The Quantitative Proteome of the Cement and Adhesive Gland of the Pedunculate Barnacle, Pollicipes pollicipes

 ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results
2.1. Protein Identification
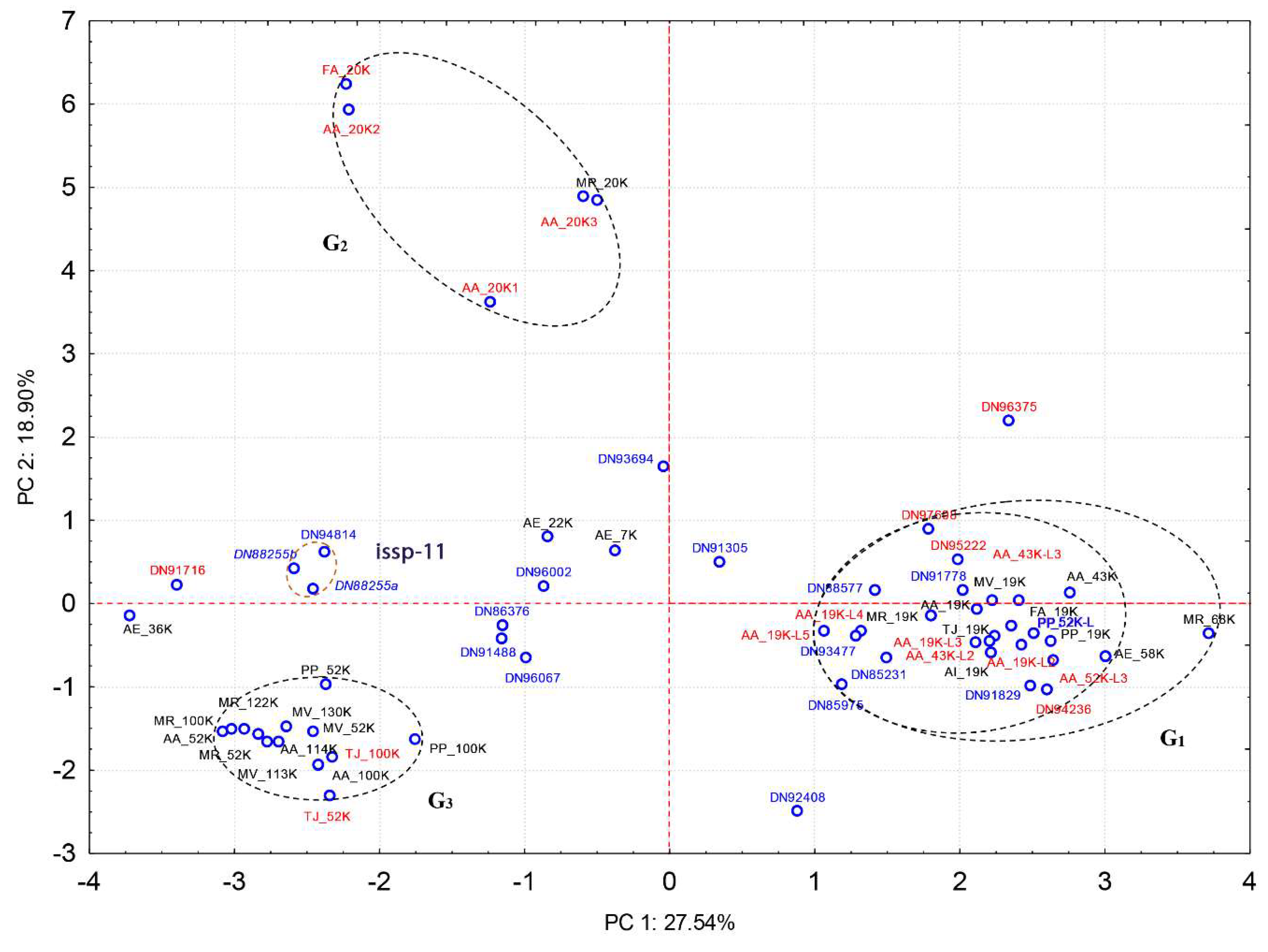
2.2. Quantitative Proteomic Analyses
Unannotated Proteins from the Cement Proteome
3. Discussion
3.1. The Proteome Profile of Pollicipes pollicipes Cement Gland
3.2. Unraveling the Pollicipes pollicipes Cement Proteome
Unannotated Proteins
4. Materials and Methods
4.1. Sampling, Protein Solubilization, and Extraction
4.2. LC-MS/MS Analyses
4.3. Protein Identification and Quantitative Proteomic Analyses
4.4. Data Filtration and Downstream Analyses
4.5. Characterization of Unannotated Cement Proteins
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Foster, B.A. Barnacle ecology and adaptation. In Barnacle Biology; Southward, A.J., Crisp, D.J., Eds.; AA Balkema: Rotterdam, The Netherlands, 1987; p. 443. ISBN 9061916283. [Google Scholar]
- Borja, A.; Liria, P.; Muxika, I.; Bald, J. Relationships between wave exposure and biomass of the goose barnacle (Pollicipes pollicipes, Gmelin, 1790) in the Gaztelugatxe Marine Reserve (Basque Country, northern Spain). ICES J. Mar. Sci. 2016, 63, 226–236. [Google Scholar] [CrossRef]
- Anderson, D.T. Reproductive system, mating and oviposition. In Barnacles. Structure, Function, Development and Evolution; Anderson, D.T., Ed.; Chapman & Hall: London, UK, 1994; pp. 127–170. ISBN 0-412-44420-8. [Google Scholar]
- Stewart, R.J. Protein-based underwater adhesives and the prospects for their biotechnological production. Appl. Microbiol. Biotechnol. 2011, 89, 27–33. [Google Scholar] [CrossRef]
- Schultz, M.P.; Bendick, J.A.; Holm, E.R.; Hertel, W.M. Economic impact of biofouling on a naval surface ship. Biofouling 2010, 27, 87–98. [Google Scholar] [CrossRef]
- Kamino, K. Underwater Adhesive of Marine Organisms as the Vital Link between Biological Science and Material Science. Mar. Biotechnol. 2008, 10, 111–121. [Google Scholar] [CrossRef]
- Bhagat, V.; Becker, M.L. Degradable Adhesives for Surgery and Tissue Engineering. Biomacromolecules 2017, 18, 3009–3039. [Google Scholar] [CrossRef]
- Meredith, H.J.; Jenkins, C.L.; Wilker, J.J. Enhancing the Adhesion of a Biomimetic Polymer Yields Performance Rivaling Commercial Glues. Adv. Funct. Mater. 2014, 24, 3259–3267. [Google Scholar] [CrossRef]
- Kamino, K. Barnacle underwater attachment. In Biological Adhesives; Springer: Berlin/Heidelberg, Germany, 2006; pp. 145–166. [Google Scholar]
- So, C.R.; Scancella, J.M.; Fears, K.P.; Essock-Burns, T.; Haynes, S.E.; Leary, D.H.; Diana, Z.; Wang, C.; North, S.; Oh, C.S.; et al. Oxidase Activity of the Barnacle Adhesive Interface Involves Peroxide-Dependent Catechol Oxidase and Lysyl Oxidase Enzymes. ACS Appl. Mater. Interfaces 2017, 9, 11493–11505. [Google Scholar] [CrossRef]
