Cholesterol Transport in Wild-Type NPC1 and P691S: Molecular Dynamics Simulations Reveal Changes in Dynamical Behavior


Abstract
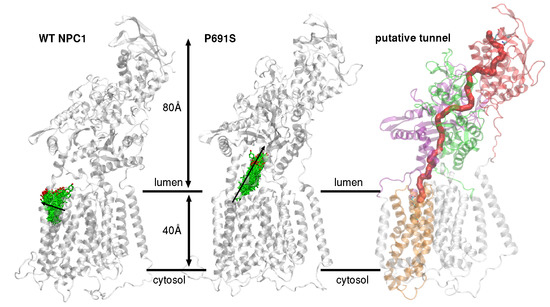
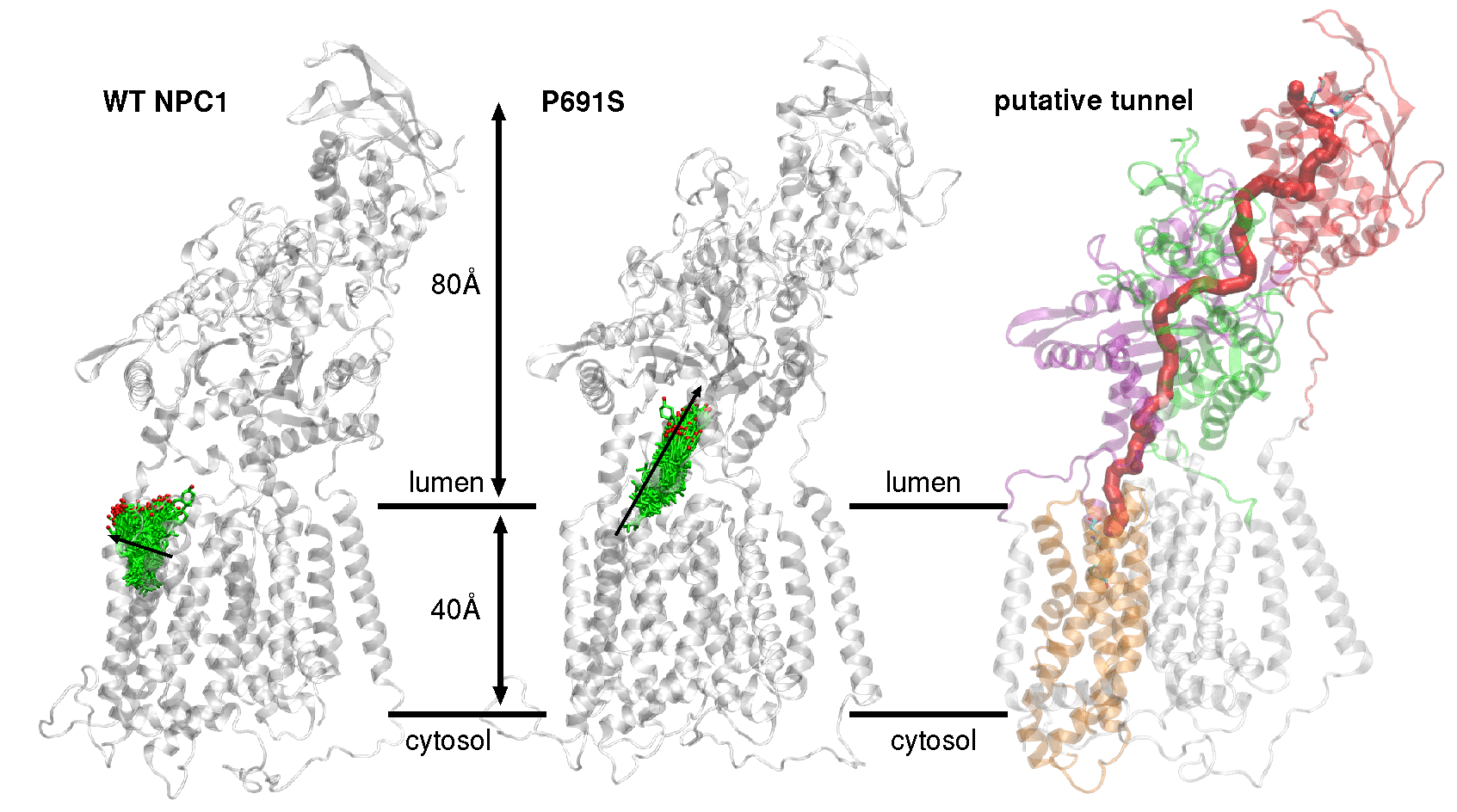
1. Introduction
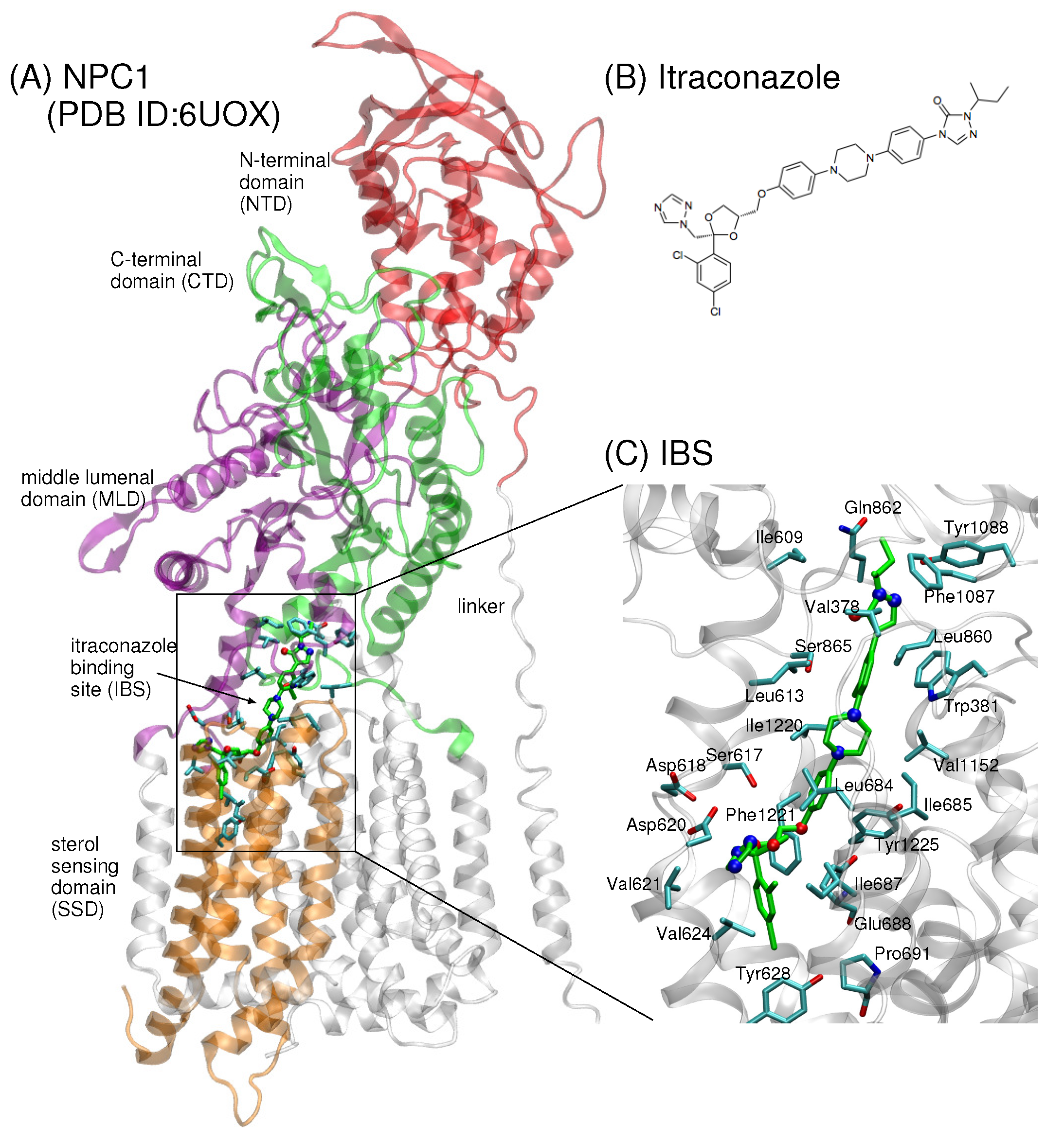
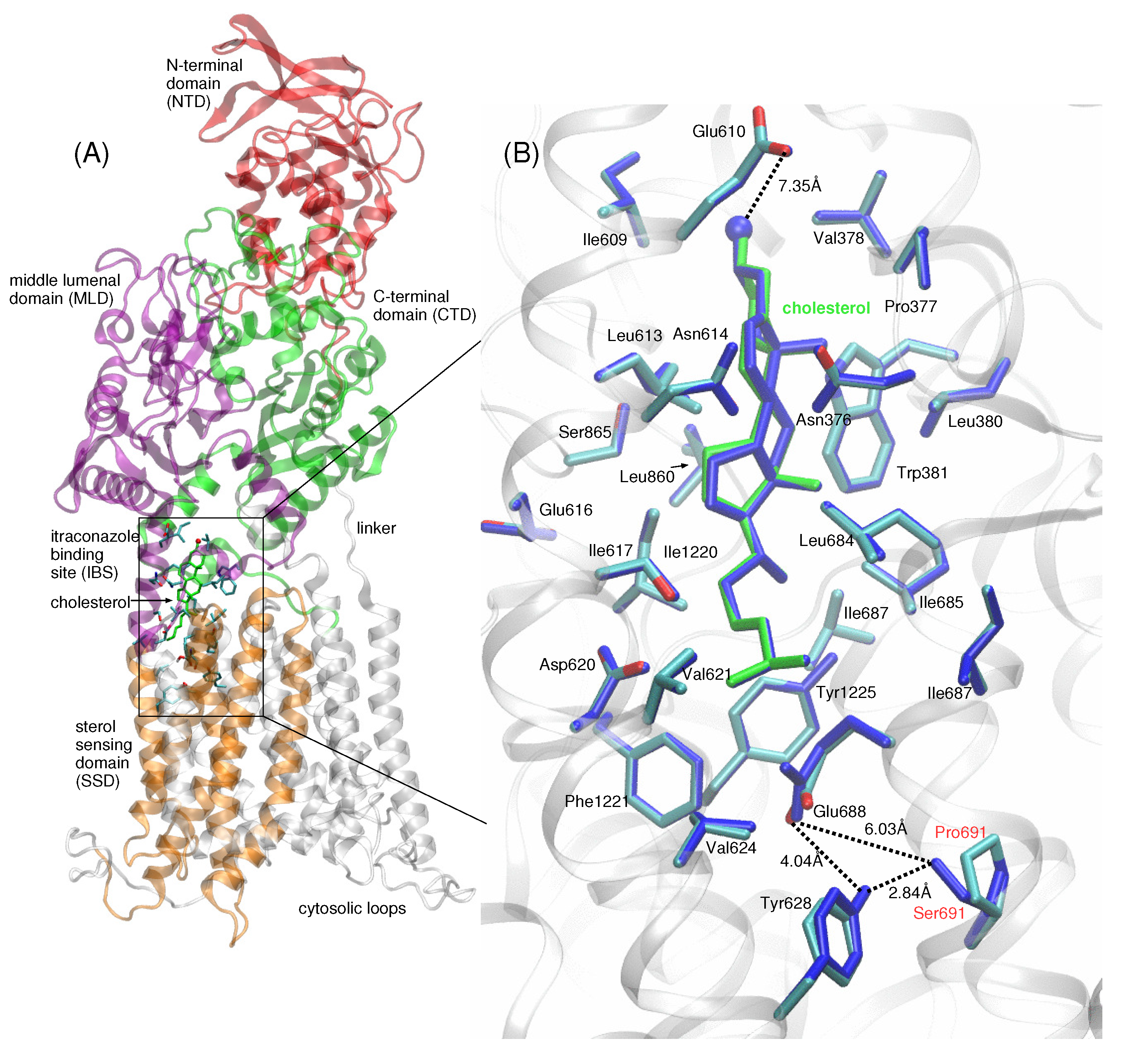
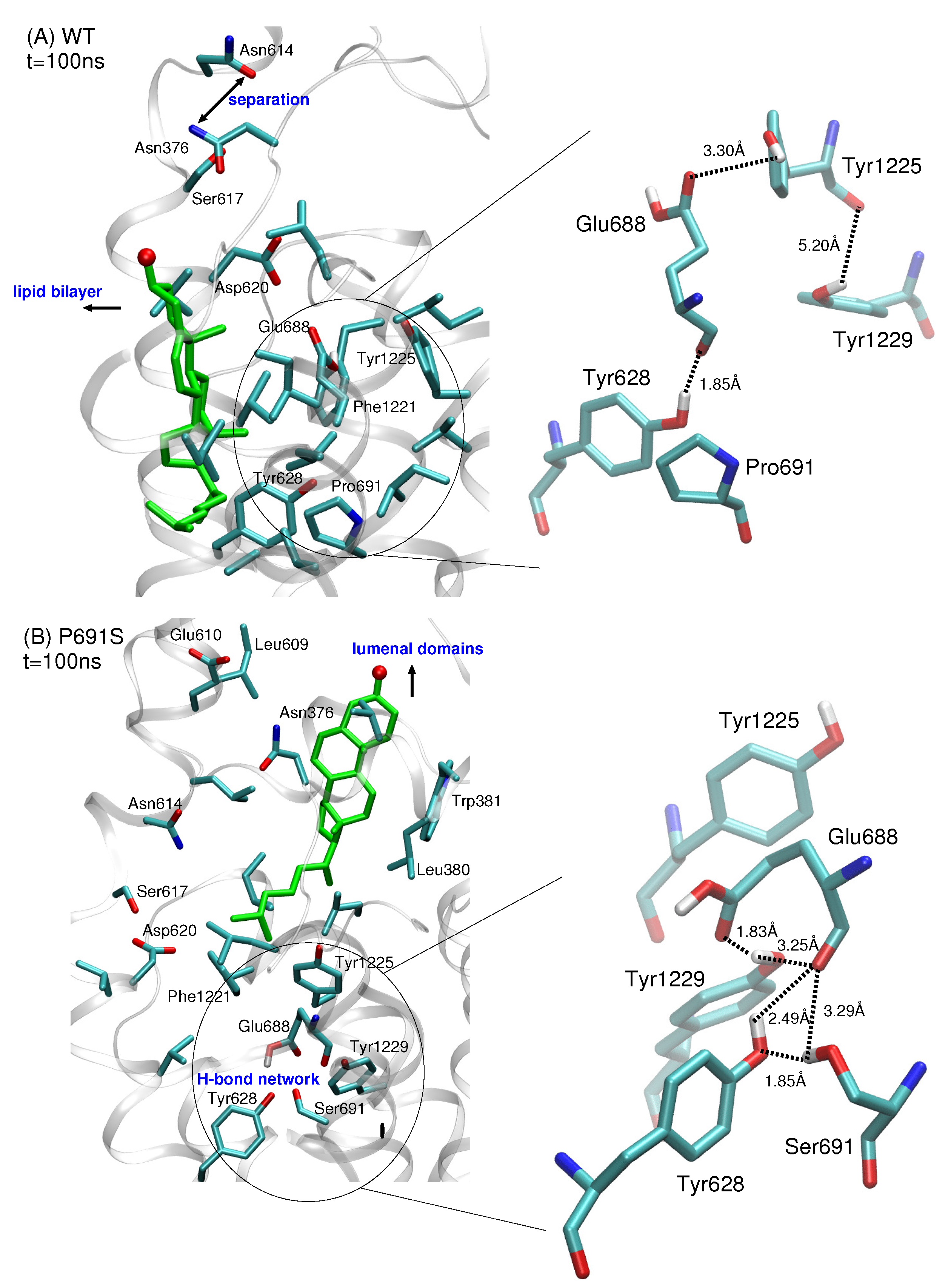
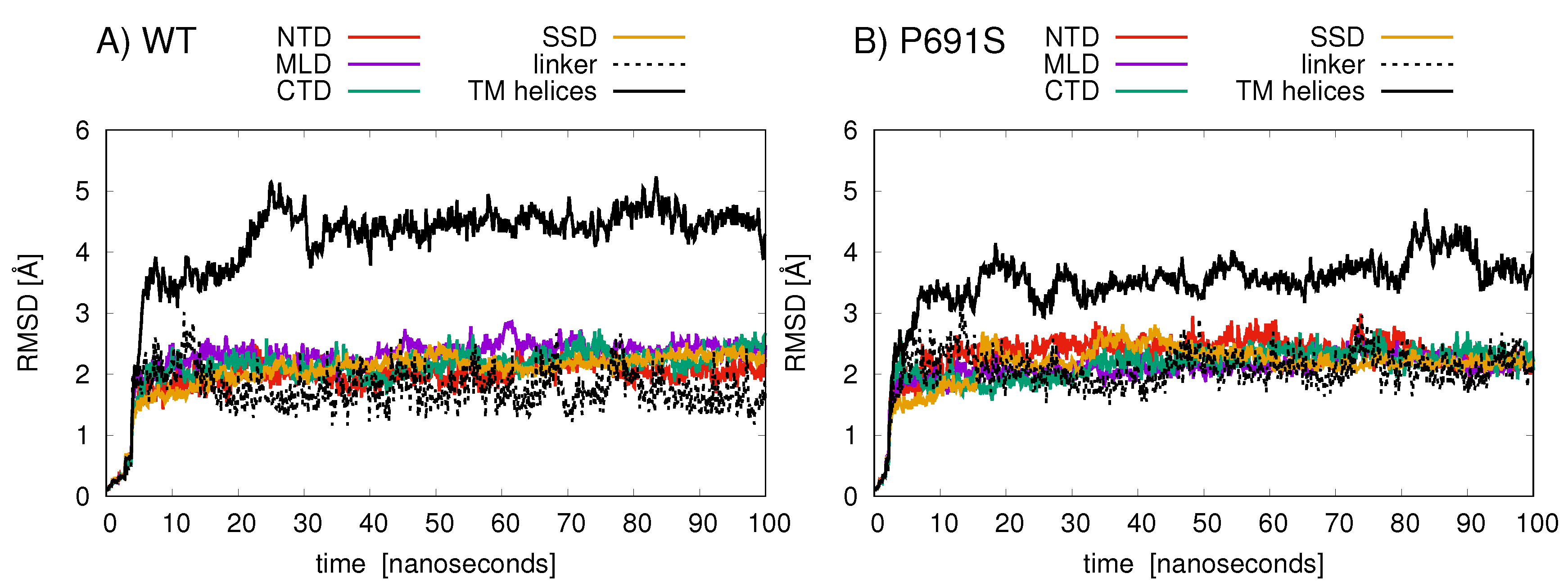
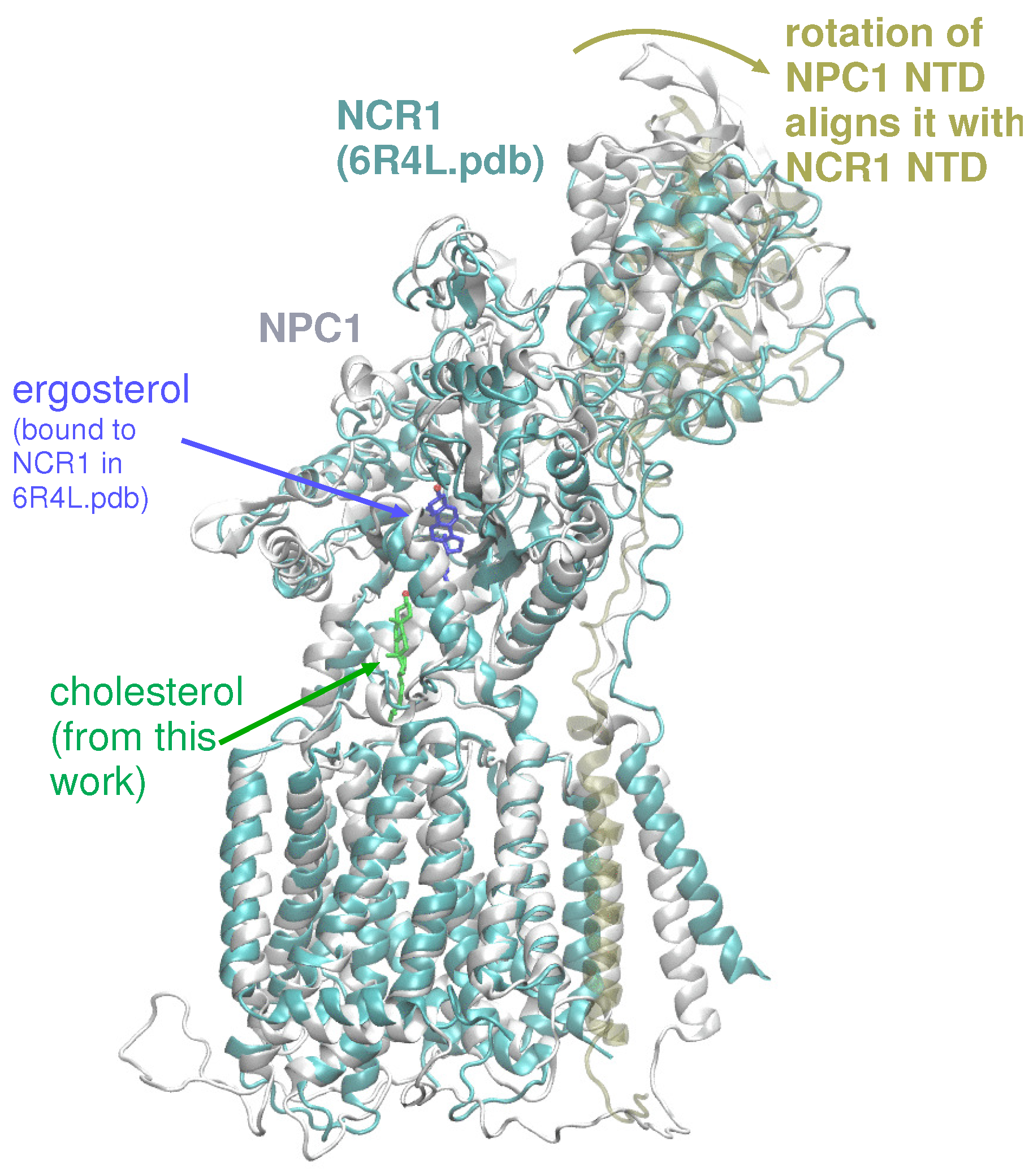
2. Results
3. Conclusions
4. Methods
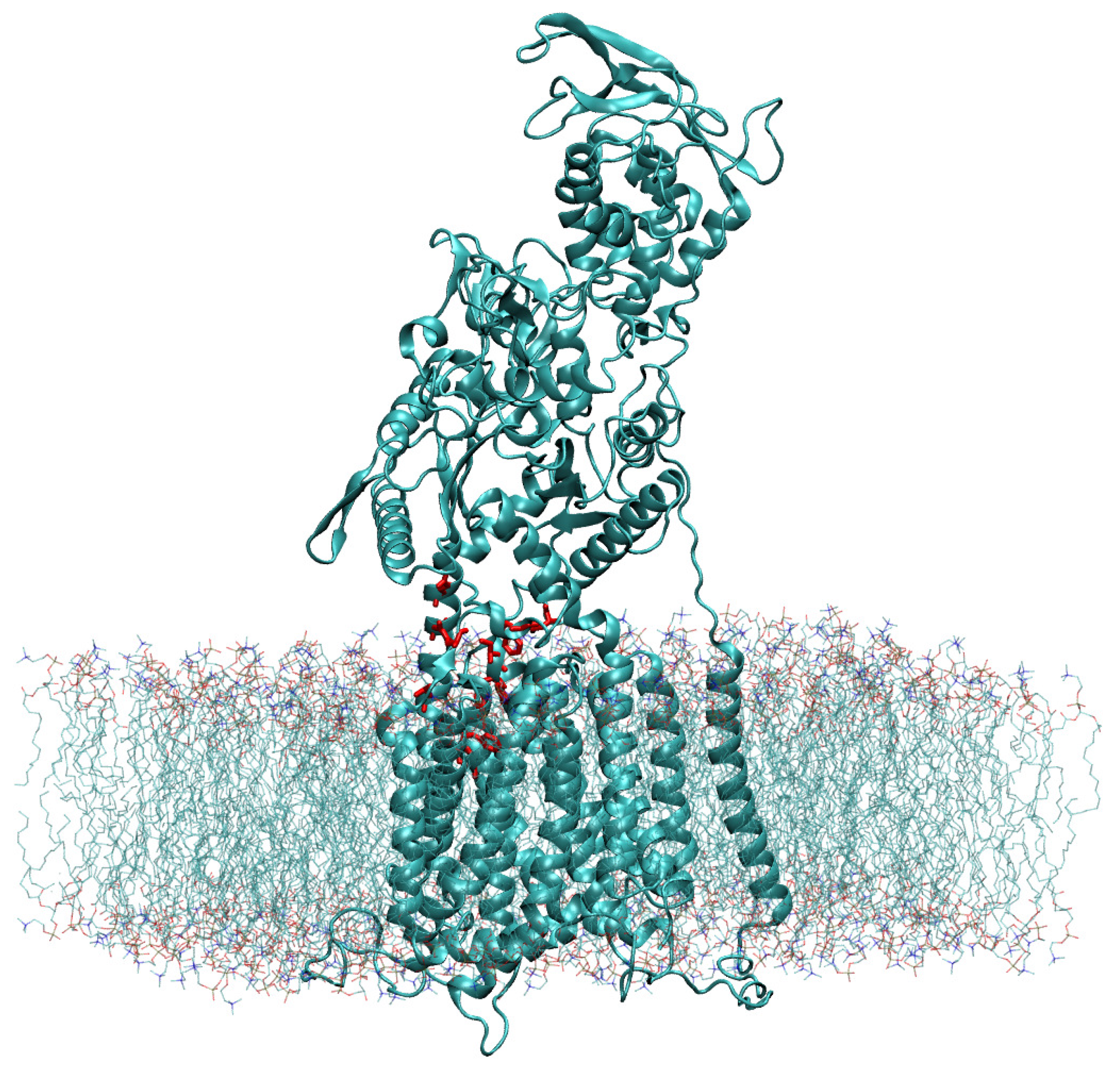
4.1. Construction of the Model
4.2. Molecular Dynamics Simulations
4.3. Calculation of Distance Correlation Coefficients
4.4. Tunnel Identification
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Vanier, M. Niemann–Pick disease type C. Orphanet Rare Dis. 2010, 5, 1–18. [Google Scholar] [CrossRef]
- Wheeler, S.; Sillence, D.J. Niemann–Pick type C disease: Cellular pathology and pharmacotherapy. J. Neurochem. 2019, 152. [Google Scholar] [CrossRef] [PubMed]
- Infante, R.E.; Wang, M.L.; Radhakrishnan, A.; Kwon, H.J.; Brown, M.S.; Goldstein, J.L. NPC2 facilitates bidirectional transfer of cholesterol between NPC1 and lipid bilayers, a step in cholesterol egress from lysosomes. Proc. Natl. Acad. Sci. USA 2008, 105, 15287–15292. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.J.; Abi-Mosleh, L.; Wang, M.L.; Deisenhofer, J.; Goldstein, J.L.; Brown, M.S.; Infante, R.E. Structure of N-terminal domain of NPC1 reveals distinct subdomains for binding and transfer of cholesterol. Cell 2009, 137, 1213–1224. