Underlying Ossification Phenotype in a Murine Model of Metastatic Synovial Sarcoma




Abstract
1. Introduction
2. Results
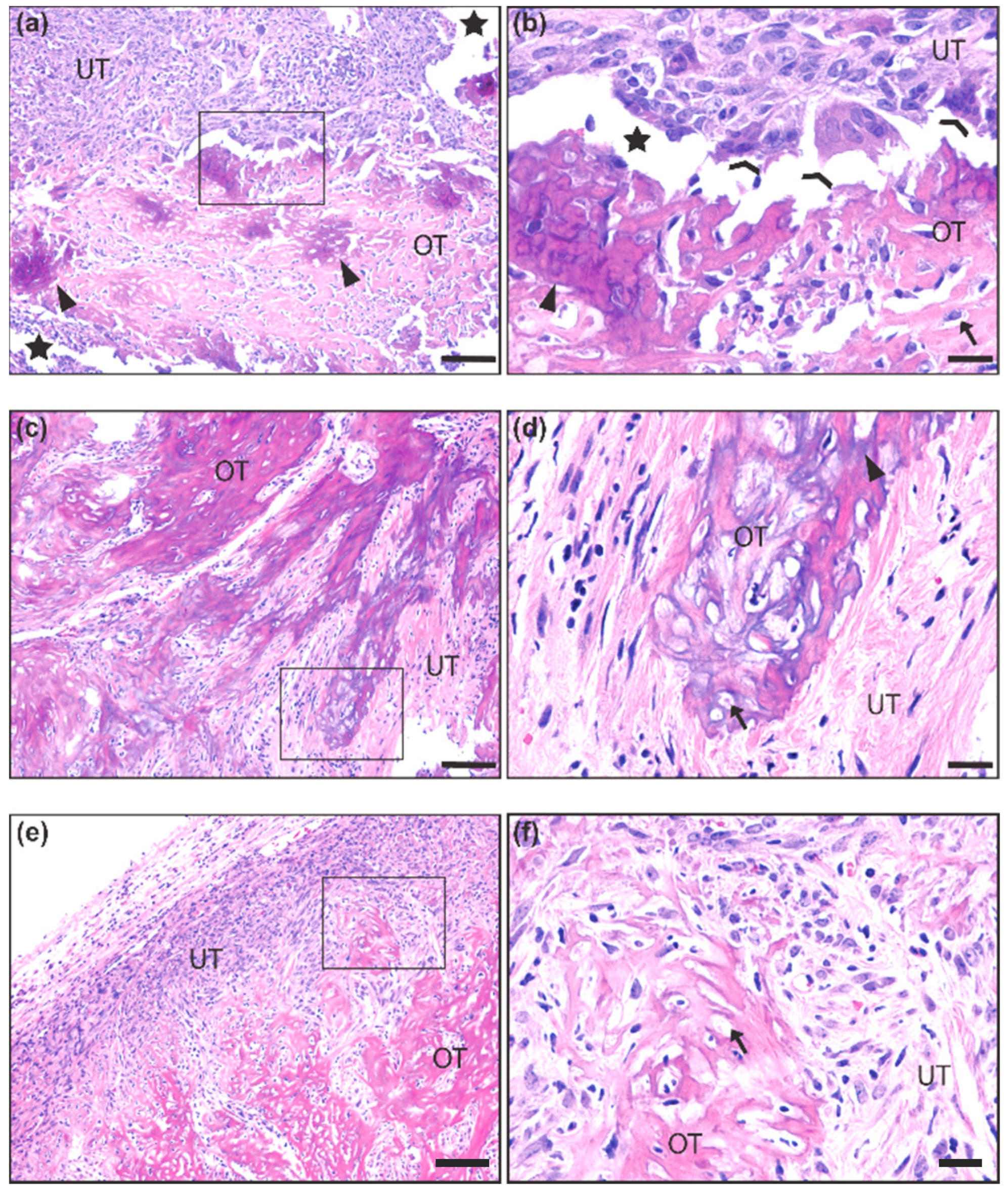
2.1. Gross and Histological Analysis of Mouse Synovial Sarcoma Demonstrates Overt Ossification in a Minority of Cases
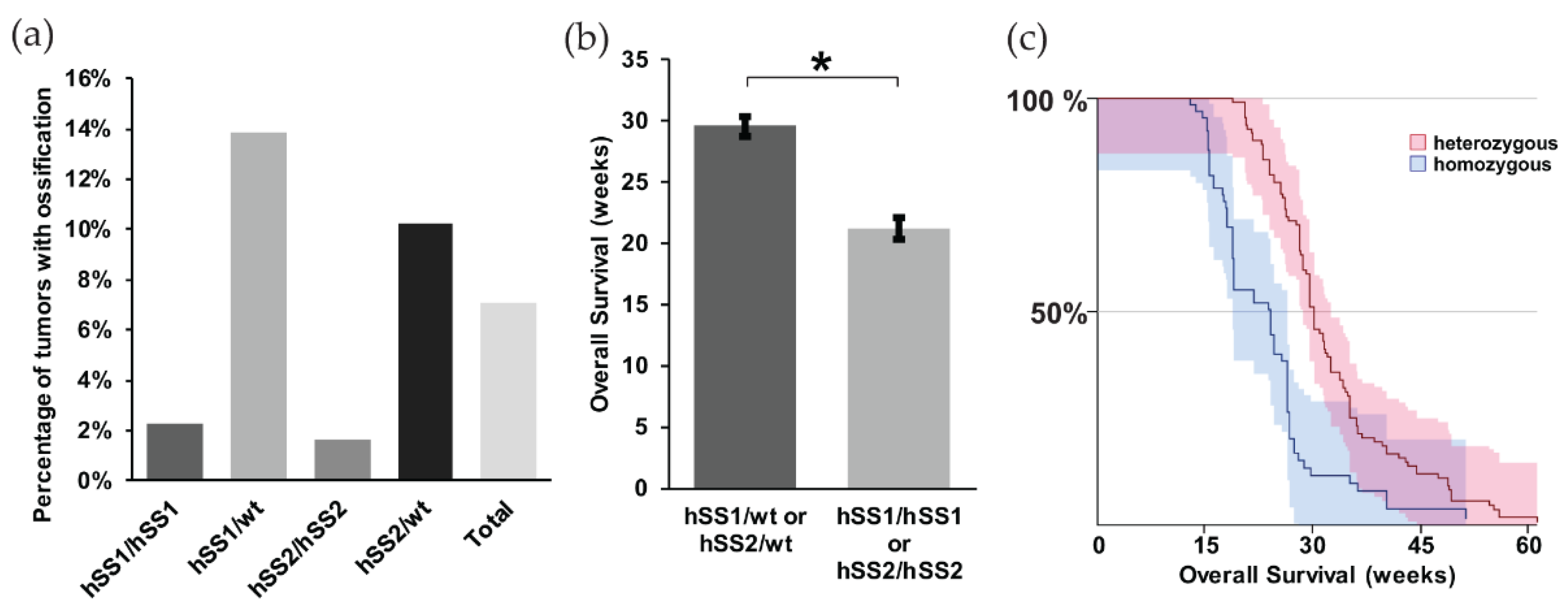
2.2. Differences in the Ossification Phenotype between Heterozygous and Homozygous Fusion Gene Expression
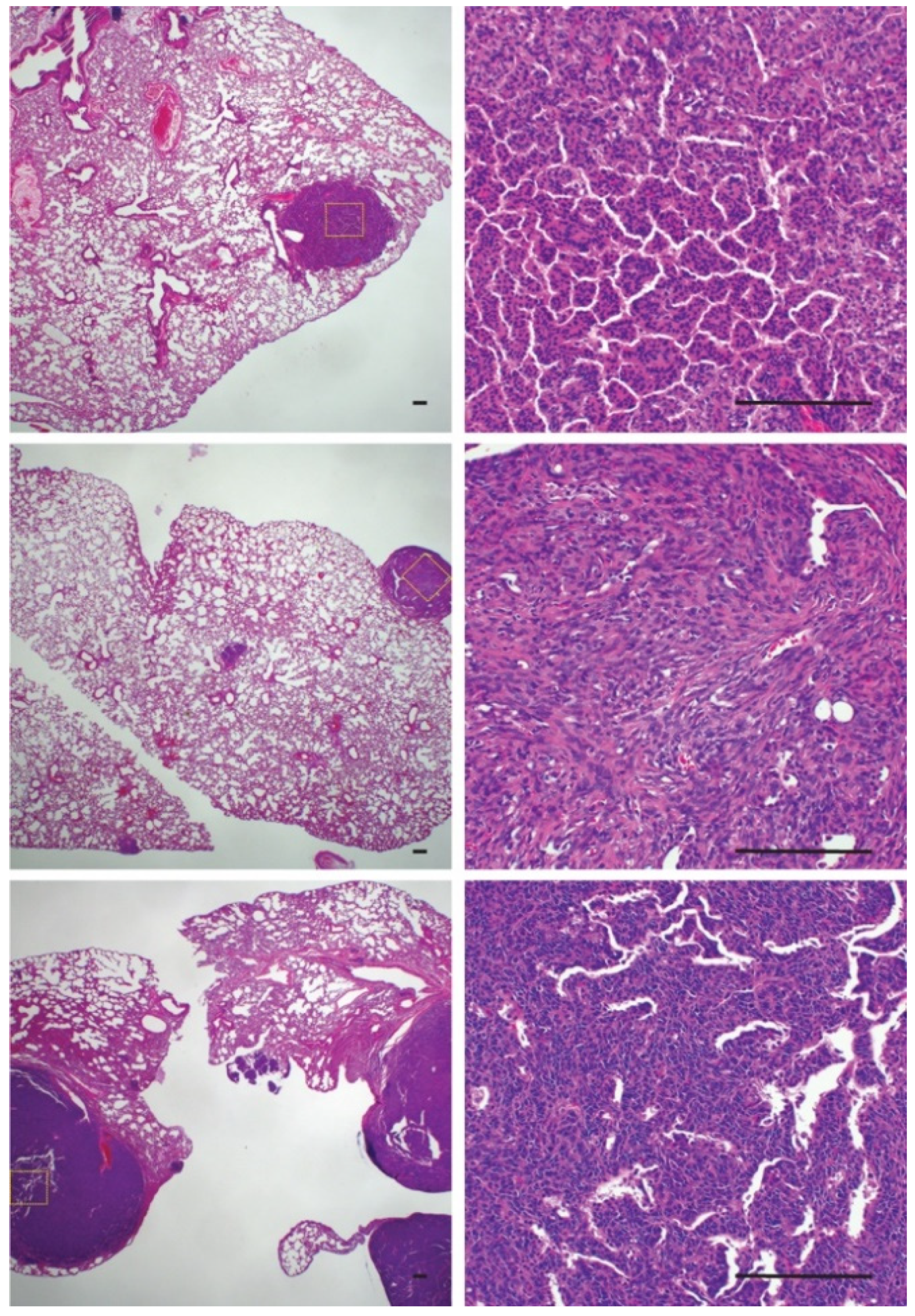
2.3. Bone Reactivity Detected by Radiography at a Higher Incidence than Histological Ossification
2.4. Underlying Gene Expression of Bone Development Genes in Metastatic Model of Synovial Sarcoma
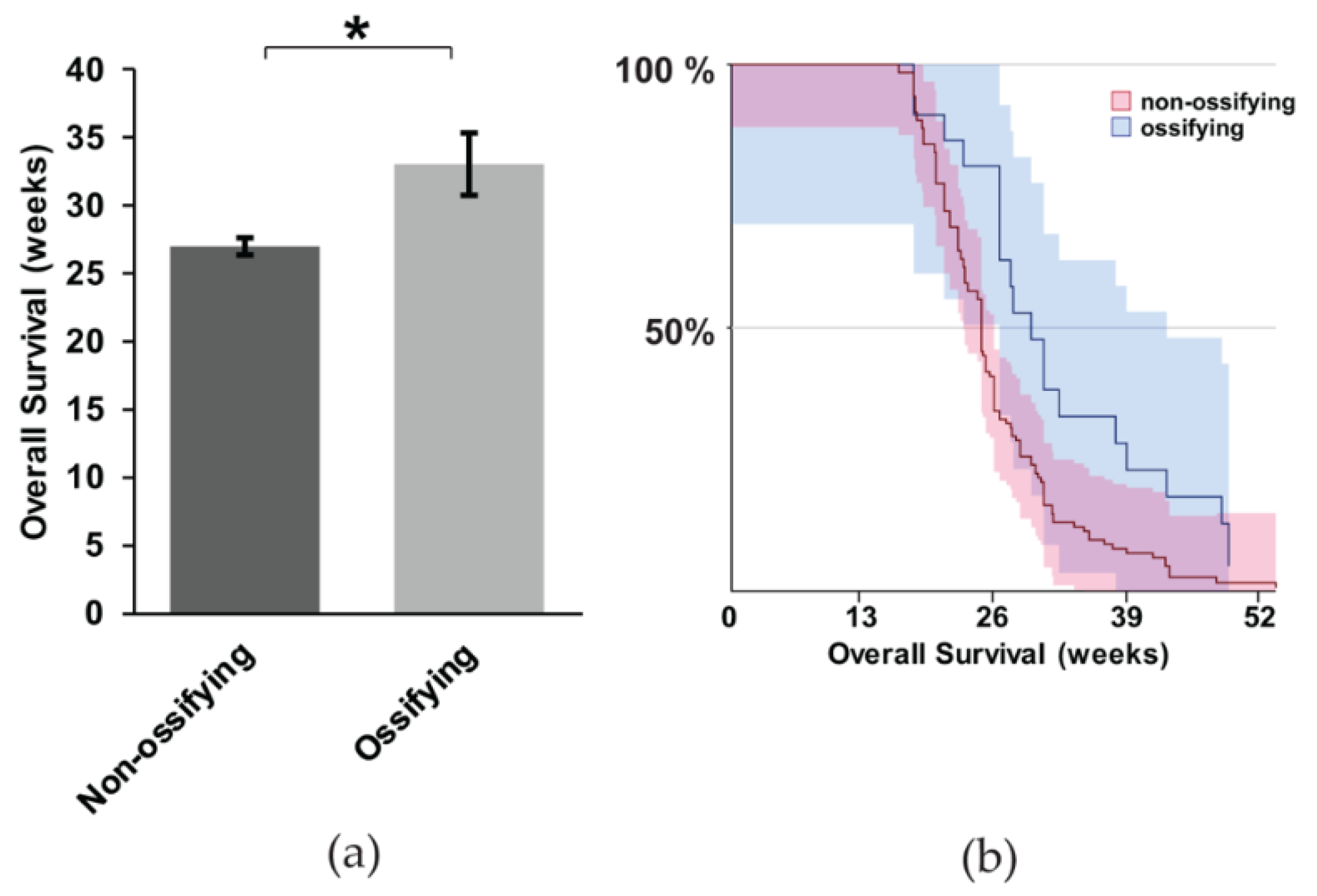
2.5. Gross Metastasis is Loosely Correlated with the Presence of Ossification, but Survival is More Favorable in Mice with Ossification
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Histology
4.3. Transcriptome Analyses
4.4. Statistics and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SS | Synovial Sarcoma |
| OPG | Osteoprotegerin |
| Pten | Phosphatase and tensin homolog |
| HO | Heterotopic Ossification |
| CEBPB | CCAAT/enhancer binding protein |
| BMP6 | Bone morphogenic protein 6 |
| PTGS2 | Prostaglandin-endoperoxide synthase 2 |
| DMP1 | Dentin matrix protein 1 |
| FGF2 | Fibroblast growth factor 2 |
| H2AFV | H2A.Z Variant Histone 2 |
| PTHLH | Parathyroid hormone-like hormone |
| MDSC | Myeloid derived suppressor cells |
| RUNX2 | Runt-related transcription factor 2 |
| Tyrobp | Transmembrane Immune Signaling Adaptor |
| Ifng | Interferon gamma |
| Il17 | Interleukin 17 |
