Colorectal Cancer Invasion and Atrophy of the Enteric Nervous System: Potential Feedback and Impact on Cancer Progression

Abstract
1. Introduction: General Features of Cancer Invasion
2. Morphology and Neurotransmission within the Enteric Nervous System
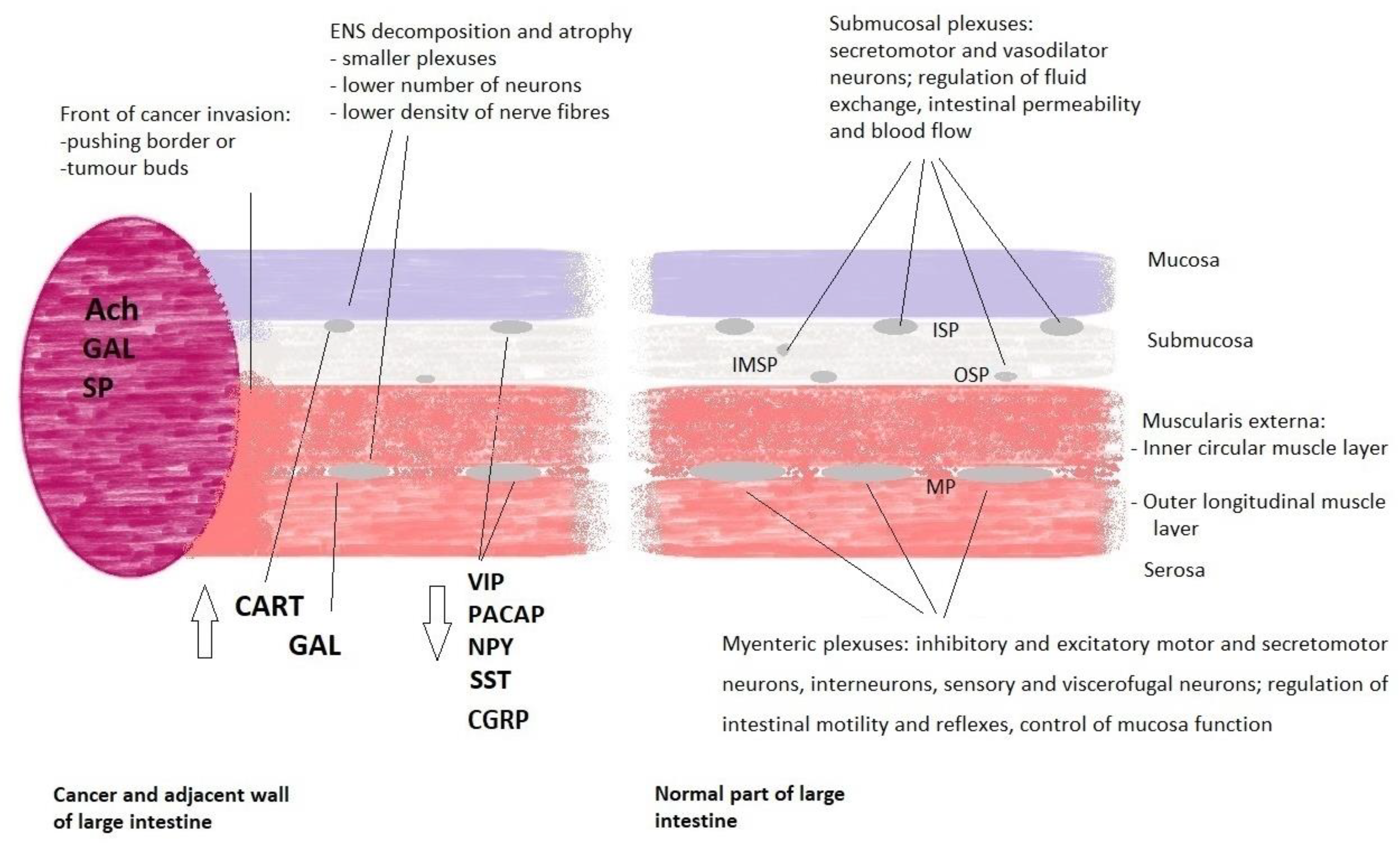
3. Cancer and the ENS Decomposition
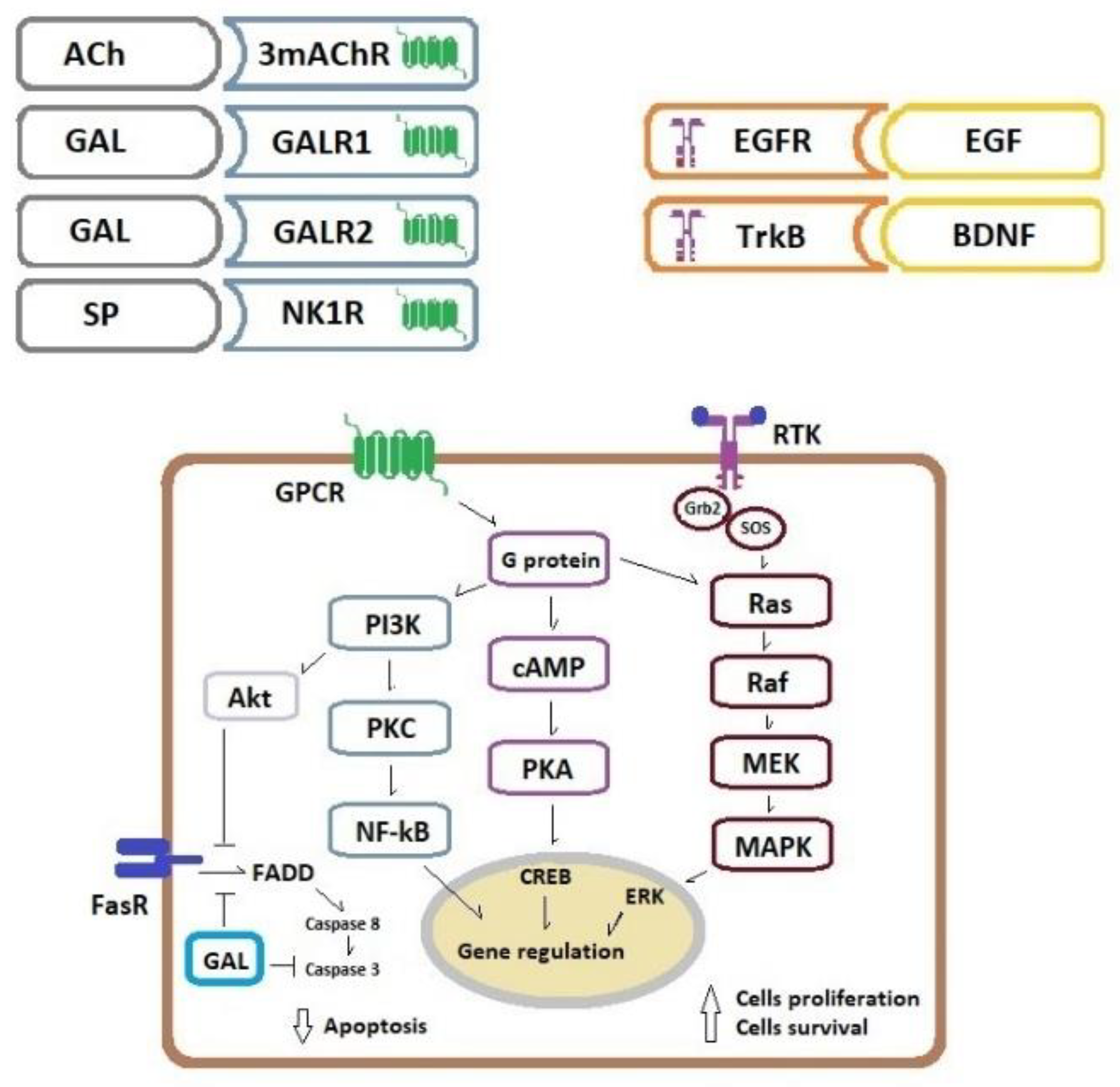
4. Cancer Cell Proliferation and Apoptosis Inhibition Related to the ENS Neurotransmitters/Neuromodulators
5. Cancer Perineural Invasion
6. Conclusions
Funding
Conflicts of Interest
References
- Koelzer, V.H.; Lugli, A. The tumor border configuration of colorectal cancer as histomorphological prognostic indicator. Front. Oncol. 2014, 4, 29. [Google Scholar] [CrossRef] [PubMed]
- Dawson, H.; Lugli, A. Molecular and pathogenetic aspects of tumor budding in colorectal cancer. Front. Med. (Lausanne) 2015, 2, 11. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Yu, B.; Gao, H.; Mu, J.; Ji, C. Matrix Metalloproteinase 2 Overexpression and Prognosis in Colorectal Cancer: A Meta-Analysis. Mol. Biol. Rep. 2013, 40, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Araújo, R.F., Jr.; Lira, G.A.; Vilaça, J.A.; Guedes, H.G.; Leitão, M.C.A.; Lucena, H.F.; Ramos, C.C.O. Prognostic and Diagnostic Implications of MMP-2, MMP-9, and VEGF-α Expressions in Colorectal Cancer. Pathol. Res. Pract. 2015, 211, 71–77. [Google Scholar] [CrossRef]
- Herszényi, L.; Sipos, F.; Galamb, O.; Solymosi, N.; Hritz, I.; Miheller, P.; Berczi, L.; Molnár, B.; Tulassay, Z. Matrix metalloproteinase-9 expression in the normal mucosa-adenoma-dysplasia-adenocarcinoma sequence of the colon. Pathol. Oncol. Res. 2008, 14, 31–37. [Google Scholar] [CrossRef]
- Bendardaf, R.; Buhmeida, A.; Hilska, M.; Laato, M.; Syrjänen, S.; Syrjänen, K.; Collan, Y.; Pyrhönen, S. MMP-9 (gelatinase B) expression is associated with disease-free survival and disease-specific survival in colorectal cancer patients. Cancer Investig. 2010, 28, 38–43. [Google Scholar] [CrossRef]
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix metalloproteinases: Regulators of the tumor microenvironment. Cell 2010, 141, 52–67. [Google Scholar] [CrossRef]
- Timmermans, J.P.; Adriaensen, D.; Cornelissen, W.; Scheuermann, D.W. Structural organization and neuropeptide distribution in the mammalian enteric nervous system, with special attention to those components involved in mucosal reflexes. Comp. Biochem. Physiol. 1997, 118, 331–340. [Google Scholar] [CrossRef]
- Hansen, M.B. The enteric nervous system I: Organisation and classification. Pharmacol. Toxicol. 2003, 92, 105–113. [Google Scholar] [CrossRef]
- Hansen, M.B. The enteric nervous system II: Gastrointestinal functions. Pharmacol. Toxicol. 2003, 92, 249–257. [Google Scholar] [CrossRef]
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.J. The Enteric Nervous System and Gastrointestinal Innervation: Integrated Local and Central Control. In Microbial Endocrinology: The Microbiota-Gut-Brain Axis in Health and Disease (Advances in Experimental Medicine and Biology); Lyte, M., Cryan, J., Eds.; Springer: New York, NY, USA, 2014; Volume 817. [Google Scholar]
- Natale, G.; Ryskalin, L.; Busceti, C.L.; Biagioni, F.; Fornai, F. The Nature of Catecholamine-Containing Neurons in the Enteric Nervous System in Relationship with Organogenesis, Normal Human Anatomy and Neurodegeneration. Arch. Ital. Biol. 2017, 155, 118–130. [Google Scholar] [PubMed]
- Palmer, J.M.; Greenwood-Van Meerveld, B. Integrative neuroimmunomodulation of gastrointestinal function during enteric parasitism. J. Parasitol. 2001, 87, 483–504. [Google Scholar] [CrossRef]
- El-Salhy, M.; Solomon, T.; Hausken, T.; Gilja, O.H.; Hatlebakk, J.G. Gastrointestinal neuroendocrine peptides/amines in inflammatory bowel disease. World J. Gastroenterol. 2017, 28, 5068–5085. [Google Scholar] [CrossRef]
- Brinkman, D.J.; Ten Hove, A.S.; Vervoordeldonk, M.J.; Luyer, M.D.; de Jonge, W.J. Neuroimmune Interactions in the Gut and Their Significance for Intestinal Immunity. Cells 2019, 8, 670. [Google Scholar] [CrossRef] [PubMed]
- Rademakers, G.; Vaes, N.; Schonkeren, S.; Koch, A.; Sharkey, K.A.; Melotte, V. The role of enteric neurons in the development and progression of colorectal cancer. Biochim. Biophys. Acta. Rev. Cancer 2017, 1868, 420–434. [Google Scholar] [CrossRef]
- Godlewski, J.; Lakomy, I.M. Changes in vasoactive intestinal peptide, pituitary adenylate cyclase-activating polypeptide and neuropeptide Y-ergic structures of the enteric nervous system in the carcinoma of the human large intestine. Folia Histochem. Cytobiol. 2010, 48, 208–216. [Google Scholar] [CrossRef]
