Human Melanoma-Cell Metabolic Profiling: Identification of Novel Biomarkers Indicating Metastasis

 , ,
, ,  ,
,  ,
,  and
and 
Abstract
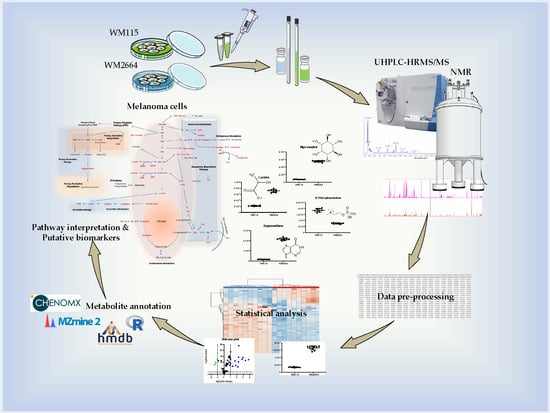
1. Introduction
2. Results
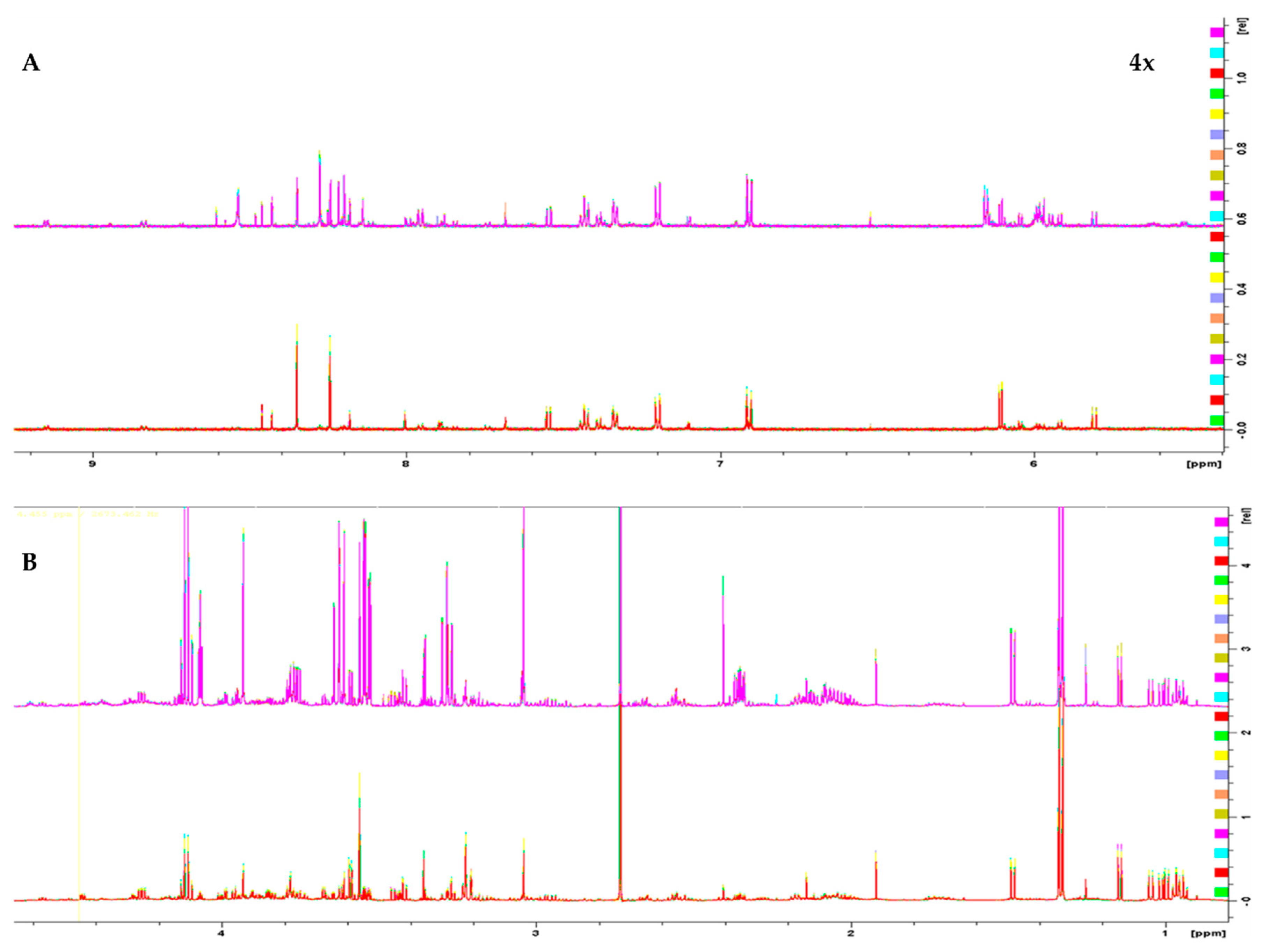
2.1. NMR
2.1.1. Metabolite Identification
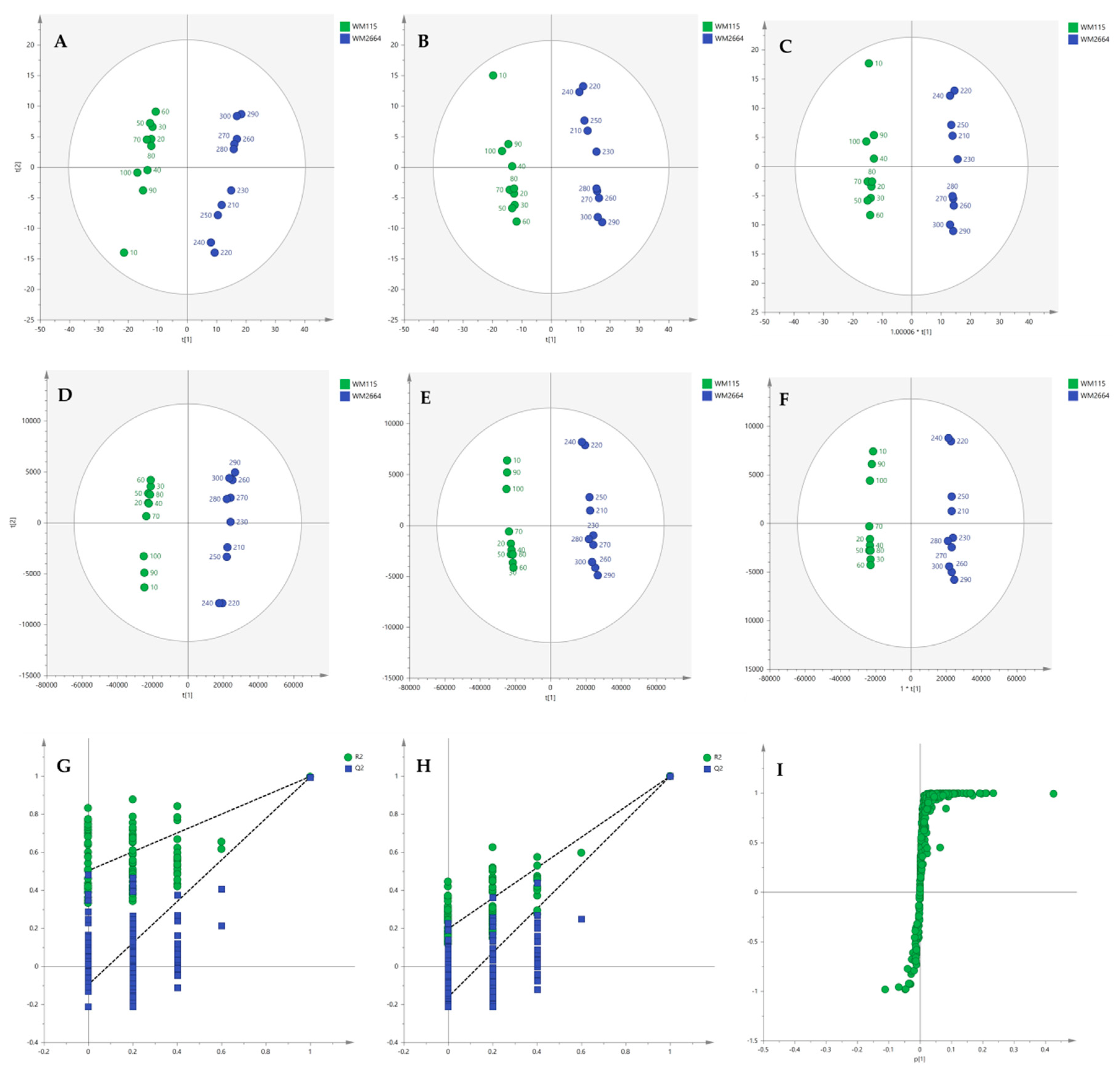
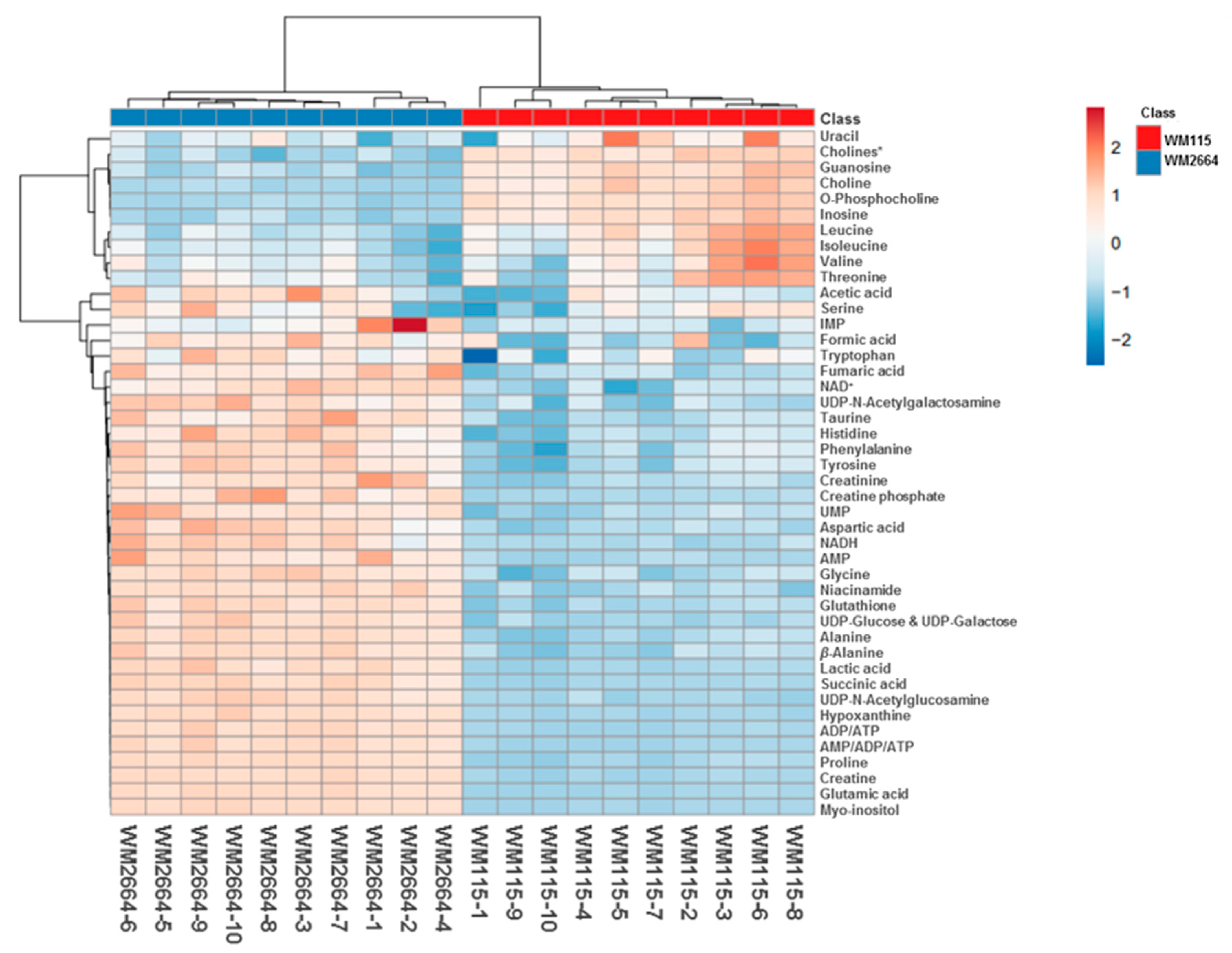
2.1.2. Chemometric Analysis of NMR Data
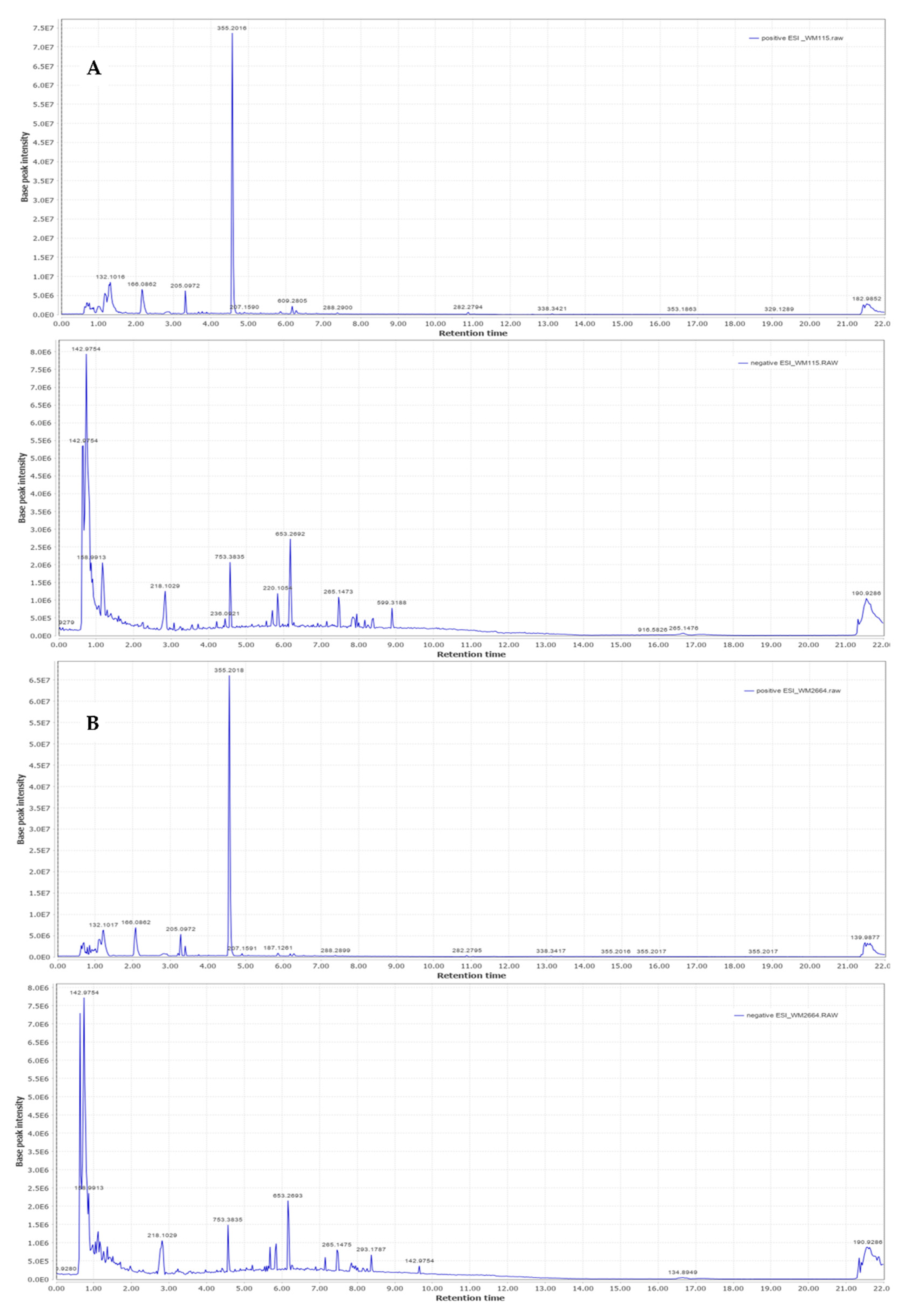
2.2. UHPLC-HRMS/MS
2.2.1. MS Spectra Processing
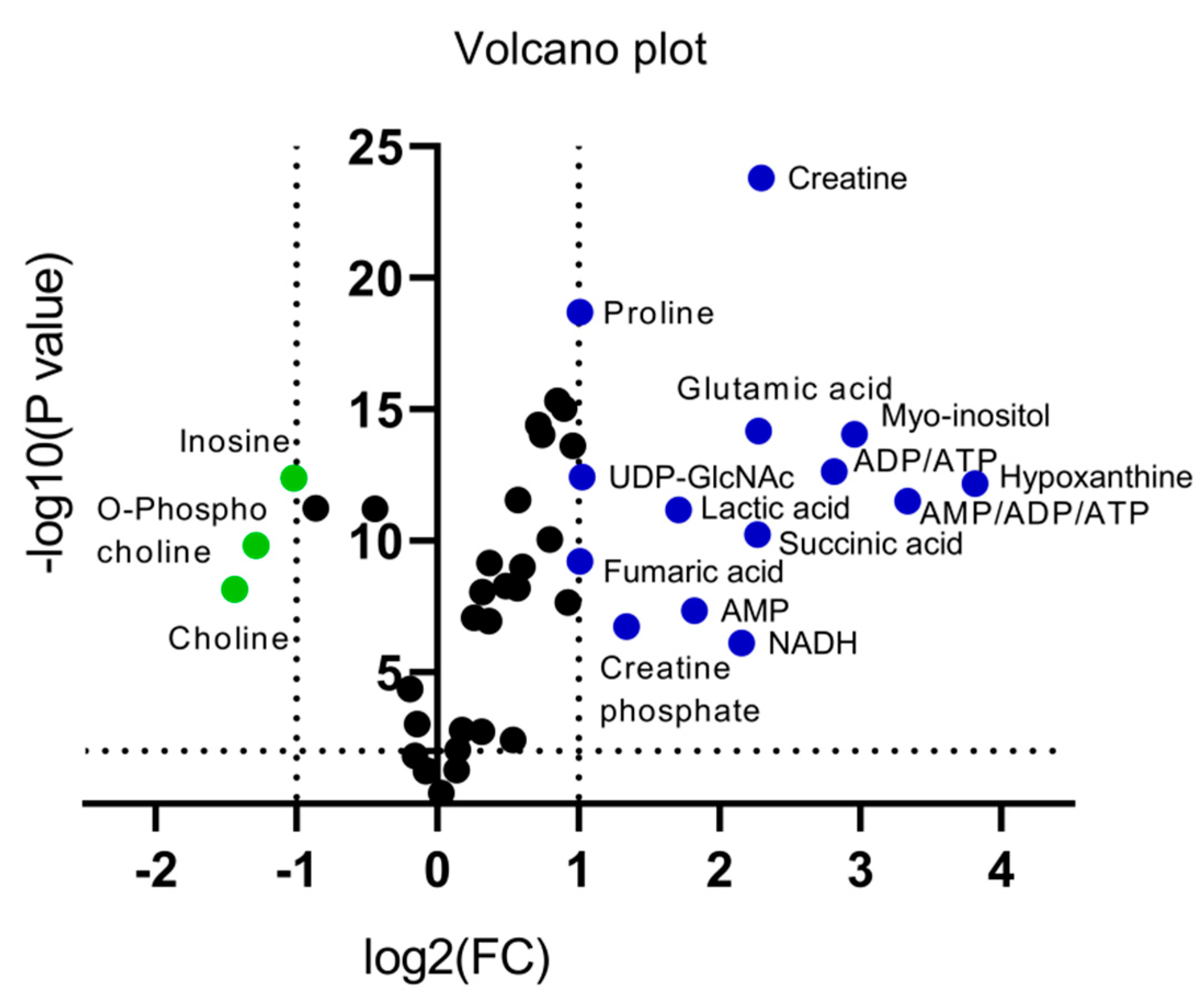
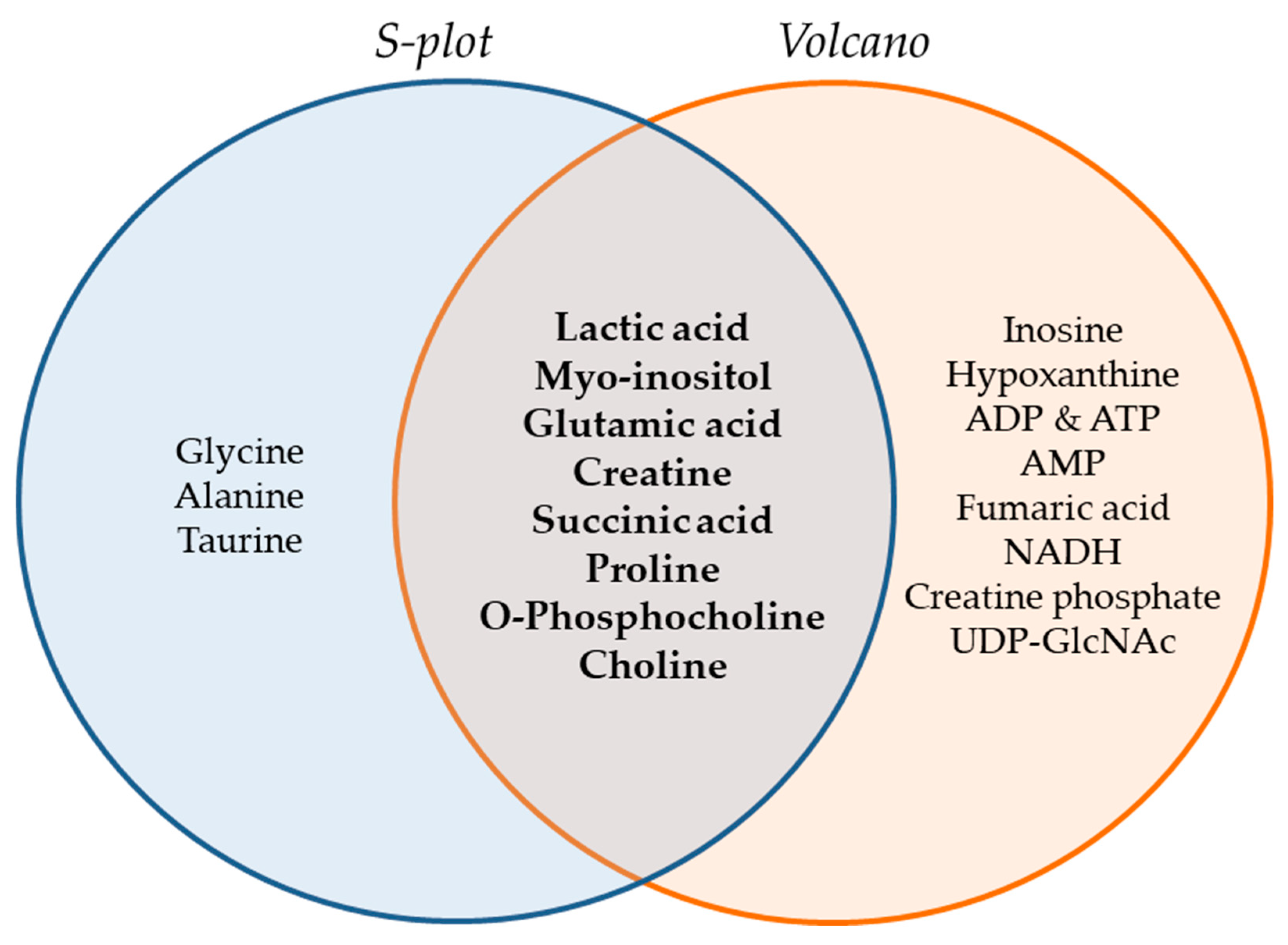
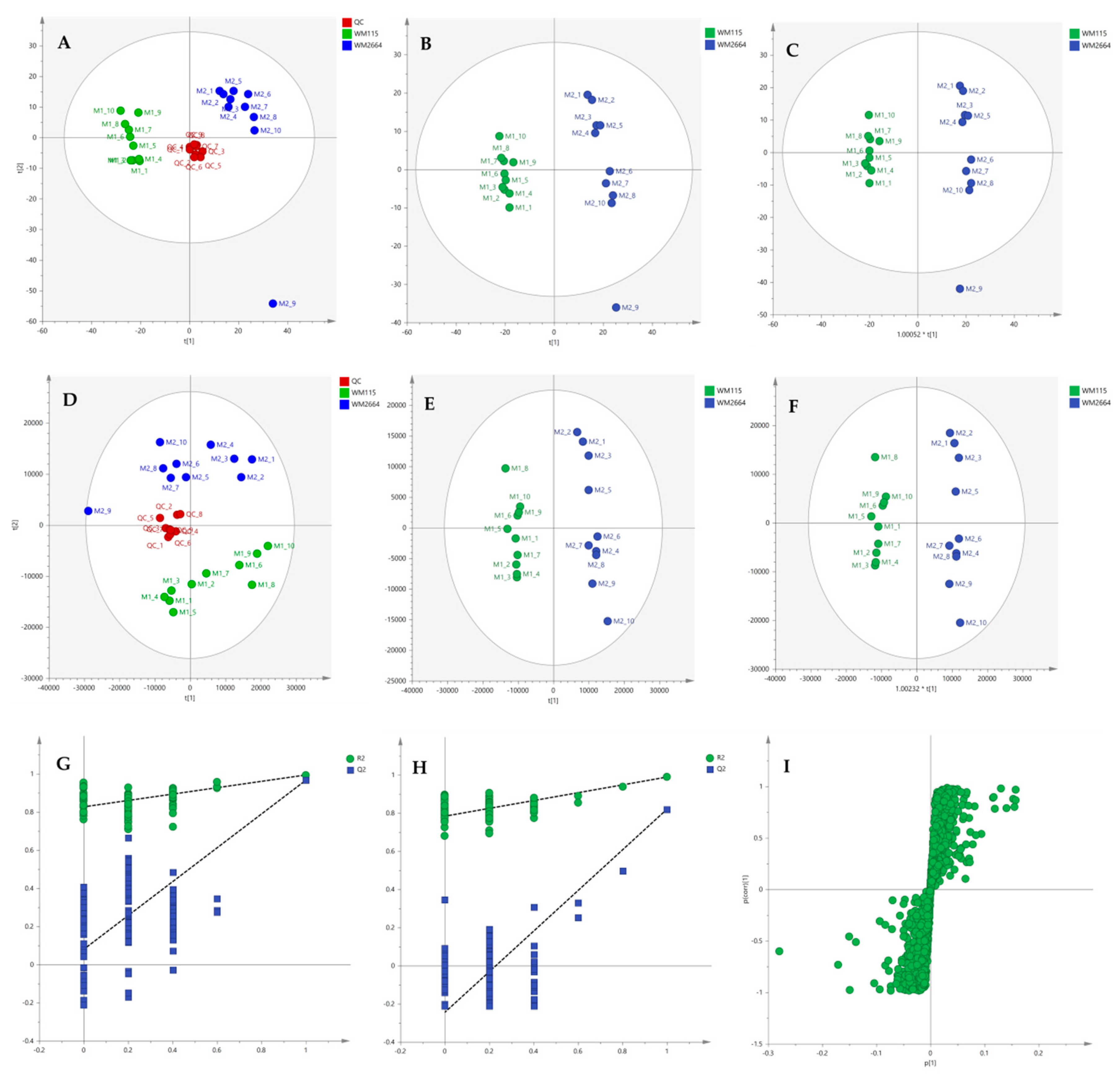
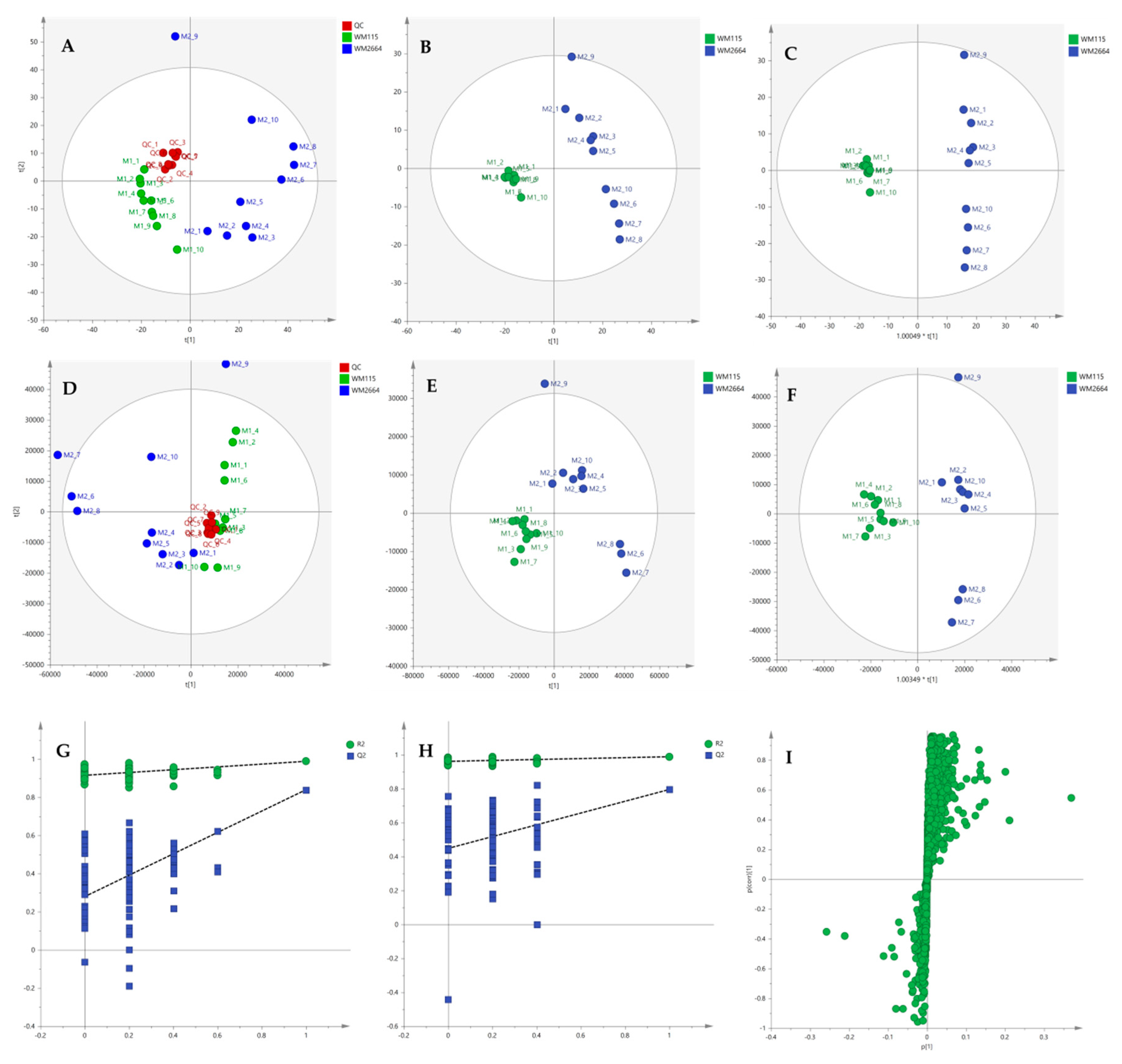
2.2.2. Chemometric Analysis of MS Data
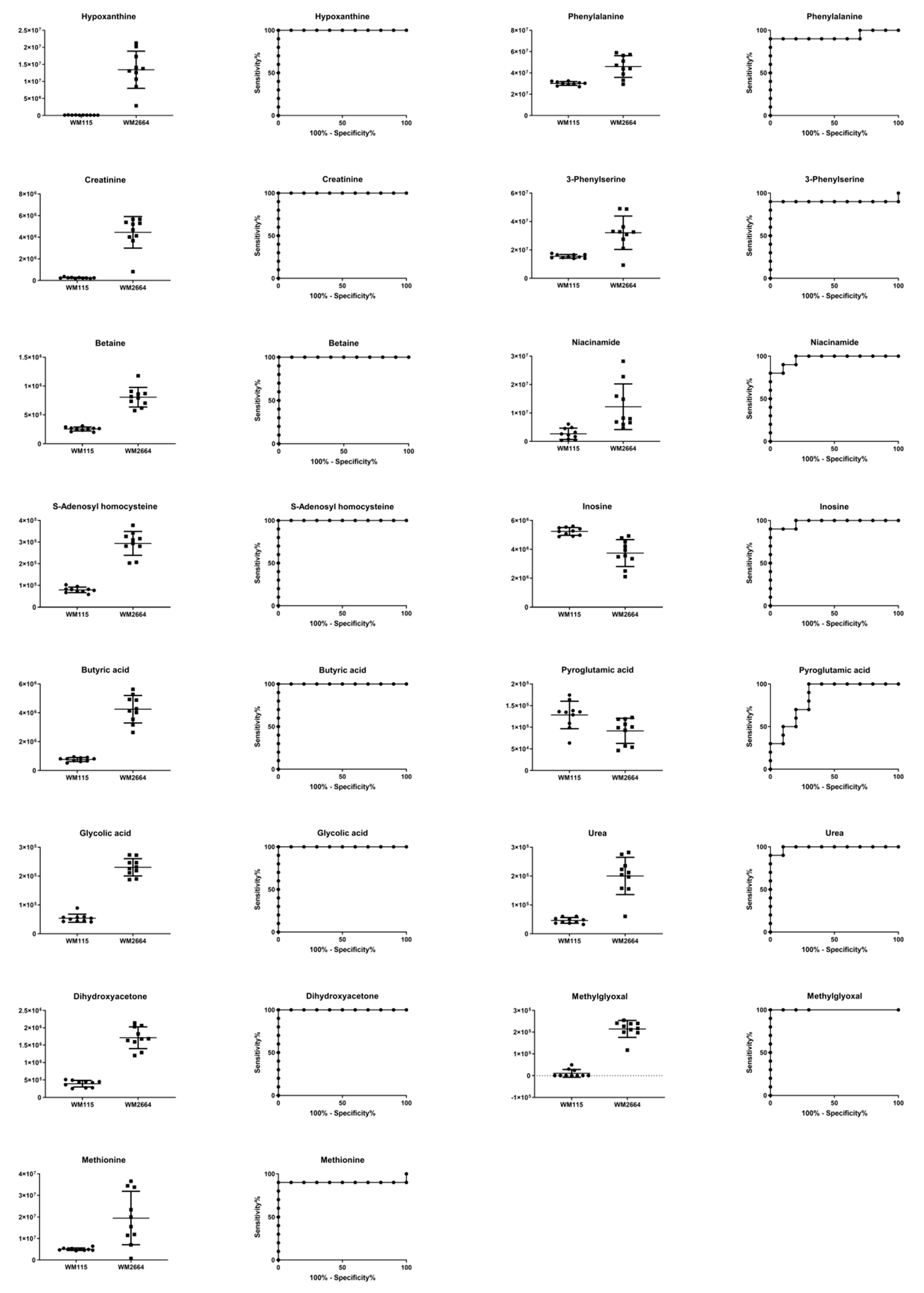
2.2.3. Identification of Selected MS Features
2.2.4. Targeted Metabolite Screening Based on Biochemical Pathways
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Lines and Culture Conditions