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Saha, P.; Li, J.; Blobel, G.; Pfeffer, S.R. Clues to the mechanism of choelsterol transfer from the structure of NPC1 middle lumenal domain bound to NPC2. Proc. Natl. Acad. Sci. USA 2016, 113, 10079–10084. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Qiang, H.W.; Zhou, X.H.; Wu, J.P.; Zhou, Q.; Yan, N. Structural insights into the Niemann–Pick C1 protein(NPC1) mediated cholesterol transfer and Ebola infection. Cell 2016, 165, 1467–1478. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lu, F.; Trinh, M.N.; Schmiege, P.; Seemann, J.; Wang, J.; Blobel, G. 3.3 Å structure of Niemann–Pick C1 protein reveals insights into the function of the C-terminal luminal domain in cholesterol transport. Proc. Natl. Acad. Sci. USA 2017, 114, 9116–9121. [Google Scholar] [CrossRef]
- Wang, C.; Scott, S.M.; Subramanian, K.; Loguercio, S.; Zhao, P.; Hutt, D.M.; Farhat, N.Y.; Porter, F.D.; Balch, W.E. Quantitating the epigenetic transformation contributing to cholesterol homeostasis using Gaussian process. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Millat, G.; Marcais, C.; Tomasetto, C.; Chikh, K.; Fensom, A.H.; Harzer, K.; Wenger, D.A.; Ohno, K.; Vanier, M.T. Niemann–Pick C1 disease: Correlations between NPC1 mutations, levels of NPC1 protein, and phenotypes emphasize the functional significance of the putative sterol-sensing domain and of the cysteine-rich luminal loop. Am. J. Hum. Genet. 2001, 68, 1373–1385. [Google Scholar] [CrossRef]