- Fears, K.P.; Orihuela, B.; Rittschof, D.; Wahl, K.J. Acorn Barnacles Secrete Phase-Separating Fluid to Clear Surfaces Ahead of Cement Deposition. Adv. Sci. 2018, 5, 1700762. [Google Scholar] [CrossRef]
- Liang, C.; Strickland, J.; Ye, Z.; Wu, W.; Hu, B.; Rittschof, D. Biochemistry of Barnacle Adhesion: An Updated Review. Front. Mar. Sci. 2019, 6, 565. [Google Scholar] [CrossRef]
- Kamino, K. Barnacle underwater attachment. In Biological Adhesives; Springer: Cham, Switzerland, 2016; pp. 153–176. [Google Scholar]
- So, C.R.; Fears, K.P.; Leary, D.H.; Scancella, J.M.; Wang, Z.; Liu, J.L.; Orihuela, B.; Rittschof, D.; Spillmann, C.M.; Wahl, K.J. Sequence basis of Barnacle Cement Nanostructure is Defined by Proteins with Silk Homology. Sci. Rep. 2016, 6, 36219. [Google Scholar] [CrossRef]
- Lobo-da-Cunha, A.; Alves, Ï.; Oliveira, E.; Cunha, I. The cement apparatus of the stalked barnacle Pollicipes pollicipes. Mar. Biol. 2017, 164, 11. [Google Scholar] [CrossRef]
- Gohad, N.V.; Aldred, N.; Hartshorn, C.M.; Lee, Y.J.; Cicerone, M.T.; Orihuela, B.; Clare, A.S.; Rittschof, D.; Mount, A.S. Synergistic roles for lipids and proteins in the permanent adhesive of barnacle larvae. Nat. Commun. 2014, 5, 4414. [Google Scholar] [CrossRef] [PubMed]
- Kamino, K.; Nakano, M.; Kanai, S. Significance of the conformation of building blocks in curing of barnacle underwater adhesive. FEBS J. 2012, 279, 1750–1760. [Google Scholar] [CrossRef] [PubMed]
- Barnes, M.K.S. MarLIN—The Marine Life Information Network. Available online: https://www.marlin.ac.uk/ (accessed on 4 April 2020).
- Rocha, M.; Antas, P.; Castro, L.F.C.; Campos, A.; Vasconcelos, V.; Pereira, F.; Cunha, I. Comparative Analysis of the Adhesive Proteins of the Adult Stalked Goose Barnacle Pollicipes pollicipes (Cirripedia: Pedunculata). Mar. Biotechnol. 2019, 21, 38–51. [Google Scholar] [CrossRef]
- Naldrett, M.J.; Kaplan, D.L. Characterization of barnacle (Balanus eburneus and B. cenatus) adhesive proteins. Mar. Biol. 1997, 127, 629–635. [Google Scholar] [CrossRef]
- Guruprasad, K.; Reddy, B.V.; Pandit, M.W. Correlation between stability of a protein and its dipeptide composition: A novel approach for predicting in vivo stability of a protein from its primary sequence. Protein Eng. 1990, 4, 155–161. [Google Scholar] [CrossRef]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef]
- Ikai, A. Thermostability and aliphatic index of globular proteins. J. Biochem. 1980, 88, 1895–1898. [Google Scholar]
- Schlessinger, A.; Rost, B. Protein flexibility and rigidity predicted from sequence. Proteins 2005, 61, 115–126. [Google Scholar] [CrossRef]
- Chen, Z.-F.; Zhang, H.; Wang, H.; Matsumura, K.; Wong, Y.H.; Ravasi, T.; Qian, P.-Y. Quantitative Proteomics Study of Larval Settlement in the Barnacle Balanus amphitrite. PLoS ONE 2014, 9, e88744. [Google Scholar] [CrossRef]
