Comparative Analysis Delineates the Transcriptional Resistance Mechanisms for Pod Borer Resistance in the Pigeonpea Wild Relative Cajanus scarabaeoides (L.) Thouars

 , , ,
, , ,
Abstract
1. Introduction
2. Results
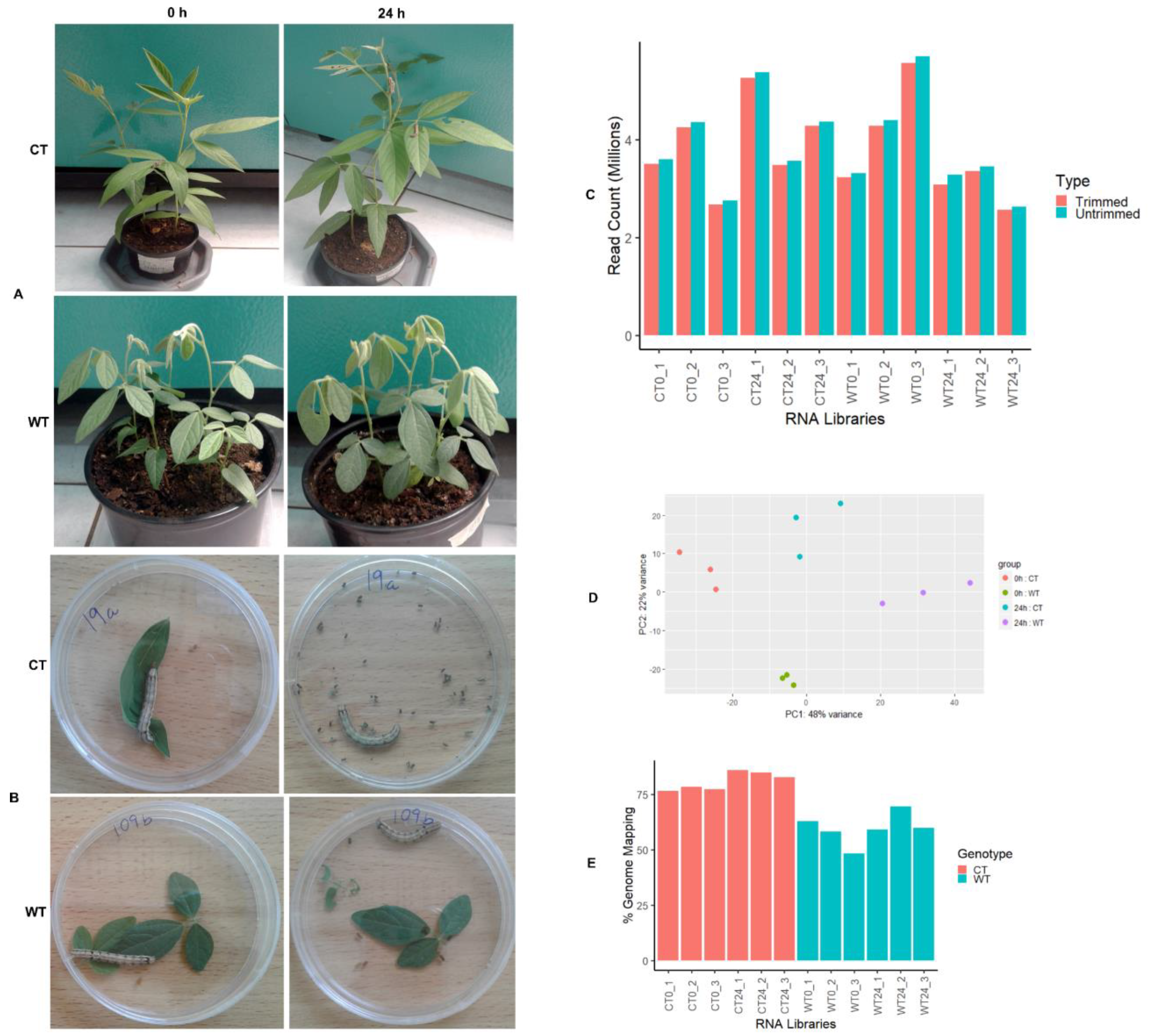
2.1. Wild Pigeonpea Cajanus scarabaeoides Is Resistant to Helicoverpa armigera Herbivory
2.2. Generation of Transcriptomes from Susceptible and Resistant Pigeonpea Accessions
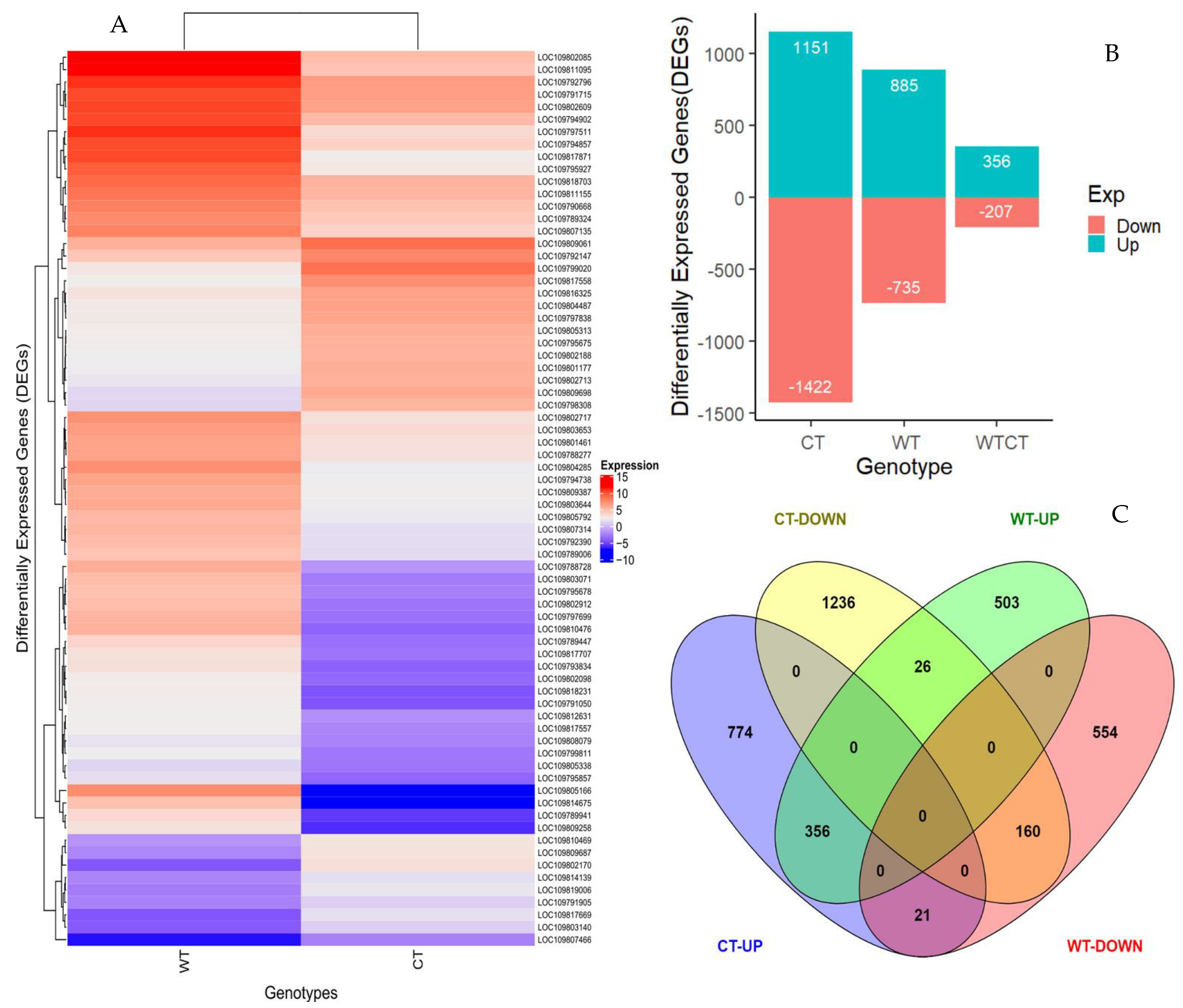
2.3. Herbivory-Induced Transcriptome Changes between Susceptible Cultivated and Tolerant Wild Pigeonpea Genotypes
2.4. Plant Hormones and Their Role in Plant Defense during Insect Herbivory
2.5. Transcription Factors Associated with H. armigera Herbivory
2.6. Calcium/Calmodulin-Mediated Signaling in H. armigera Resistance
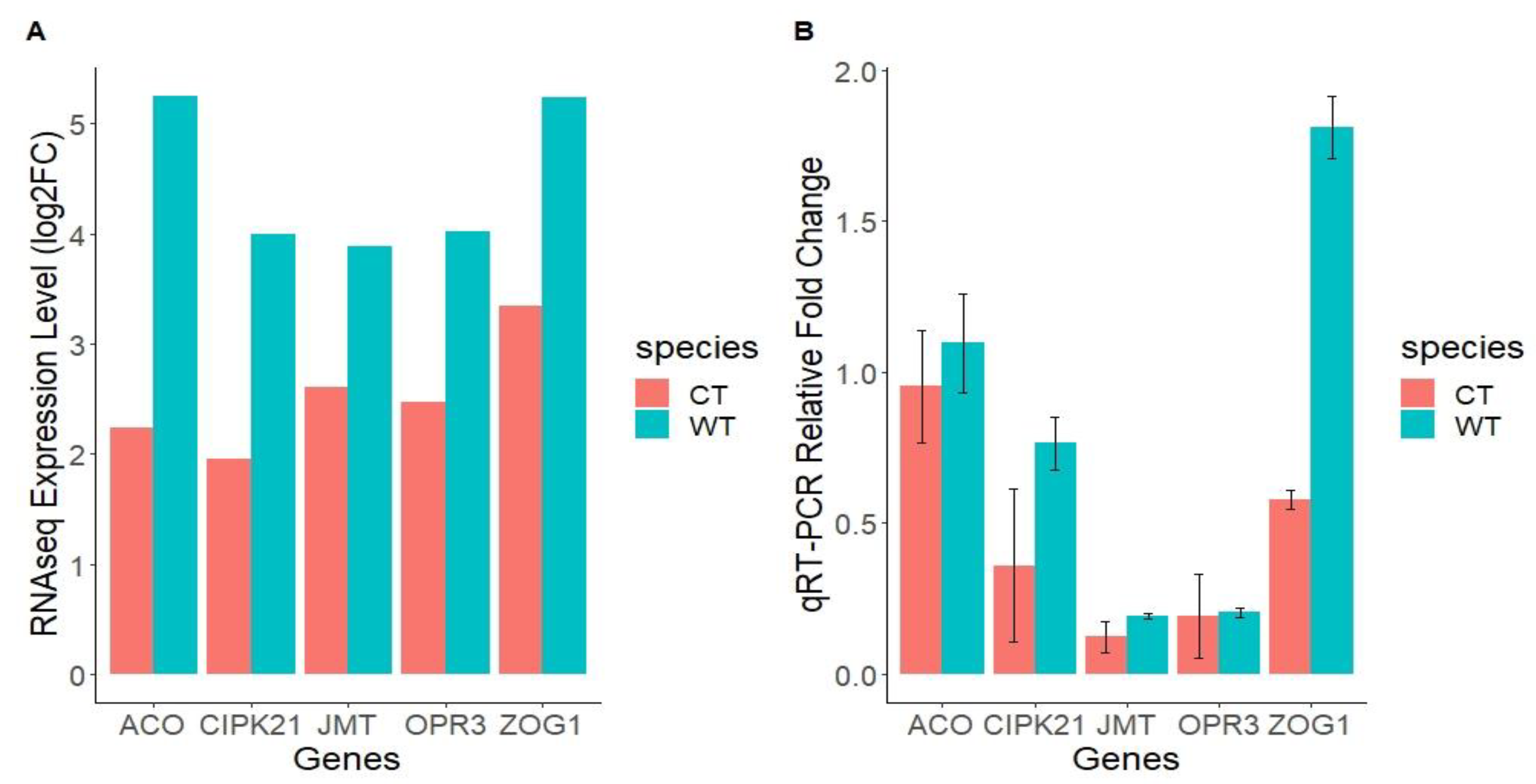
2.7. Validation of RNA-Seq Analysis by qRT-PCR
2.8. Antinutritional Factors and Herbivore-Induced Plant Volatiles and Metabolites
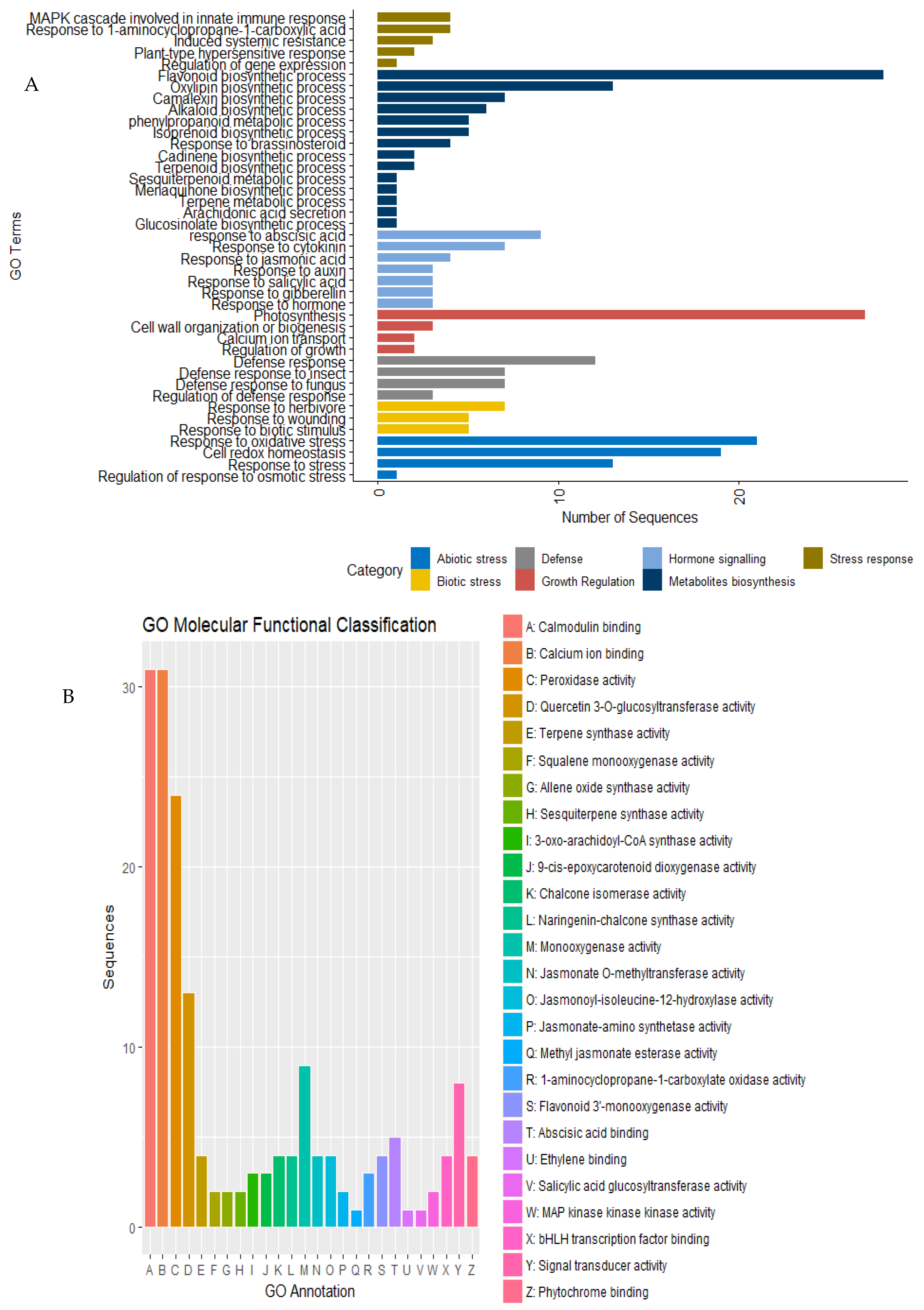
2.9. Gene Ontology Analysis Revealed Genes Enriched in Signaling and Defense-Related Pathways
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Insect Bioassay
4.3. Total RNA Extraction and High-Throughput Sequencing
4.4. Gene Expression Validation with Real-Time Quantitative Reverse Transcription PCR (qRT-PCR)
4.5. Differential Expression and GO Enrichment Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TF | Transcription factor |
| ET | Ethylene |
| JA | Jasmonic acid |
| DEG | Differentially Expressed Genes |
| CPM | Counts per million |
| ERF | Ethylene-responsive factor |
| CHS | Chalcone synthase |
| CWRs | Crop Wild Relatives |
References
- FAO. FAO Statistics Division; FAO: Rome, Italy, 2018. [Google Scholar]
- Peoples, M.; Herridge, D.; Ladha, J. Biological nitrogen fixation: An efficient source of nitrogen for sustainable agricultural production? Plant Soil 1995, 174, 3–28. [Google Scholar] [CrossRef]
- Shanower, T.; Romeis, J.; Minja, E. Insect pests of pigeonpea and their management. Annu. Rev. Entomol. 1999, 44, 77–96. [Google Scholar] [CrossRef] [PubMed]
- Kambrekar, D. Management of Legume Podborer, Helicoverpa armigera with Host Plant Resistance. Legume Genom. Genet. 2016, 7. [Google Scholar] [CrossRef]