- Shammas, H.; Kuech, E.M.; Rizk, S.; Das, A.M.; Naim, H.Y. Genotypes Generate Protein Phenotypes that Vary in their Intracellular Processing, Trafficking and Localization. Sci. Rep. 2019, 9, 5292–5304. [Google Scholar] [CrossRef]
- Ohgami, N.; Ko, D.C.; Thomas, M.; Scott, M.P.; Chang, C.C.Y.; Chang, T.Y. Binding between the Niemann–Pick C1 protein and a photoactivatable cholesterol analog requires a functional sterol-sensing domain. Proc. Natl. Acad. Sci. USA 2004, 101, 12473–12478. [Google Scholar] [CrossRef]
- Lu, F.; Liang, Q.L.; Abi-Mosleh, A.D.; Brabander, J.K.D.; Goldstein, J.L.; Brown, M.S. Identification of NPC1 as the target of U18666A, an inhibitor of lysosomal cholesterol export and Ebola infection. eLife 2015, 4, e12177. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.B.L.; Kidmose, R.T.; Szomek, M.; Thaysen, K.; Rawson, S.; Muench, S.P.; Wüstner, D.; Panyella Pedersen, B. Structural insight into eukaryotic sterol transport through Niemann–Pick Type C proteins. Cell 2019, 179, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.P.; Ioannou, Y.A. Topological analysis of Niemann–Pick C1 protein reveals that the membrane orientation of the putative sterol-sending domain is identical to those of 3-hydroxy-3-methylglutaryl-CoA reductase and sterol regulatory element binding protein cleavage-activating protein. J. Biol. Chem. 2000, 275, 24367–24373. [Google Scholar] [PubMed]