- Mithieux, S.M.; Weiss, A.S. Elastin. Adv. Protein Chem. 2005, 70, 437–461. [Google Scholar]
- Ritossa, F. Discovery of the heat shock response. Cell Stress Chaperones 1996, 1, 97–98. [Google Scholar] [CrossRef]
- Heikkila, J.J.; Ohan, N.; Tam, Y.; Ali, A. Heat shock protein gene expression during Xenopus development. Cell. Mol. Life Sci. 1997, 53, 114–121. [Google Scholar] [CrossRef]
- Gunter, H.M.; Degnan, B.M. Developmental expression of Hsp90, Hsp70 and HSF during morphogenesis in the vetigastropod Haliotis asinina. Dev. Genes Evol. 2007, 217, 603–612. [Google Scholar] [CrossRef]
- Arbeitman, M.N.; Hogness, D.S. Molecular chaperones activate the Drosophila ecdysone receptor, an RXR heterodimer. Cell 2000, 101, 67–77. [Google Scholar] [CrossRef]
- Bishop, C.D.; Brandhorst, B.P. NO/cGMP signaling and HSP90 activity represses metamorphosis in the sea urchin Lytechinus pictus. Biol. Bull. 2001, 201, 394–404. [Google Scholar] [CrossRef]
- Mahroof, R.; Kun, Y.Z.; Neven, L.; Subramanyam, B.; Bai, J. Expression patterns of three heat shock protein 70 genes among developmental stages of the red flour beetle, Tribolium castaneum (Coleoptera: Tenebrionidae). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2005, 141, 247–256. [Google Scholar] [CrossRef]
- Anil, A.; Desai, D.; Khandeparker, L.; Gaonkar, C.A. Barnacles and Their Significance in Biofouling. In Operational and Environmental Consequences of Large Industrial Cooling Water Systems; Rajagopal, S., Jenner, H., Venugopalan, V., Eds.; Springer: Boston, MA, USA, 2012; pp. 65–93. [Google Scholar]
- Nakano, M.; Kamino, K. Amyloid-like conformation and interaction for the self-assembly in barnacle underwater cement. Biochemistry 2015, 54, 826–835. [Google Scholar] [CrossRef]
- Kamino, K.; Inoue, K.; Maruyama, T.; Takamatsu, N.; Harayama, S.; Shizuri, Y. Barnacle Cement Proteins. Importance of disulfide bonds in their insolubility. J. Biol. Chem. 2000, 275, 27360–27365. [Google Scholar] [CrossRef]
- Urushida, Y.; Nakano, M.; Matsuda, S.; Inoue, N.; Kanai, S.; Kitamura, N.; Nishino, T.; Kamino, K. Identification and functional characterization of a novel barnacle cement protein. FEBS J. 2007, 274, 4336–4346. [Google Scholar] [CrossRef]
- Mori, Y.; Urushida, Y.; Nakano, M.; Uchiyama, S.; Kamino, K. Calcite-specific coupling protein in barnacle underwater cement. FEBS J. 2007, 274, 6436–6446. [Google Scholar] [CrossRef]
- He, L.-S.; Zhang, G.; Qian, P.-Y. Characterization of Two 20kDa-Cement Protein (cp20k) Homologues in Amphibalanus amphitrite. PLoS ONE 2013, 8, e64130. [Google Scholar] [CrossRef]
- Lin, H.C.; Wong, Y.H.; Tsang, L.M.; Chu, K.H.; Qian, P.Y.; Chan, B.K.K. First study on gene expression of cement proteins and potential adhesion-related genes of a membranous-based barnacle as revealed from Next-Generation Sequencing technology. Biofouling 2014, 30, 169–181. [Google Scholar] [CrossRef]