4.3. Cell Collection and Storage
4.4. Extraction Protocol
4.5. Sample Preparation
4.6. NMR Analysis
4.7. MS Analysis
4.8. Data Pre-Processing
4.8.1. NMR
4.8.2. MS
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ali, Z.; Yousaf, N.; Larkin, J. Melanoma epidemiology, biology and prognosis. EJC Suppl. 2013, 11, 81–91. [Google Scholar] [CrossRef]
- Cummins, D.L.; Cummins, J.M.; Pantle, H.; Silverman, M.A.; Leonard, A.L.; Chanmugam, A. Cutaneous Malignant Melanoma. Mayo Clin. Proc. 2006, 81, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Ratnikov, B.I.; Scott, D.A.; Osterman, A.L.; Smith, J.W.; Ronai, Z.A. Metabolic rewiring in melanoma. Oncogene 2017, 36, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Beger, R.D. A Review of Applications of Metabolomics in Cancer. Metabolites 2013, 3, 552–574. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.K. Global systems biology, personalized medicine and molecular epidemiology. Mol. Syst. Biol. 2006, 2, 52. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. “Metabonomics”: Understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Lindon, J.C. Systems biology: Metabonomics. Nature 2008, 455, 1054–1056. [Google Scholar] [CrossRef]
- Zhang, A.; Sun, H.; Xu, H.; Qiu, S.; Wang, X. Cell Metabolomics. OMICS 2013, 17, 495–501. [Google Scholar] [CrossRef]
- Griffin, J.L.; Shockcor, J.P. Metabolic profiles of cancer cells. Nat. Rev. Cancer 2004, 4, 551–561. [Google Scholar] [CrossRef]
- Scott, D.A.; Richardson, A.D.; Filipp, F.V.; Knutzen, C.A.; Chiang, G.G.; Ronai, Z.A.; Osterman, A.L.; Smith, J.W. Comparative Metabolic Flux Profiling of Melanoma Cell Lines. J. Biol. Chem. 2011, 286, 42626–42634. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Lee, H.; Kim, S.-H.; Jin, H.; Bae, J.; Choi, H.-K. Discovery of potential biomarkers in human melanoma cells with different metastatic potential by metabolic and lipidomic profiling. Sci. Rep. 2017, 7, 8864. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo Rodrigues, M.; Obre, E.; Melo, F.; Santos, G.; Galina, A.; Jasiulionis, M.; Rossignol, R.; Rumjanek, F.; Amoêdo, N. Enhanced OXPHOS, glutaminolysis and beta-oxidation constitute the metastatic phenotype of melanoma cells. Biochem. J. 2015, 473. [Google Scholar]
- Fedele, T.A.; Galdos-Riveros, A.C.; Jose de Farias e Melo, H.; Magalhães, A.; Maria, D.A. Prognostic relationship of metabolic profile obtained of melanoma B16F10. Biomed. Pharmacother. 2013, 67, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, G.K.; Yeow, C.H. Proton NMR characterization of intact primary and metastatic melanoma cells in 2D & 3D cultures. Biol. Res. 2017, 50, 12. [Google Scholar] [PubMed]
- Yu, Z.; Huang, M.; Clowers, B.H. Comparative metabolite profiling of a metastatic and primary melanoma cell line using untargeted metabolomics: A case study. Clin. Mass Spectrom. 2018, 10, 16–24. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef]
- Cui, X.; Churchill, G.A. Statistical tests for differential expression in cDNA microarray experiments. Genome Biol. 2003, 4, 210. [Google Scholar] [CrossRef]
- De Hertogh, B.; De Meulder, B.; Berger, F.; Pierre, M.; Bareke, E.; Gaigneaux, A.; Depiereux, E. A benchmark for statistical microarray data analysis that preserves actual biological and technical variance. BMC Bioinform. 2010, 11, 17. [Google Scholar] [CrossRef]
- Katajamaa, M.; Orešič, M. Data processing for mass spectrometry-based metabolomics. J. Chromatogr. A 2007, 1158, 318–328. [Google Scholar] [CrossRef]
- Alonso, A.; Marsal, S.; Julià, A. Analytical Methods in Untargeted Metabolomics: State of the Art in 2015. Front. Bioeng. Biotechnol. 2015, 3, 23. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling The False Discovery Rate—A Practical And Powerful Approach To Multiple Testing. J. Royal Statist. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Benjamini, Y.; Krieger, A.M.; Yekutieli, D. Adaptive linear step-up procedures that control the false discovery rate. Biometrika 2006, 93, 491–507. [Google Scholar] [CrossRef]
- Uppal, K.; Walker, D.I.; Jones, D.P. xMSannotator: An R package for network-based annotation of high-resolution metabolomics data. Anal. Chem. 2017, 89, 1063–1067. [Google Scholar] [CrossRef] [PubMed]
- R: The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 29 September 2018).
- Smith, C.A.; O’Maille, G.; Want, E.J.; Qin, C.; Trauger, S.A.; Brandon, T.R.; Custodio, D.E.; Abagyan, R.; Siuzdak, G. METLIN: A metabolite mass spectral database. Ther. Drug Monit. 2005, 27, 747–751. [Google Scholar] [CrossRef] [PubMed]
- EUR-Lex, Access to European Union Law, 2002/657/EC. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?qid=1585663763029&uri=CELEX:32002D0657 (accessed on 31 March 2020).