- Godlewski, J.; Kaleczyc, J. Somatostatin, substance P and calcitonin gene-related peptide-positive intramural nerve structures of the human large intestine affected by carcinoma. Folia Histochem. Cytobiol. 2010, 48, 475–483. [Google Scholar] [CrossRef]
- Furness, J.B. Integrated Neural and Endocrine Control of Gastrointestinal Function. Adv. Exp. Med. Biol. 2016, 891, 159–173. [Google Scholar]
- Harington, A.M.; Hutson, J.M.; Southwell, B.R. Cholinergic neurotransmission and muscarinic receptors in the enteric nervous system. Prog. Histochem. Cytochem. 2010, 44, 173–202. [Google Scholar] [CrossRef]
- Xue, J.; Askwith, C.; Javed, N.H.; Cooke, H.J. Autonomic nervous system and secretion across the intestinal surface. Auton. Neurosci. 2007, 133, 55–63. [Google Scholar] [CrossRef]
- Cheng, K.; Samimi, R.; Xie, G.; Shant, J.; Drachenberg, C.; Wade, M.; Davis, R.J.; Nomikos, G.; Raufman, J.P. Acetylcholine release by human colon cancer cells mediates autocrine stimulation of cell proliferation. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Belo, A.; Cheng, K.; Chahdi, A.; Shant, J.; Xie, G.; Khurana, S.; Raufman, J.P. Muscarinic receptor agonists stimulate human colon cancer cell migration and invasion. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Raufman, J.P.; Samimi, R.; Shah, N.; Khurana, S.; Shant, J.; Drachenberg, C.; Xie, G.; Wess, J.; Cheng, K. Genetic ablation of M3 muscarinic receptors attenuates murine colon epithelial cell proliferation and neoplasia. Cancer Res. 2008, 68, 3573–3578. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.; Shang, A.C.; Drachenberg, C.B.; Zhan, M.; Raufman, J.P. Differential expression of M3 muscarinic receptors in progressive colon neoplasia and metastasis. Oncotarget 2017, 8, 21106–21114. [Google Scholar] [CrossRef]
- Iwasaki, M.; Akiba, Y.; Kaunitz, J.D. Recent advances in vasoactive intestinal peptide physiology and pathophysiology: Focus on the gastrointestinal system. F1000Research 2019, 8(F1000 Faculty Rev), 1629. [Google Scholar] [CrossRef]
- Krueger, D.; Michel, K.; Zeller, F.; Demir, I.E.; Ceyhan, G.O.; Slotta-Huspenina, J.; Schemann, M. Neural Influences on Human Intestinal Epithelium in Vitro. J. Physiol. 2016, 594, 357–372. [Google Scholar] [CrossRef]
- Chandrasekharan, B.; Nezami, B.G.; Srinivasan, S. Emerging neuropeptide targets in inflammation: NPY and VIP. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, 949–957. [Google Scholar] [CrossRef]
- Jayawardena, D.; Guzman, G.; Gill, R.K.; Alrefai, W.A.; Onyuksel, H.; Dudeja, P.K. Expression and Localization of VPAC1, the Major Receptor of Vasoactive Intestinal Peptide Along the Length of the Intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, 16–25. [Google Scholar] [CrossRef]
- Alleaume, C.; Eychène, A.; Caigneaux, E.; Muller, J.M.; Philippe, M. Vasoactive intestinal peptide stimulates proliferation in HT29 human colonic adenocarcinoma cells: Concomitant activation of Ras/Rap1-B-Raf-ERK signalling pathway. Neuropeptides 2003, 37, 98–104. [Google Scholar] [CrossRef]
- Levy, A.; Gal, R.; Granoth, R.; Dreznik, Z.; Fridkin, M.; Gozes, I. In vitro and in vivo treatment of colon cancer by VIP antagonists. Regul. Pept. 2002, 109, 127–133. [Google Scholar] [CrossRef]
- Ogasawara, M.; Murata, J.; Ayukawa, K.; Saimi, I. Differential effect of intestinal neuropeptides on invasion and migration of colon carcinoma cells in vitro. Cancer Lett. 1997, 119, 123. [Google Scholar] [CrossRef]
- Liu, S.; Zeng, Y.; Li, Y.; Guo, W.; Liu, J.; Ouyang, N. VPAC1 overexpression is associated with poor differentiation in colon cancer. Tumour Biol. 2014, 35, 6397–6404. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, M.; Murata, J.; Kamitani, Y.; Hayashi, K.; Saiki, I. Inhibition by vasoactive intestinal polypeptide (VIP) of angiogenesis induced by murine Colon 26-L5 carcinoma cells metastasized in liver. Clin. Exp. Metastasis. 1999, 17, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, M.; Murata, J.; Ayukawa, K.; Saiki, I. Inhibitory effect of vasoactive intestinal polypeptide (VIP) on experimental liver metastasis by murine colon 26-L5 carcinoma cells. Oncol. Res. 1998, 10, 361–370. [Google Scholar]
- Iishi, H.; Tatsuta, M.; Baba, M.; Okuda, S.; Taniguchi, H. Enhacement by vasoactive intestinal peptide of experimental carcinogenesis induced by azoxymethane in rat colon. Cancer Res. 1987, 47, 4890–4893. [Google Scholar]
