In Utero Exposure to Bisphenol a Promotes Mammary Tumor Risk in MMTV-Erbb2 Transgenic Mice Through the Induction of ER-erbB2 Crosstalk



Abstract
1. Introduction
2. Results
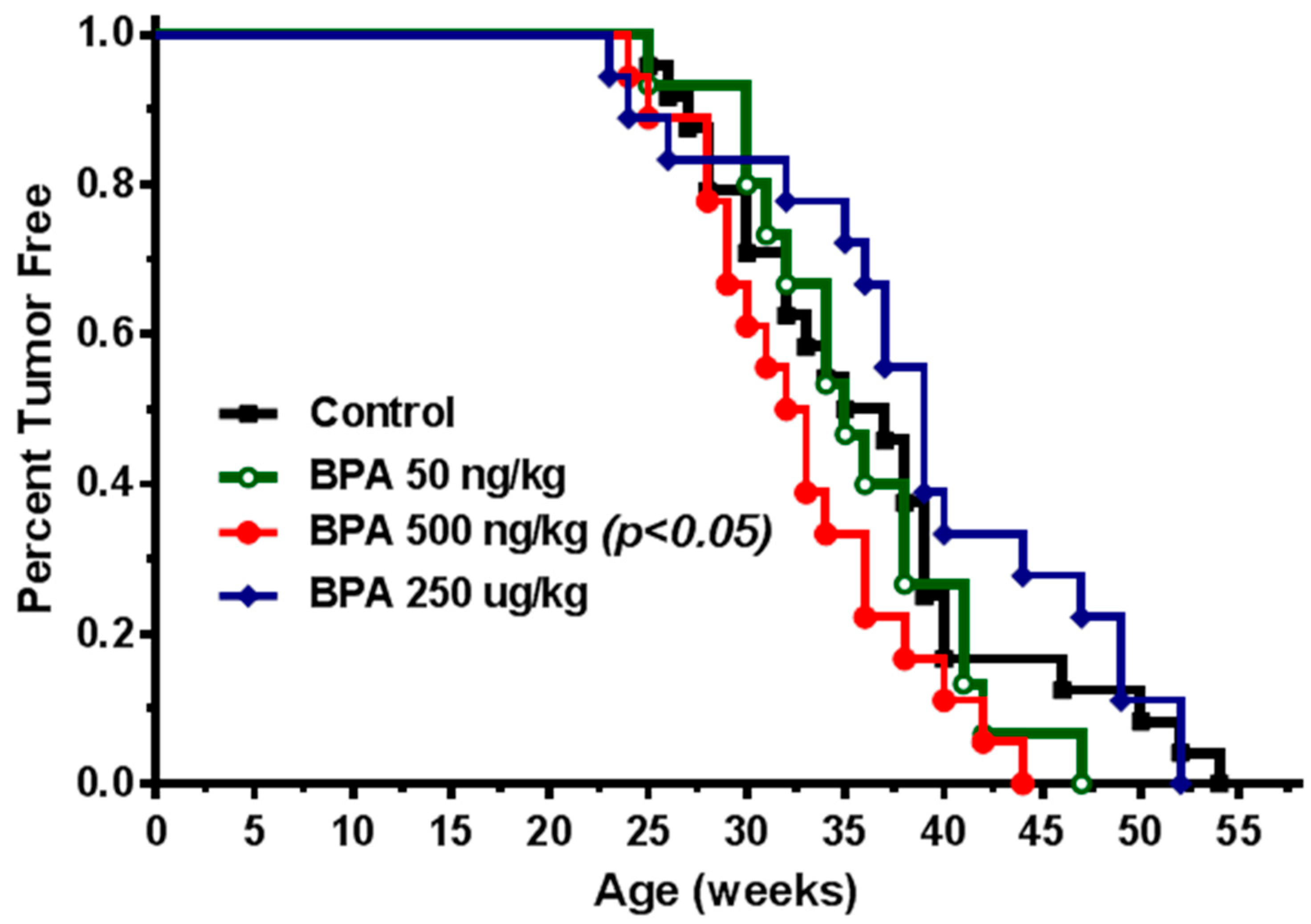
2.1. In Utero Exposure to Low Doses of BPA Promotes Mammary Tumor Development in MMTV-erbB2 Mice
2.2. In Utero BPA Exposure Alters Vaginal Opening Dates and Estrous Cycle in MMTV-erbB2 Transgenic Mice
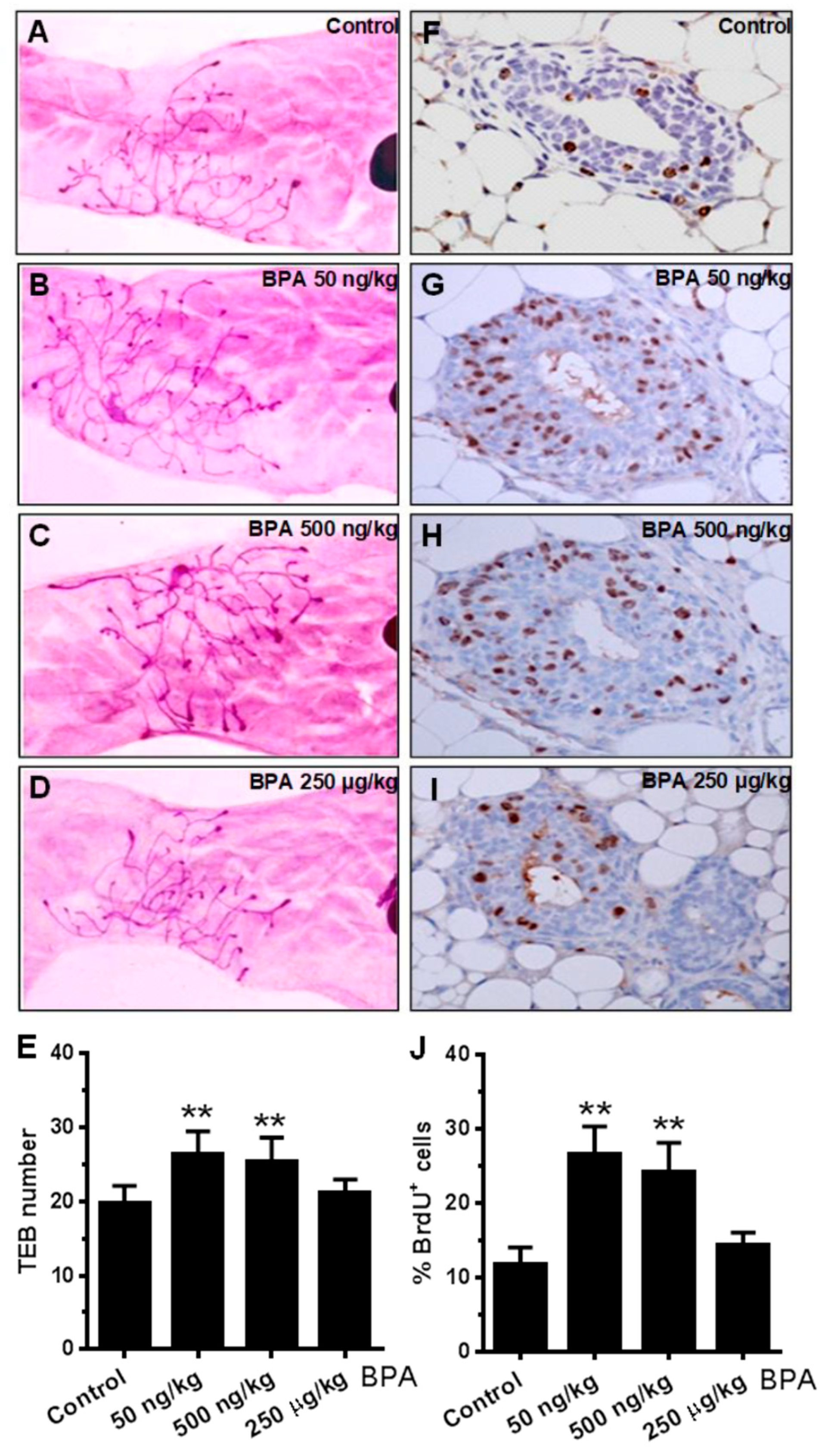
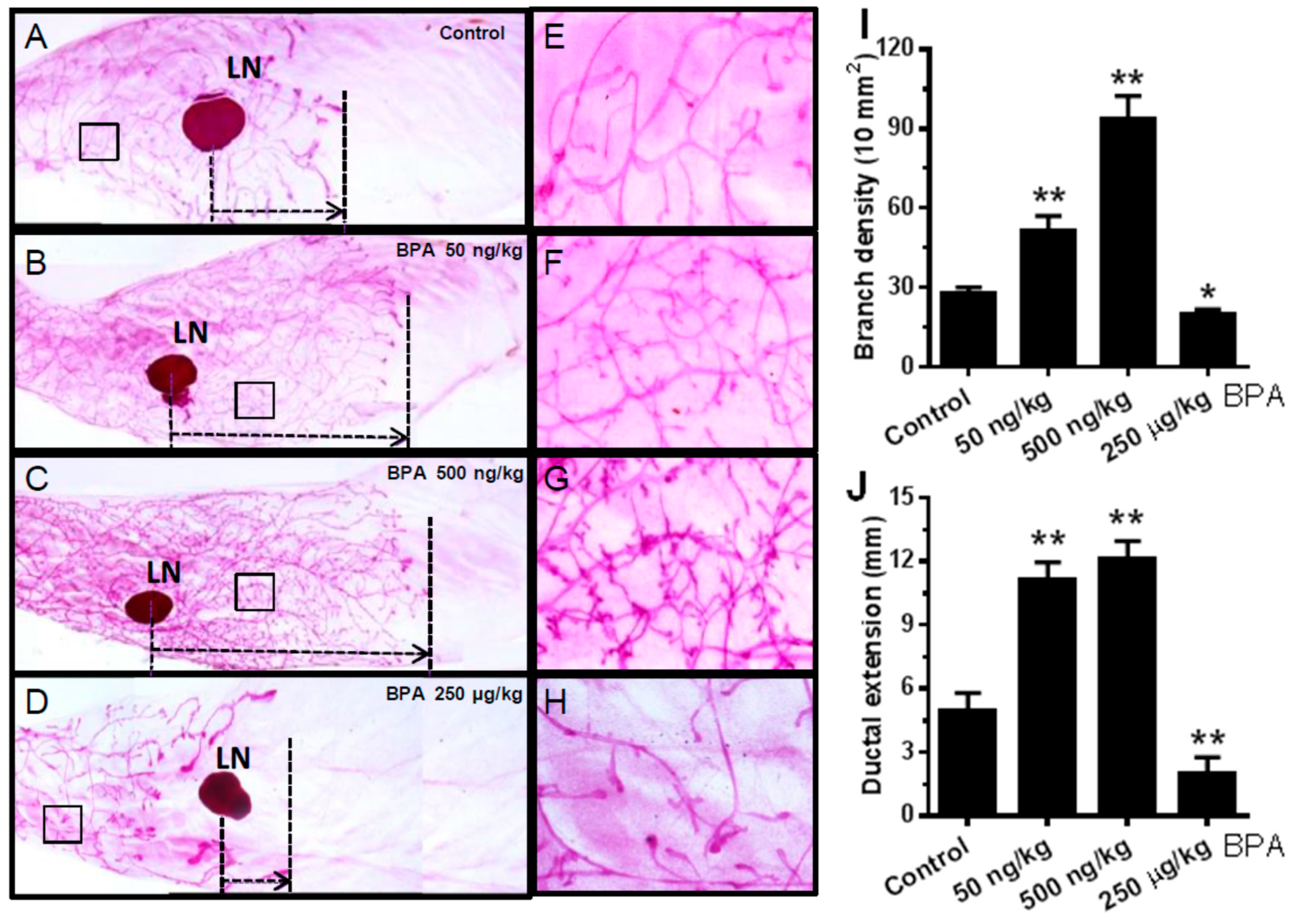
2.3. In Utero Exposure to BPA Induces Proliferative Mammary Morphogenesis
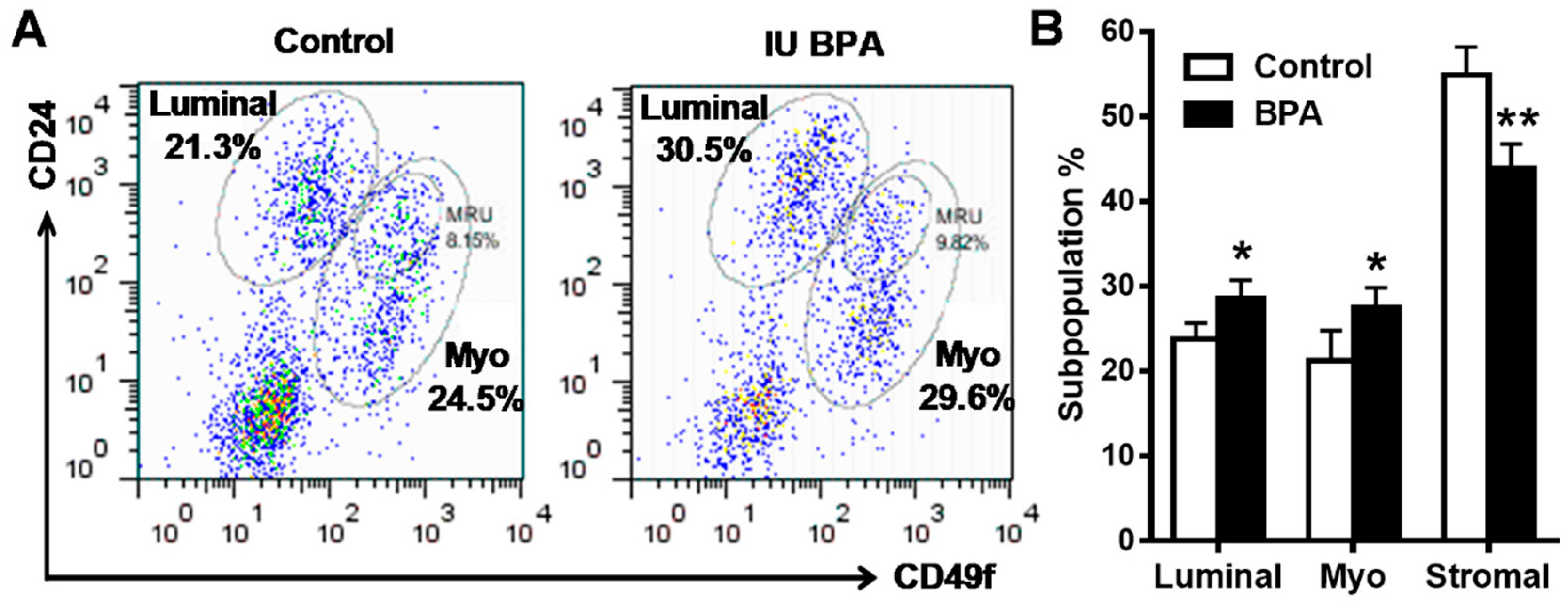
2.4. In Utero Exposure to BPA Induces Mammary Epithelial Cell Repopulation
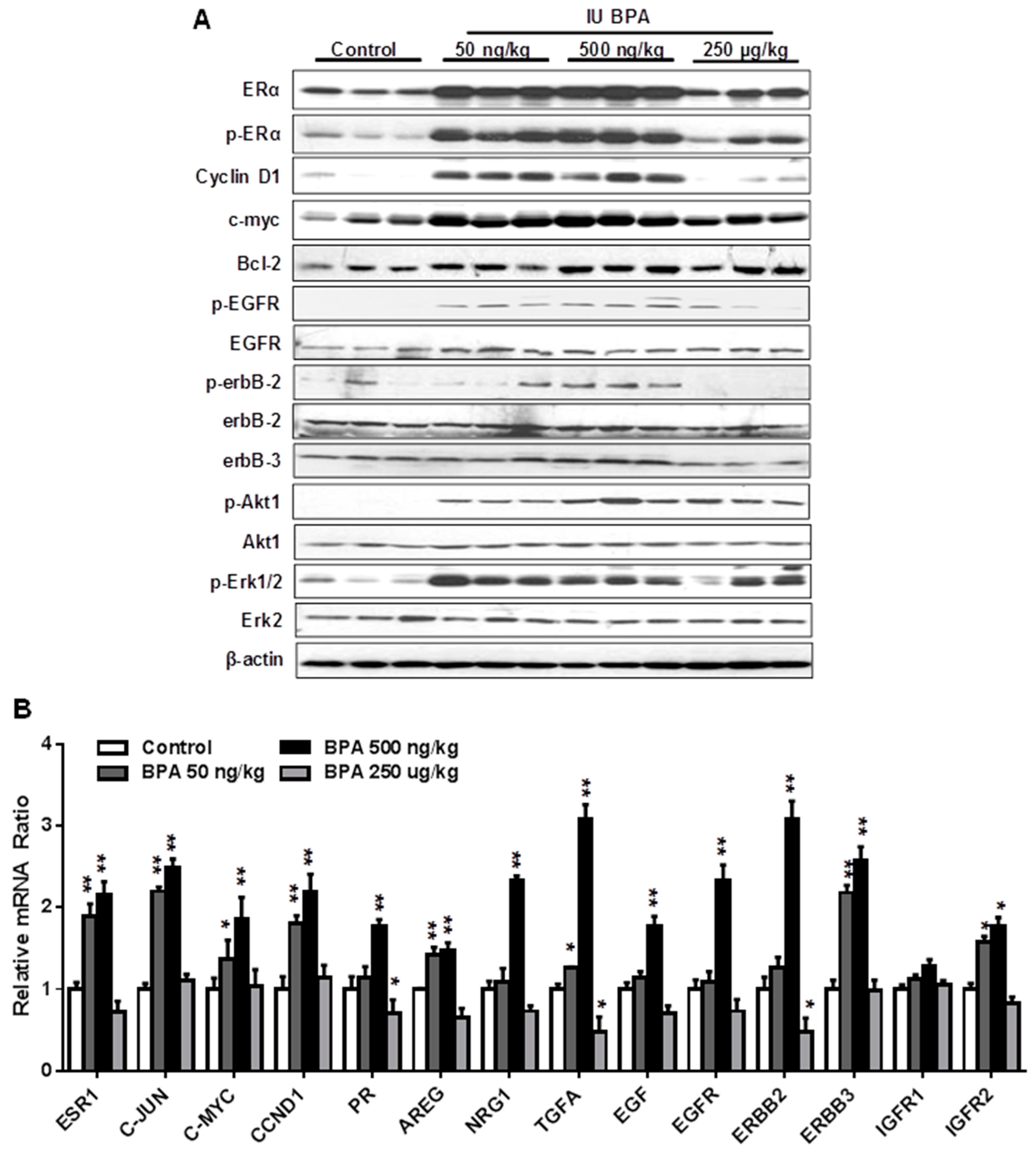
2.5. In Utero Exposure to Low Dose BPA Induces Crosstalk between ER and EGFR/erbB2 Pathways
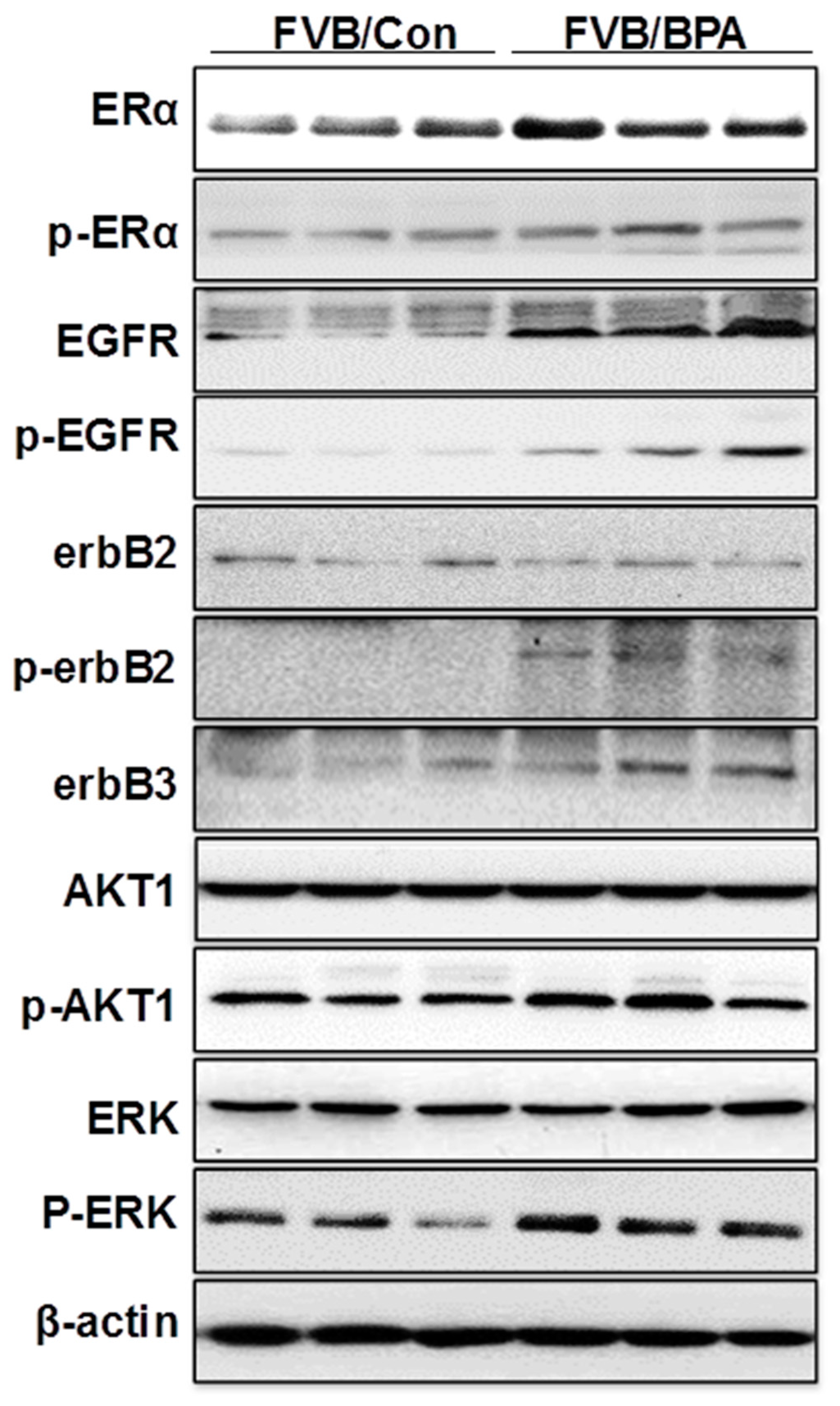
2.6. In Utero Exposure to Low Dose BPA Also Induces ER-EGFR/erbB2 Crosstalk in FVB/N Mice
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Animals and Treatments
4.3. Whole Mount Analysis
4.4. 5-Bromo-2′-Deoxyuridine (BrdU) Incorporation and Immunohistochemistry (IHC)
4.5. FACS Analysis of Mammary Epithelial Cell Subpopulations with CD24 and CD49f
4.6. Western Blotting
4.7. RNA Isolation, Reverse Transcription, and Real-Time PCR
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BPA | Bisphenol A |
| DES | diethylstilbestrol |
| EDCs | Endocrine disrupting compounds |
| ER | estrogen receptor |
| GD | gestational day |
| TICs | tumor initiation cells |
| TEBs | terminal end buds |
| RTK | receptor tyrosine kinase |
| PND | postnatal day |
| MRU | mammary repopulating unit |
| MaSCs | mammary stem cells |
References
- Hilakivi-Clarke, L.; Clarke, R.; Lippman, M.E. Perinatal factors increase breast cancer risk. Breast Cancer Res. Treat. 1994, 31, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Grotmol, T.; Weiderpass, E.; Tretli, S. Conditions in utero and cancer risk. Eur. J. Epidemiol. 2006, 21, 561–570. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ekbom, A. Growing evidence that several human cancers may originate in utero. Semin. Cancer Biol. 1998, 8, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.R.; Wise, L.A.; Hatch, E.E.; Troisi, R.; Titus-Ernstoff, L.; Strohsnitter, W.; Kaufman, R.; Herbst, A.L.; Noller, K.L.; Hyer, M.; et al. Prenatal diethylstilbestrol exposure and risk of breast cancer. Cancer Epidemiol. Biomarkers Prev. 2006, 15, 1509–1514. [Google Scholar] [CrossRef]