| Tgfb | Transforming growth factor beta |
| PI3K | Phosphoinositide 3-kinase |
| AKT | Serine/Threonine Kinase |
| VEGF | Vascular endothelial growth factor |
| TGF-a | Transforming growth factor alpha |
| IHH | Indian Hedgehog |
| PTH1R | Parathyroid hormone 1 receptor |
| CI | Confidence interval |
| FPKM | Fragments Per Kilobase of transcript per Million mapped reads |
References
- Deshmukh, R.; Mankin, H.J.; Singer, S. Synovial sarcoma: The importance of size and location for survival. Clin. Orthop. Relat. Res. 2004, 419, 155–161. [Google Scholar] [CrossRef]
- Haldar, M.; Hancock, J.D.; Coffin, C.M.; Lessnick, S.L.; Capecchi, M.R. A conditional mouse model of synovial sarcoma: Insights into a myogenic origin. Cancer Cell 2007, 11, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Herzog, C.E. Overview of sarcomas in the adolescent and young adult population. J. Pediatr. Hematol. Oncol. 2005, 27, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Barrott, J.J.; Illum, B.E.; Jin, H.; Hedberg, M.L.; Wang, Y.; Grossmann, A.; Haldar, M.; Capecchi, M.R.; Jones, K.B. Paracrine osteoprotegerin and beta-catenin stabilization support synovial sarcomagenesis in periosteal cells. J. Clin. Investig. 2018, 128, 207–218. [Google Scholar] [CrossRef]
- Hara, S.; Hatori, M.; Hosaka, M.; Komatsu, T.; Tsuchiya, T.; Kimura, N. Synovial sarcoma with massive ossification—A case report. Ups. J. Med. Sci. 2003, 108, 151–158. [Google Scholar] [CrossRef]
- Hisaoka, M.; Matsuyama, A.; Shimajiri, S.; Akiba, J.; Kusano, H.; Hiraoka, K.; Shoda, T.; Hashimoto, H. Ossifying synovial sarcoma. Pathol. Res. Pract. 2009, 205, 195–198. [Google Scholar] [CrossRef]