- Vu, J.P.; Benhammou, J.N.; Goyal, D.; Luong, L.; Oh, S.; Germano, P.; Pisegna, J.R. PACAP regulation of gastrointestinal function and obesity. Current. In Topics of Neurotoxicity 11 Pituitary Adenylate Cyclase Activating Polypeptide-PACAP; Reglodi, D., Tamas, A., Eds.; Springer International Publishing AG: Cham, Switzerland, 2016; pp. 261–270. [Google Scholar]
- Wu, M.J.; Kee, K.H.; Na, J.; Kim, S.W.; Bae, Y.; Shin, D.H.; Choi, S.; Jun, J.Y.; Jeong, H.-S.; Park, J.-S. Pituitary adenylate cyclase-activating polypeptide inhibits pacemaker activity of colonic interstitial cells of Cajal. Korean J. Physiol. Pharmacol. 2015, 19, 435–440. [Google Scholar] [CrossRef]
- Le, S.V.; Yamaguchi, D.J.; McArdle, C.A.; Tachiki, K.; Pisegna, J.R.; Germano, P. PAC1 and PACAP expression, signaling, and effect on the growth of HCT8, human colonic tumor cells. Regul. Pept. 2002, 109, 115–125. [Google Scholar] [CrossRef]
- Szanto, Z.; Sarszegi, Z.; Reglodi, D.; Nemeth, J.; Szabadfi, K.; Kiss, P.; Varga, A.; Banki, E.; Csanaky, K.; Gaszner, B.; et al. PACAP immunoreactivity in human malignant tumor samples and cardiac diseases. J. Mol. Neurosci. 2012, 48, 667–673. [Google Scholar] [CrossRef]
- Nemetz, N.; Abad, C.; Lawson, G.; Nobuta, H.; Chhith, S.; Duong, L.; Tse, G.; Braun, J.; Waschek, J.A. Induction of colitis and rapid development of colorectal tumors in mice deficient in neuropeptide PACAP. Int. J. Cancer 2008, 122, 1803–1809. [Google Scholar] [CrossRef]
- Holzer, P.; Reichmann, F.; Farzi, A. Neuropeptide Y, peptide YY and pancreatic polypeptide in the gut-brain axis. Neuropeptides 2012, 46, 261–274. [Google Scholar] [CrossRef]
- Roperch, J.-P.; Incitti, R.; Forbin, S.; Bard, F.; Mansour, H.; Mesli, F.; Baumgaertner, I.; Brunetti, F.; Sobhani, I. Aberrant methylation of NPY, PENK, and WIF1 as a promising marker for blood-based diagnosis of colorectal cancer. BMC Cancer 2013, 13, 566. [Google Scholar] [CrossRef] [PubMed]
- Garrigou, S.; Perkins, G.; Garlan, F.; Normand, C.; Didelot, A.; le Corre, D.; Peyvandi, S.; Mulot, C.; Niarra, R.; Aucouturier, P.; et al. A study of hypermethylated circulating tumor DNA as a universal colorectal cancer biomarker. Clin. Chem. 2016, 62, 1129–1139. [Google Scholar] [CrossRef]
- Jeppsson, S.; Srinivasan, S.; Chandrasekharan, B. Neuropeptide Y (NPY) promotes inflammation-induced tumorigenesis by enhancing epithelial cell proliferation. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G103–G111. [Google Scholar] [CrossRef]
- Rosso, M.; Robles-Frías, M.J.; Coveñas, R.; Salinas-Martín, M.V.; Muñoz, M. The NK-1 receptor is expressed in human primary gastric and colon adenocarcinomas and is involved in the antitumor action of L-733,060 and the mitogenic action of substance P on human gastrointestinal cancer cell lines. Tumour Biol. 2008, 29, 245–254. [Google Scholar] [CrossRef]
- Garnier, A.; Vykoukal, J.; Hubertus, J.; Alt, E.; von Schweinitz, D.; Kappler, R.; Berger, M.; Ilmer, M. Targeting the neurokinin-1 receptor inhibits growth of human colon cancer cell. Int. J. Oncol. 2015, 47, 151–160. [Google Scholar] [CrossRef]
- Chen, X.Y.; Ru, G.Q.; Ma, Y.Y.; Xie, J.; Chen, W.Y.; Wang, H.J.; Wang, S.B.; Li, L.; Jin, K.T.; He, X.L.; et al. High expression of substance P and its receptor neurokinin-1 receptor in colorectal cancer is associated with tumor progression and prognosis. Onco. Targets Ther. 2016, 9, 3595–3602. [Google Scholar]
- Gonkowski, S.; Rytel, L. Somatostatin as an active substance in the mammalian enteric nervous system. Int. J. Mol. Sci. 2019, 20, 4461. [Google Scholar] [CrossRef]
- Colucci, R.; Blandizzi, C.; Ghishu, N.; Florio, T.; Del Tacca, M. Somatostatin inhibits colon cancer cell growth through cyclooxygenase-2 downregulation. Br. J. Pharmacol. 2008, 155, 198–209. [Google Scholar] [CrossRef]
- Wang, S.; Bao, Z.; Liang, Q.M.; Long, J.W.; Xiao, Z.S.; Jiang, Z.J.; Liu, B.; Yang, J.; Long, Z.X. Octreotide stimulates somatostatin receptor-induces apoptosis of SW480 colon cancer cells by activation of glycogen synthase kinase-3β, a WNT/β pathway modulator. Hepatogastroenterology 2013, 60, 1639–1646. [Google Scholar] [PubMed]
- Chen, J.S.; Liang, Q.M.; Li, H.S.; Yang, J.; Wang, S.; Long, J.W. Octrotide inhibits growth of colonic cancer SW480 cells by modulating the Wnt/P pathway. Pharmazie 2009, 64, 126–131. [Google Scholar]
