PASylated Thymosin α1: A Long-Acting Immunostimulatory Peptide for Applications in Oncology and Virology



Abstract
1. Introduction
2. Results
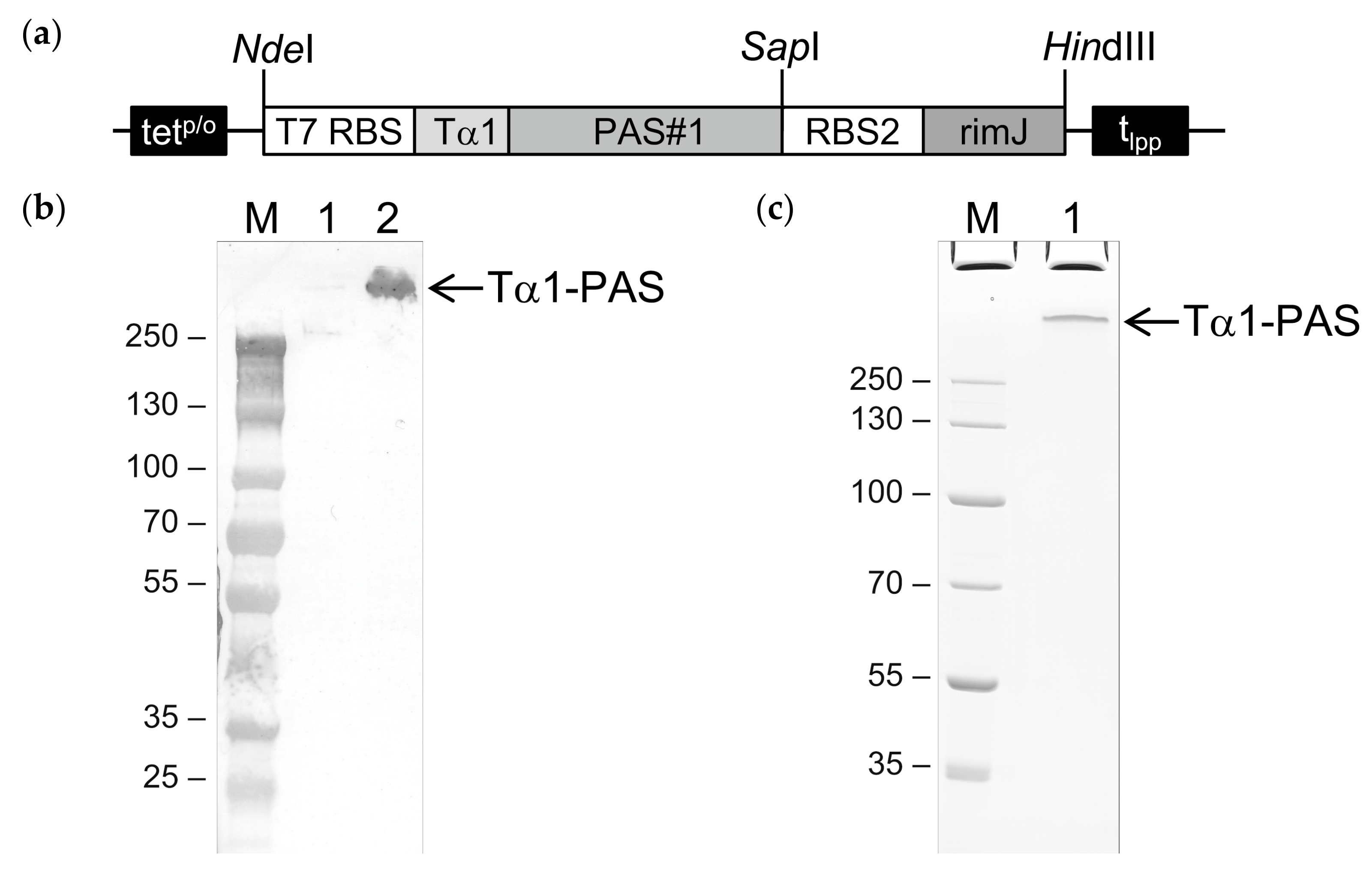
2.1. Cloning and Bacterial Production of PASylated Tα1
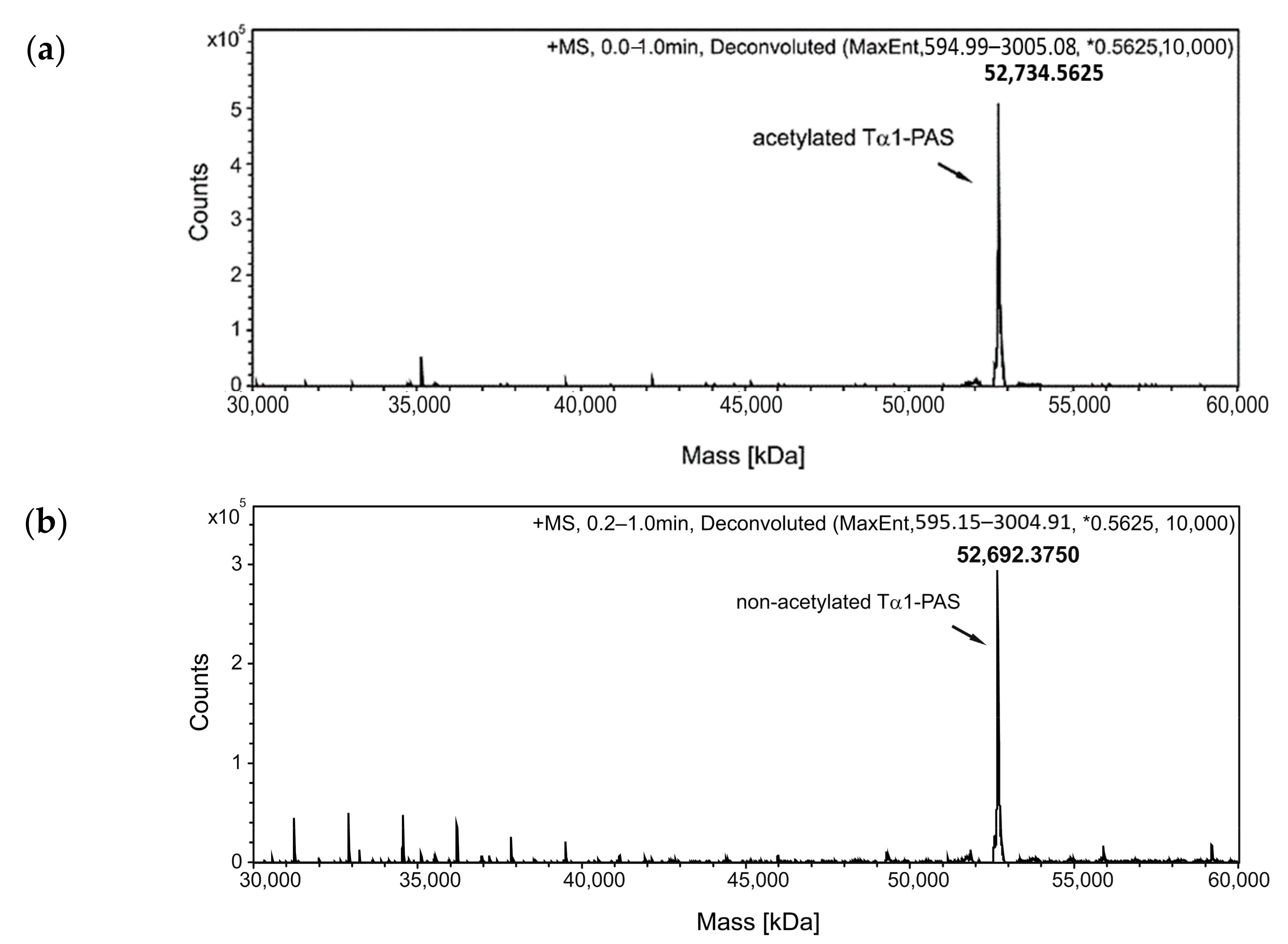
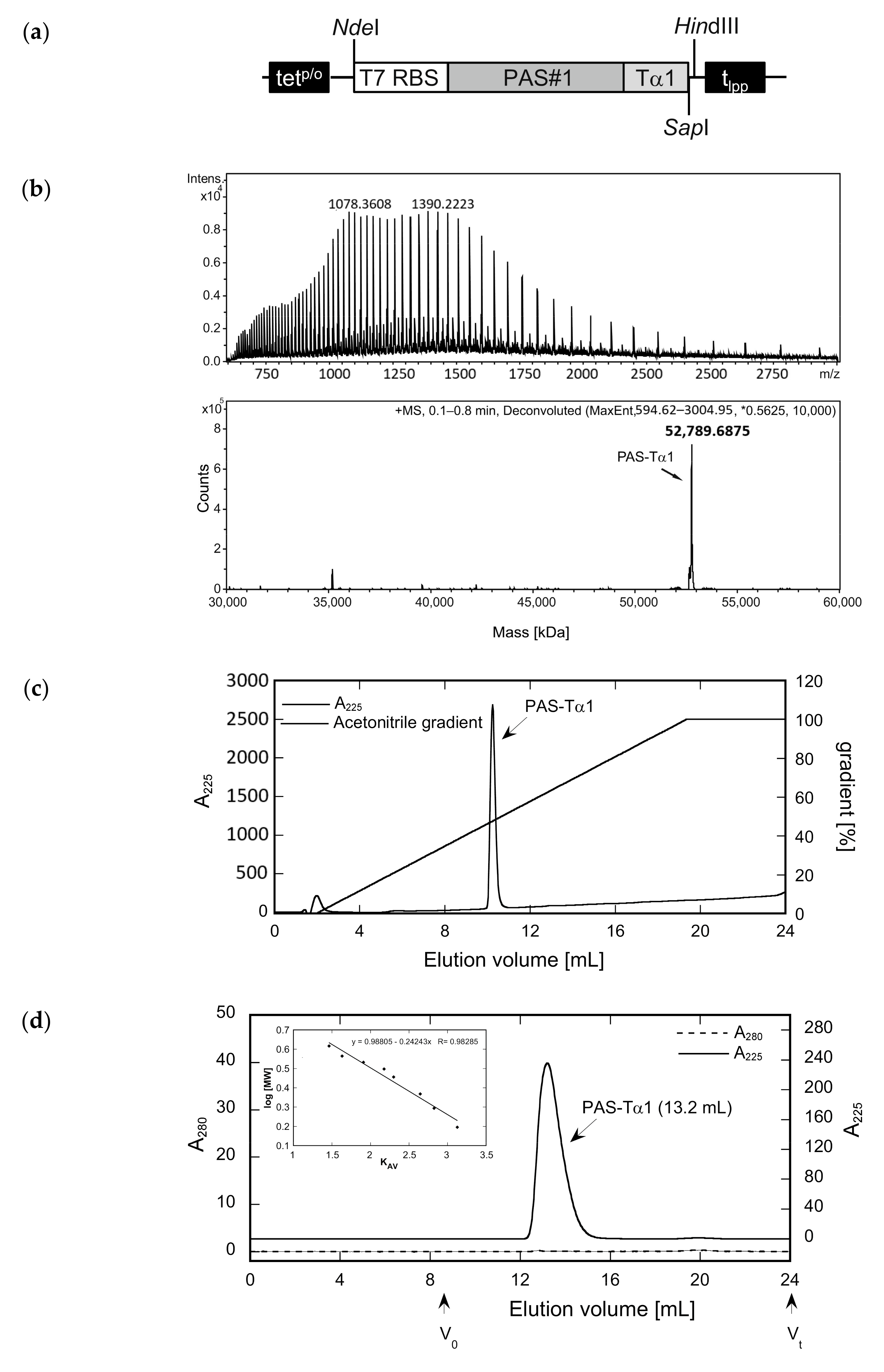
2.2. Purification and In Vitro Characterization of PASylated Tα1
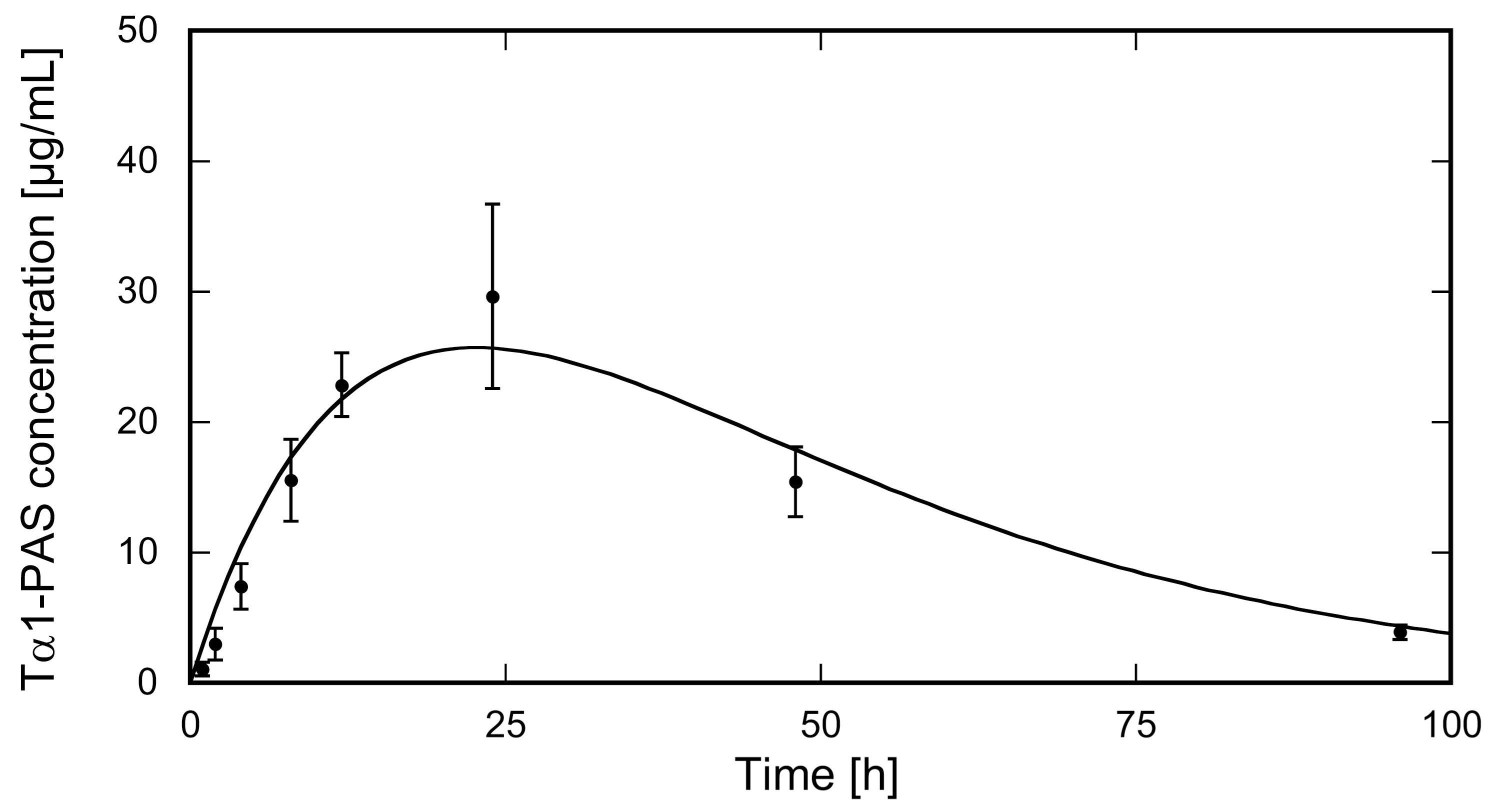
2.3. PASylation Strongly Prolongs Tα1 Pharmacokinetics in Rats
3. Discussion
4. Materials and Methods
4.1. Construction of the Expression Plasmid
4.2. Bacterial Production of Recombinant PASylated Tα1
4.3. Purification of PASylated Tα1
4.4. Analytical Size Exclusion Chromatography
4.5. Endotoxin Quantification
4.6. Western Blot Analysis
4.7. Reverse-Phase Chromatography (RPC) and ESI Mass Spectrometry
4.8. Pharmacokinetic Analysis in Rats
4.9. Quantification of Purified Tα1-PAS
4.10. ELISA Quantification of PASylated Tα1
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AEX | Anion exchange |
| AP | Alkaline phosphatase |
| aTc | Anhydrotetracycline hydrochloride |
| AUC | Area und the curve |
| BCIP | 5-Bromo-4-chloro-3-indoxyl phosphate |
| BSA | Bovine serum albumin |
| CEX | Cation exchange |
| CL | Clearance |
| Cmax | Maximum (or peak) serum concentration |
| CNP | C-type natriuretic peptide |
| CORYNEX | Corynebacterium glutamicum protein expression system |
| COVID-19 | Severe acute respiratory syndrome coronavirus 2 |
| CTLA-4 | Cytotoxic T-lymphocyte-associated protein 4 |
| CV | Column volume |
| DCs | Dendritic cells |
| DPP-IV | Dipeptidyl peptidase-4 |
| ELISA | Enzyme-linked immunosorbent assay |
| ESETEC | E. coli secretion technology |
| ESI-MS | Electrospray ionization mass spectrometry |
| FPLC | Fast protein liquid chromatography |
| GCC | Glucagon |
| GIP | Glucose-dependent insulinotropic polypeptide |
| GLP-1 | Glucagon-like peptide 1 |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| GSH | γ-L-Glutamyl-L-cysteinyl-glycine |
| HBV | Hepatitis B virus |
| HCV | Hepatitis C virus |
| hGH | Human growth hormone |
| hIFN-α | Human interferon 1 |
| HIV-1 | Human immunodeficiency virus I |
| iRGD | Internalizing arginylglycylaspartic acid peptide |
| K3-EDTA | Tri-potassium ethylenediaminetetraacetic acid |
| MHC | Major histocompatibility complex |
| MOPS | 3-(N-morpholino)propanesulfonic acid |
| NBT | Nitro blue tetrazolium |
| OD | Optical density at 550 nm |
| PAS | Proline/alanine-rich sequence |
| PAS-Tα1 | N-terminally PASylated Tα1 |
| PBS | Phosphate-buffered saline |
| PD | Pharmacodynamics |
| PEG | Polyethylene glycol |
| p.i. | Post injection |
| PK | Pharmacokinetics |
| ProTα | Prothymosin alpha |
| PTH | Parathyroid hormone |
| Q-TOF | Quadrupole time of flight |
| RBS | Ribosome-binding site |
| RGD | Arginylglycylaspartic acid |