- Milchgrub, S.; Ghandur-Mnaymneh, L.; Dorfman, H.D.; Albores-Saavedra, J. Synovial sarcoma with extensive osteoid and bone formation. Am. J. Surg. Pathol. 1993, 17, 357–363. [Google Scholar] [CrossRef]
- Ud Din, N.; Hopkins, A.; Memon, A.; Ahmad, Z.; Ahmed, R. Calcifying/ossifying synovial sarcoma: A clinicopathologic and molecular study of 5 cases. Indian J. Pathol. Microbiol. 2015, 58, 55–58. [Google Scholar] [CrossRef]
- Winnepenninckx, V.; De Vos, R.; Debiec-Rychter, M.; Samson, I.; Brys, P.; Hagemeijer, A.; Sciot, R. Calcifying/ossifying synovial sarcoma shows t(X;18) with SSX2 involvement and mitochondrial calcifications. Histopathology 2001, 38, 141–145. [Google Scholar] [CrossRef]
- Varela-Duran, J.; Enzinger, F.M. Calcifying synovial sarcoma. Cancer 1982, 50, 345–352. [Google Scholar] [CrossRef]
- Barrott, J.J.; Illum, B.E.; Jin, H.; Zhu, J.F.; Mosbruger, T.; Monument, M.J.; Smith-Fry, K.; Cable, M.G.; Wang, Y.; Grossmann, A.H.; et al. beta-catenin stabilization enhances SS18-SSX2-driven synovial sarcomagenesis and blocks the mesenchymal to epithelial transition. Oncotarget 2015, 6, 22758–22766. [Google Scholar] [CrossRef] [PubMed]
- Barrott, J.J.; Kafchinski, L.A.; Jin, H.; Potter, J.W.; Kannan, S.D.; Kennedy, R.; Mosbruger, T.; Wang, W.L.; Tsai, J.W.; Araujo, D.M.; et al. Modeling synovial sarcoma metastasis in the mouse: PI3’-lipid signaling and inflammation. J. Exp. Med. 2016, 213, 2989–3005. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.B.; Barrott, J.J.; Xie, M.; Haldar, M.; Jin, H.; Zhu, J.F.; Monument, M.J.; Mosbruger, T.L.; Langer, E.M.; Randall, R.L.; et al. The impact of chromosomal translocation locus and fusion oncogene coding sequence in synovial sarcomagenesis. Oncogene 2016, 35, 5021–5032. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Barrott, J.J.; Cable, M.G.; Monument, M.J.; Lerman, D.M.; Smith-Fry, K.; Nollner, D.; Jones, K.B. The Impact of Microenvironment on the Synovial Sarcoma Transcriptome. Cancer Microenviron. 2017. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.M.; Lin, Y.F.; Su, C.Y.; Peng, H.Y.; Chang, Y.C.; Hsiao, J.R.; Chen, C.L.; Chang, J.Y.; Shieh, Y.S.; Hsiao, M.; et al. Parathyroid Hormone-Like Hormone is a Poor Prognosis Marker of Head and Neck Cancer and Promotes Cell Growth via RUNX2 Regulation. Sci. Rep. 2017, 7, 41131. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, V.; Mula, R.V.; Weigel, N.L.; Falzon, M. Parathyroid hormone-related protein regulates cell survival pathways via integrin alpha6beta4-mediated activation of phosphatidylinositol 3-kinase/Akt signaling. Mol. Cancer Res. 2009, 7, 1119–1131. [Google Scholar] [CrossRef]
- Mula, R.V.; Bhatia, V.; Falzon, M. PTHrP promotes colon cancer cell migration and invasion in an integrin alpha6beta4-dependent manner through activation of Rac1. Cancer Lett. 2010, 298, 119–127. [Google Scholar] [CrossRef]
- Park, S.I.; Lee, C.; Sadler, W.D.; Koh, A.J.; Jones, J.; Seo, J.W.; Soki, F.N.; Cho, S.W.; Daignault, S.D.; McCauley, L.K. Parathyroid hormone-related protein drives a CD11b+Gr1+ cell-mediated positive feedback loop to support prostate cancer growth. Cancer Res. 2013, 73, 6574–6583. [Google Scholar] [CrossRef]