- Hohla, F.; Buchholz, S.; Schally, A.V.; Krishan, A.; Rick, F.G.; Szalontay, L.; Papadia, A.; Halmos, G.; Koster, F.; Aigner, E.; et al. Targeted cytotoxic somatostatin analog AN-162 ihibits growth of human colon carcinomas and increases sensitivity of doxorubicin resistant murine leukemia cells. Cancer Lett. 2010, 294, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Evangelou, I.; Petraki, C.; Msaouel, P.; Scorilas, A.; Sdrolia, E.; Padazi, G.; Koborozos, V.; Koutsilieris, M. Immunohistochemical expression of somatostatin receptor subtypes 2 and 5 in colorectal cancer. Eur. J. Clin. Investig. 2012, 42, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chew, M.H.; Tham, C.K.; Tang, C.L.; Ong, S.Y.K.; Zhao, Y. Methylation of serum SST gene in an independent prognostic marker in colorectal cancer. Am. J. Cancer Res. 2016, 6, 2098–2108. [Google Scholar] [PubMed]
- Benya, R.V.; Matkowskyj, K.A.; Danilkovich, A.; Hecht, G. Galanin causes Cl-secretion in the human colon. Potential significance of inflammation-associated NF-kappa B activation on galanin-1 receptor expression and function. Ann. N. Y. Acad. Sci. 1998, 863, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Botella, A.; Delvaux, M.; Frexinos, J.; Bueno, L. Comparative effects of galanin on isolated smooth muscle cells from ileum in five mammalian species. Life Sci. 1992, 50, 1253–1261. [Google Scholar] [CrossRef]
- Kim, K.Y.; Kee, M.K.; Chong, S.A.; Nam, M.J. Galanin is up-regulated in colon adenocarcinoma. Cancer Epidemiol. Biomark. Prev. 2007, 16, 2373–2378. [Google Scholar] [CrossRef]
- Stevenson, L.; Allen, W.L.; Turkington, R.; Jithesh, P.V.; Proutski, I.; Stewart, G.; Lenz, H.-J.; Schaeybroeck, S.V.; Longley, D.B.; Johnston, P.G. Identification of galanin and its receptor GalR1 as novel determinants of resistance to chemotherapy and potential biomarkers in colorectal cancer. Clin. Cancer Res. 2012, 18, 5412–5426. [Google Scholar] [CrossRef]
- Kwiatkowski, P.; Godlewski, J.; Kieżun, J.; Kraziński, B.E.; Kmieć, Z. Colorectal cancer patients exhibit increased levels of galanin in serum and colon tissues. Oncol. Lett. 2016, 12, 3323–3329. [Google Scholar] [CrossRef]
- Nagayoshi, K.; Ueki, T.; Tashiro, K.; Mizuuchi, Y.; Manabe, T.; Araki, H.; Oda, Y.; Kuhara, S.; Tanaka, M. Galanin plays an important role in cancer invasiveness and is associated with poor prognosis in stage II colorectal cancer. Oncol. Rep. 2015, 33, 539–546. [Google Scholar] [CrossRef]
- Tell, R.; Rivera, C.A.; Eskra, J.; Taglia, L.N.; Blunier, A.; Wang, Q.T.; Benya, R.V. Gastrin-realising peptide signaling alters colon cancer invasiveness via heterochromatin protein 1HSβ. Am. J. Pathol. 2011, 178, 672–678. [Google Scholar] [CrossRef]
- Carrol, R.E.; Matkowskyj, K.A.; Chakrabarti, S.; McDonald, T.J.; Benya, R.V. Aberrant expression of gastrin-releasing peptide and its receptor by well-differentiated colon cancers in humans. Am. J. Physiol. 1999, 276, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Rivera, C.A.; Ahlberg, N.C.; Taglia, L.; Kumar, M.; Blunier, A.; Benya, R.V. Expression of GRP and its receptor is associated with improved survival in patients with colon cancer. Clin. Exp. Metastasis. 2009, 26, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Godlewski, J.; Pidsudko, Z. Characteristic of galaninergic components of the enteric nervous system in the cancer invasion of human large intestine. Ann. Anat. 2012, 194, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Oponowicz, A.; Kozłowska, A.; Gonkowski, S.; Godlewski, J.; Majewski, M. Changes in the Distribution of Cocaine- And Amphetamine-Regulated Transcript-Containing Neural Structures in the Human Colon Affected by the Neoplastic Process. Int. J. Mol. Sci. 2018, 19, 414. [Google Scholar] [CrossRef]
- Godlewski, J. Morphological changes in the enteric nervous system caused by carcinoma of the human large intestine. Folia Histochem. Cytobiol. 2010, 48, 157–162. [Google Scholar] [CrossRef]
- Zauszkiewicz-Pawlak, A.; Godlewski, J.; Kwiatkowski, P.; Kmiec, Z. Ultrastructural characteristics of myenteric plexus in patients with colorectal cancer. Folia Histochem. Cytobiol. 2017, 55, 6–10. [Google Scholar] [CrossRef]
- Ciurea, R.N.; Rogoveanu, I.; Pirici, D.; Tartea, G.C.; Streba, C.T.; Florescu, C.; Catalin, B.; Puiu, I.; Tartea, E.A.; Vere, C.C. B2 adrenic receptors and morphological changes of the enteric nervous system in colorectal adenocarcinoma. World J. Gastroenterol. 2017, 23, 1250–1261. [Google Scholar] [CrossRef]
- Kozlowska, A.; Kwiatkowski, P.; Oponowicz, A.; Majewski, M.; Kmiec, Z.; Godlewski, J. Myenteric plexuses atrophy in the vicinity of colorectal cancer tissue is not caused by apoptosis or necrosis. Folia Histochem. Cytobiol. 2016, 54, 99–107. [Google Scholar] [CrossRef]