| RimJ | Ribosomal-protein-S5-alanine N-acetyltransferase |
| RPC | Reverse phase chromatography |
| RT | Room temperature |
| SEC | Size exclusion chromatography |
| SDS-PAGE | Sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| τ1/2α | Half-life of distribution |
| τ1/2β | Elimination half-life |
| Tα1 | Thymosin alpha 1 |
| Tα1-PAS | C-terminally PASylated Thymosin alpha 1 |
| tetp/o | Tetracycline promoter/operator |
| Th | T helper cell |
| tlpp | Lipoprotein terminator |
| TLR-2 | Toll-like receptor 1 |
| TLR-9 | Toll-like receptor 2 |
| TMAE | Trimethylaminoethyl |
| tmax | Time to reach Cmax |
References
- Goldstein, A.L.; Low, T.L.; McAdoo, M.; McClure, J.; Thurman, G.B.; Rossio, J.; Lai, C.Y.; Chang, D.; Wang, S.S.; Harvey, C.; et al. Thymosin alpha1: Isolation and sequence analysis of an immunologically active thymic polypeptide. Proc. Natl. Acad. Sci. USA 1977, 74, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Hoch, K.; Volk, D.E. Structures of thymosin proteins. Vitam. Horm. 2016, 102, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Gast, K.; Damaschun, H.; Eckert, K.; Schulze-Forster, K.; Maurer, H.R.; Muller-Frohne, M.; Zirwer, D.; Czarnecki, J.; Damaschun, G. Prothymosin alpha: A biologically active protein with random coil conformation. Biochemistry 1995, 34, 13211–13218. [Google Scholar] [CrossRef] [PubMed]
- Sarandeses, C.S.; Covelo, G.; Diaz-Jullien, C.; Freire, M. Prothymosin alpha is processed to thymosin alpha 1 and thymosin alpha 11 by a lysosomal asparaginyl endopeptidase. J. Biol. Chem. 2003, 278, 13286–13293. [Google Scholar] [CrossRef]
- Camerini, R.; Garaci, E. Historical review of thymosin alpha 1 in infectious diseases. Expert Opin. Biol. Ther. 2015, 15 (Suppl. S1), S117–S127. [Google Scholar] [CrossRef] [PubMed]
- Romani, L.; Bistoni, F.; Gaziano, R.; Bozza, S.; Montagnoli, C.; Perruccio, K.; Pitzurra, L.; Bellocchio, S.; Velardi, A.; Rasi, G.; et al. Thymosin alpha 1 activates dendritic cells for antifungal Th1 resistance through toll-like receptor signaling. Blood 2004, 103, 4232–4239. [Google Scholar] [CrossRef]
- King, R.; Tuthill, C. Immune modulation with thymosin alpha 1 treatment. Vitam. Horm. 2016, 102, 151–178. [Google Scholar] [CrossRef]
- Baumann, C.A.; Badamchian, M.; Goldstein, A.L. Thymosin alpha 1 antagonizes dexamethasone and CD3-induced apoptosis of CD4+ CD8+ thymocytes through the activation of cAMP and protein kinase C dependent second messenger pathways. Mech. Ageing Dev. 1997, 94, 85–101. [Google Scholar] [CrossRef]
- Giuliani, C.; Napolitano, G.; Mastino, A.; Di Vincenzo, S.; D’Agostini, C.; Grelli, S.; Bucci, I.; Singer, D.S.; Kohn, L.D.; Monaco, F.; et al. Thymosin-alpha1 regulates MHC class I expression in FRTL-5 cells at transcriptional level. Eur. J. Immunol. 2000, 30, 778–786. [Google Scholar] [CrossRef]
