
Optogenetic Approaches for the Spatiotemporal Control of Signal Transduction Pathways

Abstract
1. Introduction
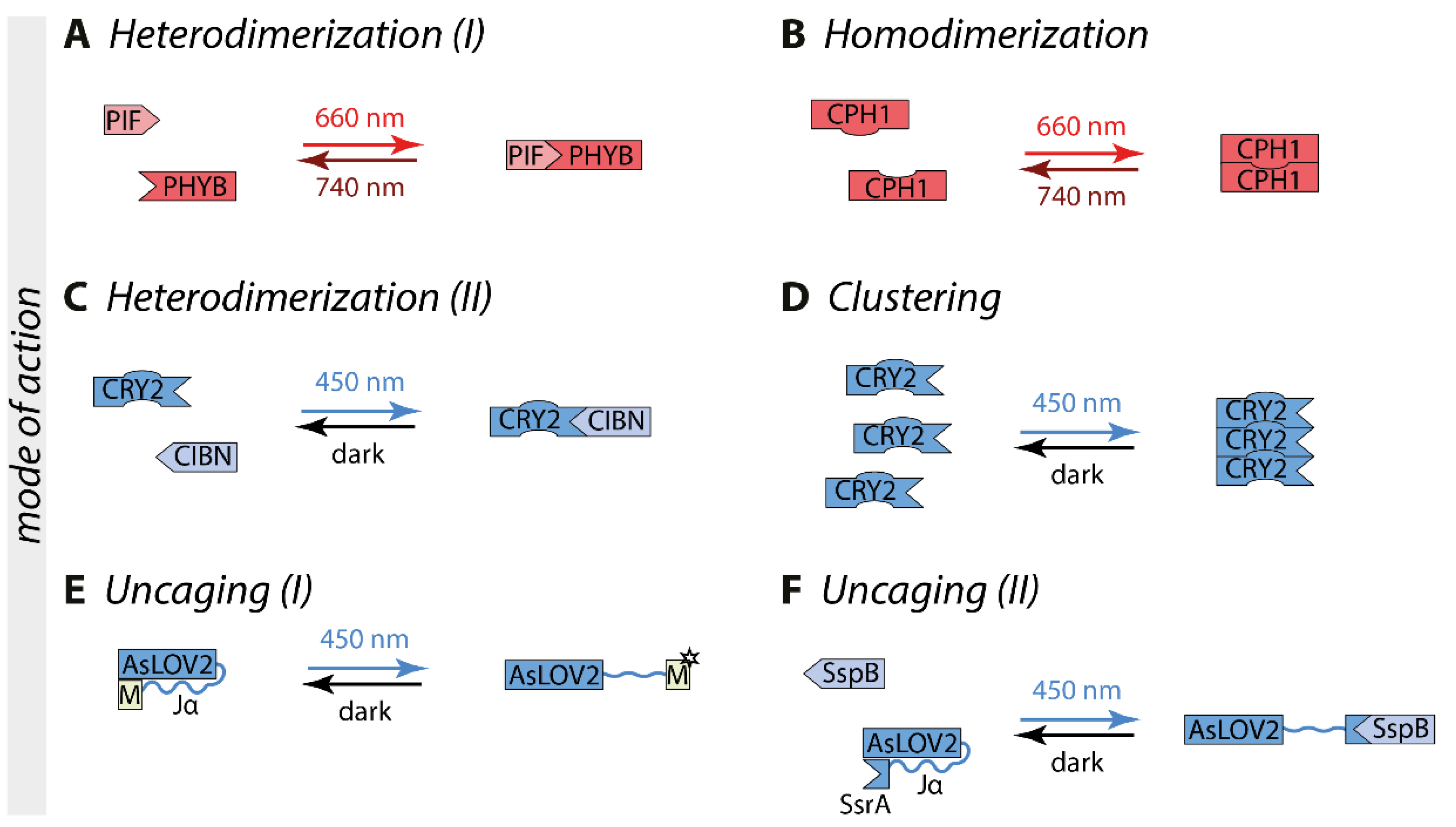
2. Optogenetic Switches as Basis for Light Control
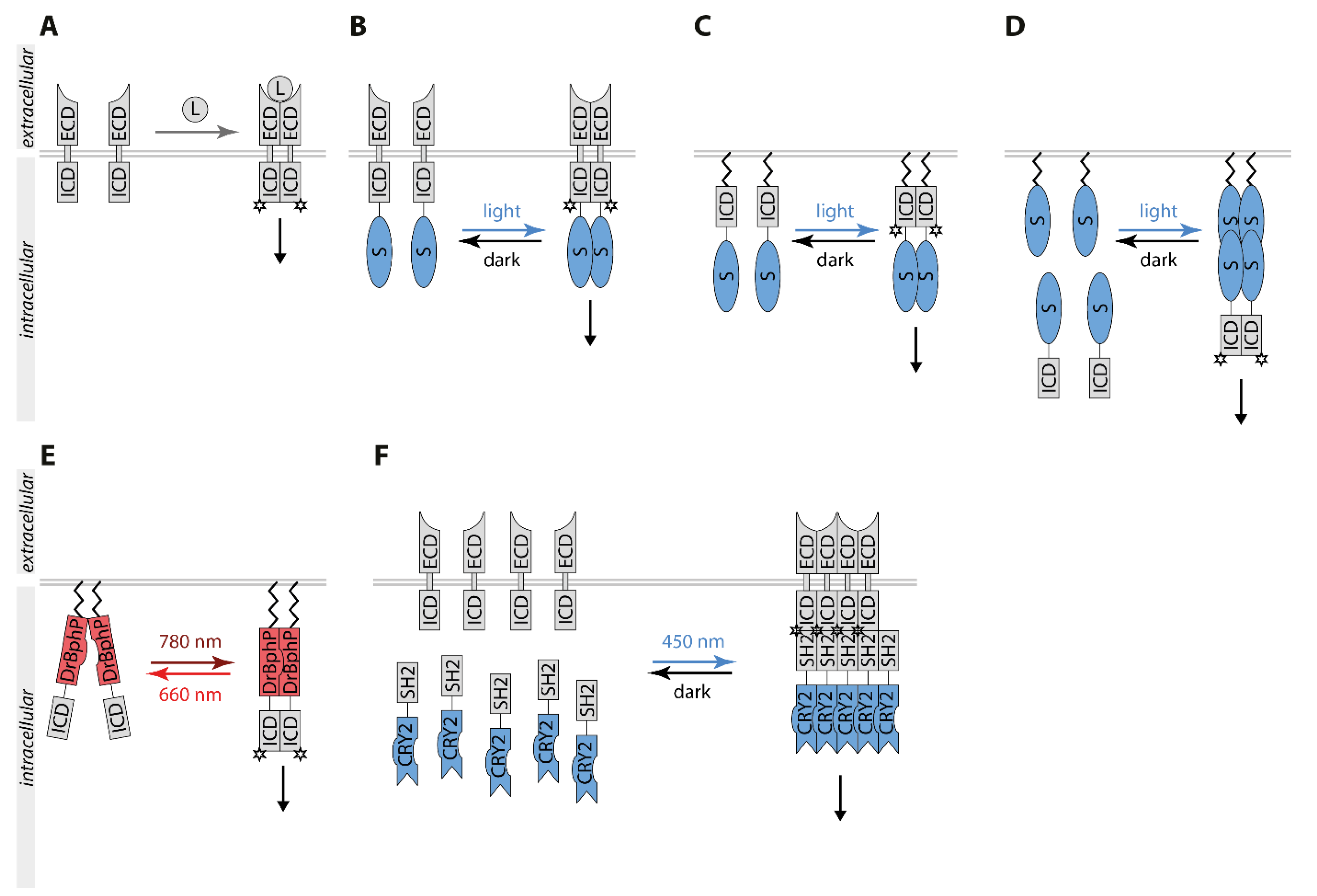
3. Optogenetic Control of Receptor Tyrosine Kinase Signaling
3.1. Light Control of Receptor Tyrosine Kinases
3.2. Optogenetic Control of the RAF/MEK/ERK Axis
3.3. Optogenetic Control of the PI3K/AKT Axis
3.4. Simultaneous Control of Two Pathways by Light
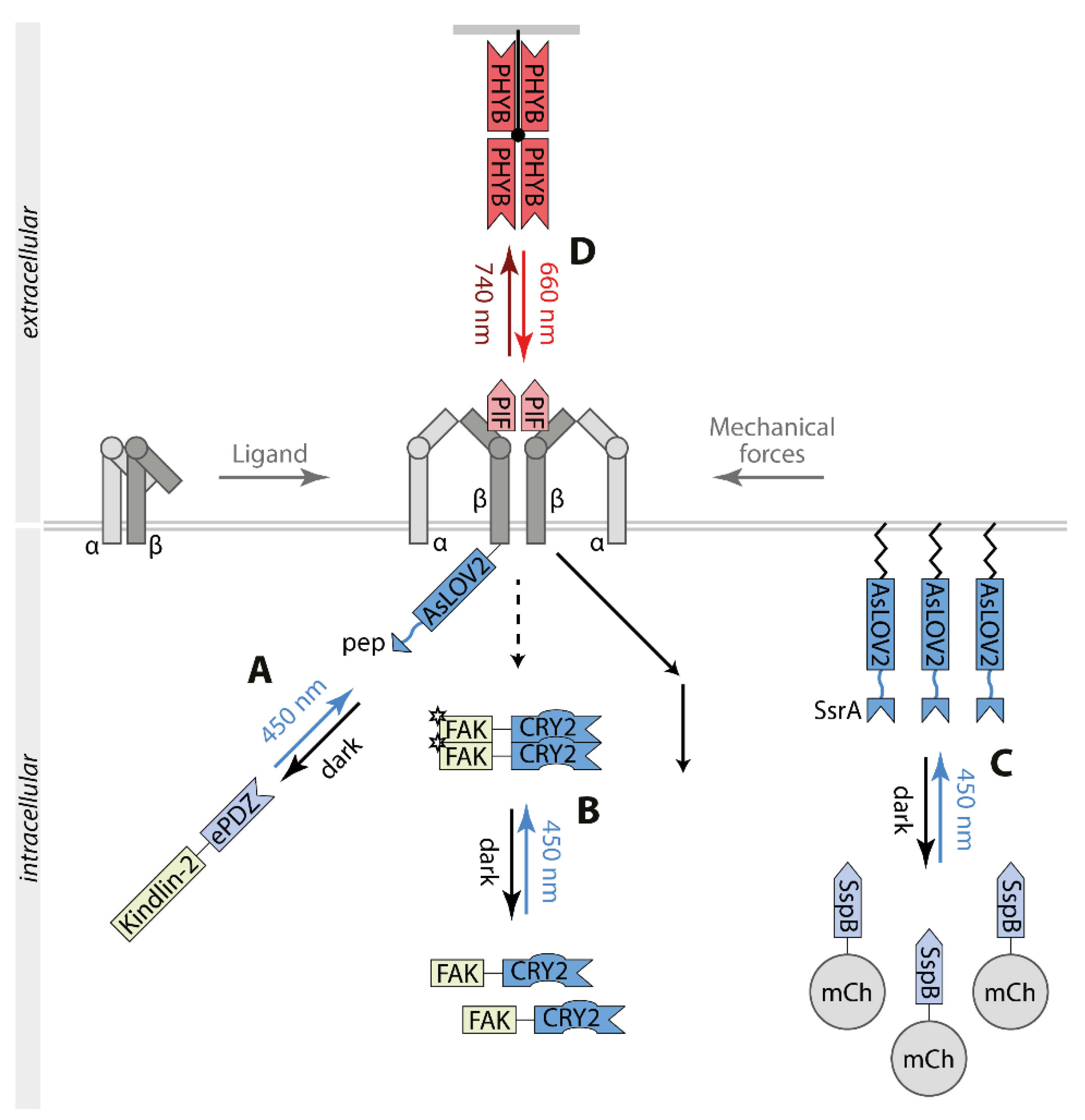
4. Light Control of Integrin Signaling
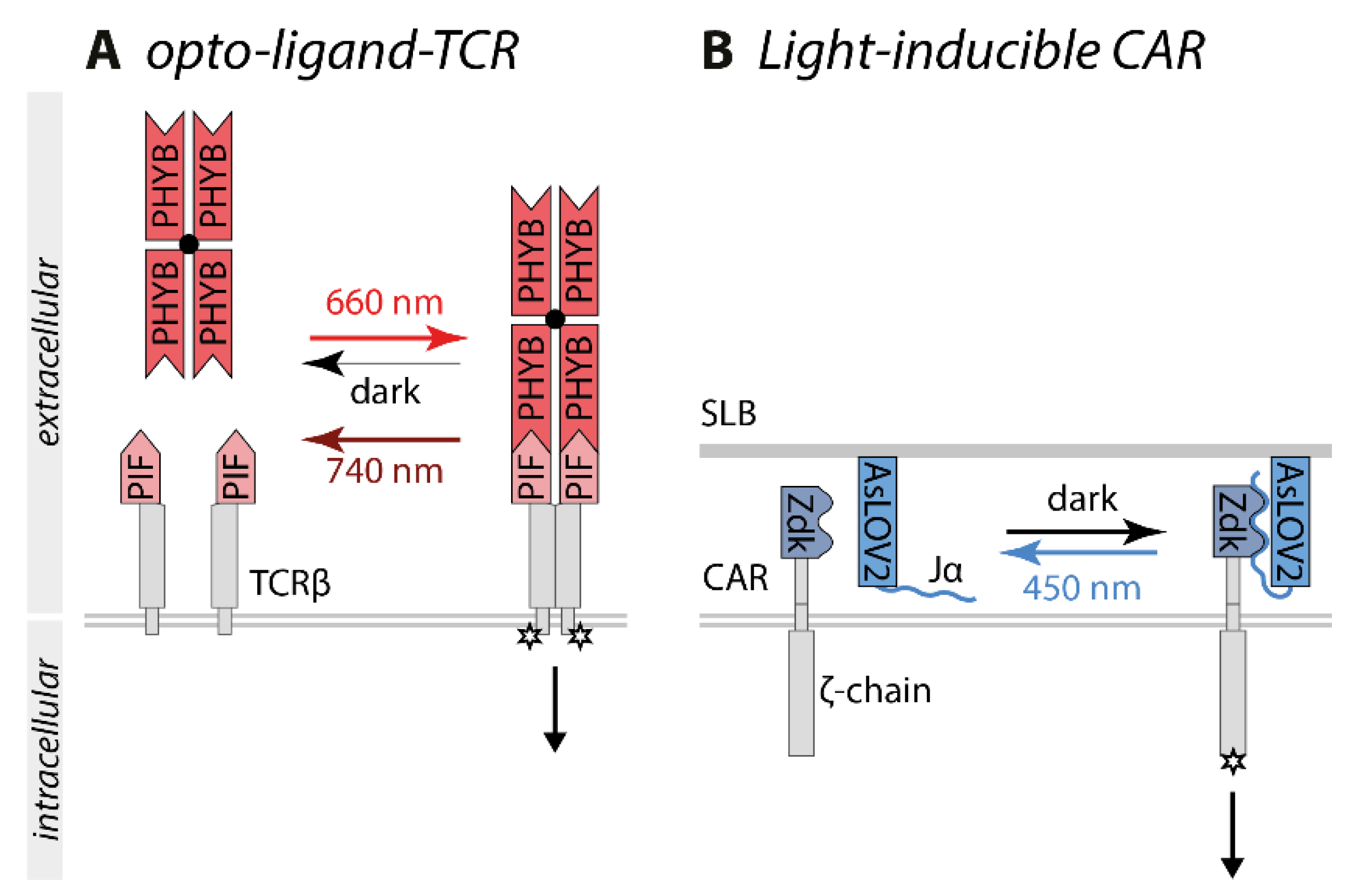
5. Optogenetic Control of T Cell Signaling
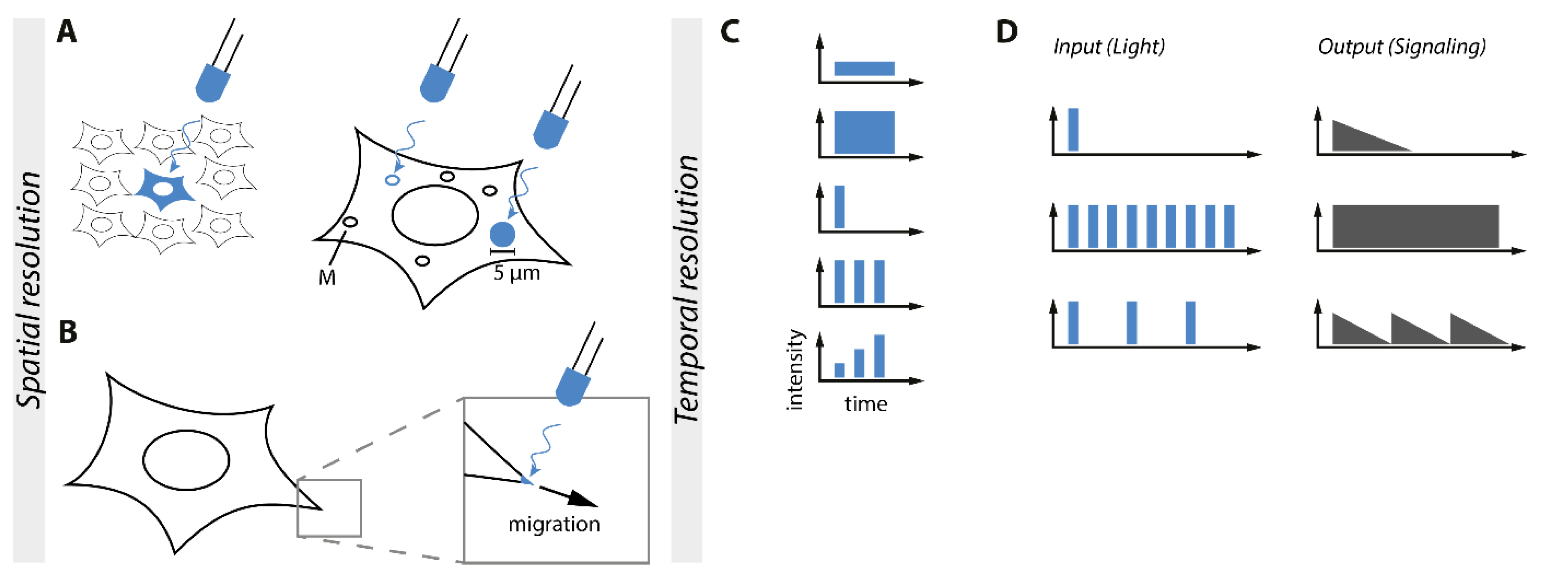
6. Control of Subcellular Localization and Spatial Resolution of Signaling Processes
7. Temporal and Signal Strength Control of Signaling Pathways
8. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AKT | Serine/threonine-protein kinase, protein kinase B |
| AuLOV | Aureochrome 1 light-oxygen-voltage |
| BNDF | Brain-derived growth factor |
| CAR | Chimeric antigen receptor |
| CBD | Cobalamin binding domain |
| CDC42 | Cell division control protein 42 homolog |
| CIB1 | Cryptochrome-interacting basic helix-loop-helix 1 |
| CLICR | Clustering indirectly using CRY2 |
| CMT | Cytoplasm to membrane |
| CNK1 | Connector enhancer of kinase suppressor of ras 1 |
| CPH1 | Cyanobacterial phytochrome-like protein 1 |
| CREB | cAMP-responsive element-binding |
| CRY2 | Cryptochrome 2 |
| DrBphP | Deinococcus radiodurans bacterial phytochrome |
| DTT | Dithiothreitol |
| ECD | Extracellular domain |
| ECM | Extracellular matrix |
| EGFR | Epidermal growth factor receptor |
| EphB2 | Ephrin Type-B receptor 2 |
| ERK | Extracellular signal-regulated protein kinase |
| FAD | Flavin adenine dinucleotide |
| FAK | Focal adhesion kinase |
| FGFR1 | Fibroblast growth factor receptor 1 |
| FRET | Förster resonance energy transfer |
| FYN | Fibroblast yes-related non-receptor kinase |
| GFP | Green fluorescent protein |
| GTP | Guanosine-5′-triphosphate |
| H2O2 | Hydrogen peroxide |
| ICD | Intracellular domain |
| iLID | Improved light-induced dimer |
| iTrkA | Intracellular domain of tropomyosin receptor kinase A |
| LCK | Lymphocyte cell-specific tyrosine-protein kinase |
| LEXY | Light-inducible nuclear export system |
| LINuS | Light-inducible nuclear localization signal |
| LOV | Light-oxygen-voltage |
| LRP6 | Lipoprotein receptor-related protein 6 |
| MAPK | Mitogen-activated protein kinase |
| MEK1 | Dual specificity mitogen-activated protein kinase kinase 1 |
| mTORC2 | Mechanistic target of rapamycin complex 2 |
| NGF | Nerve growth factor |
| OpEN-Tag | Optogenetic endomembrane targeting |
| P2A | 2A self-cleaving peptide |
| pdDronpa | fluorescent protein Dronpa |
| PDK1 | PIP3-dependent kinase 1 |
| PH | Pleckstrin homology |
| PHYB | Phytochrome B |
| PI3K | Phosphatidylinositol-3-kinase |
| PIF | Phytochrome interacting factor |
| PIP2 | Phosphatidylinositol-4,5-bisphosphate |
| PIP3 | Phosphatidylinositol-3,4,5-triphosphate |
| PKA | Protein kinase A |
| PKC | Protein kinase C |
| PLCγ | Phospholipase C γ |
| pMHC | peptide-major histocompatibility complex |
| PTP1B | Protein tyrosine phosphatase 1B |
| Rac1 | Ras-related C3 botulinum toxin substrate 1 |
| RAF | serine/threonine protein kinase, rapidly accelerated fibrosarcoma |
| RhoA | Ras homolog family member A |
| ROS1 | Proto-oncogene tyrosine-protein kinase ROS |
| RTK | Receptor protein kinase |
| SH2 | Src-homology region 2 |
| SOS | Son-of-sevenless |
| STAT3 | Signal transducer and activator of transcription 3 |
| STIM1 | Stromal interaction molecule 1 |
| TCR | T cell receptor |
| TGF | Transforming growth factor |
| TRK | Tropomyosin-related kinase |
| TULIP | Tunable light-inducible dimerization tag |
| TβRI | TGFβ receptor type I |