- Tseng, T.T.; Gratwick, K.S.; Kollman, J.; Park, D.; Nies, D.H.; Goffeau, A.; Saier, M.H., Jr. The RND permease superfamily: An ancient, ubiquitous and diverse family that includes human disease and development proteins. J. Mol. Microbiol. Biotechnol. 1999, 1, 107–125. [Google Scholar] [PubMed]
- Qi, X.; Schmiege, P.; Coutavas, E.; Wang, J.; Li, X. Structures of human Patched and its complex with native palmitoylated sonic hedgehog. Nature 2018, 560, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Qiang, H.W.; Cao, P.; Zhou, X.H.; Zhou, Q.; Lei, J.; Yan, N. Structural basis for the recognition of Sonic Hedgehog by human Patched1. Science 2018, 568, 1–11. [Google Scholar] [CrossRef]
- Long, T.; Qi, X.; Hassan, A.; Liang, Q.; De Brabander, J.K.; Li, X. Structural basis for itraconazole-mediated NPC1 inhibition. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Trinh, M.N.; Lu, F.; Li, X.; Das, A.; Liang, Q.; De Brabander, J.K.; Brown, M.S.; Goldstein, J.L. Triazoles inhibit cholesterol export from lysosomes by binding to NPC1. Proc. Natl. Acad. Sci. USA 2017, 114, 89–94. [Google Scholar] [CrossRef]
- Elghobashi-Meinhardt, N. Computational tools unravel putative sterol binding sites in the lysosomal NPC1 protein. JCIM 2019, 59, 2432–2441. [Google Scholar] [CrossRef]
- Qi, X.; Schmiege, P.; Coutavas, E.; Li, X. Two Patched molecules engage distinct sites on Hedgehog yielding a signaling-competent complex. Science 2018, 362, eaas8843. [Google Scholar] [CrossRef] [PubMed]