- Matthews, J.B.; Twomey, K.; Zacharewski, T.R. In vitro and in vivo interactions of bisphenol A and its metabolite, bisphenol A glucuronide, with estrogen receptors alpha and beta. Chem. Res. Toxicol. 2001, 14, 149–157. [Google Scholar] [CrossRef]
- Washington, W.; Hubert, L.; Jones, D.; Gray, W.G. Bisphenol a binds to the low-affinity estrogen binding site. In Vitr. Mol. Toxicol. 2001, 14, 43–51. [Google Scholar] [CrossRef]
- Matsushima, A.; Kakuta, Y.; Teramoto, T.; Koshiba, T.; Liu, X.; Okada, H.; Tokunaga, T.; Kawabata, S.; Kimura, M.; Shimohigashi, Y. Structural evidence for endocrine disruptor bisphenol A binding to human nuclear receptor ERR gamma. J. Biochem. 2007, 142, 517–524. [Google Scholar] [CrossRef]
- Gertz, J.; Reddy, T.E.; Varley, K.E.; Garabedian, M.J.; Myers, R.M. Genistein and bisphenol A exposure cause estrogen receptor 1 to bind thousands of sites in a cell type-specific manner. Genome Res. 2012, 22, 2153–2162. [Google Scholar] [CrossRef]
- Thomas, P.; Dong, J. Binding and activation of the seven-transmembrane estrogen receptor GPR30 by environmental estrogens: A potential novel mechanism of endocrine disruption. J. Steroid Biochem. Mol. Biol. 2006, 102, 175–179. [Google Scholar] [CrossRef]
- Watson, C.S.; Bulayeva, N.N.; Wozniak, A.L.; Alyea, R.A. Xenoestrogens are potent activators of nongenomic estrogenic responses. Steroids 2007, 72, 124–134. [Google Scholar] [CrossRef]
- Calafat, A.M.; Kuklenyik, Z.; Reidy, J.A.; Caudill, S.P.; Ekong, J.; Needham, L.L. Urinary concentrations of bisphenol A and 4-nonylphenol in a human reference population. Environ. Health Persp. 2005, 113, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Newbold, R.R.; Jefferson, W.N.; Padilla-Banks, E. Long-term adverse effects of neonatal exposure to bisphenol A on the murine female reproductive tract. Reprod. Toxicol. 2007, 24, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Newbold, R.R.; Jefferson, W.N.; Padilla-Banks, E. Prenatal exposure to bisphenol a at environmentally relevant doses adversely affects the murine female reproductive tract later in life. Environ. Health Persp. 2009, 117, 879. [Google Scholar] [CrossRef] [PubMed]