| TβRII | TGFβ receptor type II |
| VEGF-A | Vascular endothelial growth factor-A |
| VVD | Vivid |
References
- Stanton, B.Z.; Chory, E.J.; Crabtree, G.R. Chemically induced proximity in biology and medicine. Science 2018, 359. [Google Scholar] [CrossRef]
- Oh, T.-J.; Fan, H.; Skeeters, S.S.; Zhang, K. Steering Molecular Activity with Optogenetics: Recent Advances and Perspectives. Adv. Biol. 2021, 2000180. [Google Scholar] [CrossRef]
- Lee, H.N.; Mehta, S.; Zhang, J. Recent advances in the use of genetically encodable optical tools to elicit and monitor signaling events. Curr. Opin. Cell Biol. 2020, 63, 114–124. [Google Scholar] [CrossRef]
- Repina, N.A.; McClave, T.; Johnson, H.J.; Bao, X.; Kane, R.S.; Schaffer, D.V. Engineered Illumination Devices for Optogenetic Control of Cellular Signaling Dynamics. Cell Rep. 2020, 31, 107737. [Google Scholar] [CrossRef]
- Kramer, M.M.; Mühlhäuser, W.W.D.; Weber, W.; Radziwill, G. Multichromatic Control of Signaling Pathways in Mammalian Cells. Adv. Biosyst. 2020, e2000196. [Google Scholar] [CrossRef]
- Hongdusit, A.; Liechty, E.T.; Fox, J.M. Optogenetic interrogation and control of cell signaling. Curr. Opin. Biotechnol. 2020, 66, 195–206. [Google Scholar] [CrossRef]
- Li, Y.; Lee, M.; Kim, N.; Wu, G.; Deng, D.; Kim, J.M.; Liu, X.; Heo, W.D.; Zi, Z. Spatiotemporal Control of TGF-β Signaling with Light. Acs Synth. Biol. 2018, 7, 443–451. [Google Scholar] [CrossRef]
- Kolar, K.; Knobloch, C.; Stork, H.; Žnidarič, M.; Weber, W. OptoBase: A Web Platform for Molecular Optogenetics. ACS Synth. Biol. 2018, 7, 1825–1828. [Google Scholar] [CrossRef]
- Abreu, N.; Levitz, J. Optogenetic Techniques for Manipulating and Sensing G Protein-Coupled Receptor Signaling. Methods Mol. Biol. (Clifton NJ) 2020, 2173, 21–51. [Google Scholar] [CrossRef]
- Rost, B.R.; Schneider-Warme, F.; Schmitz, D.; Hegemann, P. Optogenetic Tools for Subcellular Applications in Neuroscience. Neuron 2017, 96, 572–603. [Google Scholar] [CrossRef]
- Spangler, S.M.; Bruchas, M.R. Optogenetic approaches for dissecting neuromodulation and GPCR signaling in neural circuits. Curr. Opin. Pharmacol. 2017, 32, 56–70. [Google Scholar] [CrossRef]
- Tsukamoto, H.; Furutani, Y. Optogenetic Modulation of Ion Channels by Photoreceptive Proteins. Adv. Exp. Med. Biol. 2021, 1293, 73–88. [Google Scholar] [CrossRef]
- Somuncu, Ö.S.; Berns, H.M.; Sanchez, J.G. New Pioneers of Optogenetics in Neuroscience. Adv. Exp. Med. Biol. 2020, 1288, 47–60. [Google Scholar] [CrossRef]
- Deisseroth, K. Optogenetics: 10 years of microbial opsins in neuroscience. Nat. Neurosci. 2015, 18, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Müller, K.; Siegel, D.; Jahnke, F.R.; Gerrer, K.; Wend, S.; Decker, E.L.; Reski, R.; Weber, W.; Zurbriggen, M.D. A red light-controlled synthetic gene expression switch for plant systems. Mol. Biosyst. 2014, 10, 1679–1688. [Google Scholar] [CrossRef] [PubMed]
- Mühlhäuser, W.W.D.; Weber, W.; Radziwill, G. OpEn-Tag-A Customizable Optogenetic Toolbox To Dissect Subcellular Signaling. ACS Synth. Biol. 2019, 8, 1679–1684. [Google Scholar] [CrossRef]
- Niopek, D.; Wehler, P.; Roensch, J.; Eils, R.; Di Ventura, B. Optogenetic control of nuclear protein export. Nat. Commun. 2016, 7, 10624. [Google Scholar] [CrossRef]
- Levskaya, A.; Weiner, O.D.; Lim, W.A.; Voigt, C.A. Spatiotemporal control of cell signalling using a light-switchable protein interaction. Nature 2009, 461, 997–1001. [Google Scholar] [CrossRef]
- Levskaya, A.; Chevalier, A.A.; Tabor, J.J.; Simpson, Z.B.; Lavery, L.A.; Levy, M.; Davidson, E.A.; Scouras, A.; Ellington, A.D.; Marcotte, E.M.; et al. Engineering Escherichia coli to see light. Nature 2005, 438, 441–442. [Google Scholar] [CrossRef]
- Kennedy, M.J.; Hughes, R.M.; Peteya, L.A.; Schwartz, J.W.; Ehlers, M.D.; Tucker, C.L. Rapid blue-light–mediated induction of protein interactions in living cells. Nat. Methods 2010, 7, 973–975. [Google Scholar] [CrossRef]