- Sharma, H.; Sujana, G.; Rao, D.M. Morphological and chemical components of resistance to pod borer, Helicoverpa armigera in wild relatives of pigeonpea. Arthropod Plant Interact. 2009, 3, 151–161. [Google Scholar] [CrossRef]
- Aruna, R.; Rao, D.M.; Reddy, L.; Upadhyaya, H.D.; Sharma, H.C. Inheritance of trichomes and resistance to pod borer (Helicoverpa armigera) and their association in interspecific crosses between cultivated pigeonpea (Cajanus cajan) and its wild relative C. scarabaeoides. Euphytica 2005, 145, 247–257. [Google Scholar] [CrossRef]
- Sharma, H.C.; Saxena, K.; Bhagwat, V. The Legume Pod Borer, Maruca Vitrata: Bionomics and Management; ICRISAT: Patancheru, India, 1999. [Google Scholar]
- Reed, W.; Lateef, S. Pigeonpea: Pest Management; Pigeonpea, CABI: Wallingford, UK, 1990; pp. 349–374. [Google Scholar]
- Green, P.; Sharma, H.; Stevenson, P.; Simmonds, M. Susceptibility of pigeonpea and some of its wild relatives to predation by Helicoverpa armigera: Implications for breeding resistant cultivars. Aust. J. Agric. Res. 2006, 57, 831–836. [Google Scholar] [CrossRef]
- Sahu, A.R.; Mishra, R.R.; Panigrahi, J. Estimation of genetic diversity among 34 genotypes in the genus Cajanus with contrasting host response to the pod borer and its allied pests. J. Crop Sci. Biotechnol. 2016, 19, 17–28. [Google Scholar] [CrossRef]
- Upadhyaya, H. Crop germplasm and wild relatives: A source of novel variation for crop improvement. Korean J. Crop Sci. 2008, 53, 12–17. [Google Scholar]
- Muñoz, N.; Liu, A.; Kan, L.; Li, M.-W.; Lam, H.-M. Potential uses of wild germplasms of grain legumes for crop improvement. Int. J. Mol. Sci. 2017, 18, 328. [Google Scholar] [CrossRef]
- Zhang, H.; Mittal, N.; Leamy, L.J.; Barazani, O.; Song, B.H. Back into the wild—Apply untapped genetic diversity of wild relatives for crop improvement. Evol. Appl. 2017, 10, 5–24. [Google Scholar] [CrossRef]
- Hyten, D.L.; Cannon, S.B.; Song, Q.; Weeks, N.; Fickus, E.W.; Shoemaker, R.C.; Specht, J.E.; Farmer, A.D.; May, G.D.; Cregan, P.B. High-throughput SNP discovery through deep resequencing of a reduced representation library to anchor and orient scaffolds in the soybean whole genome sequence. BMC Genom. 2010, 11, 38. [Google Scholar] [CrossRef]