- Garaci, E.; Pica, F.; Serafino, A.; Balestrieri, E.; Matteucci, C.; Moroni, G.; Sorrentino, R.; Zonfrillo, M.; Pierimarchi, P.; Sinibaldi-Vallebona, P. Thymosin α1 and cancer: Action on immune effector and tumor target cells. Ann. N. Y. Acad. Sci. 2012, 1269, 26–33. [Google Scholar] [CrossRef]
- Palamara, A.; Bue, M.; Savini, P. Thymosin alpha 1 inhibits Sendai virus replication: Involvement of intracellular redox state. In Proceedings of the 6th International Expert Forum of Immunotherapy and Gene Therapy, Florence, Italy, 6–8 May 1998. [Google Scholar]
- Moody, T.W.; Fagarasan, M.; Zia, F.; Cesnjaj, M.; Goldstein, A.L. Thymosin alpha 1 down-regulates the growth of human non-small cell lung cancer cells in vitro and in vivo. Cancer Res. 1993, 53, 5214–5218. [Google Scholar] [PubMed]
- Sgarbanti, R.; Nencioni, L.; Amatore, D.; Coluccio, P.; Fraternale, A.; Sale, P.; Mammola, C.L.; Carpino, G.; Gaudio, E.; Magnani, M.; et al. Redox regulation of the influenza hemagglutinin maturation process: A new cell-mediated strategy for anti-influenza therapy. Antioxid. Redox Signal. 2011, 15, 593–606. [Google Scholar] [CrossRef] [PubMed]
- Tuthill, C.W.; King, R.S. Thymosin alpha 1—A peptide immune modulator with a broad range of clinical applications. Clin. Exp. Pharmacol. 2013, 3, 1000133. [Google Scholar]
- Costantini, C.; Bellet, M.M.; Pariano, M.; Renga, G.; Stincardini, C.; Goldstein, A.L.; Garaci, E.; Romani, L. A reappraisal of thymosin alpha1 in cancer therapy. Front. Oncol. 2019, 9, 873. [Google Scholar] [CrossRef] [PubMed]
- Danielli, R.; Cisternino, F.; Giannarelli, D.; Calabro, L.; Camerini, R.; Savelli, V.; Bova, G.; Dragonetti, R.; Di Giacomo, A.M.; Altomonte, M.; et al. Long-term follow up of metastatic melanoma patients treated with Thymosin alpha-1: Investigating immune checkpoints synergy. Expert Opin. Biol. Ther. 2018, 18, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Renga, G.; Bellet, M.M.; Pariano, M.; Gargaro, M.; Stincardini, C.; D’Onofrio, F.; Mosci, P.; Brancorsini, S.; Bartoli, A.; Goldstein, A.L.; et al. Thymosin α1 protects from CTLA-4 intestinal immunopathology. Life Sci. Alliance 2020, 3, e202000662. [Google Scholar] [CrossRef]
- Romani, L.; Oikonomou, V.; Moretti, S.; Iannitti, R.G.; D’Adamo, M.C.; Villella, V.R.; Pariano, M.; Sforna, L.; Borghi, M.; Bellet, M.M.; et al. Thymosin alpha1 represents a potential potent single-molecule-based therapy for cystic fibrosis. Nat. Med. 2017, 23, 590–600. [Google Scholar] [CrossRef]
- Severa, M.; Zhang, J.; Giacomini, E.; Rizzo, F.; Etna, M.P.; Cruciani, M.; Garaci, E.; Chopp, M.; Coccia, E.M. Thymosins in multiple sclerosis and its experimental models: Moving from basic to clinical application. Mult. Scler. Relat. Disord. 2019, 27, 52–60. [Google Scholar] [CrossRef]
- Pei, F.; Guan, X.; Wu, J. Thymosin alpha 1 treatment for patients with sepsis. Expert Opin. Biol. Ther. 2018, 18, 71–76. [Google Scholar] [CrossRef]