- Vogel, S.A. The politics of plastics: The making and unmaking of bisphenol a “safety”. Am. J. Public Health 2009, 99, S559–S566. [Google Scholar] [CrossRef]
- Richter, C.A.; Birnbaum, L.S.; Farabollini, F.; Newbold, R.R.; Rubin, B.S.; Talsness, C.E.; Vandenbergh, J.G.; Walser-Kuntz, D.R.; vom Saal, F.S. In vivo effects of bisphenol A in laboratory rodent studies. Reprod. Toxicol. 2007, 24, 199–224. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Maffini, M.V.; Sonnenschein, C.; Rubin, B.S.; Soto, A.M. Bisphenol-A and the great divide: A review of controversies in the field of endocrine disruption. Endocr. Rev. 2009, 30, 75–95. [Google Scholar] [CrossRef]
- Rubin, B.S. Bisphenol A: An endocrine disruptor with widespread exposure and multiple effects. J. Steroid Biochem. 2011, 127, 27–34. [Google Scholar] [CrossRef]
- Seachrist, D.D.; Bonk, K.W.; Ho, S.M.; Prins, G.S.; Soto, A.M.; Keri, R.A. A review of the carcinogenic potential of bisphenol A. Reprod. Toxicol. 2016, 59, 167–182. [Google Scholar] [CrossRef]
- Yang, M.; Ryu, J.H.; Jeon, R.; Kang, D.; Yoo, K.Y. Effects of bisphenol A on breast cancer and its risk factors. Arch. Toxicol. 2009, 83, 281–285. [Google Scholar] [CrossRef]
- Wang, D.; Gao, H.; Bandyopadhyay, A.; Wu, A.; Yeh, I.T.; Chen, Y.; Zou, Y.; Huang, C.; Walter, C.A.; Dong, Q.; et al. Pubertal bisphenol A exposure alters murine mammary stem cell function leading to early neoplasia in regenerated glands. Cancer Prev. Res. 2014, 7, 445–455. [Google Scholar] [CrossRef]
- Dairkee, S.H.; Seok, J.; Champion, S.; Sayeed, A.; Mindrinos, M.; Xiao, W.; Davis, R.W.; Goodson, W.H. Bisphenol A induces a profile of tumor aggressiveness in high-risk cells from breast cancer patients. Cancer Res. 2008, 68, 2076–2080. [Google Scholar] [CrossRef] [PubMed]
- Goodson, W.H.; Luciani, M.G.; Sayeed, S.A.; Jaffee, I.M.; Moore, D.H.; Dairkee, S.H. Activation of the mTOR pathway by low levels of xenoestrogens in breast epithelial cells from high-risk women. Carcinogenesis 2011, 32, 1724–1733. [Google Scholar] [CrossRef] [PubMed]
- Maffini, M.V.; Rubin, B.S.; Sonnenschein, C.; Soto, A.M. Endocrine disruptors and reproductive health: The case of bisphenol-A. Mol. Cell. Endocrinol. 2006, 254–255, 79–186. [Google Scholar] [CrossRef]