- Akhunov, E.; Nicolet, C.; Dvorak, J. Single nucleotide polymorphism genotyping in polyploid wheat with the Illumina GoldenGate assay. Appl. Genet. 2009, 119, 507–517. [Google Scholar] [CrossRef]
- Mithöfer, A.; Boland, W. Recognition of herbivory-associated molecular patterns. Plant Physiol. 2008, 146, 825–831. [Google Scholar] [CrossRef]
- Arimura, G.-I.; Ozawa, R.; Maffei, M.E. Recent advances in plant early signaling in response to herbivory. Int. J. Mol. Sci. 2011, 12, 3723–3739. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Baldwin, I.T. New insights into plant responses to the attack from insect herbivores. Annu. Rev. Genet. 2010, 44, 1–24. [Google Scholar] [CrossRef]
- Loake, G.; Grant, M. Salicylic acid in plant defence—the players and protagonists. Curr. Opin. Plant Biol. 2007, 10, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.-Q.; Jiang, H.-L.; Li, C.-Y. Systemin/jasmonate-mediated systemic defense signaling in tomato. Mol. Plant 2011, 4, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Kessler, A.; Baldwin, I.T. Plant responses to insect herbivory: The emerging molecular analysis. Annu. Rev. Plant Biol. 2002, 53, 299–328. [Google Scholar] [CrossRef] [PubMed]
- Boller, T.; Felix, G. A renaissance of elicitors: Perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Zhuo, K.; Chen, S.; Hu, L.; Sun, L.; Wang, X.; Zhang, L.H.; Liao, J. A novel nematode effector suppresses plant immunity by activating host reactive oxygen species-scavenging system. New Phytol. 2016, 209, 1159–1173. [Google Scholar] [CrossRef]
- Wang, Y.; Ju, D.; Yang, X.; Ma, D.; Wang, X. Comparative Transcriptome Analysis Between Resistant and Susceptible Rice Cultivars Responding to Striped Stem Borer (SSB), Chilo suppressalis (Walker) Infestation. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef]
- Hoseinzadeh, A.H.; Soorni, A.; Shoorooei, M.; Torkzadeh Mahani, M.; Maali Amiri, R.; Allahyari, H.; Mohammadi, R. Comparative transcriptome provides molecular insight into defense-associated mechanisms against spider mite in resistant and susceptible common bean cultivars. PLoS ONE 2020, 15, e0228680. [Google Scholar] [CrossRef]
- Cheboi, J.; Kimurto, P.; Kinyua, M.; Kiplagat, O.; Towett, B.; Kiptoo, J.; Kirui, S.; Kimno, S.; Gangarao, N. Evaluation of Selected Pigeonpea (Cajanus cajan (L.) Millsp.) Genotypes for Resistance to Insect Pest Complex in Dry Areas of North Rift Valley, Kenya. Am. J. Exp. Agric. 2016, 10, 9. [Google Scholar] [CrossRef]