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Kroemer, G.; Pouyssegur, J. Tumor Cell Metabolism: Cancer’s Achilles’ Heel. Cancer Cell 2008, 13, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Romero-Garcia, S.; Moreno-Altamirano, M.M.B.; Prado-Garcia, H.; Sánchez-García, F.J. Lactate Contribution to the Tumor Microenvironment: Mechanisms, Effects on Immune Cells and Therapeutic Relevance. Front Immunol. 2016, 7, 52. [Google Scholar] [CrossRef]
- Warburg, O. On the Origin of Cancer Cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef]
- Antognelli, C.; Talesa, V.N. Glyoxalases in Urological Malignancies. Int. J. Mol. Sci. 2018, 19, 415. [Google Scholar] [CrossRef]
- Bair, W.B.; Cabello, C.M.; Uchida, K.; Bause, A.S.; Wondrak, G.T. GLO1 Overexpression in Human Malignant Melanoma. Melanoma Res. 2010, 20, 85–96. [Google Scholar] [CrossRef]
- Filipp Fabian, V.; Ratnikov, B.; De Ingeniis, J.; Smith Jeffrey, W.; Osterman Andrei, L.; Scott David, A. Glutamine-fueled mitochondrial metabolism is decoupled from glycolysis in melanoma. Pigment Cell Melanoma Res. 2012, 25, 732–739. [Google Scholar] [CrossRef] [PubMed]
- De Ingeniis, J.; Ratnikov, B.; Richardson, A.D.; Scott, D.A.; Aza-Blanc, P.; De, S.K.; Kazanov, M.; Pellecchia, M.; Ronai, Z.; Osterman, A.L.; et al. Functional Specialization in Proline Biosynthesis of Melanoma. PLoS ONE 2012, 7, 5190. [Google Scholar] [CrossRef] [PubMed]
- Dillon, B.J.; Prieto, V.G.; Curley, S.A.; Ensor, C.M.; Holtsberg, F.W.; Bomalaski, J.S.; Clark, M.A. Incidence and distribution of argininosuccinate synthetase deficiency in human cancers. Cancer 2004, 100, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Locasale, J.W.; Grassian, A.R.; Melman, T.; Lyssiotis, C.A.; Mattaini, K.R.; Bass, A.J.; Heffron, G.; Metallo, C.M.; Muranen, T.; Sharfi, H.; et al. Phosphoglycerate dehydrogenase diverts glycolytic flux and contributes to oncogenesis. Nat. Genet. 2011, 43, 869–874. [Google Scholar] [CrossRef]
- Locasale, J.W. Serine, glycine and the one-carbon cycle: Cancer metabolism in full circle. Nat. Rev. Cancer 2013, 13, 572–583. [Google Scholar] [CrossRef]
- Bessman, S.P.; Carpenter, C.L. The Creatine-Creatine Phosphate Energy Shuttle. Annu. Rev. Biochem. 1985, 54, 831–862. [Google Scholar] [CrossRef]
- de Santana-Filho, A.P.; Jacomasso, T.; Riter, D.S.; Barison, A.; Iacomini, M.; Winnischofer, S.M.B.; Sassaki, G.L. NMR metabolic fingerprints of murine melanocyte and melanoma cell lines: Application to biomarker discovery. Sci. Rep. 2017, 7, 42324. [Google Scholar] [CrossRef]
- Granovsky, M.; Fata, J.; Pawling, J.; Muller, W.J.; Khokha, R.; Dennis, J.W. Suppression of tumor growth and metastasis in Mgat5-deficient mice. Nat. Med. 2000, 6, 306–312. [Google Scholar] [CrossRef]
- Dennis, J.W.; Laferté, S. Oncodevelopmental Expression of—GlcNAcβ1–6Manα1-6Manβ1—Branched Asparagine-linked Oligosaccharides in Murine Tissues and Human Breast Carcinomas. Cancer Res. 1989, 49, 945–950. [Google Scholar]
- Sasai, K.; Ikeda, Y.; Fujii, T.; Tsuda, T.; Taniguchi, N. UDP-GlcNAc concentration is an important factor in the biosynthesis of β1,6-branched oligosaccharides: Regulation based on the kinetic properties of N-acetylglucosaminyltransferase V. Glycobiology 2002, 12, 119–127. [Google Scholar] [CrossRef]
- Croze, M.L.; Soulage, C.O. Potential role and therapeutic interests of myo-inositol in metabolic diseases. Biochimie 2013, 95, 1811–1827. [Google Scholar] [CrossRef] [PubMed]
- Novogrodskyl, A.; Nordenberg, J. The Anti-Proliferative Effect of Lithium Chloride on Melanoma Cells and Its Reversion by Myo-Inositol. Available online: https://www.researchgate.net/figure/The-effect-of-myo-inositol-on-tumorigenicity-of-untreated-and-LiCI-pretreated-B-16_fig1_20126741 (accessed on 16 December 2018).
- Ripps, H.; Shen, W. Review: Taurine: A “very essential” amino acid. Mol. Vis. 2012, 18, 2673–2686. [Google Scholar] [PubMed]
- Liesenfeld, D.B.; Habermann, N.; Owen, R.W.; Scalbert, A.; Ulrich, C.M. Review of Mass Spectrometry–Based Metabolomics in Cancer Research. Cancer Epidemiol. Biomark. Prev. 2013, 22, 2182–2201. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Xue, R.; Tang, Z.; Deng, C.; Liu, T.; Zeng, H.; Sun, Y.; Shen, X. Metabolomic investigation of gastric cancer tissue using gas chromatography/mass spectrometry. Anal. Bioanal. Chem. 2010, 396, 1385–1395. [Google Scholar] [CrossRef] [PubMed]
- SMPDB v.2.0, Purine Metabolism Pathway. Available online: http://smpdb.ca/view/SMP0000050 (accessed on 31 March 2020).
- Sugimoto, M.; Wong, D.T.; Hirayama, A.; Soga, T.; Tomita, M. Capillary electrophoresis mass spectrometry-based saliva metabolomics identified oral, breast and pancreatic cancer-specific profiles. Metabolomics 2010, 6, 78–95. [Google Scholar] [CrossRef]
- Wang, X.; Hu, M.; Liu, M.; Hu, J. Metastatic Melanoma Induced Metabolic Changes in C57BL/6J Mouse Stomach Measured by 1H NMR Spectroscopy. Metabolomics (Los Angel) 2014, 4, 1000135. [Google Scholar]
- Galons, J.P.; Job, C.; Gillies, R.J. Increase of GPC levels in cultured mammalian cells during acidosis. A 31P MR spectroscopy study using a continuous bioreactor system. Magn. Reson. Med. 1995, 33, 422–426. [Google Scholar] [CrossRef]
- Holeček, M. Branched-chain amino acids in health and disease: Metabolism, alterations in blood plasma, and as supplements. Nutr. Metab. 2018, 15, 33. [Google Scholar] [CrossRef]
- 7-Hydroxy-6-methyl-8-ribityl lumazine-Biomarker|BiomarkerBase. Available online: https://www.biomarkerbase.com/7-hydroxy-6-methyl-8-ribityl-lumazine?utm_source=adis (accessed on 26 January 2020).
- SMPDB v.2.0, Riboflavin Metabolism Pathway. Available online: http://smpdb.ca/view/SMP0000070 (accessed on 31 March 2020).