- Lin, H.Y. The severe COVID-19: A sepsis induced by viral infection? And its immunomodulatory therapy. Chin. J. Traumatol. 2020, 23, 190–195. [Google Scholar] [CrossRef]
- Meng, Z.; Wang, T.; Li, C.; Chen, X.; Li, L.; Qin, X.; Li, H.; Luo, J. An experimental trial of recombinant human interferon alpha nasal drops to prevent coronavirus disease 2019 in medical staff in an epidemic area. medRxiv 2020. [Google Scholar] [CrossRef]
- Liu, Y.; Pang, Y.; Hu, Z.; Wu, M.; Wang, C.; Feng, Z.; Mao, C.; Tan, Y.; Chen, L.; Li, M.; et al. Thymosin alpha 1 (Talpha1) reduces the mortality of severe COVID-19 by restoration of lymphocytopenia and reversion of exhausted T cells. Clin. Infect. Dis. 2020, 71, 2150–2157. [Google Scholar] [CrossRef] [PubMed]
- Toniolo, C.; Bonora, G.M.; Heimer, E.P.; Felix, A.M. Structure, solubility and reactivity of peptides. A conformational study of two protected key intermediates from a large-scale synthesis of thymosin alpha 1. Int. J. Pept. Protein Res. 1987, 30, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Toplak, A.; Rozeboom, H.J.; Wijma, H.J.; Quaedflieg, P.; van Maarseveen, J.H.; Janssen, D.B.; Nuijens, T. Design of a substrate-tailored peptiligase variant for the efficient synthesis of thymosin-alpha1. Org. Biomol. Chem. 2018, 16, 609–618. [Google Scholar] [CrossRef]
- Liu, B.; Gong, X.; Chang, S.; Sun, P.; Wu, J. Generation of mature Nalpha-terminal acetylated thymosin alpha 1 by cleavage of recombinant prothymosin alpha. Sci. World J. 2013, 2013, 387282. [Google Scholar] [CrossRef]
- Zhou, L.; Lai, Z.T.; Lu, M.K.; Gong, X.G.; Xie, Y. Expression and hydroxylamine cleavage of thymosin alpha 1 concatemer. J. Biomed. Biotechnol. 2008, 2008, 736060. [Google Scholar] [CrossRef]
- Ren, Y.; Yao, X.; Dai, H.; Li, S.; Fang, H.; Chen, H.; Zhou, C. Production of Nalpha-acetylated thymosin alpha1 in Escherichia coli. Microb. Cell Fact. 2011, 10, 26. [Google Scholar] [CrossRef]
- Chen, P.F.; Zhang, H.Y.; Fu, G.F.; Xu, G.X.; Hou, Y.Y. Overexpression of soluble human thymosin alpha 1 in Escherichia coli. Acta Biochim. Biophys. Sin. 2005, 37, 147–151. [Google Scholar] [CrossRef]
- Rost, K.L.; Wierich, W.; Masayuki, F.; Tuthill, C.W.; Horwitz, D.L.; Herrmann, W.M. Pharmacokinetics of thymosin alpha1 after subcutaneous injection of three different formulations in healthy volunteers. Int. J. Clin. Pharmacol. Ther. 1999, 37, 51–57. [Google Scholar]
- Schlapschy, M.; Binder, U.; Börger, C.; Theobald, I.; Wachinger, K.; Kisling, S.; Haller, D.; Skerra, A. PASylation: A biological alternative to PEGylation for extending the plasma half-life of pharmaceutically active proteins. Protein Eng. Des. Sel. 2013, 26, 489–501. [Google Scholar] [CrossRef]