- Bugaj, L.J.; Choksi, A.T.; Mesuda, C.K.; Kane, R.S.; Schaffer, D.V. Optogenetic protein clustering and signaling activation in mammalian cells. Nat. Methods 2013, 10, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Swartz, T.E.; Corchnoy, S.B.; Christie, J.M.; Lewis, J.W.; Szundi, I.; Briggs, W.R.; Bogomolni, R.A. The Photocycle of a Flavin-binding Domain of the Blue Light Photoreceptor Phototropin. J. Biol. Chem. 2001, 276, 36493–36500. [Google Scholar] [CrossRef] [PubMed]
- Salomon, M.; Christie, J.M.; Knieb, E.; Lempert, U.; Briggs, W.R. Photochemical and Mutational Analysis of the FMN-Binding Domains of the Plant Blue Light Receptor, Phototropin. Biochemistry 2000, 39, 9401–9410. [Google Scholar] [CrossRef] [PubMed]
- Guntas, G.; Hallett, R.A.; Zimmerman, S.P.; Williams, T.; Yumerefendi, H.; Bear, J.E.; Kuhlman, B. Engineering an improved light-induced dimer (iLID) for controlling the localization and activity of signaling proteins. Proc. Natl. Acad. Sci. USA 2015, 112, 112–117. [Google Scholar] [CrossRef]
- Taslimi, A.; Vrana, J.D.; Chen, D.; Borinskaya, S.; Mayer, B.J.; Kennedy, M.J.; Tucker, C.L. An optimized optogenetic clustering tool for probing protein interaction and function. Nat. Commun. 2014, 5, 4925. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Kim, N.Y.; Lee, S.; Kim, N.; Kim, J.; Heo, W.D. Optogenetic protein clustering through fluorescent protein tagging and extension of CRY2. Nat. Commun. 2017, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Hope, J.; Ong, Q.; Lou, H.-Y.; Kim, N.; McCarthy, C.; Acero, V.; Lin, M.Z.; Cui, B. Understanding CRY2 interactions for optical control of intracellular signaling. Nat. Commun. 2017, 8, 547. [Google Scholar] [CrossRef]
- Wang, H.; Vilela, M.; Winkler, A.; Tarnawski, M.; Schlichting, I.; Yumerefendi, H.; Kuhlman, B.; Liu, R.; Danuser, G.; Hahn, K.M. LOVTRAP: An optogenetic system for photoinduced protein dissociation. Nat. Methods 2016, 13, 755–758. [Google Scholar] [CrossRef]
- Kawano, F.; Suzuki, H.; Furuya, A.; Sato, M. Engineered pairs of distinct photoswitches for optogenetic control of cellular proteins. Nat. Commun. 2015, 6, 6256. [Google Scholar] [CrossRef]
- Strickland, D.; Lin, Y.; Wagner, E.; Hope, C.M.; Zayner, J.; Antoniou, C.; Sosnick, T.R.; Weiss, E.L.; Glotzer, M. TULIPs: Tunable, light-controlled interacting protein tags for cell biology. Nat. Methods 2012, 9, 379–384. [Google Scholar] [CrossRef]
- Grusch, M.; Schelch, K.; Riedler, R.; Reichhart, E.; Differ, C.; Berger, W.; Inglés-Prieto, Á.; Janovjak, H. Spatio-temporally precise activation of engineered receptor tyrosine kinases by light. EMBO J. 2014, 33, 1713–1726. [Google Scholar] [CrossRef]
- Zhou, X.X.; Fan, L.Z.; Li, P.; Shen, K.; Lin, M.Z. Optical control of cell signaling by single-chain photoswitchable kinases. Science 2017, 355, 836–842. [Google Scholar] [CrossRef] [PubMed]
- Kainrath, S.; Stadler, M.; Reichhart, E.; Distel, M.; Janovjak, H. Green-Light-Induced Inactivation of Receptor Signaling Using Cobalamin-Binding Domains. Angew. Chem. Int. Ed. 2017, 56, 4608–4611. [Google Scholar] [CrossRef] [PubMed]
- Leopold, A.V.; Chernov, K.G.; Shemetov, A.A.; Verkhusha, V.V. Neurotrophin receptor tyrosine kinases regulated with near-infrared light. Nat. Commun. 2019, 10, 1129. [Google Scholar] [CrossRef]
- Müller, K.; Engesser, R.; Timmer, J.; Nagy, F.; Zurbriggen, M.D.; Weber, W. Synthesis of phycocyanobilin in mammalian cells. Chem. Commun. 2013, 49, 8970–8972. [Google Scholar] [CrossRef]
- Uda, Y.; Miura, H.; Goto, Y.; Yamamoto, K.; Mii, Y.; Kondo, Y.; Takada, S.; Aoki, K. Improvement of Phycocyanobilin Synthesis for Genetically Encoded Phytochrome-Based Optogenetics. Acs Chem. Biol. 2020, 15, 2896–2906. [Google Scholar] [CrossRef]