- McCormack, E.; Tsai, Y.-C.; Braam, J. Handling calcium signaling: Arabidopsis CaMs and CMLs. Trends Plant Sci. 2005, 10, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Holopainen, J.K.; Blande, J.D. Where do herbivore-induced plant volatiles go? Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef]
- Diamond, J. Evolution, consequences and future of plant and animal domestication. Nature 2002, 418, 700. [Google Scholar] [CrossRef]
- Ranger, C.; Singh, A.; Johnson-Cicalese, J.; Polavarapu, S.; Vorsa, N. Intraspecific variation in aphid resistance and constitutive phenolics exhibited by the wild blueberry Vaccinium darrowi. J. Chem. Ecol. 2007, 33, 711. [Google Scholar] [CrossRef]
- Koenig, D.; Jiménez-Gómez, J.M.; Kimura, S.; Fulop, D.; Chitwood, D.H.; Headland, L.R.; Kumar, R.; Covington, M.F.; Devisetty, U.K.; Tat, A.V. Comparative transcriptomics reveals patterns of selection in domesticated and wild tomato. Proc. Natl. Acad. Sci. USA 2013, 110, E2655–E2662. [Google Scholar] [CrossRef]
- Bari, R.; Jones, J.D. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Koo, A.J.; Howe, G.A. The wound hormone jasmonate. Phytochemistry 2009, 70, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, O.; Piqueras, R.; Sánchez-Serrano, J.J.; Solano, R. ETHYLENE RESPONSE FACTOR1 integrates signals from ethylene and jasmonate pathways in plant defense. Plant Cell 2003, 15, 165–178. [Google Scholar] [CrossRef] [PubMed]
- McGrath, K.C.; Dombrecht, B.; Manners, J.M.; Schenk, P.M.; Edgar, C.I.; Maclean, D.J.; Scheible, W.-R.; Udvardi, M.K.; Kazan, K. Repressor-and activator-type ethylene response factors functioning in jasmonate signaling and disease resistance identified via a genome-wide screen of Arabidopsis transcription factor gene expression. Plant Physiol. 2005, 139, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Kiep, V.; Vadassery, J.; Lattke, J.; Maaß, J.P.; Boland, W.; Peiter, E.; Mithöfer, A.J.N.P. Systemic cytosolic Ca2+ elevation is activated upon wounding and herbivory in Arabidopsis. New Phytol. 2015, 207, 996–1004. [Google Scholar] [CrossRef]