- Durando, M.; Kass, L.; Piva, J.; Sonnenschein, C.; Soto, A.M.; Luque, E.H.; Muñoz-de-Toro, M. Prenatal bisphenol A exposure induces preneoplastic lesions in the mammary gland in Wistar rats. Environ. Health Perspect. 2007, 115, 80–86. [Google Scholar] [CrossRef]
- Markey, C.M.; Luque, E.H.; de Toro, M.M.; Sonnenschein, C.; Soto, A.M. In utero exposure to bisphenol A alters the development and tissue organization of the mouse mammary gland. Biol. Reprod. 2001, 65, 1215–1223. [Google Scholar] [CrossRef]
- Betancourt, A.M.; Eltoum, I.A.; Desmond, R.A.; Russo, J.; Lamartiniere, C.A. In utero exposure to bisphenol A shifts the window of susceptibility for mammary carcinogenesis in the rat. Environ. Health Perspect. 2010, 118, 1614. [Google Scholar] [CrossRef]
- Moral, R.; Wang, R.; Russo, I.H.; Lamartiniere, C.A.; Pereira, J.; Russo, J. Effect of prenatal exposure to the endocrine disruptor bisphenol A on mammary gland morphology and gene expression signature. J. Endocrinol. 2008, 196, 101–112. [Google Scholar] [CrossRef]
- Kass, L.; Altamirano, G.A.; Bosquiazzo, V.L.; Luque, E.H.; Muñoz-de-Toro, M. Perinatal exposure to xenoestrogens impairs mammary gland differentiation and modifies milk composition in Wistar rats. Reprod. Toxicol. 2012, 33, 390–400. [Google Scholar] [CrossRef]
- Hindman, A.R.; Mo, X.M.; Helber, H.L.; Kovalchin, C.E.; Ravichandran, N.; Murphy, A.R.; Fagan, A.M.; St John, P.M.; Burd, C.J. Varying Susceptibility of the Female Mammary Gland to In Utero Windows of BPA Exposure. Endocrinology 2017, 158, 3435–3447. [Google Scholar] [CrossRef]
- Acevedo, N.; Davis, B.; Schaeberle, C.M.; Sonnenschein, C.; Soto, A.M. Perinatally administered bisphenol a as a potential mammary gland carcinogen in rats. Environ. Health Perspect. 2013, 121, 1040–1046. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Yang, B.J.; Li, N.; Feng, L.M.; Shi, X.Y.; Zhao, W.H.; Liu, S.J. Bisphenol A and hormone-associated cancers: Current progress and perspectives. Medicine 2015, 94, e211. [Google Scholar] [CrossRef] [PubMed]
- Paulose, T.; Speroni, L.; Sonnenschein, C.; Soto, A.M. Estrogens in the wrong place at the wrong time: Fetal BPA exposure and mammary cancer. Reproduct. Toxicol. 2015, 54, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.; Thor, A.; He, M.; Barcos, M.; Ljung, B.; Benz, C. The HER2 (c-erbB-2) oncogene is frequently amplified in in situ carcinomas of the breast. Oncogene 1992, 7, 1027–1032. [Google Scholar] [PubMed]