- Wu, Y.I.; Frey, D.; Lungu, O.I.; Jaehrig, A.; Schlichting, I.; Kuhlman, B.; Hahn, K.M. A genetically encoded photoactivatable Rac controls the motility of living cells. Nature 2009, 461, 104–108. [Google Scholar] [CrossRef]
- Younas, T.; Vidallon, M.L.P.; Tabor, R.F.; He, L. Open–Closed Structure of Light-Responsive Protein LOV2 Regulates Its Molecular Interaction with a Binding Partner. J. Phys. Chem. Lett. 2020, 11, 8647–8653. [Google Scholar] [CrossRef]
- Niopek, D.; Benzinger, D.; Roensch, J.; Draebing, T.; Wehler, P.; Eils, R.; Di Ventura, B. Engineering light-inducible nuclear localization signals for precise spatiotemporal control of protein dynamics in living cells. Nat. Commun. 2014, 5, 4404. [Google Scholar] [CrossRef]
- Schlessinger, J. Receptor tyrosine kinases: Legacy of the first two decades. Cold Spring Harb. Perspect. Biol. 2014, 6. [Google Scholar] [CrossRef]
- Ferguson, K.M.; Hu, C.; Lemmon, M.A. Insulin and epidermal growth factor receptor family members share parallel activation mechanisms. Protein Sci.: A Publ. Protein Soc. 2020, 29, 1331–1344. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Lovly, C.M. Mechanisms of receptor tyrosine kinase activation in cancer. Mol Cancer 2018, 17, 58. [Google Scholar] [CrossRef] [PubMed]
- Leopold, A.V.; Verkhusha, V.V. Light control of RTK activity: From technology development to translational research. Chem. Sci. 2020, 11, 10019–10034. [Google Scholar] [CrossRef]
- Chang, K.-Y.; Woo, D.; Jung, H.; Lee, S.; Kim, S.; Won, J.; Kyung, T.; Park, H.; Kim, N.; Yang, H.W.; et al. Light-inducible receptor tyrosine kinases that regulate neurotrophin signalling. Nat. Commun. 2014, 5, 4057. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Kim, J.M.; Lee, M.; Kim, C.Y.; Chang, K.-Y.; Heo, W.D. Spatiotemporal control of fibroblast growth factor receptor signals by blue light. Chem. Biol. 2014, 21, 903–912. [Google Scholar] [CrossRef]
- Locke, C.; Machida, K.; Tucker, C.L.; Wu, Y.; Yu, J. Optogenetic activation of EphB2 receptor in dendrites induced actin polymerization by activating Arg kinase. Biol. Open 2017, 6, 1820–1830. [Google Scholar] [CrossRef] [PubMed]
- Alapin, J.M.; Dines, M.; Vassiliev, M.; Tamir, T.; Ram, A.; Locke, C.; Yu, J.; Lamprecht, R. Activation of EphB2 Forward Signaling Enhances Memory Consolidation. Cell Rep. 2018, 23, 2014–2025. [Google Scholar] [CrossRef]
- Inglés-Prieto, Á.; Reichhart, E.; Muellner, M.K.; Nowak, M.; Nijman, S.M.B.; Grusch, M.; Janovjak, H. Light-assisted small-molecule screening against protein kinases. Nat. Chem. Biol. 2015, 11, 952–954. [Google Scholar] [CrossRef]
- Csanaky, K.; Hess, M.W.; Klimaschewski, L. Membrane-Associated, Not Cytoplasmic or Nuclear, FGFR1 Induces Neuronal Differentiation. Cells 2019, 8, 243. [Google Scholar] [CrossRef]
- Reichhart, E.; Ingles-Prieto, A.; Tichy, A.-M.; McKenzie, C.; Janovjak, H. A Phytochrome Sensory Domain Permits Receptor Activation by Red Light. Angew. Chem. Int. Ed. 2016, 55, 6339–6342. [Google Scholar] [CrossRef]
- Duan, L.; Hope, J.M.; Guo, S.; Ong, Q.; François, A.; Kaplan, L.; Scherrer, G.; Cui, B. Optical Activation of TrkA Signaling. Acs Synth. Biol. 2018, 7, 1685–1693. [Google Scholar] [CrossRef]
- Krishnamurthy, V.V.; Fu, J.; Oh, T.-J.; Khamo, J.; Yang, J.; Zhang, K. A Generalizable Optogenetic Strategy to Regulate Receptor Tyrosine Kinases during Vertebrate Embryonic Development. J. Mol. Biol. 2020, 432, 3149–3158. [Google Scholar] [CrossRef]
- Hope, J.M.; Liu, A.; Calvin, G.J.; Cui, B. Construction of Light-Activated Neurotrophin Receptors Using the Improved Light-Induced Dimerizer (iLID). J. Mol. Biol. 2020, 432, 3739–3748. [Google Scholar] [CrossRef] [PubMed]
- Leopold, A.V.; Pletnev, S.; Verkhusha, V.V. Bacterial Phytochrome as a Scaffold for Engineering of Receptor Tyrosine Kinases Controlled with Near-Infrared Light. J. Mol. Biol. 2020, 432, 3749–3760. [Google Scholar] [CrossRef] [PubMed]