- Diaz-Montero, C.M.; Salem, M.L.; Nishimura, M.I.; Garrett-Mayer, E.; Cole, D.J.; Montero, A.J. Increased circulating myeloid-derived suppressor cells correlate with clinical cancer stage, metastatic tumor burden, and doxorubicin-cyclophosphamide chemotherapy. Cancer Immunol. Immunother. 2009, 58, 49–59. [Google Scholar] [CrossRef]
- Soragna, D.; Papi, L.; Ratti, M.T.; Sestini, R.; Tupler, R.; Montalbetti, L. An Italian family affected by Nasu-Hakola disease with a novel genetic mutation in the TREM2 gene. J. Neurol. Neurosurg. Psychiatry 2003, 74, 825–826. [Google Scholar] [CrossRef]
- Gu, Q.; Yang, H.; Shi, Q. Macrophages and bone inflammation. J. Orthop. Transl. 2017, 10, 86–93. [Google Scholar] [CrossRef]
- Oda, Y.; Hashimoto, H.; Tsuneyoshi, M.; Takeshita, S. Survival in synovial sarcoma. A multivariate study of prognostic factors with special emphasis on the comparison between early death and long-term survival. Am. J. Surg. Pathol. 1993, 17, 35–44. [Google Scholar] [CrossRef]
- Guo, Y.; Collaco, C.R.; Bruera, E. Heterotopic ossification in critical illness and cancer: A report of 2 cases. Arch. Phys. Med. Rehabil. 2002, 83, 855–859. [Google Scholar] [CrossRef] [PubMed]
- Carriaga, M.T.; Henson, D.E. The histologic grading of cancer. Cancer 1995, 75, 406–421. [Google Scholar] [CrossRef]
- Baht, G.S.; Vi, L.; Alman, B.A. The Role of the Immune Cells in Fracture Healing. Curr. Osteoporos. Rep. 2018, 16, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Ponzetti, M.; Rucci, N. Updates on Osteoimmunology: What’s New on the Cross-Talk Between Bone and Immune System. Front. Endocrinol. 2019, 10, 236. [Google Scholar] [CrossRef]