- Gamett, D.C.; Pearson, G.; Cerione, R.A.; Friedberg, I. Secondary dimerization between members of the epidermal growth factor receptor family. J. Biol. Chem. 1997, 272, 12052–12056. [Google Scholar] [CrossRef] [PubMed]
- Hynes, N.E.; Lane, H.A. ERBB receptors and cancer: The complexity of targeted inhibitors. Nat. Rev. Cancer 2005, 5, 341–354. [Google Scholar] [CrossRef]
- Shen, K.; Novak, R.F. DDT stimulates c-erbB2, c-met, and STATS tyrosine phosphorylation, Grb2-Sos association, MAPK phosphorylation, and proliferation of human breast epithelial cells. Biochem. Biophys. Res. Commun. 1997, 231, 17–21. [Google Scholar] [CrossRef]
- Jin, Z.; MacDonald, R.S. Soy isoflavones increase latency of spontaneous mammary tumors in mice. J. Nutr. 2002, 132, 3186–3190. [Google Scholar] [CrossRef]
- Sacco, M.; Soldati, S.; Cato, E.M.; Cattaneo, L.; Pratesi, G.; Scanziani, E.; Verzzoni, P. Combined effects on tumor growth and metastasis by anti-estrogenic and antiangiogenic therapies in MMTV-neu mice. Gene Ther. 2002, 9, 1338–1341. [Google Scholar] [CrossRef]
- Nanni, P.; Nicoletti, G.; De Giovanni, C.; Landuzzi, L.; Di Carlo, E.; Iezzi, M.; Ricci, C.; Astolfi, A.; Croci, S.; Marangoni, F. Prevention of HER-2/neu transgenic mammary carcinoma by tamoxifen plus interleukin 12. Int. J. Cancer 2003, 105, 384–389. [Google Scholar] [CrossRef]
- Zhang, G.; Han, D.; Liu, G.; Gao, S.; Cai, X.; Duan, R.; Feng, X. Effects of soy isoflavone and endogenous oestrogen on breast cancer in MMTV-erbB2 transgenic mice. J. Int. Med. Res. 2012, 40, 2073–2082. [Google Scholar] [CrossRef]
- Munoz-de-Toro, M.; Markey, C.M.; Wadia, P.R.; Luque, E.H.; Rubin, B.S.; Sonnenschein, C.; Soto, A.M. Perinatal exposure to bisphenol-A alters peripubertal mammary gland development in mice. Endocrinol. 2005, 146, 4138–4147. [Google Scholar] [CrossRef] [PubMed]
- Boyd, N.F.; Guo, H.; Martin, L.J.; Sun, L.; Stone, J.; Fishell, E.; Jong, R.A.; Hislop, G.; Chiarelli, A.; Minkin, S. Mammographic density and the risk and detection of breast cancer. N. Engl. J. Med. 2007, 356, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Habel, L.A.; Lipson, J.A.; Achacoso, N.; Rothstein, J.H.; Yaffe, M.J.; Liang, R.Y.; Acton, L.; McGuire, V.; Whittemore, A.S.; Rubin, D.L.; et al. Case-control study of mammographic density and breast cancer risk using processed digital mammograms. Breast Cancer Res. 2016, 18, 53. [Google Scholar] [CrossRef]