- Van Neerven, S.M.; Vermeulen, L. The interplay between intrinsic and extrinsic Wnt signaling in controlling intestinal transformation. Differentiation 2019, 108, 17–23. [Google Scholar] [CrossRef]
- Konishi, M.; Hayakawa, Y.; Koike, K. Role of Muscarinic Acetylcholine Signaling in Gastrointestinal Cancers. Biomedicines 2019, 7, 58. [Google Scholar] [CrossRef]
- Von Rosenvinge, E.C.; Raufman, J.P. Muscarinic receptor signaling in colon cancer. Cancers 2011, 3, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.; Khurana, S.; Cheng, K.; Raufman, J.P. Muscarinic receptors and ligands in cancer. Am. J. Physiol. Cell Physiol. 2009, 296, C221–C232. [Google Scholar] [CrossRef] [PubMed]
- Labed, S.A.; Wani, K.A.; Jagadeesan, S.; Hakkim, A.; Najibi, M.; Irazoqui, J.E. Intestinal Epithelial Wnt Signaling Mediates Acetylcholine-Triggered Host Defense against Infection. Immunity 2018, 48, 963–978. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Cheng, K.; Shant, J.; Raufman, J.P. Acethylcholine induced activation of M3 muscarinic receptors stimulates robust matrix metalloproteinase gene expression in human colon cancer cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, 755–763. [Google Scholar] [CrossRef]
- Tolaymat, M.; Larabee, S.M.; Hu, S.; Xie, G.; Raufman, J.-P. The Role of M3 Muscarinic Receptor Ligand-Induced Kinase Signaling in Colon Cancer Progression. Cancers 2019, 11, 308. [Google Scholar] [CrossRef]
- Carethers, J.M.; Jung, B.H. Genetics and Genetic Biomarkers in Sporadic Colorectal Cancer. Gastroenterology 2015, 149, 1177–1190. [Google Scholar] [CrossRef]
- Lakomy, M.; Sienkiewicz, W.; Zmudzki, J.; Wasowicz, K. Changes in the expression of some neuropeptide in the intestines and nerve ganglia during the porcine dysyntery. Bull. Vet. Inst. Pulawy. 2005, 49, 393–398. [Google Scholar]
- Skobowiat, C.; Calka, J.; Majewski, M. Axotomy induced changes in neuronal plasticity of sympathetic chain ganglia (SChG) neurons supplying descending colon in the pig. Exp. Mol. Pathol. 2011, 90, 13–18. [Google Scholar] [CrossRef]
- Lang, R.; Gundlach, A.L.; Kofler, B. The galanin peptide family: Receptor pharmacology, pleiotropic biological actions, and implications in health and disease. Pharmacol. Ther. 2007, 115, 177–207. [Google Scholar] [CrossRef]
- Gudermann, T.; Roelle, S. Calcium-dependent growth regulation of small cell lung cancer cells by neuropeptides. Endocr. Relat. Cancer 2006, 13, 1069–1084. [Google Scholar] [CrossRef]
- Banerjee, R.; Henson, B.S.; Russo, N.; Tsodikov, A.; D’Silva, N.J. Rap1 mediates galanin receptor 2-induced proliferation and survival in squamous cell carcinoma. Cell Signal. 2011, 23, 1110–1118. [Google Scholar] [CrossRef] [PubMed]
- Mashaghi, A.; Marmalidou, A.; Tehrani, M.; Grace, P.M.; Pothoulakis, C.; Dana, R. Neuropeptide substance P and the immune response. Cell Mol. Life Sci. 2016, 73, 4249–4264. [Google Scholar] [CrossRef]
- Muñoz, M.; Coveñas, R. Neurokinin-1 receptor antagonists as antitumor drugs in gastrointestinal cancer: A new approach. Saudi J. Gastroenterol. 2016, 22, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Knijn, N.; Mogk, S.C.; Teerenstra, S.; Simmer, F.; Nagtegaal, I.D. Perineural Invasion is a Strong Prognostic Factor in Colorectal Cancer: A Systematic Review. Am. J. Surg. Pathol. 2016, 40, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Liebig, C.; Ayala, G.; Wilks, J.; Verstovsek, G.; Liu, H.; Agarwal, N.; Berger, D.H.; Albo, D. Perineural invasion is an independent predictor of outcome in colorectal cancer. J. Clin. Oncol. 2009, 27, 5131–5137. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, H.; Gong, H.; Cao, M.; Zhang, G.; Wang, Y. Clinical significance of perineural invasion in stages II and III colorectal cancer. Pathol. Res. Pract. 2015, 211, 839–844. [Google Scholar] [CrossRef]
- Duchalais, E.; Guilluy, C.; Steven Nedellec, S.; Touvron, M.; Bessard, A.; Touchefeu, Y.; Bossard, C.; Boudin, H.; Louarn, G.; Neunlist, M.; et al. Colorectal Cancer Cells Adhere to and Migrate Along the Neurons of the Enteric Nervous System. Cell Mol. Gastroenterol. Hepatol. 2017, 5, 31–49. [Google Scholar] [CrossRef]
- Adad, S.J.; Etchebehere, R.M.; Araujo, J.R.; Madureira, A.B.; Lima, V.G.; Silva, A.A.; Eduardo, C. Association of chagasic megacolon and cancer of the colon: Case report and review of the literature. Rev. Soc. Bras. Med. Trop. 2002, 35, 63–68. [Google Scholar] [CrossRef][Green Version]
- Garcia, S.B.; Aranha, A.L.; Garcia, F.R.; Basile, F.V.; Pinto, A.P.; de Oliveira, E.C.; Zucoloto, S. A retrospective study of histopathological findings in 894 cases of megacolon: What is the relationship between megacolon and colonic cancer? Rev. Inst. Med. Trop. Sao Paulo 2003, 45, 91–93. [Google Scholar] [CrossRef]
