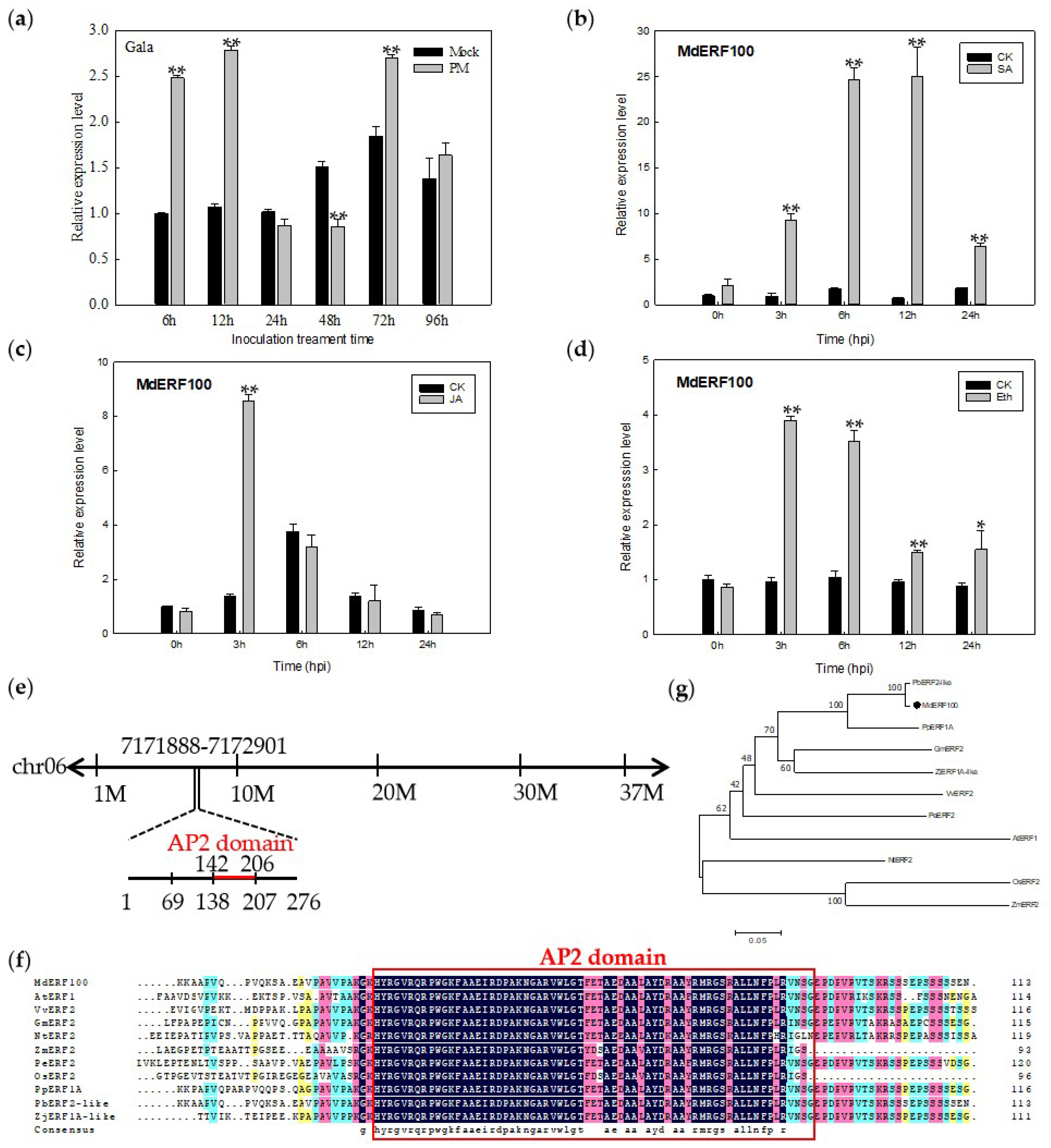{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Expression Pattern and Characterization of MdERF100
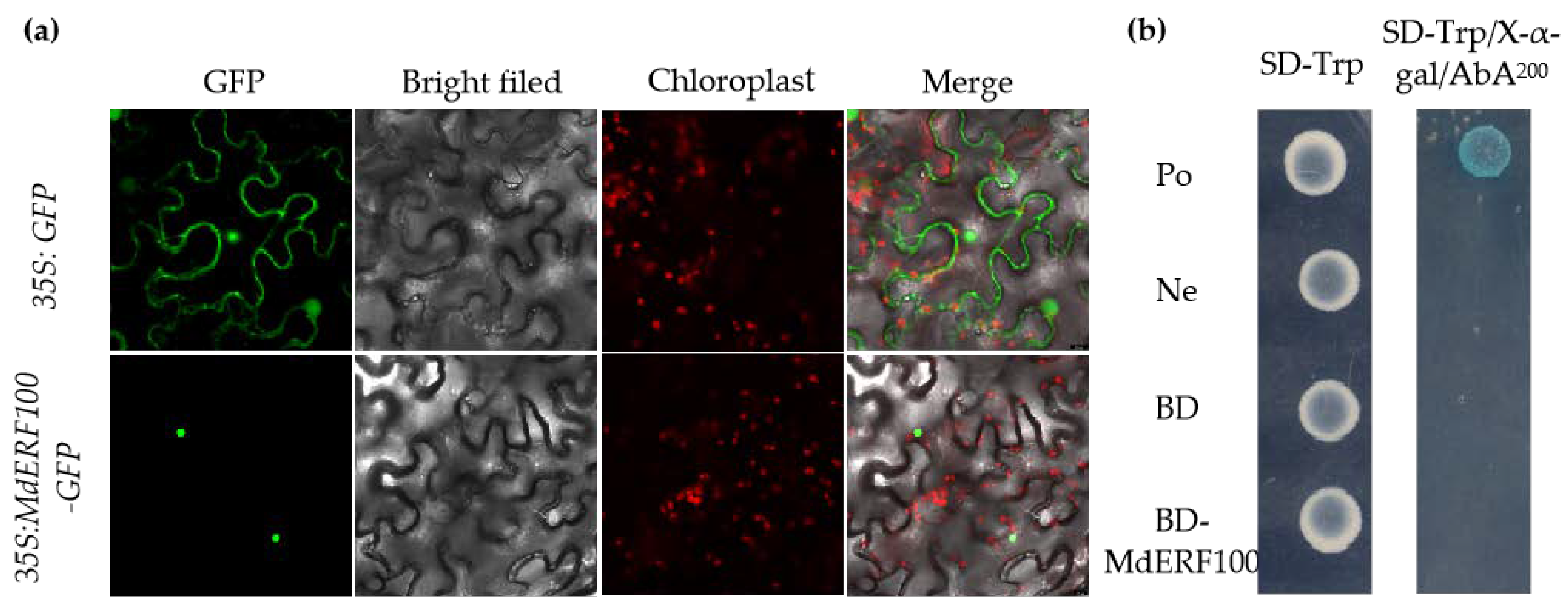
2.2. MdERF100 Shows Nuclear Localization but No Transcriptional Activation Activity
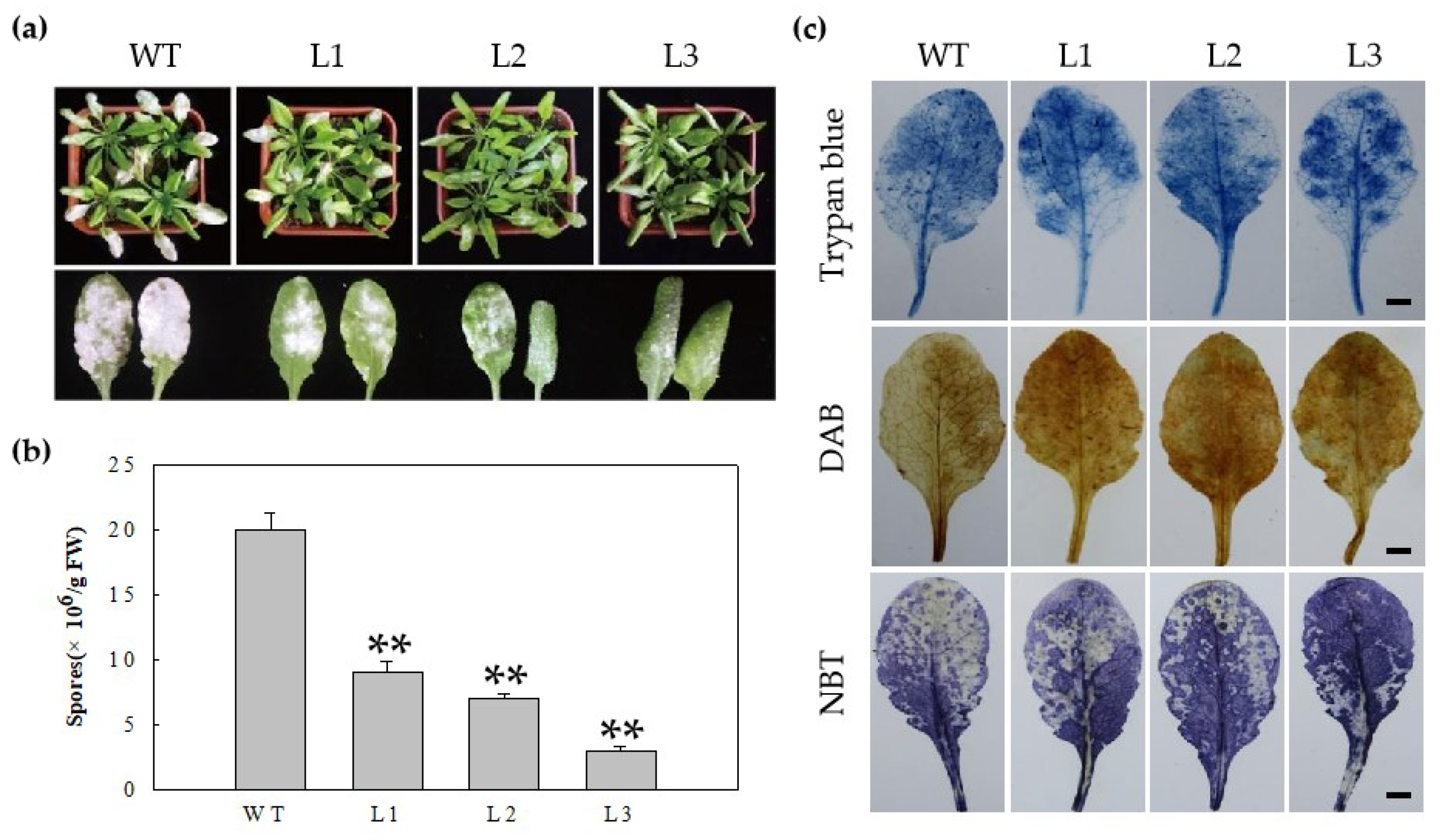