- Kenney, N.J.; Smith, G.H.; Lawrence, E.; Barrett, J.C.; Salomon, D.S. Identification of Stem Cell Units in the Terminal End Bud and Duct of the Mouse Mammary Gland. J. Biomed. Biotechnol. 2001, 1, 133–143. [Google Scholar] [CrossRef]
- Farnie, G.; Clarke, R.B. Mammary stem cells and breast cancer--role of Notch signalling. Stem Cell Rev. 2007, 3, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Stingl, J. Detection and analysis of mammary gland stem cells. J. Pathol. 2009, 217, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Asselin-Labat, M.L.; Vaillant, F.; Shackleton, M.; Bouras, T.; Lindeman, G.J.; Visvader, J.E. Delineating the epithelial hierarchy in the mouse mammary gland. Cold Spring Harb. Symp. Quant. Biol. 2008, 73, 469–478. [Google Scholar] [CrossRef]
- Arpino, G.; Wiechmann, L.; Osborne, C.K.; Schiff, R. Crosstalk between the estrogen receptor and the HER tyrosine kinase receptor family: Molecular mechanism and clinical implications for endocrine therapy resistance. Endocr. Rev. 2008, 29, 217–233. [Google Scholar] [CrossRef]
- Smith, C.L. Cross-talk between peptide growth factor and estrogen receptor signaling pathways. Biol. Reprod. 1998, 58, 627–632. [Google Scholar] [CrossRef]
- Hanson, M.A.; Poston, L.; Gluckman, P.D. DOHaD—The challenge of translating the science to policy. J Dev. Orig. Health Dis. 2019, 10, 263–267. [Google Scholar] [CrossRef]
- Jenkins, S.; Wang, J.; Eltoum, I.; Desmond, R.; Lamartiniere, C.A. Chronic oral exposure to bisphenol A results in a nonmonotonic dose response in mammary carcinogenesis and metastasis in MMTV-erbB2 mice. Environ. Health Perspect. 2011, 119, 1604–1609. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Edgerton, S.M.; Kosanke, S.D.; Mason, T.L.; Alvarez, K.M.; Liu, N.; Chatterton, R.T.; Liu, B.; Wang, Q.; Kim, A.; et al. Hormonal and dietary modulation of mammary carcinogenesis in mouse mammary tumor virus-c-erbB-2 transgenic mice. Cancer Res. 2003, 63, 2425–2433. [Google Scholar] [PubMed]
- Liu, B.; Edgerton, S.; Yang, X.; Kim, A.; Ordonez-Ercan, D.; Mason, T.; Alvarez, K.; McKimmey, C.; Liu, N.; Thor, A. Low-dose dietary phytoestrogen abrogates tamoxifen-associated mammary tumor prevention. Cancer Res. 2005, 65, 879–886. [Google Scholar] [PubMed]