- Kudla, J.; Batistič, O.; Hashimoto, K. Calcium signals: The lead currency of plant information processing. Plant Cell 2010, 22, 541–563. [Google Scholar] [CrossRef]
- Seybold, H.; Trempel, F.; Ranf, S.; Scheel, D.; Romeis, T.; Lee, J. Ca2+ signalling in plant immune response: From pattern recognition receptors to Ca2+ decoding mechanisms. New Phytol. 2014, 204, 782–790. [Google Scholar] [CrossRef]
- Vadassery, J.; Reichelt, M.; Hause, B.; Gershenzon, J.; Boland, W.; Mithöfer, A. CML42-mediated calcium signaling co-ordinates responses to Spodoptera herbivory and abiotic stresses in Arabidopsis. Plant Physiol. 2012, 159, 1159–1175. [Google Scholar] [CrossRef]
- Zhang, H.; Kjemtrup-Lovelace, S.; Li, C.; Luo, Y.; Chen, L.P.; Song, B.-H. Comparative RNA-Seq Analysis Uncovers a Complex Regulatory Network for Soybean Cyst Nematode Resistance in Wild Soybean (Glycine soja). Sci. Rep. 2017, 7, 9699. [Google Scholar] [CrossRef]
- Ranf, S.; Eschen-Lippold, L.; Pecher, P.; Lee, J.; Scheel, D. Interplay between calcium signalling and early signalling elements during defence responses to microbe-or damage-associated molecular patterns. Plant J. 2011, 68, 100–113. [Google Scholar] [CrossRef]
- Bowles, D.J. Defense-related proteins in higher plants. Annu. Rev. Biochem. 1990, 59, 873–907. [Google Scholar] [CrossRef]
- Dao, T.T.H.; Linthorst, H.J.M.; Verpoorte, R. Chalcone synthase and its functions in plant resistance. Phytochem. Rev. 2011, 10, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.E.; Rashotte, A.M.; Murphy, A.S.; Normanly, J.; Tague, B.W.; Peer, W.A.; Taiz, L.; Muday, G.K. Flavonoids act as negative regulators of auxin transport in vivo in Arabidopsis. Plant Physiol. 2001, 126, 524–535. [Google Scholar] [CrossRef] [PubMed]
- Davis, D. The use of intergeneric grafts to demonstrate toxins in the Fusarium wilt disease of tomato. Am. J. Bot. 1954, 395–398. [Google Scholar] [CrossRef]
- Major, I.T.; Constabel, C.P. Functional Analysis of the Kunitz Trypsin Inhibitor Family in Poplar Reveals Biochemical Diversity and Multiplicity in Defense against Herbivores. Plant Physiol. 2008, 146, 888. [Google Scholar] [CrossRef] [PubMed]
- Aharoni, A.; Giri, A.P.; Deuerlein, S.; Griepink, F.; de Kogel, W.-J.; Verstappen, F.W.; Verhoeven, H.A.; Jongsma, M.A.; Schwab, W.; Bouwmeester, H.J. Terpenoid metabolism in wild-type and transgenic Arabidopsis plants. Plant Cell 2003, 15, 2866–2884. [Google Scholar] [CrossRef] [PubMed]