- Matsumura, K.; Nagano, M.; Fusetani, N. Purification of a larval settlement-inducing protein complex (SIPC) of the barnacle, Balanus amphitrite. J. Exp. Zool. 1998, 281, 12–20. [Google Scholar] [CrossRef]
- Dreanno, C.; Kirby, R.R.; Clare, A.S. Locating the barnacle settlement pheromone: Spatial and ontogenetic expression of the settlement-inducing protein complex of Balanus amphitrite. Proc. R. Soc. B Biol. Sci. 2006, 273, 2721–2728. [Google Scholar] [CrossRef]
- Clare, A.S. Toward a characterization of the chemical cue to barnacle gregariousness. In Chemical Communication in Crustaceans; Springer: New York, NY, USA, 2011; pp. 431–450. ISBN 9780387771014. [Google Scholar]
- Zhang, G.; Yang, X.X.; Leung, P.M.; He, L.S.; Chan, T.Y.; Yan, G.Y.; Zhang, Y.; Sun, J.; Xu, Y.; Qian, P.Y. Secretory locations of SIPC in Amphibalanus amphitrite cyprids and a novel function of SIPC in biomineralization. Sci. Rep. 2016, 6, 29376. [Google Scholar] [CrossRef]
- Petrone, L.; Aldred, N.; Emami, K.; Enander, K.; Ederth, T.; Clare, A.S.; Clare, A.S. Chemistry-specific surface adsorption of the barnacle settlement-inducing protein complex. Interface Focus 2015, 5, 20140047. [Google Scholar] [CrossRef]
- Dreanno, C.; Matsumura, K.; Dohmae, N.; Takio, K.; Hirota, H.; Kirby, R.R.; Clare, A.S. An α2-macroglobulin-like protein is the cue to gregarious settlement of the barnacle Balanus amphitrite. Proc. Natl. Acad. Sci. USA 2006, 103, 14396–14401. [Google Scholar] [CrossRef]
- Lewis, C.A. Some observations of factors affecting embryonic and larval growth of Pollicipes polymerus (Cirripedia: Lepadomorpha) in vitro. Mar. Biol. 1975, 32, 127–139. [Google Scholar] [CrossRef]
- Hoffman, D.L. Foot-loose barnacles: Implications of the retention of mobility into the post- metamorphic stages of the goose barnacle. American Zoologist. Am. Zool. 1981, 21, 911–1052. [Google Scholar]
- Essock-Burns, T.; Soderblom, E.J.; Orihuela, B.; Moseley, M.A.; Rittschof, D.; Mount, A.S.; Reitzel, A.M.; Essock-burns, T. Hypothesis Testing with Proteomics: A Case Study Using Wound Healing Mechanisms in Fluids Associated with Barnacle Glue. Front. Mar. Sci. 2019, 6, 343. [Google Scholar] [CrossRef]
- Armstrong, P.B.; Quigley, J.P. α2-macroglobulin: An evolutionarily conserved arm of the innate immune system. Dev. Comp. Immunol. 1999, 23, 375–390. [Google Scholar] [CrossRef]
- Yorisue, T.; Matsumura, K.; Hirota, H.; Dohmae, N. Possible molecular mechanisms of species recognition by barnacle larvae inferred from multi- specific sequencing analysis of proteinaceous settlement-inducing pheromone. Biofouling 2012, 28, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Aldred, N.; Phang, I.Y.; Conlan, S.L.; Clare, A.S.; Vancso, G.J. The effects of a serine protease, Alcalase, on the adhesives of barnacle cyprids (Balanus amphitrite). Biofouling 2008, 24, 97–107. [Google Scholar] [CrossRef]