- Khamo, J.S.; Krishnamurthy, V.V.; Chen, Q.; Diao, J.; Zhang, K. Optogenetic Delineation of Receptor Tyrosine Kinase Subcircuits in PC12 Cell Differentiation. Cell Chem. Biol. 2019, 26, 400–410.e3. [Google Scholar] [CrossRef] [PubMed]
- Bugaj, L.J.; Spelke, D.P.; Mesuda, C.K.; Varedi, M.; Kane, R.S.; Schaffer, D.V. Regulation of endogenous transmembrane receptors through optogenetic Cry2 clustering. Nat. Commun. 2015, 6, 6898. [Google Scholar] [CrossRef] [PubMed]
- Toettcher, J.E.; Weiner, O.D.; Lim, W.A. Using optogenetics to interrogate the dynamic control of signal transmission by the Ras/Erk module. Cell 2013, 155, 1422–1434. [Google Scholar] [CrossRef]
- Johnson, H.E.; Goyal, Y.; Pannucci, N.L.; Schüpbach, T.; Shvartsman, S.Y.; Toettcher, J.E. The Spatiotemporal Limits of Developmental Erk Signaling. Dev. Cell 2017, 40, 185–192. [Google Scholar] [CrossRef]
- Wend, S.; Wagner, H.J.; Müller, K.; Zurbriggen, M.D.; Weber, W.; Radziwill, G. Optogenetic control of protein kinase activity in mammalian cells. ACS Synth. Biol. 2014, 3, 280–285. [Google Scholar] [CrossRef]
- Zhang, K.; Duan, L.; Ong, Q.; Lin, Z.; Varman, P.M.; Sung, K.; Cui, B. Light-mediated kinetic control reveals the temporal effect of the Raf/MEK/ERK pathway in PC12 cell neurite outgrowth. PLoS ONE 2014, 9, e92917. [Google Scholar] [CrossRef]
- Krishnamurthy, V.V.; Khamo, J.S.; Mei, W.; Turgeon, A.J.; Ashraf, H.M.; Mondal, P.; Patel, D.B.; Risner, N.; Cho, E.E.; Yang, J.; et al. Reversible optogenetic control of kinase activity during differentiation and embryonic development. Development 2016, 143, 4085–4094. [Google Scholar] [CrossRef]
- Patel, A.L.; Yeung, E.; McGuire, S.E.; Wu, A.Y.; Toettcher, J.E.; Burdine, R.D.; Shvartsman, S.Y. Optimizing photoswitchable MEK. Proc. Natl. Acad. Sci. USA 2019, 116, 25756–25763. [Google Scholar] [CrossRef] [PubMed]
- Idevall-Hagren, O.; Dickson, E.J.; Hille, B.; Toomre, D.K.; de Camilli, P. Optogenetic control of phosphoinositide metabolism. Proc. Natl. Acad. Sci. USA 2012, 109, E2316–E2323. [Google Scholar] [CrossRef]
- Kakumoto, T.; Nakata, T. Optogenetic control of PIP3: PIP3 is sufficient to induce the actin-based active part of growth cones and is regulated via endocytosis. PLoS ONE 2013, 8, e70861. [Google Scholar] [CrossRef] [PubMed]
- Toettcher, J.E.; Gong, D.; Lim, W.A.; Weiner, O.D. Light-based feedback for controlling intracellular signaling dynamics. Nat. Methods 2011, 8, 837–839. [Google Scholar] [CrossRef] [PubMed]
- Katsura, Y.; Kubota, H.; Kunida, K.; Kanno, A.; Kuroda, S.; Ozawa, T. An optogenetic system for interrogating the temporal dynamics of Akt. Sci. Rep. 2015, 5, 14589. [Google Scholar] [CrossRef]
- Mühlhäuser, W.W.D.; Hörner, M.; Weber, W.; Radziwill, G. Light-Regulated Protein Kinases Based on the CRY2-CIB1 System. Methods Mol. Biol. (Clifton NJ) 2017, 1596, 257–270. [Google Scholar] [CrossRef]
- Fischer, A.; Warscheid, B.; Weber, W.; Radziwill, G. Optogenetic clustering of CNK1 reveals mechanistic insights in RAF and AKT signalling controlling cell fate decisions. Sci. Rep. 2016, 6, 38155. [Google Scholar] [CrossRef]
- Wilson, M.Z.; Ravindran, P.T.; Lim, W.A.; Toettcher, J.E. Tracing Information Flow from Erk to Target Gene Induction Reveals Mechanisms of Dynamic and Combinatorial Control. Mol. Cell 2017, 67, 757–769.e5. [Google Scholar] [CrossRef]
- Johnson, H.E.; Toettcher, J.E. Signaling Dynamics Control Cell Fate in the Early Drosophila Embryo. Dev. Cell 2019, 48, 361–370.e3. [Google Scholar] [CrossRef]
- Chatelle, C.V.; Hövermann, D.; Müller, A.; Wagner, H.J.; Weber, W.; Radziwill, G. Optogenetically controlled RAF to characterize BRAF and CRAF protein kinase inhibitors. Sci. Rep. 2016, 6, 23713. [Google Scholar] [CrossRef]