- Watari, H.; Blanchette-Mackie, E.J.; Dwyer, N.K.; Watari, M.; Neufeld, E.B.; Patel, S.; Pentchev, P.G.; Strauss, J.F., III. Mutations in the leucine zipper motif and sterol-sensing domain inactivate the Niemann–Pick C1 glycoprotein. J. Biol. Chem. 1999, 274, 21861–21866. [Google Scholar] [CrossRef] [PubMed]
- Millard, E.E.; Gale, S.E.; Dudley, N.; Zhang, J.; Schaffer, J.E.; Ory, D.S. The sterol-sensing domain of the Niemann–Pick C1 (NPC1) protein regulates trafficking of low density lipoprotein cholesterol. J. Biol. Chem. 2005, 280, 28581–28590. [Google Scholar] [CrossRef] [PubMed]
- Berka, K.; Hanák, O.; Sehnal, D.; Banáš, P.; Navátilová, V.; Jaiswal, D.; Ionescu, C.M.; Svobodová Vařekova, R.; Koča, J.; Otyepka, M. MOLEonline 2.0: Interactive web-based analysis of biomacromolecular channels. Nucleic Acids Res. 2012, 40, W222–W227. [Google Scholar] [CrossRef]
- Trinh, M.N.; Brown, M.S.; Seemann, J.; Goldstein, J.L.; Lu, F. Lysosomal cholesterol export reconstituted from fragments of Niemann–Pick C1. eLife 2018, 7, e38564. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Needleman, S.B.; Wunsch, C.D. A general method applicable to the search for similarities in the amino acid sequence of two proteins. J. Mol. Biol. 1970, 48, 443–453. [Google Scholar] [CrossRef]
- Brooks, B.R.; Bruccoleri, R.E.; Olafson, B.D.; States, D.J.; Swaminathan, S.; Karplus, M. CHARMM: A program for macromolecular energy, minimization, and dynamics calculations. J. Comput. Chem. 1983, 4, 187–217. [Google Scholar] [CrossRef]