- Ferrier, G.A.; Kim, S.J.; Kaddis, C.S.; Loo, J.A.; Ann Zimmer, C.; Zimmer, R.K. MULTIFUNCin: A Multifunctional Protein Cue Induces Habitat Selection by, and Predation on, Barnacles. In Proceedings of the Integrative and Comparative Biology; Oxford University Press: Oxford, UK, 2016; Volume 56, pp. 901–913. [Google Scholar]
- Walley, L.J.; Rees, E.I.S. Studies on the Larval Structure and Metamorphosis of Balanus balanoides (L.). Philos. Trans. R. Soc. B Biol. Sci. 1969, 256, 237–280. [Google Scholar]
- Dougherty, W.J. Zinc metalloprotease activity in the cement precursor secretion of the barnacle, Chthamalus fragilis Darwin. Tissue Cell 1996, 28, 439–447. [Google Scholar] [CrossRef]
- Wu, C.; Charoensapsri, W.; Nakamura, S.; Tassanakajon, A.; Söderhäll, I.; Söderhäll, K. An MBL-like protein may interfere with the activation of the proPO-system, an important innate immune reaction in invertebrates. Immunobiology 2013, 218, 159–168. [Google Scholar] [CrossRef]
- Amparyup, P.; Donpudsa, S.; Tassanakajon, A. Shrimp single WAP domain (SWD)-containing protein exhibits proteinase inhibitory and antimicrobial activities. Dev. Comp. Immunol. 2008, 32, 1497–1509. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, Y.T.; Li, X.J.; Wang, S.C.; Li, D.; Li, W.W.; Wang, Q. Lipopolysaccharide and beta-1, 3-glucan binding protein (LGBP) stimulates prophenoloxidase activating system in Chinese mitten crab (Eriocheir sinensis). Dev. Comp. Immunol. 2016, 61, 70–79. [Google Scholar] [CrossRef]
- Wang, C.S.; Ashton, N.N.; Weiss, R.B.; Stewart, R.J. Peroxinectin catalyzed dityrosine crosslinking in the adhesive underwater silk of a casemaker caddisfly larvae, Hysperophylax occidentalis. Insect Biochem. Mol. Biol. 2014, 54, 69–79. [Google Scholar] [CrossRef]
- Wang, C.S.; Pan, H.; Weerasekare, G.M.; Stewart, R.J. Peroxidase-catalysed interfacial adhesion of aquatic caddisworm silk. J. R. Soc. Interface 2015, 12, 20150710. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hodgeman, B.A.; Christensen, B.M. Involvement of peroxidase in chorion hardening in Aedes aegypti. Insect Biochem. Mol. Biol. 1996, 26, 309–317. [Google Scholar] [CrossRef]
- Wang, X.; Wang, C.; Xu, B.; Wei, J.; Xiao, Y.; Huang, F. Adsorption of intrinsically disordered barnacle adhesive proteins on silica surface. Appl. Surf. Sci. 2018, 427, 942–949. [Google Scholar] [CrossRef]
- Cheng, S.; Cetinkaya, M.; Gräter, F. How Sequence Determines Elasticity of Disordered Proteins. Biophys. J. 2010, 99, 3863–3869. [Google Scholar] [CrossRef] [PubMed]
- Rauscher, S.; Baud, S.; Miao, M.; Keeley, F.W.; Pomès, R. Proline and Glycine Control Protein Self-Organization into Elastomeric or Amyloid Fibrils. Structure 2006, 14, 1667–1676. [Google Scholar] [CrossRef] [PubMed]