- Hutchinson, J.N.; Muller, W.J. Transgenic mouse models of human breast cancer. Oncogene 2000, 19, 6130–6137. [Google Scholar] [CrossRef] [PubMed]
- Mandrup, K.; Boberg, J.; Isling, L.; Christiansen, S.; Hass, U. Low-dose effects of bisphenol A on mammary gland development in rats. Andrology 2016, 4, 673–683. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, H.; Liu, S. Low-Dose Bisphenol A Exposure: A Seemingly Instigating Carcinogenic Effect on Breast Cancer. Adv. Sci. 2017, 4, 1600248. [Google Scholar] [CrossRef]
- Lozada, K.W.; Keri, R.A. Bisphenol A increases mammary cancer risk in two distinct mouse models of breast cancer. Biol. Reprod. 2011, 85, 490–497. [Google Scholar] [CrossRef]
- Ayyanan, A.; Laribi, O.; Schuepbach-Mallepell, S.; Schrick, C.; Gutierrez, M.; Tanos, T.; Lefebvre, G.; Rougemont, J.; Yalcin-Ozuysal, Ö.; Brisken, C. Perinatal exposure to bisphenol a increases adult mammary gland progesterone response and cell number. Mol. Endocrinol. 2011, 25, 1915–1923. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human exposure to bisphenol A (BPA). Reprod. Toxicol. 2007, 24, 139–177. [Google Scholar] [CrossRef]
- Nikaido, Y.; Yoshizawa, K.; Danbara, N.; Tsujita-Kyutoku, M.; Yuri, T.; Uehara, N.; Tsubura, A. Effects of maternal xenoestrogen exposure on development of the reproductive tract and mammary gland in female CD-1 mouse offspring. Reprod. Toxicol. 2004, 18, 803–811. [Google Scholar] [CrossRef]
- Hilakivi-Clarke, L.; Cho, E.; deAssis, S.; Olivo, S.; Ealley, E.; Bouker, K.B.; Welch, J.N.; Khan, G.; Clarke, R.; Cabanes, A. Maternal and prepubertal diet, mammary development and breast cancer risk. J. Nutr. 2001, 131, 154S–157S. [Google Scholar] [CrossRef] [PubMed]
- Visvader, J.E.; Stingl, J. Mammary stem cells and the differentiation hierarchy: Current status and perspectives. Genes Dev. 2014, 28, 1143–1158. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Parris, A.B.; Xiao, Z.; Howard, E.W.; Kosanke, S.D.; Feng, X.; Yang, X. Short-term early exposure to lapatinib confers lifelong protection from mammary tumor development in MMTV-erbB-2 transgenic mice. J. Exp. Clin. Cancer Res. 2017, 36, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Davis, M.; Blackwelder, A.J.; Bachman, N.; Liu, B.; Edgerton, S.; Williams, L.L.; Thor, A.D.; Yang, X. Metformin selectively targets tumor-initiating cells in ErbB2-overexpressing breast cancer models. Cancer Prev. Res. 2014, 7, 199–210. [Google Scholar] [CrossRef]