- Kannen, V.; de Oliveira, E.C.; Motta, B.Z.; Chaguri, A.J.; Brunaldi, M.O.; Garcia, S.B. Trypanosomiasis-induced megacolon illustrates how myenteric neurons modulate the risk for colon cancer in rats and humans. PLoS Negl. Trop. Dis. 2015, 9, e0003744. [Google Scholar] [CrossRef]
- Amit, M.; Na’ara, S.; Gil, Z. Mechanisms of cancer dissemination along nerves. Nat. Rev. Cancer 2016, 16, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Liu, S. Neurotrophic factors in enteric physiology and pathophysiology. Neurogastroenterol. Motil. 2018, 30, e13446. [Google Scholar] [CrossRef]
- Meldolesi, J. Neurotrophin receptors in the pathogenesis, diagnosis and therapy of neurodegenerative diseases. Pharmacol. Res. 2017, 121, 129–137. [Google Scholar] [CrossRef]
- Reichardt, L.F. Neurotrophin-regulated signalling pathways. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1545–1564. [Google Scholar] [CrossRef] [PubMed]
- Brunetto de Farias, C.; Rosenberg, D.B.; Heinen, T.E.; Koehler-Santos, P.; Abujamra, A.L.; Kapczinski, F.; Brunetto, A.L.; Aston-Prolla, P.; Meurer, L.; Reis Bogo, M.; et al. BDNF/TrkB content and interaction with gastrin-releasing peptide receptor blockade in colorectal cancer. Oncology 2010, 79, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhang, S.; Wang, X.; Yang, Z.; Ou, G. Overexpression of TrkB promotes the progression of colon cancer. APMIS 2010, 118, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Liu, B.; Ji, R.; Jiang, X.; Yan, X.; Xin, Y. Targeting the BDNF/TrkB pathway for the treatment of tumors. Oncol. Lett. 2019, 17, 2031–2039. [Google Scholar] [CrossRef]
- Mazouffre, C.; Geyl, S.; Perraud, A.; Blondy, S.; Jauberteau, M.O.; Mathonnet, M.; Verdier, M. Dual inhibition of BDNF/TrkB and autophagy: A promising therapeutic approach for colorectal cancer. J. Cell Mol. Med. 2017, 21, 2610–2622. [Google Scholar] [CrossRef]
- Blondy, S.; Christou, N.; David, V.; Verdier, M.; Jauberteau, M.O.; Mathonnet, M.; Perraud, A. Neurotrophins and their involvement in digestive cancers. Cell Death Dis. 2019, 10, 123. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| ACh | Normal function in the large intestine |
| Stimulates contraction of muscularis externa smooth muscles; increase fluid secretion and blood flow in submucosa vessels, regulates proper intestinal peristalsis and reflexes [10,19,20,21]. | |
| Localization in the ENS: excitatory muscle motor neurons, secretomotor neurons, viscerofugal mechanosensory neurons and interneurons, vasodilator neurons [8,9]. | |
| Possible involvement in colorectal cancer development: ACh stimulates cancer cell proliferation in an autocrine and paracrine way. | |
| In vitro studies on CRC cell lines | |
| - H508 cells; nonselective muscarinic receptor antagonists, selective 3mAChR antagonist, and choline transport inhibitors attenuate cancer cells proliferation; acetylcholinesterase inhibitors induce cell proliferation [22]. | |
| - H508, WiDr, and Caco-2 cells and conditioned-cell culture media; choline acetyltransferase induces cell proliferation [22]; | |
| - H508 and HT29 cells; ACh induces cell migration mediated by MMP-7, which cleaves pro-EGF into EGF, EGFR activation and MAPK/ERK and PI3K signaling [23]. | |
| CRC tumour tissue and in vivo studies | |
| - 3mAChR-deficient mice, chemically-induced CRC model; a lower number of adenoma and adenocarcinoma and lower tumour size [24]. | |
| - in human CRC 3mAChR over-expression (IHC) was found in 70% of adenomas vs adjacent normal epithelium; in CRC tumour cells both intense apical and cytoplasmic immunoreactivity (Ir) vs only apical, weak Ir in normal colon epithelium; no alterations of 3mAChR-Ir in lymph node and liver metastases [25]; | |
| VIP | Normal function in the large intestine |
| As potent vasodilator increases blood flow in submucosal vessels [19,26]; is the primary pro-secretory neurotransmitter in the large intestine [19,26] by increasing colon electrogenic Cl– and HCO3– secretion [27]; affects intestinal barrier function [26]; has anti-inflammatory effects [28]; relaxes of smooth muscles of muscularis externa [9,10]. | |
| Localization in the ENS: inhibitory motor neurons, secretomotor neurons, viscerofugal neurons, part of interneurons and vasodilator neurons [8,9,21]. | |
| VIP receptor (VPAC1) mRNA expression in the human intestine, was highest in the sigmoid colon, followed by ileum and jejunum; in human colon VPAC1 showed predominantly apical localization in the intestinal epithelium [29]. | |