- Dicko, C.; Porter, D.; Bond, J.; Kenney, J.M.; Vollrath, F. Structural Disorder in Silk Proteins Reveals the Emergence of Elastomericity. Biomacromolecules 2008, 9, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Sullan, R.M.A.; Gunari, N.; Tanur, A.E.; Chan, Y.; Dickinson, G.H.; Orihuela, B.; Rittschof, D.; Walker, G.C. Nanoscale structures and mechanics of barnacle cement. Biofouling 2009, 25, 263–275. [Google Scholar] [CrossRef]
- Barlow, D.E.; Dickinson, G.H.; Orihuela, B.; Kulp, J.L.; Rittschof, D.; Wahl, K.J. Characterization of the Adhesive Plaque of the Barnacle Balanus amphitrite: Amyloid-Like Nanofibrils Are a Major Component. Langmuir 2010, 26, 6549–6556. [Google Scholar] [CrossRef]
- Zhong, C.; Gurry, T.; Cheng, A.A.; Downey, J.; Deng, Z.; Stultz, C.M.; Lu, T.K. Strong underwater adhesives made by self-assembling multi-protein nanofibres. Nat. Nanotechnol. 2014, 9, 858–866. [Google Scholar] [CrossRef]
- Liu, X.; Liang, C.; Zhang, X.; Li, J.; Huang, J.; Zeng, L.; Ye, Z.; Hu, B.; Wu, W. Amyloid fibril aggregation: An insight into the underwater adhesion of barnacle cement. Biochem. Biophys. Res. Commun. 2017, 493, 654–659. [Google Scholar] [CrossRef]
- Raman, S.; Malms, L.; Utzig, T.; Shrestha, B.R.; Stock, P.; Krishnan, S.; Valtiner, M. Adhesive barnacle peptides exhibit a steric-driven design rule to enhance adhesion between asymmetric surfaces. Colloids Surfaces B Biointerfaces 2017, 152, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Kamino, K. Novel barnacle underwater adhesive protein is a charged amino acid-rich protein constituted by a Cys-rich repetitive sequence. Biochem. J. 2001, 356, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.; Danielsson, G.; Farinha, A.P.; Kuruvilla, J.; Warholm, P.; Cristobal, S. Shotgun proteomics to unravel marine mussel (Mytilus edulis) response to long-term exposure to low salinity and propranolol in a Baltic Sea microcosm. J. Proteom. 2016, 137, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.S.; Foehr, S.; Garfield, D.A.; Furlong, E.E.; Steinmetz, L.M.; Krijgsveld, J. Ultrasensitive proteome analysis using paramagnetic bead technology. Mol. Syst. Biol. 2014, 10, 757. [Google Scholar] [CrossRef] [PubMed]
- Sielaff, M.; Kuharev, J.; Bohn, T.; Hahlbrock, J.; Bopp, T.; Tenzer, S.; Distler, U. Evaluation of FASP, SP3, and iST Protocols for Proteomic Sample Preparation in the Low Microgram Range. J. Proteome Res. 2017, 16, 4060–4072. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized ppb-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Machado, A.M.; Sarropoulou, E.; Castro, L.F.C.; Vasconcelos, V.; Cunha, I. An important resource for understanding bio-adhesion mechanisms: Cement gland transcriptomes of two goose barnacles, Pollicipes pollicipes and Lepas anatifera (Cirripedia, Thoracica). Mar. Genom. 2019, 45, 16–20. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote) omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Yachdav, G.; Kloppmann, E.; Kajan, L.; Hecht, M.; Goldberg, T.; Hamp, T.; Hönigschmid, P.; Schafferhans, A.; Roos, M.; Bernhofer, M.; et al. PredictProtein--an open resource for online prediction of protein structural and functional features. Nucleic Acids Res. 2014, 42, W337–W343. [Google Scholar] [CrossRef]