- Xu, Y.; Nan, D.; Fan, J.; Bogan, J.S.; Toomre, D. Optogenetic activation reveals distinct roles of PIP3 and Akt in adipocyte insulin action. J. Cell Sci. 2016, 129, 2085–2095. [Google Scholar] [CrossRef] [PubMed]
- Ong, Q.; Guo, S.; Duan, L.; Zhang, K.; Collier, E.A.; Cui, B. The Timing of Raf/ERK and AKT Activation in Protecting PC12 Cells against Oxidative Stress. PLoS ONE 2016, 11, e0153487. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Fan, H.; Li, F.; Skeeters, S.S.; Krishnamurthy, V.V.; Song, Y.; Zhang, K. Optical control of ERK and AKT signaling promotes axon regeneration and functional recovery of PNS and CNS in Drosophila. eLife 2020, 9. [Google Scholar] [CrossRef]
- Müller, K.; Engesser, R.; Schulz, S.; Steinberg, T.; Tomakidi, P.; Weber, C.C.; Ulm, R.; Timmer, J.; Zurbriggen, M.D.; Weber, W. Multi-chromatic control of mammalian gene expression and signaling. Nucleic Acids Res. 2013, 41, e124. [Google Scholar] [CrossRef]
- Bugaj, L.J.; Lim, W.A. High-throughput multicolor optogenetics in microwell plates. Nat. Protoc. 2019, 14, 2205–2228. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Weber, W.; Warscheid, B.; Radziwill, G. AKT-dependent phosphorylation of the SAM domain induces oligomerization and activation of the scaffold protein CNK1. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2017, 1864, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Mühlhäuser, W.W.D.; Warscheid, B.; Radziwill, G. Membrane localization of acetylated CNK1 mediates a positive feedback on RAF/ERK signaling. Sci. Adv. 2017, 3, e1700475. [Google Scholar] [CrossRef]
- Lietha, D.; Izard, T. Roles of Membrane Domains in Integrin-Mediated Cell Adhesion. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Kasirer-Friede, A.; Shattil, S.J. Optogenetic interrogation of integrin αVβ3 function in endothelial cells. J. Cell Sci. 2017, 130, 3532–3541. [Google Scholar] [CrossRef]
- Baaske, J.; Mühlhäuser, W.W.D.; Yousefi, O.S.; Zanner, S.; Radziwill, G.; Hörner, M.; Schamel, W.W.A.; Weber, W. Optogenetic control of integrin-matrix interaction. Commun. Biol. 2019, 2, 15. [Google Scholar] [CrossRef]
- Meshik, X.; O’Neill, P.R.; Gautam, N. Physical Plasma Membrane Perturbation Using Subcellular Optogenetics Drives Integrin-Activated Cell Migration. ACS Synth. Biol. 2019, 8, 498–510. [Google Scholar] [CrossRef] [PubMed]
- Hörner, M.; Chatelle, C.; Mühlhäuser, W.W.D.; Stocker, D.R.; Coats, M.; Weber, W.; Radziwill, G. Optogenetic control of focal adhesion kinase signaling. Cell. Signal. 2018, 42, 176–183. [Google Scholar] [CrossRef]
- Ricken, J. Photo-Switchable Proteins for Cell-Matrix Adhesion; Heidelberg University Library: Heidelberg, Germany, 2019. [Google Scholar]
- Xu, D.; Ricken, J.; Wegner, S.V. Turning Cell Adhesions ON or OFF with High Spatiotemporal Precision Using the Green Light Responsive Protein CarH. Chemistry (Weinh. Der Bergstr. Ger.) 2020, 26, 9859–9863. [Google Scholar] [CrossRef]
- Wu, L.; Wei, Q.; Brzostek, J.; Gascoigne, N.R.J. Signaling from T cell receptors (TCRs) and chimeric antigen receptors (CARs) on T cells. Cell. Mol. Immunol. 2020, 17, 600–612. [Google Scholar] [CrossRef]
- Yousefi, O.S.; Günther, M.; Hörner, M.; Chalupsky, J.; Wess, M.; Brandl, S.M.; Smith, R.W.; Fleck, C.; Kunkel, T.; Zurbriggen, M.D.; et al. Optogenetic control shows that kinetic proofreading regulates the activity of the T cell receptor. eLife 2019, 8, e42475. [Google Scholar] [CrossRef]
- Tischer, D.K.; Weiner, O.D.; Chakraborty, A.K. Light-based tuning of ligand half-life supports kinetic proofreading model of T cell signaling. eLife 2019, 8, e42498. [Google Scholar] [CrossRef]
- O’Donoghue, G.P.; Bugaj, L.J.; Anderson, W.; Daniels, K.G.; Rawlings, D.J.; Lim, W.A. T cells selectively filter oscillatory signals on the minutes timescale. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef]