- Kappers, I.F.; Aharoni, A.; Van Herpen, T.W.; Luckerhoff, L.L.; Dicke, M.; Bouwmeester, H.J. Genetic engineering of terpenoid metabolism attracts bodyguards to Arabidopsis. Science 2005, 309, 2070–2072. [Google Scholar] [CrossRef]
- Schweizer, F.; Bodenhausen, N.; Lassueur, S.; Masclaux, F.G.; Reymond, P. Differential contribution of transcription factors to Arabidopsis thaliana defense against Spodoptera littoralis. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef]
- Bencke-Malato, M.; Cabreira, C.; Wiebke-Strohm, B.; Bücker-Neto, L.; Mancini, E.; Osorio, M.B.; Homrich, M.S.; Turchetto-Zolet, A.C.; De Carvalho, M.C.; Stolf, R. Genome-wide annotation of the soybean WRKY family and functional characterization of genes involved in response to Phakopsora pachyrhiziinfection. BMC Plant Biol. 2014, 14, 236. [Google Scholar] [CrossRef]
- Skibbe, M.; Qu, N.; Galis, I.; Baldwin, I.T. Induced Plant Defenses in the Natural Environment: Nicotiana attenuata WRKY3 and WRKY6 Coordinate Responses to Herbivory. Plant Cell 2008, 20, 1984. [Google Scholar] [CrossRef]
- Fernández-Calvo, P.; Chini, A.; Fernández-Barbero, G.; Chico, J.-M.; Gimenez-Ibanez, S.; Geerinck, J.; Eeckhout, D.; Schweizer, F.; Godoy, M.; Franco-Zorrilla, J.M. The Arabidopsis bHLH transcription factors MYC3 and MYC4 are targets of JAZ repressors and act additively with MYC2 in the activation of jasmonate responses. Plant Cell 2011, 23, 701–715. [Google Scholar] [CrossRef]
- Nuruzzaman, M.; Sharoni, A.M.; Kikuchi, S. Roles of NAC transcription factors in the regulation of biotic and abiotic stress responses in plants. Front. Microbiol. 2013, 4, 248. [Google Scholar] [CrossRef]
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.; Giorgi, F. Trimmomatic: A flexible read trimming tool for illumina NGS data. Bioinformatics 2014, 25, 402–408. [Google Scholar]
- Varshney, R.K.; Chen, W.; Li, Y.; Bharti, A.K.; Saxena, R.K.; Schlueter, J.A.; Donoghue, M.T.; Azam, S.; Fan, G.; Whaley, A.M. Draft genome sequence of pigeonpea (Cajanus cajan), an orphan legume crop of resource-poor farmers. Nat. Biotechnol. 2012, 30, 83. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Götz, S. Blast2GO: A Comprehensive Suite for Functional Analysis in Plant Genomics. Int. J. Plant Genom. 2008, 2008, 619832. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Susceptible Cultivated (CT) | Tolerant Wild (WT) | |||||