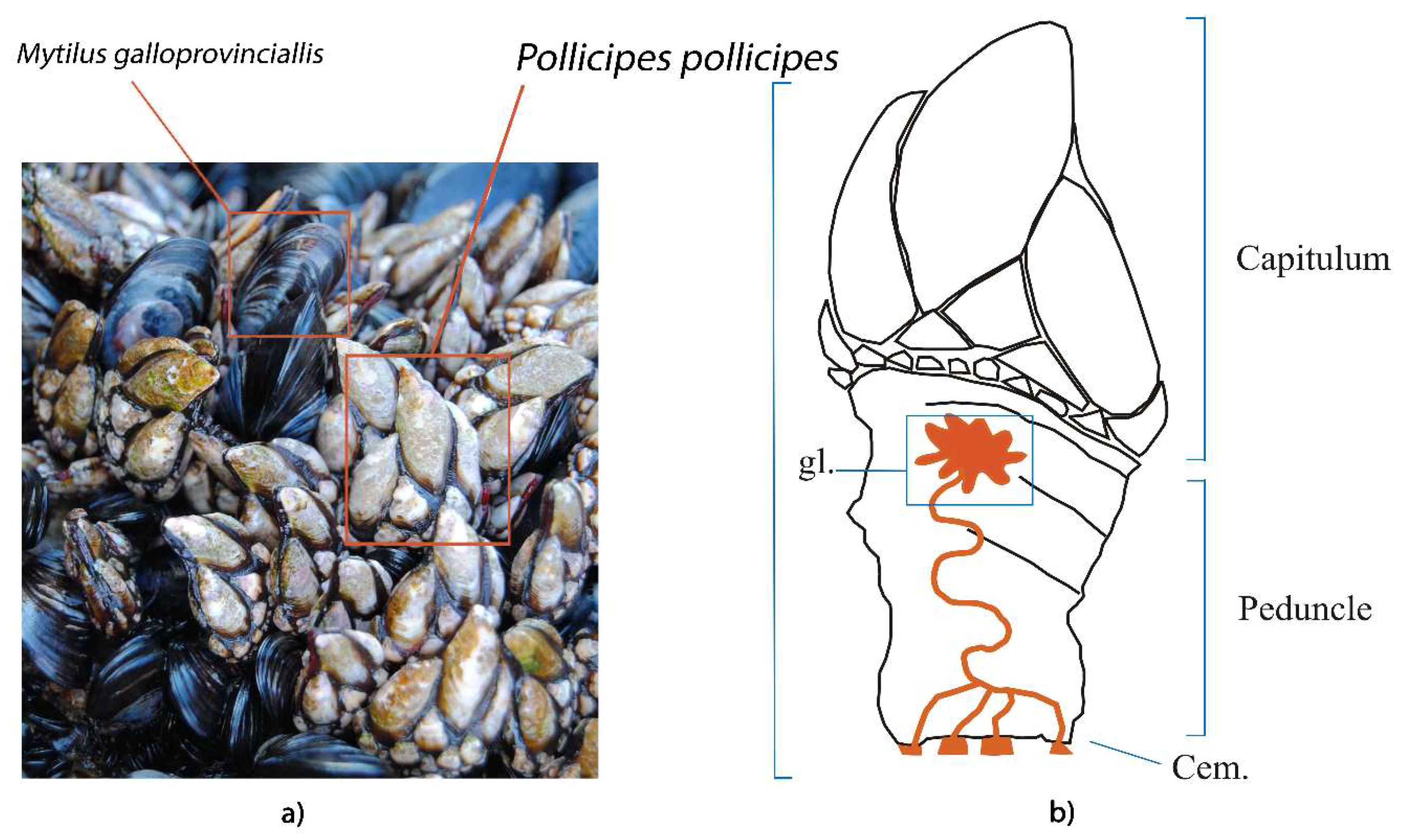{kind=link}
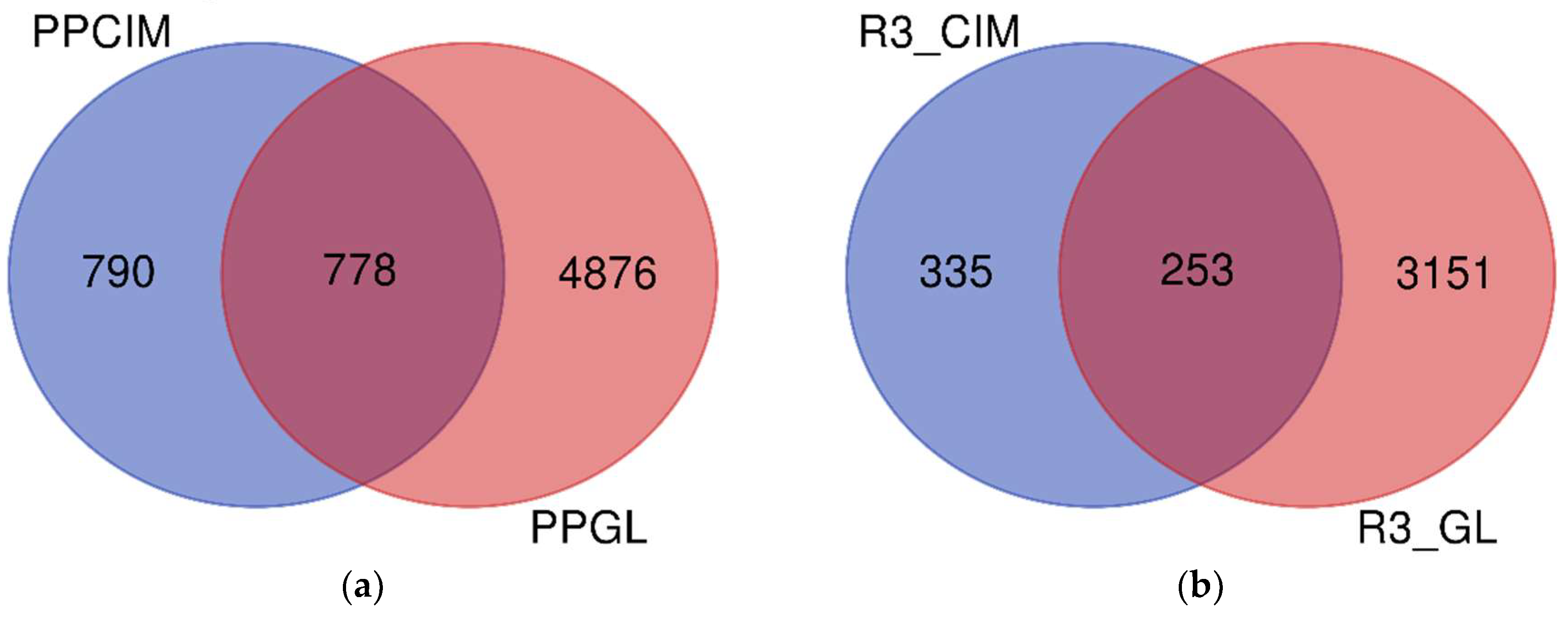{kind=link}
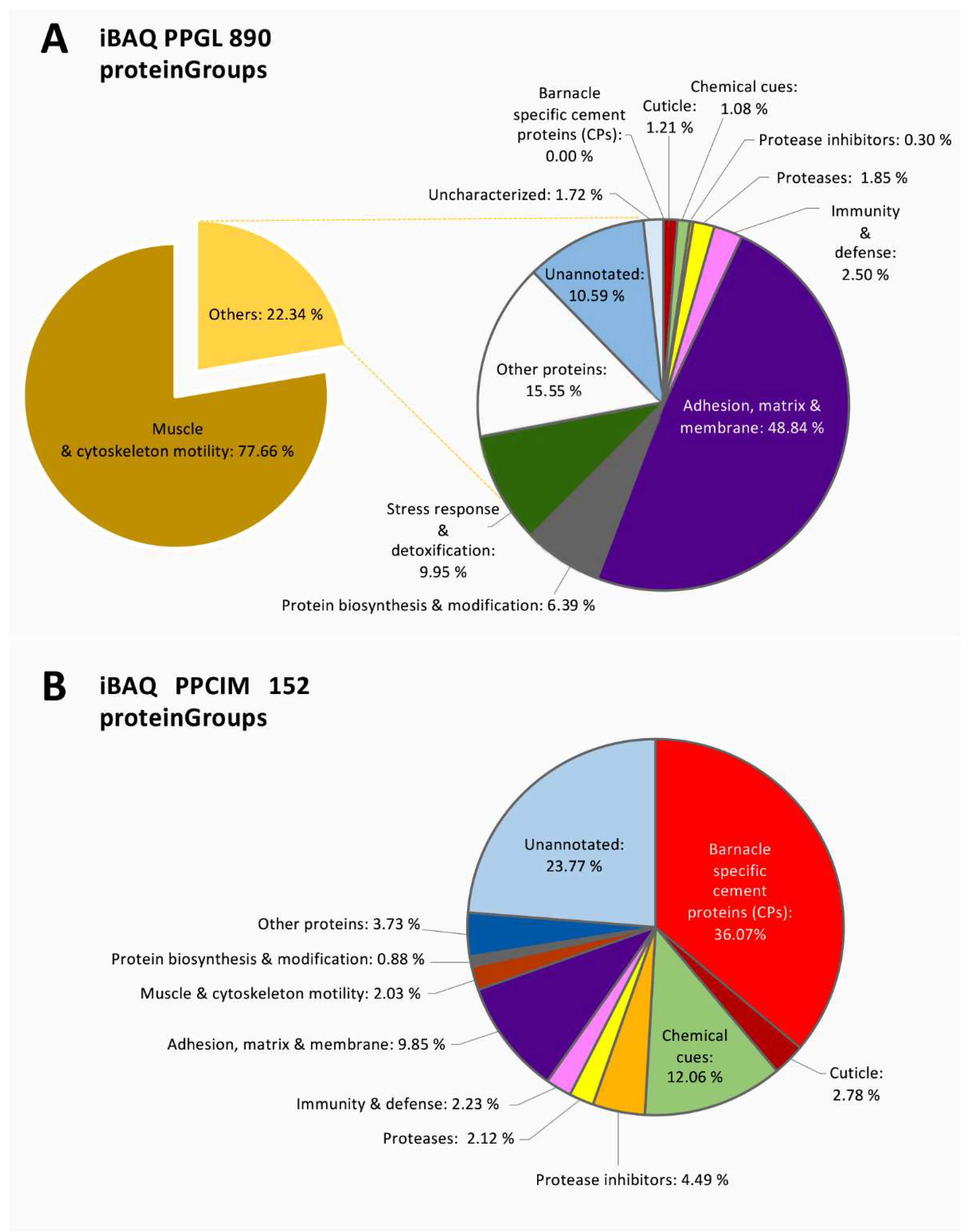{kind=link}
{kind=link}
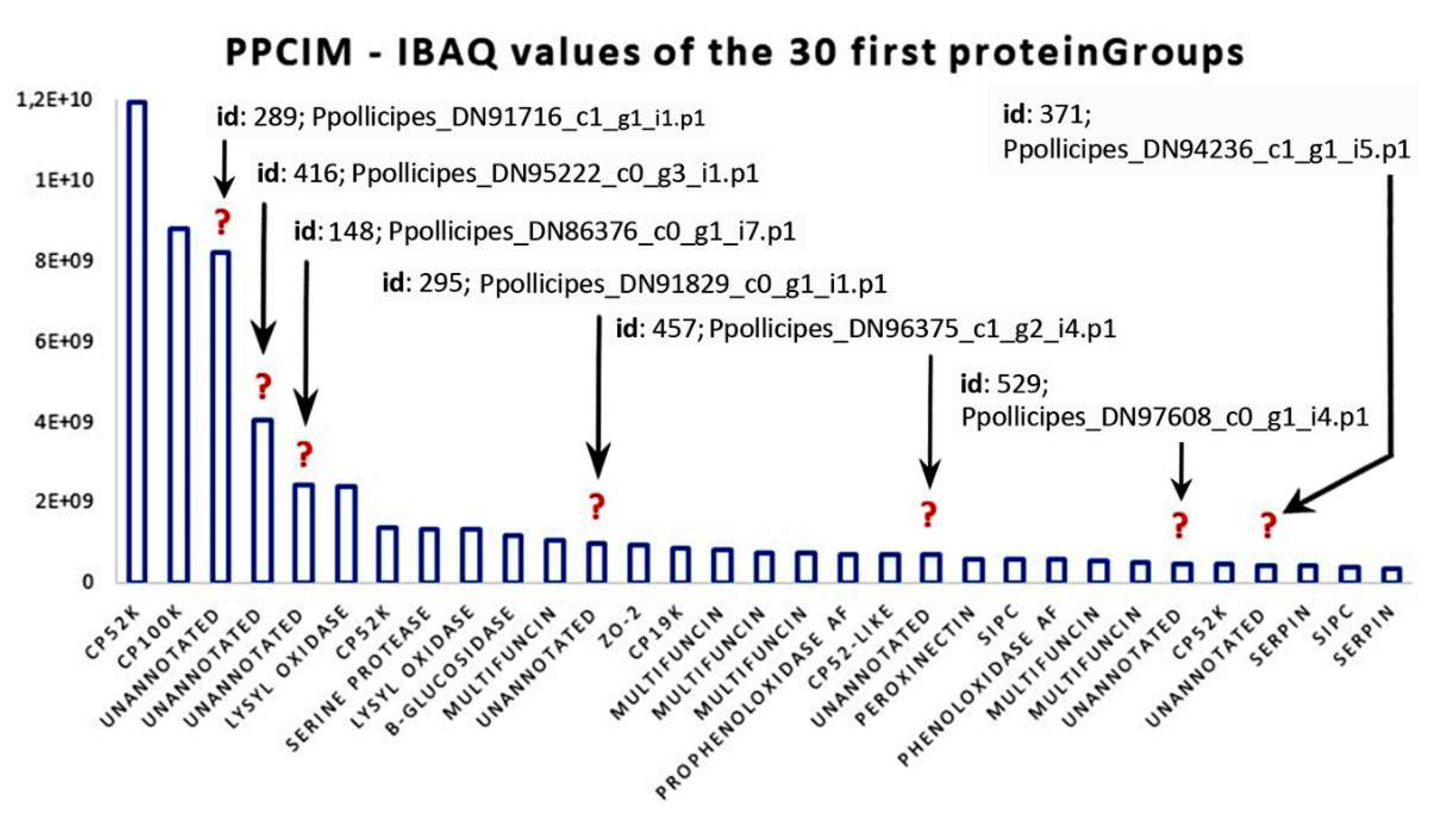{kind=link}
{kind=link}
| iBAQ Rank | Barnacle Specific Leading Cement Proteins | No. Res | MM(x1000) | pI | Instability Index a | Hydropathy (GRAVY) b | % Neg. res. | % Pos. res. | Aliphatic Index c | Aromatic res. (%) | Secondary Structure Composition (%) | Solvent Accessibility (%) | Protein Disorder (%) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Loop | α-Helix | β-Sheet | Exposed | Interm | Buried | |||||||||||||
| 3 | DN91716_c1_g1_i1.p1 | 390 | 45.4 | 10.4 | 50.95 | −0.142 | 6 | 47 | 88.69 | 17.5 | 53.13 | 25.38 | 19.49 | 26.15 | 7.44 | 66.41 | 0.51 | |
| 4 | DN95222_c0_g3_i1.p1 | 145 | 15.3 | 9.1 | 29.78 | −0.768 | 19 | 22 | 64.48 | 0.7 | 63.45 | 10.34 | 26.21 | 72.41 | 3.45 | 24.14 | 100.0 | |
| 5 | DN86376_c0_g1_i7.p1 | 398 | 42.8 | 9.57 | 41.03 | 0.260 | 7 | 11 | 104.95 | 6.8 | 40.7 | 35.18 | 24.12 | 40.20 | 0.00 | 59.80 | 1.78 | |
| 12 | DN91829_c0_g1_i1.p1 | 269 | 25.3 | 4.1 | 49.53 | 0.111 | 28 | 7 | 79.93 | 2.2 | 92.19 | 5.58 | 2.23 | 64.68 | 1.86 | 33.56 | 69.58 | |
| 19 | DN93583_c0_g1_i1.p2 | 243 | 23.7 | 5.4 | 34.24 | −0.293 | 11 | 9 | 72.3 | 0.4 | 91.8 | 6.1 | 2.1 | 68.7 | 2.1 | 29.2 | 91.77 | |
| 20 | DN96375_c1_g2_i4.p1 | 281 | 27.9 | 4.0 | 21.94 | −0.67 | 51 | 14 | 62.17 | 0.8 | 77.94 | 6.05 | 16.01 | 54.09 | 2.85 | 43.03 | 45.20 | |
| 26 | DN97608_c0_g1_i4.p1 | 146 | 14.4 | 10.7 | 20.79 | −0.738 | 9 | 21 | 52.95 | 2.1 | 92.84 | 3.42 | 2.74 | 71.92 | 2.74 | 25.34 | 100.0 | |
| 28 | DN94236_c1_g1_i5.p1 | 312 | 29.3 | 6.7 | 21.37 | −0.013 | 21 | 20 | 69.01 | 2.5 | 97.76 | 0.00 | 2.24 | 90.38 | 0.32 | 9.29 | 93.59 | |
| 36 | DN94814_c0_g2_i2.p1 | 240 | 26.3 | 10.6 | 24.38 | 0.12 | 12 | 36 | 101.58 | 8.3 | 50.00 | 43.33 | 6.67 | 37.08 | 7.08 | 55.83 | 4.58 | |
| 38 | DN93477_c2_g1_i1.p1 | 353 | 37.4 | 8.5 | 37.24 | −0.532 | 44 | 46 | 79.52 | 0.6 | 90.93 | 1.42 | 7.65 | 68.56 | 3.40 | 28.05 | 64.58 | |
| 48 | DN92408_c0_g3_i2.p1 | 392 | 43.5 | 11.5 | 25.19 | −0.148 | 13 | 37 | 89.92 | 4.3 | 96.68 | 0.00 | 3.32 | 94.90 | 0.51 | 4.59 | 96.17 | |
| 57 | DN85975_c0_g1_i3.p2 | 203 | 19.8 | 4.8 | 43.26 | −0.159 | 16 | 11 | 55.91 | 6.9 | 94.58 | 5.42 | 0.00 | 88.67 | 1.48 | 9.85 | 95.07 | |
| 60 | DN88255_c1_g1_i1.p1 | 265 | 30.0 | 11.3 | 49.76 | −0.206 | 14 | 45 | 96.45 | 6.9 | 43.77 | 41.17 | 9.06 | 50.94 | 7.55 | 41.51 | 7.17 | |