- Fang, H.; Zhang, X.; Shen, L.; Si, X.; Ren, Y.; Dai, H.; Li, S.; Zhou, C.; Chen, H. RimJ is responsible for N(alpha)-acetylation of thymosin alpha1 in Escherichia coli. Appl. Microbiol. Biotechnol. 2009, 84, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Breibeck, J.; Skerra, A. The polypeptide biophysics of proline/alanine-rich sequences (PAS): Recombinant biopolymers with PEG-like properties. Biopolymers 2018, 109, e23069. [Google Scholar] [CrossRef] [PubMed]
- Binder, U.; Skerra, A. PASylation®: A versatile technology to extend drug delivery. Curr. Opin. Colloid Int. 2017, 31, 10–17. [Google Scholar] [CrossRef]
- Gebauer, M.; Skerra, A. Prospects of PASylation@ for the design of protein and peptide therapeutics with extended half-life and enhanced action. Bioorg. Med. Chem. 2018, 26, 2882–2887. [Google Scholar] [CrossRef] [PubMed]
- Skerra, A. Use of the tetracycline promoter for the tightly regulated production of a murine antibody fragment in Escherichia coli. Gene 1994, 151, 131–135. [Google Scholar] [CrossRef]
- Morath, V.; Bolze, F.; Schlapschy, M.; Schneider, S.; Sedlmayer, F.; Seyfarth, K.; Klingenspor, M.; Skerra, A. PASylation of murine leptin leads to extended plasma half-life and enhanced in vivo efficacy. Mol. Pharm. 2015, 12, 1431–1442. [Google Scholar] [CrossRef] [PubMed]
- Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook, 1st ed.; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar] [CrossRef]
- Xiao, Q.; Zhang, F.; Nacev, B.A.; Liu, J.O.; Pei, D. Protein N-terminal processing: Substrate specificity of Escherichia coli and human methionine aminopeptidases. Biochemistry 2010, 49, 5588–5599. [Google Scholar] [CrossRef]
- Garrett, E.R. The Bateman function revisited: A critical reevaluation of the quantitative expressions to characterize concentrations in the one compartment body model as a function of time with first-order invasion and first-order elimination. J. Pharmacokinet. Biopharm. 1994, 22, 103–128. [Google Scholar] [CrossRef]
- Wang, F.; Yu, T.; Zheng, H.; Lao, X. Thymosin alpha1-Fc modulates the immune system and down-regulates the progression of melanoma and breast cancer with a prolonged half-life. Sci. Rep. 2018, 8, 12351. [Google Scholar] [CrossRef]
- Wegmüller, S.; Schmid, S. Recombinant peptide production in microbial cells. Curr. Org. Chem. 2014, 18, 1005–2019. [Google Scholar] [CrossRef]
- Goeddel, D.V.; Kleid, D.G.; Bolivar, F.; Heyneker, H.L.; Yansura, D.G.; Crea, R.; Hirose, T.; Kraszewski, A.; Itakura, K.; Riggs, A.D. Expression in Escherichia coli of chemically synthesized genes for human insulin. Proc. Natl. Acad. Sci. USA 1979, 76, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Li, Q.; Wang, F.; Bao, J.; Dai, M.; Zheng, H.; Lao, X. Generation of a novel long-acting thymosin alpha1-Fc fusion protein and its efficacy for the inhibition of breast cancer in vivo. Biomed. Pharmacother. 2018, 108, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Mücke, M.O.R.; Leonhartsberger, S. E. coli secretion technologies enable production of high yields of active human antibody fragments. BioProcess Int. 2009, 7, 40–47. [Google Scholar]