- Ma, Y.; Lim, Y.J.; Benda, A.; Lou, J.; Goyette, J.; Gaus, K. Clustering of the ζ-Chain Can Initiate T Cell Receptor Signaling. Int. J. Mol. Sci. 2020, 21, 3498. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, T.; Dema, A.; van Haren, J. Lights, cytoskeleton, action: Optogenetic control of cell dynamics. Curr. Opin. Cell Biol. 2020, 66, 1–10. [Google Scholar] [CrossRef]
- Kichuk, T.C.; Carrasco-López, C.; Avalos, J.L. Lights up on organelles: Optogenetic tools to control subcellular structure and organization. Wiley Interdiscip. Rev. Syst. Biol. Med. 2021, 13, e1500. [Google Scholar] [CrossRef] [PubMed]
- Cavanaugh, K.E.; Staddon, M.F.; Munro, E.; Banerjee, S.; Gardel, M.L. RhoA Mediates Epithelial Cell Shape Changes via Mechanosensitive Endocytosis. Dev. Cell 2020, 52, 152–166.e5. [Google Scholar] [CrossRef] [PubMed]
- Berlew, E.E.; Kuznetsov, I.A.; Yamada, K.; Bugaj, L.J.; Boerckel, J.D.; Chow, B.Y. Single-component optogenetic tools for inducible RhoA GTPase signaling. bioRxiv 2021, 2021.02.01.429147. [Google Scholar] [CrossRef]
- Hongdusit, A.; Zwart, P.H.; Sankaran, B.; Fox, J.M. Minimally disruptive optical control of protein tyrosine phosphatase 1B. Nat. Commun. 2020, 11, 788. [Google Scholar] [CrossRef]
- Yang, L.; Ozawa, T.; Dong, H.; Zhang, X. Optogenetic Control of Phosphatidylinositol (3,4,5)-triphosphate Production by Light-sensitive Cryptochrome Proteins on the Plasma Membrane. Chin. J. Chem. 2021. [Google Scholar] [CrossRef]
- Ma, G.; He, L.; Liu, S.; Xie, J.; Huang, Z.; Jing, J.; Lee, Y.-T.; Wang, R.; Luo, H.; Han, W.; et al. Optogenetic engineering to probe the molecular choreography of STIM1-mediated cell signaling. Nat. Commun. 2020, 11, 1039. [Google Scholar] [CrossRef]
- O’Banion, C.P.; Priestman, M.A.; Hughes, R.M.; Herring, L.E.; Capuzzi, S.J.; Lawrence, D.S. Design and Profiling of a Subcellular Targeted Optogenetic cAMP-Dependent Protein Kinase. Cell Chem. Biol. 2018, 25, 100–109.e8. [Google Scholar] [CrossRef]
- Bugaj, L.J.; Sabnis, A.J.; Mitchell, A.; Garbarino, J.E.; Toettcher, J.E.; Bivona, T.G.; Lim, W.A. Cancer mutations and targeted drugs can disrupt dynamic signal encoding by the Ras-Erk pathway. Science 2018, 361. [Google Scholar] [CrossRef]
- Thomas, O.S.; Hörner, M.; Weber, W. A graphical user interface to design high-throughput optogenetic experiments with the optoPlate-96. Nat. Protoc. 2020, 15, 2785–2787. [Google Scholar] [CrossRef]
- Gil, A.A.; Carrasco-López, C.; Zhu, L.; Zhao, E.M.; Ravindran, P.T.; Wilson, M.Z.; Goglia, A.G.; Avalos, J.L.; Toettcher, J.E. Optogenetic control of protein binding using light-switchable nanobodies. Nat. Commun. 2020, 11, 4044. [Google Scholar] [CrossRef]
- Godwin, W.C.; Hoffmann, G.F.; Gray, T.J.; Hughes, R.M. Imaging of morphological and biochemical hallmarks of apoptosis with optimized optogenetic tools. J. Biol. Chem. 2019, 294, 16918–16929. [Google Scholar] [CrossRef]
- Zhang, M.; Lin, X.; Zhang, J.; Su, L.; Ma, M.; Ea, V.L.; Liu, X.; Wang, L.; Chang, J.; Li, X.; et al. Blue light-triggered optogenetic system for treating uveal melanoma. Oncogene 2020, 39, 2118–2124. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zheng, Y.; Bhusari, S.; Villiou, M.; Pearson, S.; Campo, A. Printed Degradable Optical Waveguides for Guiding Light into Tissue. Adv. Funct. Mater. 2020, 30, 2004327. [Google Scholar] [CrossRef]
- Ye, H.; Daoud-El Baba, M.; Peng, R.-W.; Fussenegger, M. A synthetic optogenetic transcription device enhances blood-glucose homeostasis in mice. Science 2011, 332, 1565–1568. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Hyun, Y.-M.; Lim, K.; Lee, H.; Cummings, R.J.; Gerber, S.A.; Bae, S.; Cho, T.Y.; Lord, E.M.; Kim, M. Optogenetic control of chemokine receptor signal and T-cell migration. Proc. Natl. Acad. Sci. USA 2014, 111, 6371–6376. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Shou, C.; He, M.; Xu, J.; Cheng, Y.; Yuan, Z.; Lan, M.; Zhao, Y.; Yang, Y.; Chen, X.; et al. A combination of LightOn gene expression system and tumor microenvironment-responsive nanoparticle delivery system for targeted breast cancer therapy. Acta Pharm. Sin. B 2020, 10, 1741–1753. [Google Scholar] [CrossRef]