| 79 | DN85231_c0_g1_i1.p1 | 246 | 23.7 | 12.4 | 25.83 | 0.049 | 3 | 20 | 77.03 | 1.6 | 96.34 | 0.00 | 3.66 | 67.48 | 1.22 | 31.30 | 87.80 | |
| 87 | DN88577_c1_g1_i6.p1 | 105 | 10.3 | 6.8 | 4.04 | 0.25 | 5 | 5 | 75.24 | 4.5 | 60.00 | 0.00 | 40.00 | 60.95 | 4.76 | 34.29 | 62.86 | |
| 89 | DN96067_c1_g1_i3.p1 | 183 | 20.2 | 10.8 | 19.96 | 0.138 | 8 | 28 | 106.94 | 7.6 | 34.97 | 19.13 | 45.9 | 43.72 | 8.74 | 47.54 | 2.73 | |
| 95 | DN96002_c6_g1_i8.p1 | 238 | 26.9 | 10.0 | 57.57 | −0.786 | 23 | 37 | 57.77 | 7.6 | 59.24 | 12.61 | 26.15 | 59.24 | 4.62 | 36.13 | 51.68 | |
| 102 | DN91305_c0_g1_i7.p3 | 190 | 20.5 | 6.9 | 38.75 | −0.453 | 21 | 21 | 69.68 | 4.2 | 46.84 | 14.21 | 38.95 | 55.79 | 5.79 | 38.42 | 26.32 | |
| 103 | DN91778_c0_g1_i1.p1 | 606 | 59.3 | 5.0 | 55.78 | −0.332 | 49 | 32 | 67.79 | 0.9 | 99.05 | 0.00 | 0.50 | 96.20 | 0.00 | 3.80 | 97.95 | |
| 106 | DN91488_c0_g1_i1.p1 | 204 | 22.3 | 9.0 | 46.8 | −0.23 | 17 | 22 | 68.63 | 7.4 | 39.71 | 53.92 | 6.37 | 52.94 | 2.45 | 44.61 | 29.90 | |
| 108 | DN88255_c1_g2_i2.p1 | 265 | 29.9 | 10.8 | 54.04 | −0.186 | 18 | 42 | 101.62 | 5.7 | 46.79 | 37.36 | 15.85 | 41.51 | 5.66 | 52.83 | 7.55 | |
| 141 | DN93694_c2_g1_i2.p1 | 302 | 30.9 | 8.6 | 36.49 | −0.364 | 20 | 24 | 70.13 | 4.0 | 3.31 | 96.69 | 0 | 56.95 | 4.97 | 38.08 | 84.44 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domínguez-Pérez, D.; Almeida, D.; Wissing, J.; Machado, A.M.; Jänsch, L.; Castro, L.F.; Antunes, A.; Vasconcelos, V.; Campos, A.; Cunha, I. The Quantitative Proteome of the Cement and Adhesive Gland of the Pedunculate Barnacle, Pollicipes pollicipes. Int. J. Mol. Sci. 2020, 21, 2524. https://doi.org/10.3390/ijms21072524
Domínguez-Pérez D, Almeida D, Wissing J, Machado AM, Jänsch L, Castro LF, Antunes A, Vasconcelos V, Campos A, Cunha I. The Quantitative Proteome of the Cement and Adhesive Gland of the Pedunculate Barnacle, Pollicipes pollicipes. International Journal of Molecular Sciences. 2020; 21(7):2524. https://doi.org/10.3390/ijms21072524
Chicago/Turabian StyleDomínguez-Pérez, Dany, Daniela Almeida, Josef Wissing, André M. Machado, Lothar Jänsch, Luís Filipe Castro, Agostinho Antunes, Vitor Vasconcelos, Alexandre Campos, and Isabel Cunha. 2020. "The Quantitative Proteome of the Cement and Adhesive Gland of the Pedunculate Barnacle, Pollicipes pollicipes" International Journal of Molecular Sciences 21, no. 7: 2524. https://doi.org/10.3390/ijms21072524
APA StyleDomínguez-Pérez, D., Almeida, D., Wissing, J., Machado, A. M., Jänsch, L., Castro, L. F., Antunes, A., Vasconcelos, V., Campos, A., & Cunha, I. (2020). The Quantitative Proteome of the Cement and Adhesive Gland of the Pedunculate Barnacle, Pollicipes pollicipes. International Journal of Molecular Sciences, 21(7), 2524. https://doi.org/10.3390/ijms21072524