2.3. Heterologous Expression of MdERF100 in Arabidopsis Confers Powdery Mildew Resistance
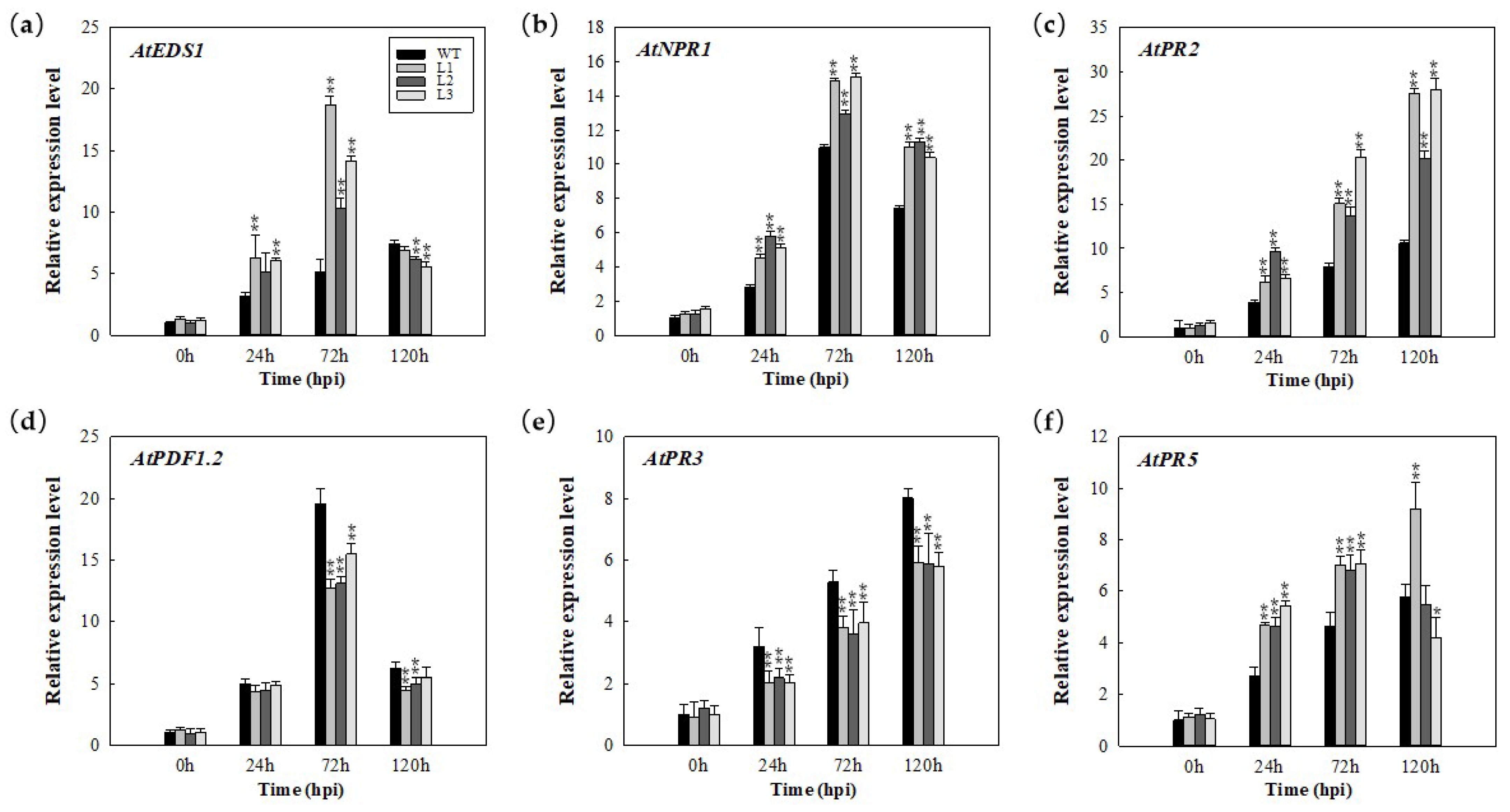
2.4. Expression Analysis of Disease Resistance (R) Genes in MdERF100-Overexpressing Plants in Response to E. necator Inoculation
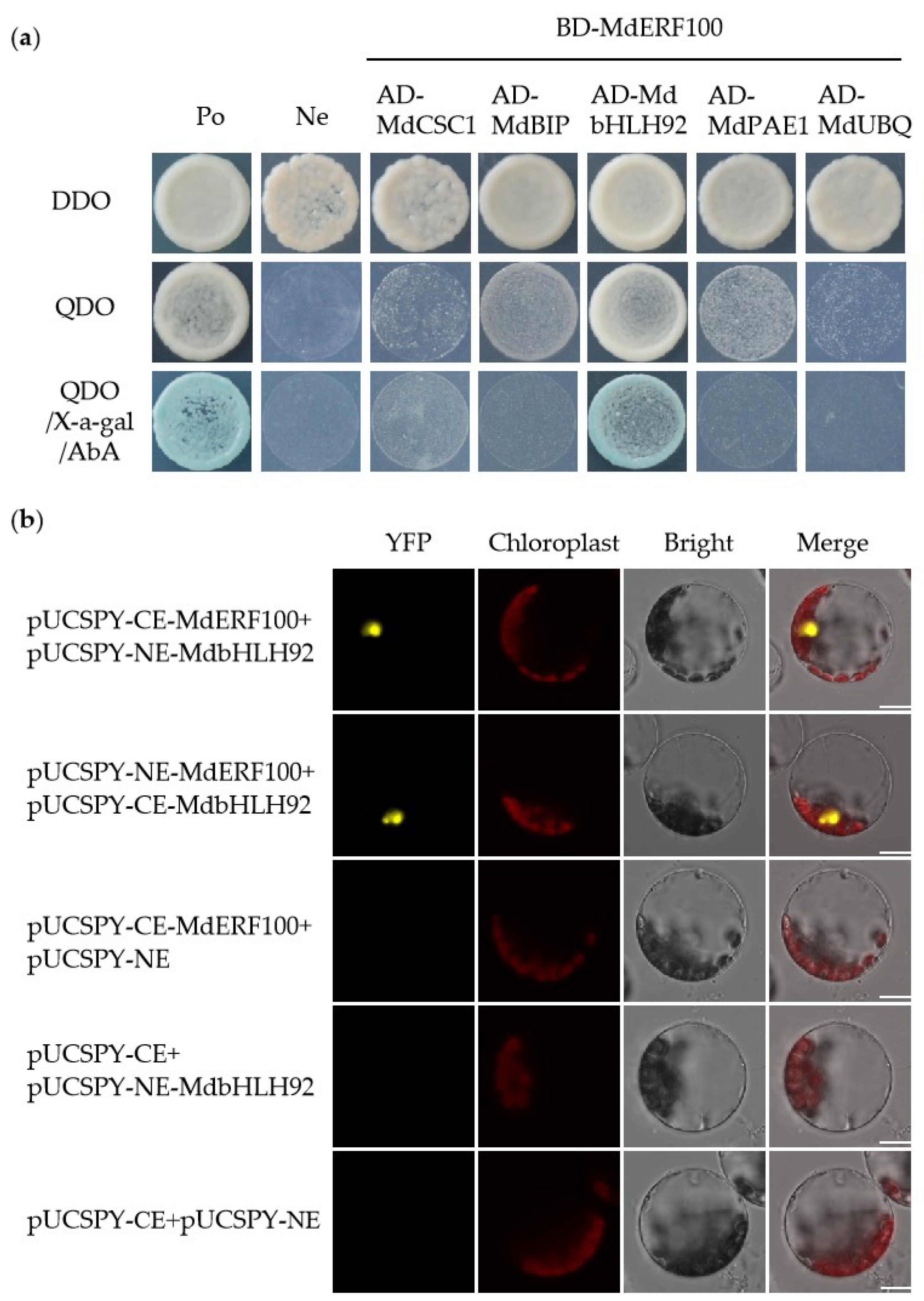
2.5. MdERF100 Physically Interacts with MdbHLH92
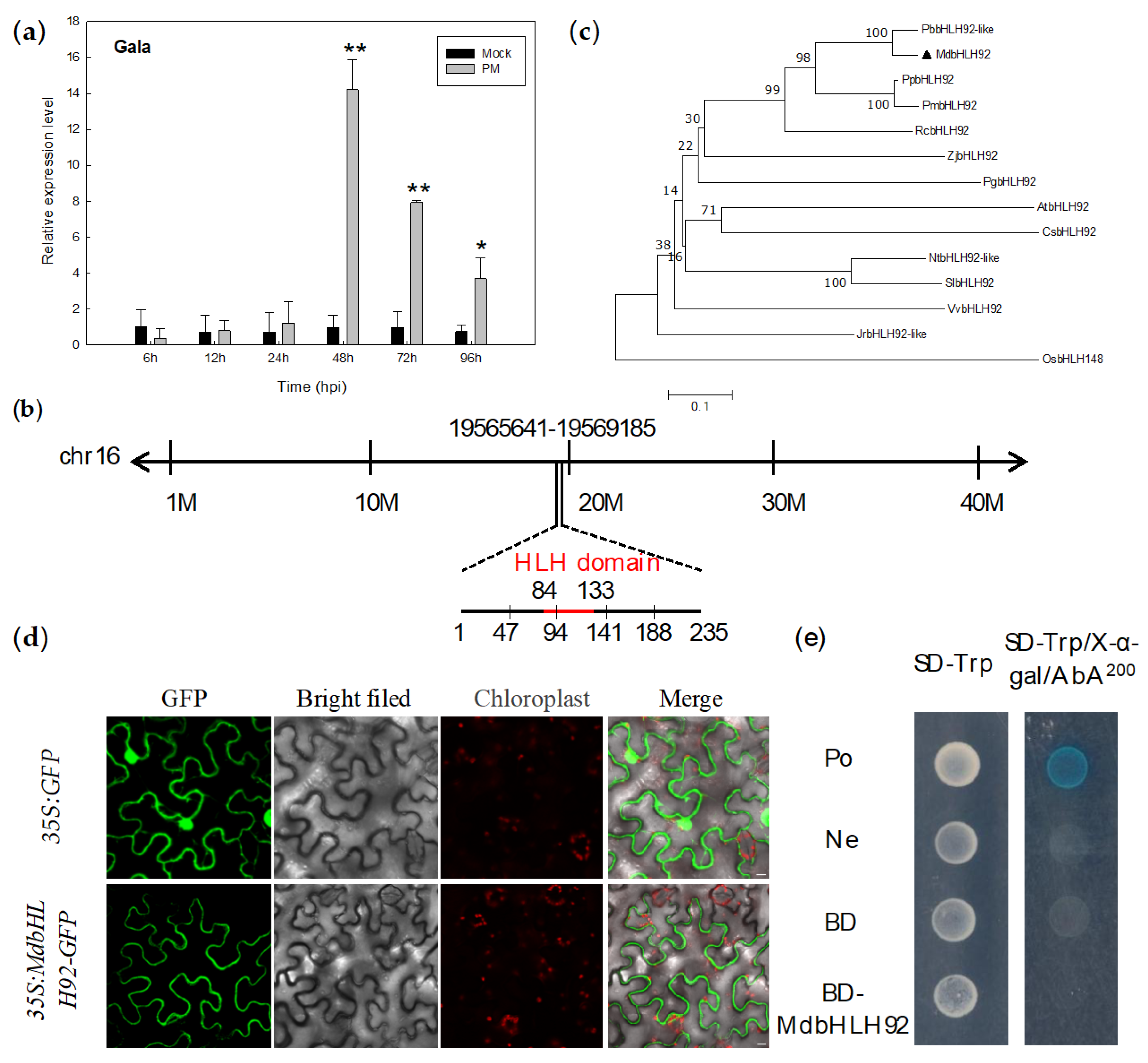