- Kikuchi, Y.; Date, M.; Yokoyama, K.; Umezawa, Y.; Matsui, H. Secretion of active-form Streptoverticillium mobaraense transglutaminase by Corynebacterium glutamicum: Processing of the pro-transglutaminase by a cosecreted subtilisin-Like protease from Streptomyces albogriseolus. Appl. Environ. Microbiol. 2003, 69, 358–366. [Google Scholar] [CrossRef]
- Matsuda, Y.; Itaya, H.; Kitahara, Y.; Theresia, N.M.; Kutukova, E.A.; Yomantas, Y.A.; Date, M.; Kikuchi, Y.; Wachi, M. Double mutation of cell wall proteins CspB and PBP1a increases secretion of the antibody Fab fragment from Corynebacterium glutamicum. Microb. Cell. Fact. 2014, 13, 56. [Google Scholar] [CrossRef]
- Di Cesare, S.; Binder, U.; Maier, T.; Skerra, A. High-yield production of PASylated human growth hormone using secretory E. coli technology. BioProcess Int. 2013, 11, 30–38. [Google Scholar]
- Chen, J.; Li, H.; Wang, T.; Sun, S.; Liu, J. Production of N(alpha)-acetyl Talpha1-HSA through in vitro acetylation by RimJ. Oncotarget 2017, 8, 95247–95255. [Google Scholar] [CrossRef]
- Lao, X.; Liu, M.; Chen, J.; Zheng, H. A tumor-penetrating peptide modification enhances the antitumor activity of thymosin alpha 1. PLoS ONE 2013, 8, e72242. [Google Scholar] [CrossRef]
- Gao, D.; Zhang, X.; Zhang, J.; Cao, J.; Wang, F. Expression of thymosin alpha1-thymopentin fusion peptide in Pichia pastoris and its characterization. Arch. Pharm. Res. 2008, 31, 1471–1476. [Google Scholar] [CrossRef]
- Moretti, S.; Oikonomou, V.; Garaci, E.; Romani, L. Thymosin alpha1: Burying secrets in the thymus. Expert Opin. Biol. Ther. 2015, 15 (Suppl. S1), S51–S58. [Google Scholar] [CrossRef][Green Version]
- Kubetzko, S.; Sarkar, C.A.; Plückthun, A. Protein PEGylation decreases observed target association rates via a dual blocking mechanism. Mol. Pharmacol. 2005, 68, 1439–1454. [Google Scholar] [CrossRef] [PubMed]
- Powers, N.E.; Swartzwelter, B.; Marchetti, C.; de Graaf, D.M.; Lerchner, A.; Schlapschy, M.; Datar, R.; Binder, U.; Edwards, C.K., 3rd; Skerra, A.; et al. PASylation of IL-1 receptor antagonist (IL-1Ra) retains IL-1 blockade and extends its duration in mouse urate crystal-induced peritonitis. J. Biol. Chem. 2020, 295, 868–882. [Google Scholar] [CrossRef] [PubMed]
- Peng, G.; Pan, X.; Hu, H.; Xu, Y.; Wu, C. N-terminal site-specific PEGylation enhances the circulation half-life of Thymosin alpha 1. J. Drug Deliv. Sci. Technol. 2019, 49, 405–412. [Google Scholar] [CrossRef]