- Schabbauer, G.; Matt, U.; Gunzl, P.; Warszawska, J.; Furtner, T.; Hainzl, E.; Elbau, I.; Mesteri, I.; Doninger, B.; Binder, B.R.; et al. Myeloid PTEN promotes inflammation but impairs bactericidal activities during murine pneumococcal pneumonia. J. Immunol. 2010, 185, 468–476. [Google Scholar] [CrossRef]
- Nakanishi, A.; Kitagishi, Y.; Ogura, Y.; Matsuda, S. The tumor suppressor PTEN interacts with p53 in hereditary cancer (Review). Int. J. Oncol. 2014, 44, 1813–1819. [Google Scholar] [CrossRef]
- Yu, C.Y.; Chang, W.C.; Zheng, J.H.; Hung, W.H.; Cho, E.C. Transforming growth factor alpha promotes tumorigenesis and regulates epithelial-mesenchymal transition modulation in colon cancer. Biochem Biophys. Res. Commun. 2018, 506, 901–906. [Google Scholar] [CrossRef]
- Dubansky, B.H.; Dubansky, B.D. Natural development of dermal ectopic bone in the american alligator (Alligator mississippiensis) resembles heterotopic ossification disorders in humans. Anat. Rec. 2018, 301, 56–76. [Google Scholar] [CrossRef]
- Sorkin, M.; Huber, A.K.; Hwang, C.; Carson, W.F.t.; Menon, R.; Li, J.; Vasquez, K.; Pagani, C.; Patel, N.; Li, S.; et al. Regulation of heterotopic ossification by monocytes in a mouse model of aberrant wound healing. Nat. Commun. 2020, 11, 722. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ossification Phenotype | N | |
|---|---|---|
| Rosa-LSL-SS18-SSX1/wt; Ptenfl/fl | ++ | 94 |
| Rosa-LSL-SS18-SSX1/SS18-SSX1; Ptenfl/fl | + | 88 |
| Rosa-LSL-SS18-SSX2/wt; Ptenfl/fl | ++ | 157 |