2.6. Isolation of MdbHLH92 and Its Response to Powdery Mildew
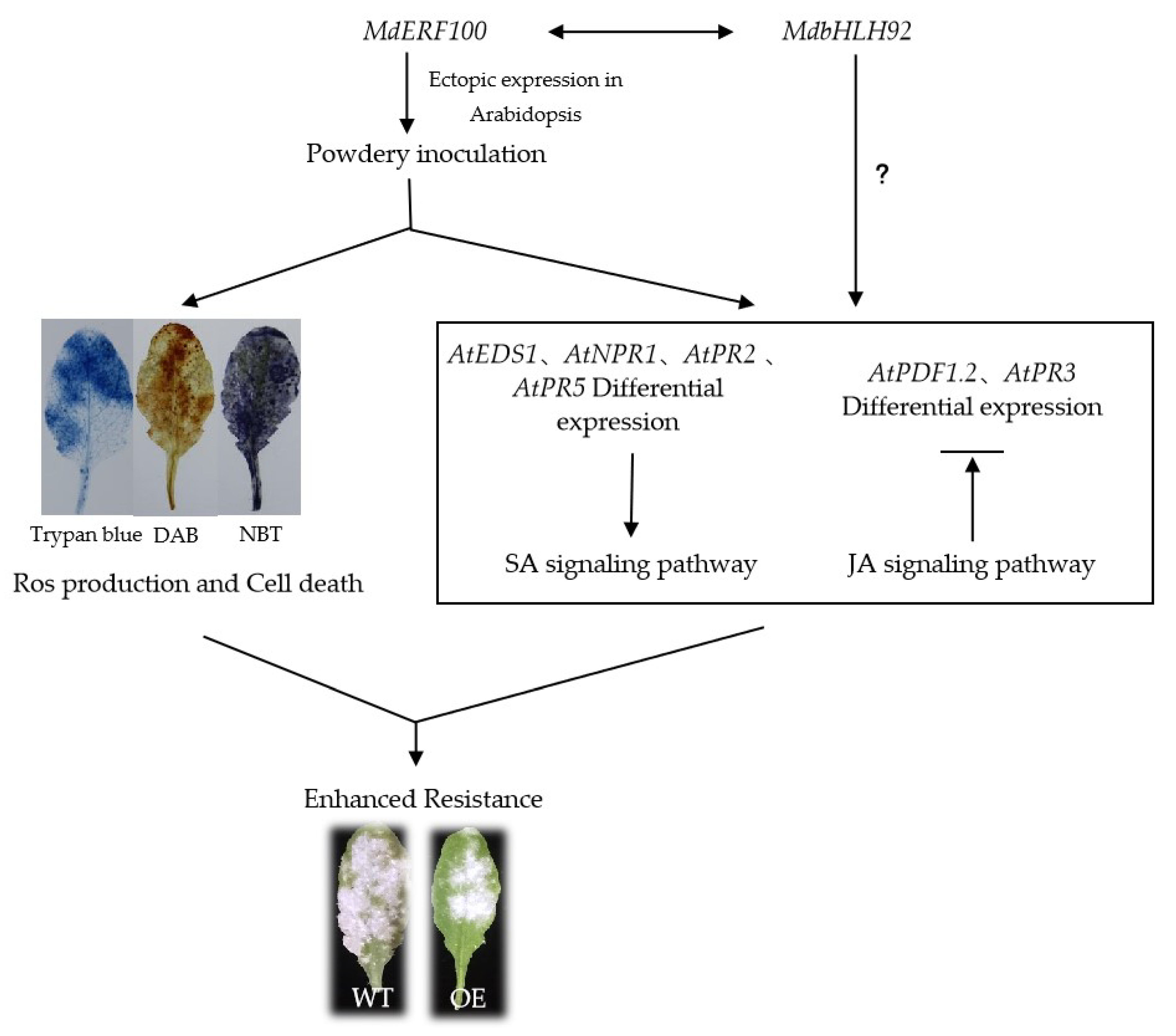
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Bioinformatics Analysis
4.3. Subcellular Localization and Transcriptional Activation Assays
4.4. Inoculation of Arabidopsis Plants with Powdery Mildew Pathogen
4.5. RNA Isolation and Gene Expression Analysis
4.6. Y2H and BiFC Assays
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tian, X.; Zhang, L.; Feng, S.; Zhao, Z.; Wang, X.; Gao, H. Transcriptome Analysis of Apple Leaves in Response to Powdery Mildew (Podosphaera leucotricha) Infection. Int. J. Mol. Sci. 2019, 20, 2326. [Google Scholar] [CrossRef]
- Gañán, L.; White, I.R.; Friesen, M.; Peever, T.; Amiri, A. A Genome Resource for the Apple Powdery Mildew Pathogen Podo-sphaera leucotricha. Phytopathology 2020, 110, 1756–1758. [Google Scholar] [CrossRef]
- Turechek, W.W.; Carroll, J.E.; Rosenberger, D.A. Powdery Mildew of Apple. 2005. Available online: https://ecommons.cornell.edu/handle/1813/43120 (accessed on 10 May 2019).