- Caldwell, G.W.; Masucci, J.A.; Yan, Z.; Hageman, W. Allometric scaling of pharmacokinetic parameters in drug discovery: Can human CL, Vss and t1/2 be predicted from in-vivo rat data? Eur. J. Drug Metab. Pharmacokinet. 2004, 29, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, I. Interspecies Pharmacokinetic Scaling: Principles and Application of Allometric Scaling, 1st ed.; Pine House Publishers: Rockville, MD, USA, 2005. [Google Scholar]
- Matteucci, C.; Grelli, S.; Balestrieri, E.; Minutolo, A.; Argaw-Denboba, A.; Macchi, B.; Sinibaldi-Vallebona, P.; Perno, C.F.; Mastino, A.; Garaci, E. Thymosin alpha 1 and HIV-1: Recent advances and future perspectives. Future Microbiol. 2017, 12, 141–155. [Google Scholar] [CrossRef]
- King, R.S.; Tuthill, C. Evaluation of thymosin alpha 1 in nonclinical models of the immune-suppressing indications melanoma and sepsis. Expert Opin. Biol. Ther. 2015, 15 (Suppl. S1), S41–S49. [Google Scholar] [CrossRef]
- Garaci, E.; Pica, F.; Matteucci, C.; Gaziano, R.; D’Agostini, C.; Miele, M.T.; Camerini, R.; Palamara, A.T.; Favalli, C.; Mastino, A.; et al. Historical review on thymosin alpha1 in oncology: Preclinical and clinical experiences. Expert Opin. Biol. Ther. 2015, 15 (Suppl. S1), S31–S39. [Google Scholar] [CrossRef]
- Fosgerau, K.; Hoffmann, T. Peptide therapeutics: Current status and future directions. Drug Discov. Today 2015, 20, 122–128. [Google Scholar] [CrossRef]
- Pipes, G.T.; Yang, J. Cardioprotection by thymosin beta 4. Vitam. Horm. 2016, 102, 209–226. [Google Scholar] [CrossRef]
- Sosne, G. Thymosin beta 4 and the eye: The journey from bench to bedside. Expert Opin. Biol. Ther. 2018, 18, 99–104. [Google Scholar] [CrossRef]
- Lumsden, N.G.; Khambata, R.S.; Hobbs, A.J. C-type natriuretic peptide (CNP): Cardiovascular roles and potential as a therapeutic target. Curr. Pharm. Des. 2010, 16, 4080–4088. [Google Scholar] [CrossRef] [PubMed]
- Leder, B.Z. Parathyroid hormone and parathyroid hormone-related protein analogs in osteoporosis therapy. Curr. Osteoporos. Rep. 2017, 15, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Jelinic, M.; Marshall, S.A.; Stewart, D.; Unemori, E.; Parry, L.J.; Leo, C.H. Peptide hormone relaxin: From bench to bedside. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 314, R753–R760. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Verma, S.; Vaidya, S.; Kalia, K.; Tiwari, V. Recent updates on GLP-1 agonists: Current advancements & challenges. Biomed. Pharmacother. 2018, 108, 952–962. [Google Scholar] [CrossRef] [PubMed]
- John, H.; Maronde, E.; Forssmann, W.G.; Meyer, M.; Adermann, K. N-terminal acetylation protects glucagon-like peptide GLP-1-(7-34)-amide from DPP-IV-mediated degradation retaining cAMP- and insulin-releasing capacity. Eur. J. Med. Res. 2008, 13, 73–78. [Google Scholar] [PubMed]
- Eggenstein, E.; Richter, A.; Skerra, A. FluoroCalins: Engineered lipocalins with novel binding functions fused to a fluorescent protein for applications in biomolecular imaging and detection. Protein Eng. Des. Sel. 2019, 32, 289–296. [Google Scholar] [CrossRef]