| Rosa-LSL-SS18-SSX2/SS18-SSX2; Ptenfl/fl | + | 124 |
| Non-Reactive with Bone | Reactive with Bone | |
|---|---|---|
| Nonmetastatic synovial sarcoma | 7 | 3 |
| Metastatic synovial sarcoma | 14 | 36 |
| Nonmetastatic | Metastatic | |
|---|---|---|
| Non-ossifying synovial sarcoma | 60 | 44 |
| Ossifying synovial sarcoma | 7 | 13 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirkham, M.; Kalivas, A.; Fatema, K.; Luelling, S.; Dubansky, B.H.; Dubansky, B.; Jones, K.B.; Barrott, J.J. Underlying Ossification Phenotype in a Murine Model of Metastatic Synovial Sarcoma. Int. J. Mol. Sci. 2020, 21, 2636. https://doi.org/10.3390/ijms21072636
Kirkham M, Kalivas A, Fatema K, Luelling S, Dubansky BH, Dubansky B, Jones KB, Barrott JJ. Underlying Ossification Phenotype in a Murine Model of Metastatic Synovial Sarcoma. International Journal of Molecular Sciences. 2020; 21(7):2636. https://doi.org/10.3390/ijms21072636
Chicago/Turabian StyleKirkham, Matthew, Austen Kalivas, Kaniz Fatema, Sarah Luelling, Brooke H. Dubansky, Benjamin Dubansky, Kevin B. Jones, and Jared J. Barrott. 2020. "Underlying Ossification Phenotype in a Murine Model of Metastatic Synovial Sarcoma" International Journal of Molecular Sciences 21, no. 7: 2636. https://doi.org/10.3390/ijms21072636
APA StyleKirkham, M., Kalivas, A., Fatema, K., Luelling, S., Dubansky, B. H., Dubansky, B., Jones, K. B., & Barrott, J. J. (2020). Underlying Ossification Phenotype in a Murine Model of Metastatic Synovial Sarcoma. International Journal of Molecular Sciences, 21(7), 2636. https://doi.org/10.3390/ijms21072636