- Pessina, S.; Angeli, D.; Martens, S.; Visser, R.G.; Bai, Y.; Salamini, F.; Velasco, R.; Schouten, H.J.; Malnoy, M. The knock-down of the expression of MdMLO19 reduces susceptibility to powdery mildew (Podosphaera leucotricha) in apple (Malus domestica). Plant Biotechnol. J. 2016, 14, 2033–2044. [Google Scholar] [CrossRef] [PubMed]
- Li, M.-Y.; Xu, Z.-S.; Huang, Y.; Tian, C.; Wang, F.; Xiong, A.-S. Genome-wide analysis of AP2/ERF transcription factors in carrot (Daucus carota L.) reveals evolution and expression profiles under abiotic stress. Mol. Genet. Genom. 2015, 290, 2049–2061. [Google Scholar] [CrossRef]
- Wang, L. Studies on transcription factor ERF regulating disease resistance-related genes in Chinese wild Vitis quinquangularis. Ph.D. Thesis, Northwest A&F University, Xianyang, China, 2019. [Google Scholar]
- Gutterson, N.; Reuber, T.L. Regulation of disease resistance pathways by AP2/ERF transcription factors. Curr. Opin. Plant Biol. 2004, 7, 465–471. [Google Scholar] [CrossRef]
- Liu, M.; Ma, F.; Wu, F.; Jiang, C.; Wang, Y. Expression of stilbene synthase VqSTS6 from wild Chinese Vitis quinquangularis in grapevine enhances resveratrol production and powdery mildew resistance. Planta 2019, 250, 1997–2007. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, J.; Zhang, S.; Yu, Q.; Lan, H. Investigation of the Nature of CgCDPK and CgbHLH001 Interaction and the Function of bHLH Transcription Factor in Stress Tolerance in Chenopodium glaucum. Front. Plant Sci. 2021, 11, 603298. [Google Scholar] [CrossRef]
- Toledo-Ortiz, G.; Huq, E.; Quail, P.H. The Arabidopsis Basic/Helix-Loop-Helix Transcription Factor Family. Plant Cell 2003, 15, 1749–1770. [Google Scholar] [CrossRef] [PubMed]
- Feller, A.; Machemer, K.; Braun, E.L.; Grotewold, E. Evolutionary and comparative analysis of MYB and bHLH plant transcription factors. Plant J. 2011, 66, 94–116. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Wen, X.; Wen, Z.; Qiu, Z.; Hou, Q.; Li, Z.; Mei, L.; Yu, H.; Qiao, G. Genome-wide identification and expression analysis of bHLH transcription factor family in response to cold stress in sweet cherry (Prunus avium L.). Sci. Hortic. 2021, 279, 109905. [Google Scholar] [CrossRef]
- Qiu, J.-R.; Huang, Z.; Xiang, X.-Y.; Xu, W.-X.; Wang, J.-T.; Chen, J.; Song, L.; Xiao, Y.; Li, X.; Ma, J.; et al. MfbHLH38, a Myrothamnus flabellifolia bHLH transcription factor, confers tolerance to drought and salinity stresses in Arabidopsis. BMC Plant Biol. 2020, 20, 1–14. [Google Scholar] [CrossRef]
- Rajappa, S.; Krishnamurthy, P.; Kumar, P.P. Regulation of AtKUP2 Expression by bHLH and WRKY Transcription Factors Helps to Confer Increased Salt Tolerance to Arabidopsis thaliana Plants. Front. Plant Sci. 2020, 11, 1311. [Google Scholar] [CrossRef]
- Lockhart, J. Personal Trainer: bHLH121 functions upstream of a transcriptional network of heavy lifters involved in bal-ancing iron levels. Plant Cell 2020, 32, 293–294. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-G.; Mudgett, M.B. Tomato bHLH132 Transcription Factor Controls Growth and Defense and Is Activated by Xanthomonas euvesicatoria Effector XopD During Pathogenesis. Mol. Plant-Microbe Interact. 2019, 32, 1614–1622. [Google Scholar] [CrossRef]
- Wang, M.; Yang, D.; Ma, F.; Zhu, M.; Shi, Z.; Miao, X. OsHLH61-OsbHLH96 influences rice defense to brown planthopper through regulating the pathogen-related genes. Rice 2019, 12, 9. [Google Scholar] [CrossRef]
- Ke, Y.; Kang, Y.; Wu, M.; Liu, H.; Hui, S.; Zhang, Q.; Li, X.; Xiao, J.; Wang, S. Jasmonic Acid-Involved OsEDS1 Signaling in Rice-Bacteria Interactions. Rice 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Matthews, B.F.; Beard, H.; Brewer, E.; Kabir, S.; MacDonald, M.H.; Youssef, R.M. Arabidopsis genes, AtNPR1, AtTGA2 and AtPR-5, confer partial resistance to soybean cyst nematode (Heterodera glycines) when overexpressed in transgenic soy-bean roots. BMC Plant Biol. 2014, 14, 96. [Google Scholar] [CrossRef] [PubMed]
- Yuan, P.; Jewell, J.B.; Behera, S.; Tanaka, K.; Poovaiah, B.W. Distinct Molecular Pattern-Induced Calcium Signatures Lead to Different Downstream Transcriptional Regulations via AtSR1/CAMTA3. Int. J. Mol. Sci. 2020, 21, 8163. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, D.; Chu, J.; Boyle, P.; Wang, Y.; Brindle, I.D.; De Luca, V.; Després, C. The Arabidopsis NPR1 protein is a re-ceptor for the plant defense hormone salicylic acid. Cell Rep. 2012, 1, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Mur, L.A.; Kenton, P.; Atzorn, R.; Miersch, O.; Wasternack, C. The Outcomes of Concentration-Specific Interactions between Salicylate and Jasmonate Signaling Include Synergy, Antagonism, and Oxidative Stress Leading to Cell Death. Plant Physiol. 2006, 140, 249–262. [Google Scholar] [CrossRef]