- Shelton, D.N.; Fernandez-Gonzalez, R.; Illa-Bochaca, I.; Ortiz-de-Solorzano, C.; Barcellos-Hoff, M.H.; Welm, B.E. Use of stem cell markers in dissociated mammary populations. Methods Molecul. Biol. 2010, 621, 49–55. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Day 26 | Day 27 | Day 28 | Day 29 | Day 30 | Day 31 | Vaginal Opening Day (Mean ± S.E.) | p-Value |
|---|---|---|---|---|---|---|---|---|
| Control | 1 | 4 | 8 | 2 | 29.7 ± 0.2 | |||
| BPA 50 ng/kg | 4 | 2 | 4 | 2 | 3 | 27.9 ± 0.4 | <0.01 | |
| BPA 500 ng/kg | 3 | 5 | 5 | 2 | 28.4 ± 0.3 | <0.01 | ||
| BPA 250 µg/kg | 2 | 3 | 4 | 5 | 1 | 29.0 ± 0.3 |
| Group | Days in Estrus Phase (Mean ± S.E.) | p-Value |
|---|---|---|
| Control | 5.2 ± 0.4 | |
| BPA 50 ng/kg | 7.4 ± 1.1 | <0.01 |
| BPA 500 ng/kg | 7.2 ± 0.8 | <0.01 |
| BPA 250 µg/kg | 5.0 ± 0.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Z.; Parris, A.B.; Howard, E.W.; Davis, M.; Cao, X.; Woods, C.; Yang, X. In Utero Exposure to Bisphenol a Promotes Mammary Tumor Risk in MMTV-Erbb2 Transgenic Mice Through the Induction of ER-erbB2 Crosstalk. Int. J. Mol. Sci. 2020, 21, 3095. https://doi.org/10.3390/ijms21093095
Ma Z, Parris AB, Howard EW, Davis M, Cao X, Woods C, Yang X. In Utero Exposure to Bisphenol a Promotes Mammary Tumor Risk in MMTV-Erbb2 Transgenic Mice Through the Induction of ER-erbB2 Crosstalk. International Journal of Molecular Sciences. 2020; 21(9):3095. https://doi.org/10.3390/ijms21093095
Chicago/Turabian StyleMa, Zhikun, Amanda B. Parris, Erin W. Howard, Meghan Davis, Xia Cao, Courtney Woods, and Xiaohe Yang. 2020. "In Utero Exposure to Bisphenol a Promotes Mammary Tumor Risk in MMTV-Erbb2 Transgenic Mice Through the Induction of ER-erbB2 Crosstalk" International Journal of Molecular Sciences 21, no. 9: 3095. https://doi.org/10.3390/ijms21093095
APA StyleMa, Z., Parris, A. B., Howard, E. W., Davis, M., Cao, X., Woods, C., & Yang, X. (2020). In Utero Exposure to Bisphenol a Promotes Mammary Tumor Risk in MMTV-Erbb2 Transgenic Mice Through the Induction of ER-erbB2 Crosstalk. International Journal of Molecular Sciences, 21(9), 3095. https://doi.org/10.3390/ijms21093095