| Possible involvement in colorectal cancer development: VIP stimulates cancer cell proliferation, but has anti-metastatic activity and attenuates angiogenesis. | |
| In vitro studies on CRC cell lines | |
| - HT29 cells; VIP stimulates cells proliferation and induced a time- and concentration-dependent activation of cAMP-Rap1/Ras-B-Raf-ERK signalling pathway [30]. | |
| - HCT-15 cells; VIP antagonists, neurotensin(6-11)VIP(7-28) - inhibitor of cancer cells growth [31]. | |
| - 26-L5 cells; VIP attenuate cancer cells motility and invasiveness potential [32]. | |
| Tumour tissue and in vivo studies | |
| - Human CRC tumours: VPAC1 receptors overexpression (IHC) is associated with poor differentiation in CRC; VPAC1 receptors prevail in blood vessels surrounding tumour CRC and are numerous in tumour-associated macrophages [33]; | |
| - Mice, experimental CRC metastasis model; VIP inhibits liver metastasis in a dose-dependent manner and attenuates angiogenesis through suppression of endothelial cells [34,35]; | |
| - Mice, chemically induced CRC; VIP increases the incidence of colonic tumours [36]; | |
| - Rat, chemically induced CRC, administration of VIP hybrid antagonist neurotensin(6-11)VIP(7-28) for 10 weeks reduced tumour volume, staging, lymphocyte infiltration and the number of dysplastic crypts [31]. | |
| PACAP | Normal function in large intestine |
| Increased intestinal secretion [9,19,37] and relaxation of smooth muscle of muscularis externa [38]. | |
| Localization in the ENS: inhibitory motor neurons and part of interneurons; PACAP is frequently colocalized with the sensory neuropeptide CGRP and also with VIP [8,9]. | |
| Possible involvement in colorectal cancer development: PACAP stimulates cancer cell proliferation and induces cell survival. | |
| In vitro studies on CRC cell lines | |
| - HCT8 cells; PACAP-38 elevates via PAC1 receptor both intracellular cAMP levels and cytosolic Ca(2+) concentration, increases the number of cancer cells, attenuates apoptotic signaling via Fas-R/Fas-L down-regulation [39]. | |
| Tumour tissue and in vivo studies | |
| PACAP38- and PACAP27-like immunoreactivity was lower in tissue samples of of CRC compared with normal colon tissue [40]. | |
| PACAP-deficient mice, rapid development of CRC tumours in chemically induced CRC experimental model [41]. | |
| NPY | Normal function in large intestine |
| Increased intestinal secretion, regulation of blood flow in the submucosa, relaxation of muscularis externa [19,21,42], proinflammatory effects [28] | |
| Localization in the ENS: inhibitory motor neurons, secretomotor neurons and innervation of submucosal blood vessels [8,9]. | |
| Possible involvement in colorectal cancer development: reduces cancer cells invasiveness. | |
| In vitro studies on CRC cell lines | |
| - 26-L5 cells; NPY suppresses cancer cell motility, inhibits cell migration, reduces invasive potential of tumour cells in a concentration-dependent manner [32]. | |
| Studies on CRC tumour and blood samples, CRC biomarker, in vivo studies | |
| - Hypermethylated NPY gene, is a marker to screen patients with high risk of colorectal cancer [43], and hypermethylated NPY circulating tumour DNA (MetctDNA), is the useful marker of cancer development [44]. | |
| - NPY promotes inflammation-induced tumorigenesis (DSS-mouse model) by increasing epithelial cell proliferation and downregulating apoptosis [45] | |
| CGRP | Normal function in large intestine |
| Intestinal sensation and modulation of intestinal secretion [19,21] | |
| Localization in the ENS: sensory neurons and secretomotor neurons and innervation of submucosal blood vessels [8,9]. | |
| Possible involvement in colorectal cancer development: reduces invasive potential of cancer cells. | |
| In vitro studies on CRC cell lines | |
| - 26-L5 cells; CGRP suppresses cancer cell motility, inhibits cell migration, reduces invasive potential of tumour cells in a concentration-dependent manner [26] | |
| SP | Normal function in large intestine |
| Modulation of intestinal secretion and intestinal sensation [19], contraction of muscularis externa [21], proinflammatory effects [28]. | |
| Localization in the ENS: excitatory muscle motor neurons, sensory neurons and part of interneurons [9]. | |
| Possible involvement in colorectal cancer development: SP increases cancer cell proliferation. | |
| In vitro studies on CRC cell lines | |