- Kieseritzky, G.; Knapp, E.W. Optimizing pKa computation in proteins wtih pH adapted conformations. Proteins 2008, 71, 1335–1348. [Google Scholar] [CrossRef]
- Rabenstein, B.; Knapp, E.W. Calculated pH-Dependent Population and Protonation of Carbon-Monoxy- Myoglobin Conformers. Biophys. J. 2001, 80, 1141–1150. [Google Scholar] [CrossRef]
- Meyer, T.; Knapp, E.W. pK(a) Values in Proteins Determined by Electrostatics Applied to Molecular Dynamics Trajectories. J. Chem. Theory Comput. 2015, 11, 2827–2840. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Tim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Kim, T.; Im, W. Automated builder and database of protein/membrane complexes for molecular dynamics simulations. PLoS ONE 2007, 2, e880. [Google Scholar] [CrossRef] [PubMed]
- Lomize, M.A.; Lomize, A.L.; Pogozheva, I.D.; Mosberg, H.I. OPM: Orientations of proteins in membranes database. Bioinformatics 2006, 22, 623–625. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, W.; Chandrasekhar, J.; Madura, J.; Impey, R.; Klein, M. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Hodošček, M.; Elghobashi-Meinhardt, N. Simulations of NPC1(NTD):NPC2 protein complex reveal cholesterol transfer pathways. Int. J. Mol. Sci. 2018, 19, 2623. [Google Scholar] [CrossRef]
- MacKerell, A.D., Jr.; Bashford, D.; Bellott, M.L.; Dunbrack, R.L., Jr.; Evanseck, J.D.; Field, M.J.; Fischer, S.; Gao, J.; Guo, H.; Ha, S.; et al. All-atom empirical potential for molecular modeling and dynamics studies of proteins. J. Phys. Chem. B 1998, 102, 3586–3616. [Google Scholar] [CrossRef]