|---|---|---|---|---|---|---|
| Accession | Log2FC | p-Value | Log2FC | p-Value | Description | Pathway |
| XM_020361379.1 | 3.31 | 4.12 × 10−7 | 3.86 | 3.81 × 10−11 | calcium uniporter protein 2 | Ca2+ signalling |
| XM_020383212.1 | −5.45 | 1.35 × 10−4 | 2.26 | 2.72 × 10−3 | calmodulin receptor-like kinase | Ca2+ signalling |
| XM_020371759.2 | 1.96 | 1.00 × 10−5 | 6.92 | 8.29 × 10−18 | CBL-interacting serine/threonine-protein kinase 21 | Ca2+ signalling |
| XM_020370652.1 | 2.47 | 6.74 × 10−8 | 4.02 | 3.08 × 10−13 | 12-oxophytodienoate reductase | Ethylene |
| XM_020349666.1 | 8.11 | 3.13 × 10−18 | 9.30 | 3.74 × 10−25 | ABR1-like | Ethylene |
| XM_020353227.2 | 2.29 | 3.76 × 10−9 | 3.81 | 1.26 × 10−10 | 1-aminocyclopropane-1-carboxylate oxidase | Ethylene |
| XM_020357438.2 | 3.34 | 4.21 × 10−4 | 5.24 | 0.000485 | zeatin O-glucosyltransferase-like | Cytokinin |
| XM_020370529.1 | −3.56 | 2.44 × 10−9 | 6.86 | 5.86 × 10−9 | chalcone synthase 1 | Flavonoid |
| XM_020357544.1 | 6.88 | 1.25 × 10−9 | 10.90 | 5.89 × 10−36 | kunitz trypsin inhibitor | Inhibitors |
| XM_020364128.1 | 3.22 | 3.17 × 10−7 | 7.18 | 1.21 × 10−10 | wound-induced trypsin inhibitor | Inhibitors |
| XM_020376230.1 | −1.27 | 3.23 × 10−3 | 6.19 | 3.60 × 10−6 | 2-hydroxyisoflavanone synthase | Isoflavonoid |
| XM_020384234.1 | 5.57 | 1.11 × 10−4 | 8.95 | 8.29 × 10−18 | isoflavone-7-O-methyltransferase | Isoflavonoid |
| XM_020377202.1 | −2.60 | 3.34 × 10−5 | 10.14 | 3.38 × 10−37 | 2-oxoglutarate-dependent dioxygenase | Jasmonic acid |
| XM_020350219.1 | 2.76 | 5.51 × 10−10 | 4.52 | 2.48 × 10−9 | allene oxide synthase | Jasmonic acid |
| XM_020355903.1 | −2.12 | 3.28 × 10−7 | 7.13 | 6.50 × 10−19 | linoleate 9S-lipoxygenase | Jasmonic acid |
| XM_020365833.2 | 2.61 | 4.25 × 10−4 | 3.89 | 0.000155 | jasmonate O-methyltransferase | Jasmonic acid |
| XM_020383219.1 | 2.68 | 2.16 × 10−4 | 10.12 | 1.02 × 10−29 | nerolidol synthase 1 | Terpene |
| XM_020353681.1 | −4.46 | 2.51 × 10−6 | 8.71 | 5.98 × 10−14 | terpene synthase 2 | Terpene |