| - 23132/87 and SW-403 cells; L-733,060, neurokinin-1 (NK-1) receptor antagonist - inhibit the growth of CRC cells in a dose-dependent manner via activation apoptosis [46]. | |
| - LiM6 and DLD1 cells; Upregulation of SP-NK1R – induce CRC progression, neurokinin-1 receptor (NK1R) antagonist - inhibit colon cancer cell lines growth via inhibition Wnt signaling [47]. | |
| Tumour tissue and in vivo studies | |
| - CRC tissue sample; SP and NK1R levels is upregulated in CRC, high expression is associated with lymph node metastasis and poor prognosis [48]. | |
| SST (SOM) | Normal function in large intestine |
| Inhibition of intestinal secretion and intestinal motility, blood flow regulation in the submucosa [19,49]; anti-inflammatory effects [28] | |
| Localization in the ENS: secretomotor neurons, part of interneurons, submucosal blood vessels [9,21]. | |
| Possible involvement in colorectal cancer development: STT inhibits growth of colon cancer cells. | |
| In vitro studies on CRC cell lines | |
| - Caco-2, HT-29 and HCT116 cells; STT inhibit cell proliferation, decreases COX-2 expression and function in CRC cells via activation of sst(3) or sst(5) receptors [50]. | |
| - SW480 cells; Octreotide, (STT analog), downregulate Wnt target genes cyclinD1 and c-Myc [51], inhibit Wnt/beta-catenin signaling pathway - arrest the cell cycle and induce apoptosis [52]. | |
| - HT-29, HCT-15, and HCT-116 cells; AN-162 (SST analog) increase the number of apoptotic cells [53]. | |
| Studies on CRC tumour and blood samples | |
| - CRC tissue samples; SST receptor type 2 and 5 - negative correlation with CRC invasion and liver metastasis. Patients with longer survival [54]. | |
| - CRC blood samples; methylated SST gene, at the preoperative time point - associated with patients shorter survival and predictor for cancer recurrence [55]. | |
| GAL | Normal function in large intestine |
| Modulates intestinal secretion, contraction of muscularis mucosae, increases blood flow in the submucosa [19,56,57], anti-inflammatory effects [28]. | |
| Localization in the ENS: secretomotor neurons and vasodilator neurons [8]. | |
| Possible involvement in colorectal cancer development: GAL induces proliferation of cancer cells and improves cell survival | |
| In vitro studies on CRC cell lines | |
| - LOVO, HCT15, SW480 and SW620 cells: high levels of galanin expression [58]; | |
| - HCT116, LS174T, RKO, HT29, SW620 and LoVo; GalR1/galanin silencing results in the downregulation of the endogenous caspase-8 inhibitor FLIP(L), which results in the induction of caspase-8-dependent apoptosis [59]. | |
| Studies on CRC tumour and blood samples, CRC biomarker. | |
| - CRC tissue: increased GAL gene expression [58] and GAL levels [60] correlate with poor disease-free survival, poor prognosis and tumour recurrence of CRC patients [59,61] | |
| - CRC blood samples; 2.4 times higher GAL concentrations than in healthy control [60]. | |
| GRP | Normal function in large intestine |
| Relaxation of muscularis externa [19]. | |
| Localization in the ENS neurons: inhibitory motor neurons and part of interneurons [9,21] | |
| Possible involvement in colorectal cancer development: GRP attenuates CRC invasiveness. | |
| In vitro studies on CRC cell lines | |
| - Caco-2 and LS-174T cells; inhibiting RP/GRPR signalling increases invasiveness of cells [62]. | |
| Tumour tissue | |
| - GRP/GRPR co-expression in all well-differentiated part of CRC tumours [63], GRP and GRPR expression (IHC) correlated with better patients survival [64]. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godlewski, J.; Kmiec, Z. Colorectal Cancer Invasion and Atrophy of the Enteric Nervous System: Potential Feedback and Impact on Cancer Progression. Int. J. Mol. Sci. 2020, 21, 3391. https://doi.org/10.3390/ijms21093391
Godlewski J, Kmiec Z. Colorectal Cancer Invasion and Atrophy of the Enteric Nervous System: Potential Feedback and Impact on Cancer Progression. International Journal of Molecular Sciences. 2020; 21(9):3391. https://doi.org/10.3390/ijms21093391
Chicago/Turabian StyleGodlewski, Janusz, and Zbigniew Kmiec. 2020. "Colorectal Cancer Invasion and Atrophy of the Enteric Nervous System: Potential Feedback and Impact on Cancer Progression" International Journal of Molecular Sciences 21, no. 9: 3391. https://doi.org/10.3390/ijms21093391
APA StyleGodlewski, J., & Kmiec, Z. (2020). Colorectal Cancer Invasion and Atrophy of the Enteric Nervous System: Potential Feedback and Impact on Cancer Progression. International Journal of Molecular Sciences, 21(9), 3391. https://doi.org/10.3390/ijms21093391