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kalé, L.; Schulten, K. Scalable molecular dynamics with NAMD. J. Comp. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A smooth particle mesh Ewald method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef]
- Roy, A.; Post, C.B. Detection of long-range concerted motions in protein by a distance covariance. J. Chem. Theory Comput. 2012, 8, 3009–3014. [Google Scholar] [CrossRef]
- Roy, A.; Hua, D.P.; Post, C.B. Analysis of multidomain protein dynamics. J. Chem. Theory Comput. 2016, 12, 274–280. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NTD | MLD | CTD | TMD Helices | Linker | Cyt Loops | |
|---|---|---|---|---|---|---|
| (A) WT | ||||||
| NTD | 1.00 | 0.901 | 0.935 | 0.906 | 0.643 | 0.742 |
| MLD | 0.901 | 1.000 | 0.797 | 0.897 | 0.610 | 0.592 |
| CTD | 0.935 | 0.797 | 1.000 | 0.890 | 0.628 | 0.766 |
| TMD | 0.906 | 0.897 | 0.890 | 1.000 | 0.628 | 0.633 |
| linker | 0.643 | 0.610 | 0.628 | 0.628 | 1.000 | 0.470 |
| Cyt loops | 0.742 | 0.592 | 0.766 | 0.633 | 0.470 | 1.000 |
| (B) P691S | ||||||
| NTD | 1.00 | 0.912 | 0.905 | 0.801 | 0.653 | 0.623 |
| MLD | 0.912 | 1.000 | 0.805 | 0.810 | 0.614 | 0.531 |
| CTD | 0.905 | 0.805 | 1.000 | 0.853 | 0.654 | 0.557 |
| TMD | 0.801 | 0.801 | 0.853 | 1.000 | 0.564 | 0.454 |
| linker | 0.653 | 0.614 | 0.654 | 0.564 | 1.000 | 0.514 |