- Jayakannan, M.; Bose, J.; Babourina, O.; Rengel, Z.; Shabala, S. Salicylic acid in plant salinity stress signalling and tolerance. Plant Growth Regul. 2015, 76, 25–40. [Google Scholar] [CrossRef]
- Zhang, X.; Ménard, R.; Li, Y.; Coruzzi, G.M.; Heitz, T.; Shen, W.-H.; Berr, A. Arabidopsis SDG8 Potentiates the Sustainable Transcriptional Induction of the Pathogenesis-Related Genes PR1 and PR2 During Plant Defense Response. Front. Plant Sci. 2020, 11, 277. [Google Scholar] [CrossRef]
- Colavolpe, M.B.; Villarreal, N.M.; Langer, S.E.; Romero, F.M.; Martínez, G.A.; Saini, A.; Ruiz, O.A.; Marina, M. Burkholderia sp. Strain AU4i Promotes Arabidopsis Growth and Increases Plant Defence Response to Necrotrophic Pathogens. J. Plant Growth Regul. 2020, 1–11. [Google Scholar] [CrossRef]
- Yu, Y.; Bian, L.; Wan, Y.; Jiao, Z.; Yu, K.; Zhang, G.; Guo, D. Grape (Vitis vinifera) VvDOF3 functions as a transcription activator and enhances powdery mildew resistance. Plant Physiol. Bioch. 2019, 143, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Di, Z.; Yang, W.; Liu, J.; Li, M.; Wang, X.; Cui, C.; Wang, X.; Wang, X.; Zhang, R.; et al. Overexpression of ERF1-V from Haynaldia villosa Can Enhance the Resistance of Wheat to Powdery Mildew and Increase the Tolerance to Salt and Drought Stresses. Front. Plant Sci. 2017, 8, 1948. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-K.; Zhang, J.-Y.; Zhang, Z.; Du, X.-L.; Du, B.-B.; Qu, S.-C. Overexpressing MhNPR1 in transgenic Fuji apples enhances resistance to apple powdery mildew. Mol. Biol. Rep. 2012, 39, 8083–8089. [Google Scholar] [CrossRef]
- Xie, Z.; Nolan, T.; Jiang, H.; Tang, B.; Zhang, M.; Li, Z.; Yin, Y. The AP2/ERF transcription factor TINY modulates brassino-steroid-regulated plant growth and drought responses in Arabidopsis. Plant Cell 2019, 31, 1788–1806. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, J.; Quan, R.; Pan, X.; Wan, L.; Huang, R. EAR motif mutation of rice OsERF3 alters the regulation of eth-ylene biosynthesis and drought tolerance. Planta 2013, 237, 1443–1451. [Google Scholar] [CrossRef]
- An, J.; Zhang, X.; Bi, S.; You, C.; Wang, X.; Hao, Y. The ERF transcription factor MdERF38 promotes drought stress-induced anthocyanin biosynthesis in apple. Plant J. 2019, 101, 573–589. [Google Scholar] [CrossRef]
- Cheng, L.; Zhao, T.; Wu, Y.; Wang, H.; Zhang, Z.; Zhang, D.; Wang, S.; Wang, Y. Identification of AP2/ERF genes in apple (Malus × domestica) and demonstration that MdERF017 enhances iron deficiency tolerance. Plant Cell Tiss. Org. 2020, 143, 465–482. [Google Scholar] [CrossRef]
- Zhao, M.-J.; Yin, L.-J.; Liu, Y.; Ma, J.; Zheng, J.-C.; Lan, J.-H.; Fu, J.-D.; Chen, M.; Xu, Z.-S.; Ma, Y.-Z. The ABA-induced soybean ERF transcription factor gene GmERF75 plays a role in enhancing osmotic stress tolerance in Arabidopsis and soybean. BMC Plant Biol. 2019, 19, 1–14. [Google Scholar] [CrossRef]
- Pan, Q.; Wang, C.; Xiong, Z.; Wang, H.; Fu, X.; Shen, Q.; Peng, B.; Ma, Y.; Sun, X.; Tang, K. CrERF5, an AP2/ERF Transcription Factor, Positively Regulates the Biosynthesis of Bisindole Alkaloids and Their Precursors in Catharanthus roseus. Front. Plant Sci. 2019, 10, 931. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-H.; Gu, K.-D.; Han, P.-L.; Yu, J.-Q.; Wang, C.-K.; Zhang, Q.-Y.; You, C.-X.; Hu, D.-G.; Hao, Y.-J. Apple ethylene response factor MdERF11 confers resistance to fungal pathogen Botryosphaeria dothidea. Plant Sci. 2020, 291, 110351. [Google Scholar] [CrossRef]
- Akagi, A.; Dandekar, A.M.; Stotz, H.U. Resistance of Malus domestica fruit to Botrytis cinerea depends on endogenous eth-ylene biosynthesis. Phytopathology 2011, 101, 1311–1321. [Google Scholar] [CrossRef]
- Zhu, Z.; Shi, J.; Xu, W.; Li, H.; He, M.; Xu, Y.; Xu, T.; Yang, Y.; Cao, J.; Wang, Y. Three ERF transcription factors from Chinese wild grapevine Vitis pseudoreticulata participate in different biotic and abiotic stress-responsive pathways. J. Plant Physiol. 2013, 170, 923–933. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, W.; Wang, Y. Heterologous expression of Chinese wild grapevine VqERFs in Arabidopsis thaliana enhance resistance to Pseudomonas syringae pv. tomato DC3000 and to Botrytis cinerea. Plant Sci. 2020, 293, 110421. [Google Scholar] [CrossRef] [PubMed]