- Mathiesen, D.S.; Bagger, J.I.; Bergmann, N.C.; Lund, A.; Christensen, M.B.; Vilsboll, T.; Knop, F.K. The effects of dual GLP-1/GIP receptor agonism on glucagon secretion–A review. Int. J. Mol. Sci. 2019, 20, 4092. [Google Scholar] [CrossRef]
- Lan, H.; Hao, Y.; Lv, Y.; Li, G.; Mo, Y.; Zheng, C.; Yuanfa, L. Synergistic effect of a combination of granulocyte macrophage colony-stimulating factor and thymosin α1 on Lewis lung cancer transplanted tumor in mice. Trop. J. Pharm. Res. 2020, 19, 759–764. [Google Scholar] [CrossRef]
- Kjeldsen, T.; Hogendorf, W.F.J.; Tornoe, C.W.; Anderson, J.; Hubalek, F.; Stidsen, C.E.; Sorensen, J.L.; Hoeg-Jensen, T. Dually reactive long recombinant linkers for bioconjugations as an alternative to PEG. ACS Omega 2020, 5, 19827–19833. [Google Scholar] [CrossRef]
- Fletcher, A.M.; Tellier, P.; Douville, J.; Mansell, P.; Graziano, M.J.; Mangipudy, R.S.; Brodie, T.A.; Achanzar, W.E. Adverse vacuolation in multiple tissues in cynomolgus monkeys following repeat-dose administration of a PEGylated protein. Toxicol. Lett. 2019, 317, 120–129. [Google Scholar] [CrossRef]
- Nganou-Makamdop, K.; Billingsley, J.M.; Yaffe, Z.; O’Connor, G.; Tharp, G.K.; Ransier, A.; Laboune, F.; Matus-Nicodemos, R.; Lerner, A.; Gharu, L.; et al. Type I IFN signaling blockade by a PASylated antagonist during chronic SIV infection suppresses specific inflammatory pathways but does not alter T cell activation or virus replication. PLoS Pathog. 2018, 14, e1007246. [Google Scholar] [CrossRef] [PubMed]
- Scopes, R.K. Protein Purification; Springer: New York, NY, USA, 1994; pp. 44–48. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Tα1-PAS |
|---|---|
| Cmax (mg/L) | 25.6 ± 4.4 |
| tmax (h) | 22.7 ± 1.1 |
| AUC0-∞ (h µg/mL) | 1586.7 ± 295.1 |
| τ1/2α (h) | 15.7 ± 0.8 |
| τ1/2β (h) | 15.9 ± 0.9 |
| CL (mL/h/kg) | 2.2 ± 0.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Binder, U.; Skerra, A. PASylated Thymosin α1: A Long-Acting Immunostimulatory Peptide for Applications in Oncology and Virology. Int. J. Mol. Sci. 2021, 22, 124. https://doi.org/10.3390/ijms22010124
Binder U, Skerra A. PASylated Thymosin α1: A Long-Acting Immunostimulatory Peptide for Applications in Oncology and Virology. International Journal of Molecular Sciences. 2021; 22(1):124. https://doi.org/10.3390/ijms22010124
Chicago/Turabian StyleBinder, Uli, and Arne Skerra. 2021. "PASylated Thymosin α1: A Long-Acting Immunostimulatory Peptide for Applications in Oncology and Virology" International Journal of Molecular Sciences 22, no. 1: 124. https://doi.org/10.3390/ijms22010124
APA StyleBinder, U., & Skerra, A. (2021). PASylated Thymosin α1: A Long-Acting Immunostimulatory Peptide for Applications in Oncology and Virology. International Journal of Molecular Sciences, 22(1), 124. https://doi.org/10.3390/ijms22010124