| Cyt loops | 0.623 | 0.531 | 0.557 | 0.454 | 0.514 | 1.000 |
| (C) | ||||||
| NTD | 0.000 | 0.011 | 0.030 | 0.105 | 0.010 | 0.119 |
| MLD | 0.011 | 0.000 | 0.008 | 0.087 | 0.004 | 0.061 |
| CTD | 0.030 | 0.008 | 0.000 | 0.037 | 0.026 | 0.209 |
| TMD | 0.105 | 0.087 | 0.037 | 0.000 | 0.064 | 0.179 |
| linker | 0.010 | 0.004 | 0.026 | 0.064 | 0.000 | 0.044 |
| Cyt loops | 0.119 | 0.061 | 0.209 | 0.179 | 0.044 | 0.000 |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elghobashi-Meinhardt, N. Cholesterol Transport in Wild-Type NPC1 and P691S: Molecular Dynamics Simulations Reveal Changes in Dynamical Behavior. Int. J. Mol. Sci. 2020, 21, 2962. https://doi.org/10.3390/ijms21082962
Elghobashi-Meinhardt N. Cholesterol Transport in Wild-Type NPC1 and P691S: Molecular Dynamics Simulations Reveal Changes in Dynamical Behavior. International Journal of Molecular Sciences. 2020; 21(8):2962. https://doi.org/10.3390/ijms21082962
Chicago/Turabian StyleElghobashi-Meinhardt, Nadia. 2020. "Cholesterol Transport in Wild-Type NPC1 and P691S: Molecular Dynamics Simulations Reveal Changes in Dynamical Behavior" International Journal of Molecular Sciences 21, no. 8: 2962. https://doi.org/10.3390/ijms21082962
APA StyleElghobashi-Meinhardt, N. (2020). Cholesterol Transport in Wild-Type NPC1 and P691S: Molecular Dynamics Simulations Reveal Changes in Dynamical Behavior. International Journal of Molecular Sciences, 21(8), 2962. https://doi.org/10.3390/ijms21082962