- Schenk, P.M.; Kazan, K.; Wilson, I.; Anderson, J.P.; Richmond, T.; Somerville, S.C.; Manners, J.M. Coordinated plant defense responses in Arabidopsis revealed by microarray analysis. Proc. Natl. Acad. Sci. USA 2000, 97, 11655–11660. [Google Scholar] [CrossRef]
- Zhang, H.; Hong, Y.; Huang, L.; Li, D.; Song, F. Arabidopsis AtERF014 acts as a dual regulator that differentially modulates immunity against Pseudomonas syringae pv. tomato and Botrytis cinerea. Sci. Rep. 2016, 6, 30251. [Google Scholar] [CrossRef]
- Zhang, J.; Qiao, Y.; Lv, D.; Gao, Z.; Qu, S.; Zhang, Z. Malus hupehensis NPR1 induces pathogenesis-related protein gene expression in transgenic tobacco. Plant Biol. 2012, 14, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.-H.; Bian, L.; Yu, K.-K.; Yang, S.-D.; Zhang, G.-H.; Guo, D.-L. Grape (Vitis davidii) VdGATA2 functions as a transcription activator and enhances powdery mildew resistance via the active oxygen species pathway. Sci. Hortic. 2020, 267, 109327. [Google Scholar] [CrossRef]
- Robinson, K.A.; Koepke, J.I.; Kharodawala, M.; Lopes, J.M. A network of yeast basic helix-loop-helix interactions. Nucleic Acids Res. 2000, 28, 4460–4466. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Yang, B.; Deyholos, M.K. Functional characterization of the Arabidopsis bHLH92 transcription factor in abiotic stress. Mol. Genet. Genom. 2009, 282, 503–516. [Google Scholar] [CrossRef] [PubMed]
- Meraj, T.A.; Fu, J.; Raza, M.A.; Zhu, C.; Shen, Q.; Xu, D.; Wang, Q. Transcriptional Factors Regulate Plant Stress Responses Through Mediating Secondary Metabolism. Genes 2020, 11, 346. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Zhu, L.; Wassan, G.M.; Wang, Y.; Miao, Y.; Shaban, M.; Hu, H.; Sun, H.; Zhang, X. GhJAZ2 attenuates cotton resistance to biotic stresses via the inhibition of the transcriptional activity of GhbHLH171. Mol. Plant Pathol. 2017, 19, 896–908. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Qi, T.; Fan, M.; Zhang, X.; Gao, H.; Huang, H.; Wu, D.; Guo, H.; Xie, D. The bHLH Subgroup IIId Factors Negatively Regulate Jasmonate-Mediated Plant Defense and Development. PLoS Genet. 2013, 9, e1003653. [Google Scholar] [CrossRef] [PubMed]
- Lui, S.; Luo, C.; Zhu, L.; Sha, R.; Qu, S.; Cai, B.; Wang, S. Identification and expression analysis of WRKY transcription factor genes in response to fungal pathogen and hormone treatments in apple (Malus domestica). J. Plant Biol. 2017, 60, 215–230. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamur, A.K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, L.; Ji, M.; Wu, Y.; Zhang, S.; Zhu, Y.; Yao, J.; Li, Z.; Gao, H.; Wang, X. Genome-wide identification and ex-pression analysis of the B-box transcription factor gene family in grapevine. BMC Genom. 2021, 22, 221. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Walter, M.; Chaban, C.; Schütze, K.; Batistic, O.; Weckermann, K.; Näke, C.; Blazevic, D.; Grefen, C.; Schumacher, K.; Oecking, C.; et al. Visualization of protein interactions in living plant cells using bimolecular fluorescence complementa-tion. Plant J. 2004, 40, 428–438. [Google Scholar] [CrossRef]
- Yoo, S.-D.; Cho, Y.-H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhang, L.; Ma, H.; Zhang, Y.; Zhang, X.; Ji, M.; van Nocker, S.; Ahmad, B.; Zhao, Z.; Wang, X.; et al. Overexpression of the Apple (Malus × domestica) MdERF100 in Arabidopsis Increases Resistance to Powdery Mildew. Int. J. Mol. Sci. 2021, 22, 5713. https://doi.org/10.3390/ijms22115713
Zhang Y, Zhang L, Ma H, Zhang Y, Zhang X, Ji M, van Nocker S, Ahmad B, Zhao Z, Wang X, et al. Overexpression of the Apple (Malus × domestica) MdERF100 in Arabidopsis Increases Resistance to Powdery Mildew. International Journal of Molecular Sciences. 2021; 22(11):5713. https://doi.org/10.3390/ijms22115713
Chicago/Turabian StyleZhang, Yiping, Li Zhang, Hai Ma, Yichu Zhang, Xiuming Zhang, Miaomiao Ji, Steve van Nocker, Bilal Ahmad, Zhengyang Zhao, Xiping Wang, and et al. 2021. "Overexpression of the Apple (Malus × domestica) MdERF100 in Arabidopsis Increases Resistance to Powdery Mildew" International Journal of Molecular Sciences 22, no. 11: 5713. https://doi.org/10.3390/ijms22115713
APA StyleZhang, Y., Zhang, L., Ma, H., Zhang, Y., Zhang, X., Ji, M., van Nocker, S., Ahmad, B., Zhao, Z., Wang, X., & Gao, H. (2021). Overexpression of the Apple (Malus × domestica) MdERF100 in Arabidopsis Increases Resistance to Powdery Mildew. International Journal of Molecular Sciences, 22(11), 5713. https://doi.org/10.3390/ijms22115713