| XM_020383551.1 | 5.62 | 5.20 × 10−11 | 5.59 | 1.75 × 10−4 | bHLH18 | Transcription factor |
| XM_020382733.1 | −1.79 | 1.97 × 10−3 | 5.47 | 1.37 × 10−4 | bZIP11 | Transcription factor |
| XM_020383974.1 | 4.52 | 4.92 × 10−9 | 5.72 | 5.05 × 10−5 | ERF RAP2 | Transcription factor |
| XM_020371421.1 | −2.72 | 9.95 × 10−4 | 5.70 | 5.97 × 10−4 | MYB4 | Transcription factor |
| XM_020351385.1 | 1.13 | 1.51 × 10−3 | 5.72 | 7.70 × 10−11 | MYC4 | Transcription factor |
| XM_020349871.1 | 4.36 | 4.93 × 10−3 | 2.59 | 1.47 × 10−9 | MYC2 | Transcription factor |
| XM_020376674.1 | 3.88 | 3.03 × 10−4 | 2.57 | 8.38 × 10−4 | WRKY 41 | Transcription factor |
| XM_020374599.1 | 2.06 | 1.89 × 10−4 | 3.00 | 6.17 × 10−5 | WRKY 51 | Transcription factor |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Njaci, I.; Ngugi-Dawit, A.; Oduor, R.O.; Kago, L.; Williams, B.; Hoang, L.T.M.; Mundree, S.G.; Ghimire, S.R. Comparative Analysis Delineates the Transcriptional Resistance Mechanisms for Pod Borer Resistance in the Pigeonpea Wild Relative Cajanus scarabaeoides (L.) Thouars. Int. J. Mol. Sci. 2021, 22, 309. https://doi.org/10.3390/ijms22010309
Njaci I, Ngugi-Dawit A, Oduor RO, Kago L, Williams B, Hoang LTM, Mundree SG, Ghimire SR. Comparative Analysis Delineates the Transcriptional Resistance Mechanisms for Pod Borer Resistance in the Pigeonpea Wild Relative Cajanus scarabaeoides (L.) Thouars. International Journal of Molecular Sciences. 2021; 22(1):309. https://doi.org/10.3390/ijms22010309
Chicago/Turabian StyleNjaci, Isaac, Abigail Ngugi-Dawit, Richard O. Oduor, Leah Kago, Brett Williams, Linh Thi My Hoang, Sagadevan G. Mundree, and Sita R. Ghimire. 2021. "Comparative Analysis Delineates the Transcriptional Resistance Mechanisms for Pod Borer Resistance in the Pigeonpea Wild Relative Cajanus scarabaeoides (L.) Thouars" International Journal of Molecular Sciences 22, no. 1: 309. https://doi.org/10.3390/ijms22010309
APA StyleNjaci, I., Ngugi-Dawit, A., Oduor, R. O., Kago, L., Williams, B., Hoang, L. T. M., Mundree, S. G., & Ghimire, S. R. (2021). Comparative Analysis Delineates the Transcriptional Resistance Mechanisms for Pod Borer Resistance in the Pigeonpea Wild Relative Cajanus scarabaeoides (L.) Thouars. International Journal of Molecular Sciences, 22(1), 309. https://doi.org/10.3390/ijms22010309