- Sankaran, S.; Becker, J.; Wittmann, C.; Del Campo, A. Optoregulated Drug Release from an Engineered Living Material: Self-Replenishing Drug Depots for Long-Term, Light-Regulated Delivery. Small 2019, 15, e1804717. [Google Scholar] [CrossRef]
- Hagihara, Y.; Sakamoto, A.; Tokuda, T.; Yamashita, T.; Ikemoto, S.; Kimura, A.; Haruta, M.; Sasagawa, K.; Ohta, J.; Takayama, K.; et al. Photoactivatable oncolytic adenovirus for optogenetic cancer therapy. Cell Death Dis. 2020, 11, 1–9. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Switch | Co-Factor | Mode of Action | λ (Excitation) | λ (Reversion) | Excitation Time | Reversion Time | Selected Publications |
|---|---|---|---|---|---|---|---|---|
| Crypto-chromes | CRY2/CIB1 | FAD | heterodimerization | 450 nm | dark | seconds | minutes | [20] |
| CRY2/CRY2 | FAD | homo-oligomerization | 450 nm | dark | seconds | minutes | [21] | |
| CRY2olig | FAD | homo-oligomerization | 450 nm | dark | seconds | minutes | [25] | |
| CRY2clust | FAD | homo-oligomerization | 450 nm | dark | seconds | minutes | [26] | |
| CRY2high | FAD | homo-oligomerization | 450 nm | dark | seconds | minutes | [27] | |
| CRY2low | FAD | heterodimerization | 450 nm | dark | seconds | minutes | [27] | |
| LOV domains | AsLOV2 | FMN | intramolecular conformational change | 450 nm | dark | seconds | tens of seconds | [22,23] |
| iLID | FMN | heterodimerization | 450 nm | dark | seconds | seconds to minutes | [24] | |
| LOVTRAP | FMN | heterodimerization, dissociation | 450 nm | dark | seconds | seconds to minutes | [28] | |
| Magnets | FAD | heterodimerization | 450 nm | dark | seconds | seconds to hours (variants available) | [29] | |
| TULIP | FMN | heterodimerization | 450 nm | dark | seconds | seconds to minutes | [30] | |
| VfAU1-LOV | FMN | homodimerization | 450 nm | dark | seconds | minutes | [31] | |
| Fluorescent proteins | pdDronpa1 | none | homodimerization, dissociation | 500 nm | 400 nm | seconds | seconds | [32] |
| Cobalamin-binding domains | MxCBD | CBD | homotetramerization, dissociation | 545 nm | dark | n.d. | n.d. | [33] |
| TtCBD | CBD | homotetramerization, dissociation | 545 nm | dark | n.d. | n.d. | [33] | |
| Phyto-chromes | CPH1 | PCB | homodimerization | 660 nm | 740 nm, dark | milliseconds | milliseconds | [19] |
| DrBphP | Biliverdin | homodimerization, dissociation | 660 nm | 780 nm, dark | n.d. | n.d. | [34] | |
| PHYB/PIF3 & PHYB/PIF6 | PCB | heterodimerization | 660 nm | 740 nm, dark | milliseconds | milliseconds | [18] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kramer, M.M.; Lataster, L.; Weber, W.; Radziwill, G. Optogenetic Approaches for the Spatiotemporal Control of Signal Transduction Pathways. Int. J. Mol. Sci. 2021, 22, 5300. https://doi.org/10.3390/ijms22105300
Kramer MM, Lataster L, Weber W, Radziwill G. Optogenetic Approaches for the Spatiotemporal Control of Signal Transduction Pathways. International Journal of Molecular Sciences. 2021; 22(10):5300. https://doi.org/10.3390/ijms22105300
Chicago/Turabian StyleKramer, Markus M., Levin Lataster, Wilfried Weber, and Gerald Radziwill. 2021. "Optogenetic Approaches for the Spatiotemporal Control of Signal Transduction Pathways" International Journal of Molecular Sciences 22, no. 10: 5300. https://doi.org/10.3390/ijms22105300
APA StyleKramer, M. M., Lataster, L., Weber, W., & Radziwill, G. (2021). Optogenetic Approaches for the Spatiotemporal Control of Signal Transduction Pathways. International Journal of Molecular Sciences, 22(10), 5300. https://doi.org/10.3390/ijms22105300