- Li, X.; Zhang, A.; Sun, H.; Liu, Z.; Zhang, T.; Qiu, S.; Liu, L.; Wang, X. Metabolic characterization and pathway analysis of berberine protects against prostate cancer. Oncotarget 2017, 8, 65022–65041. [Google Scholar] [CrossRef]
- Wu, H.; Southam, A.D.; Hines, A.; Viant, M.R. High-throughput tissue extraction protocol for NMR- and MS-based metabolomics. Anal. Biochem. 2008, 372, 204–212. [Google Scholar] [CrossRef]
- Goodacre, R.; Broadhurst, D.; Smilde, A.K.; Kristal, B.S.; Baker, J.D.; Beger, R.; Bessant, C.; Connor, S.; Capuani, G.; Craig, A.; et al. Proposed minimum reporting standards for data analysis in metabolomics. Metabolomics 2007, 3, 231–241. [Google Scholar] [CrossRef]
- Wei, X.; Shi, X.; Kim, S.; Zhang, L.; Patrick, J.S.; Binkley, J.; McClain, C.; Zhang, X. Data preprocessing method for liquid chromatography-mass spectrometry based metabolomics. Anal. Chem. 2012, 84, 7963–7971. [Google Scholar] [CrossRef]
- Cho, H.W.; Kim, S.B.; Jeong, M.K.; Park, Y.; Miller, N.G.; Ziegler, T.R.; Jones, D.P. Discovery of metabolite features for the modelling and analysis of high-resolution NMR spectra., Discovery of metabolite features for the modelling and analysis of high-resolution NMR spectra. Int. J. Data Min. Bioinform. 2008, 2, 176–192. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.; Cloarec, O.; Holmes, E.; Nicholson, J.K.; Lindon, J.C. Scaling and normalization effects in NMR spectroscopic metabonomic data sets. Anal. Chem. 2006, 78, 2262–2267. [Google Scholar] [CrossRef] [PubMed]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Mei, Z.; Zeng, C.; Liu, S. metaX: A flexible and comprehensive software for processing metabolomics data. BMC Bioinform. 2017, 18, 183. [Google Scholar] [CrossRef]
- Myers, O.D.; Sumner, S.J.; Li, S.; Barnes, S.; Du, X. One Step Forward for Reducing False Positive and False Negative Compound Identifications from Mass Spectrometry Metabolomics Data: New Algorithms for Constructing Extracted Ion Chromatograms and Detecting Chromatographic Peaks. Anal. Chem. 2017, 89, 8696–8703. [Google Scholar] [CrossRef]
- Du, P.; Kibbe, W.A.; Lin, S.M. Improved peak detection in mass spectrum by incorporating continuous wavelet transform-based pattern matching. Bioinformatics 2006, 22, 2059–2065. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
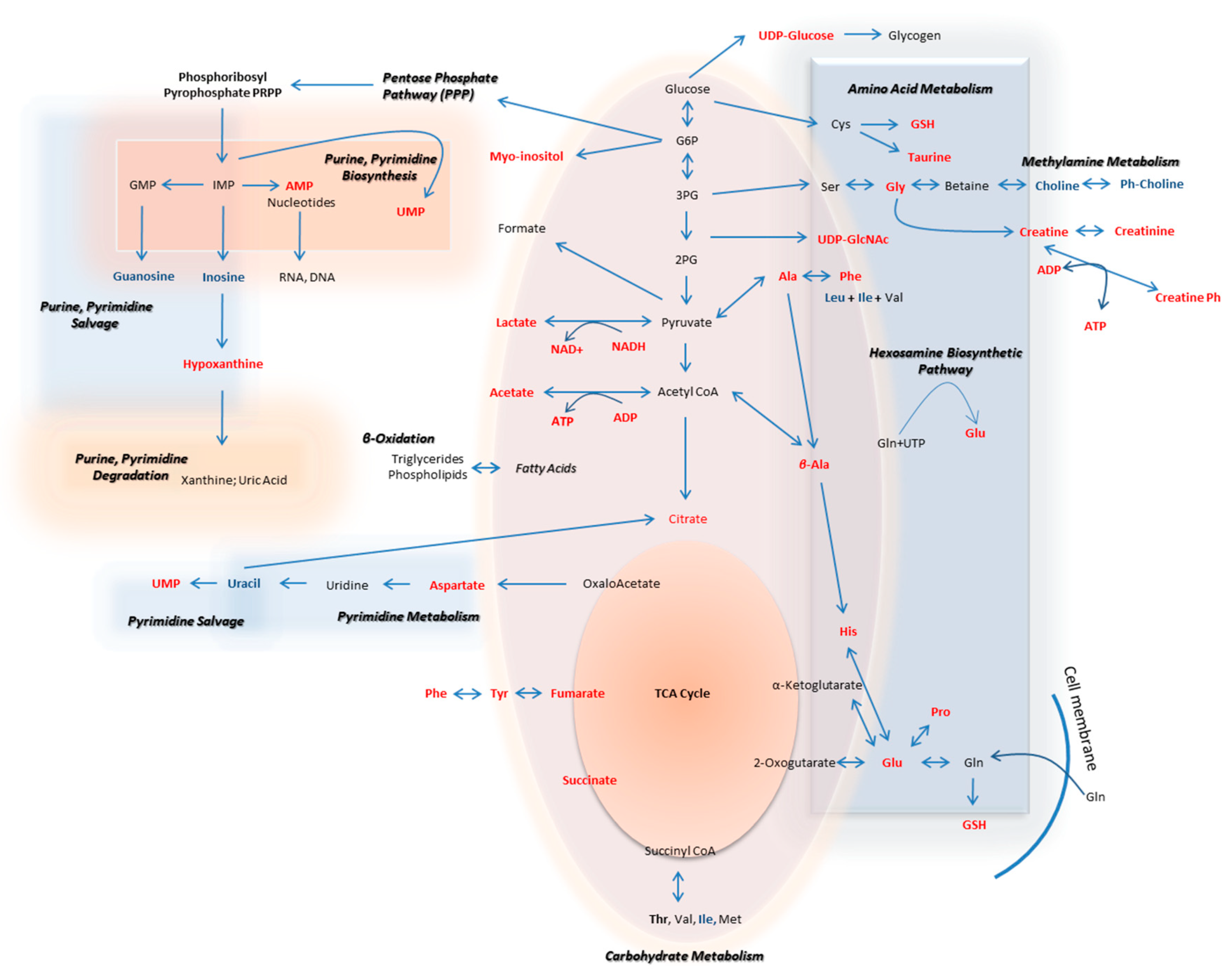{kind=link}
| Metabolites Elevated (↑) in WM115 | Metabolites Elevated (↑) in WM2664 | ||||
|---|---|---|---|---|---|
| Name | S-plot (δ) | VIP | Name | S-plot (δ) | VIP |
| o-Phosphocholine | 3.23 | 2.18 | Lactic acid | 1.33 | 8.36 |
| Choline | 3.21 | 1.34 | Myo-inositol | 3.55 | 4.60 |
| Inosine | 8.35 | 0.91 | Myo-inositol | 4.07 | 4.15 |
| Inosine | 6.11 | 0.87 | Myo-inositol | 3.63 | 4.12 |
| Leucine | 0.97 | 0.83 | Lactic acid | 4.11 | 3.83 |
| Isoleucine | 1.27 | 0.70 | Myo-inositol | 3.53 | 3.76 |
| Myo-inositol | 3.29 | 3.29 | |||
| Myo-inositol | 3.61 | 3.23 | |||
| Glutamic acid | 2.35 | 3.17 | |||
| Creatine | 3.93 | 3.16 | |||
| Creatine | 3.03 | 2.92 | |||
| Creatine | 3.05 | 2.92 | |||
| Myo-inositol | 3.65 | 2.66 | |||
| Glutamic acid | 3.77 | 2.40 | |||
| Myo-inositol | 3.27 | 2.35 | |||
| Succinic acid | 2.41 | 2.21 | |||
| Glutamic acid | 2.37 | 2.13 | |||
| Glutamic acid | 2.07 | 2.12 | |||
| Lactic acid | 4.13 | 2.07 | |||
| Proline | 3.35 | 2.06 | |||
| Glutamic acid | 3.79 | 1.90 | |||
| Glutamic acid | 2.13 | 1.86 | |||
| Glutamic acid | 2.09 | 1.79 | |||
| Glutamic acid | 3.75 | 1.77 | |||
| Glutamic acid | 2.05 | 1.77 | |||
| Lactic acid | 4.09 | 1.75 | |||
| Glutamic acid | 2.15 | 1.67 | |||
| Glycine | 3.57 | 1.66 | |||
| Glutamic acid | 2.33 | 1.65 | |||
| Alanine | 1.49 | 1.64 | |||
| Taurine and Proline 1 | 3.43 | 1.62 | |||
| Myo-inositol | 3.31 | 1.61 | |||
| Glutamic acid | 2.11 | 1.51 | |||
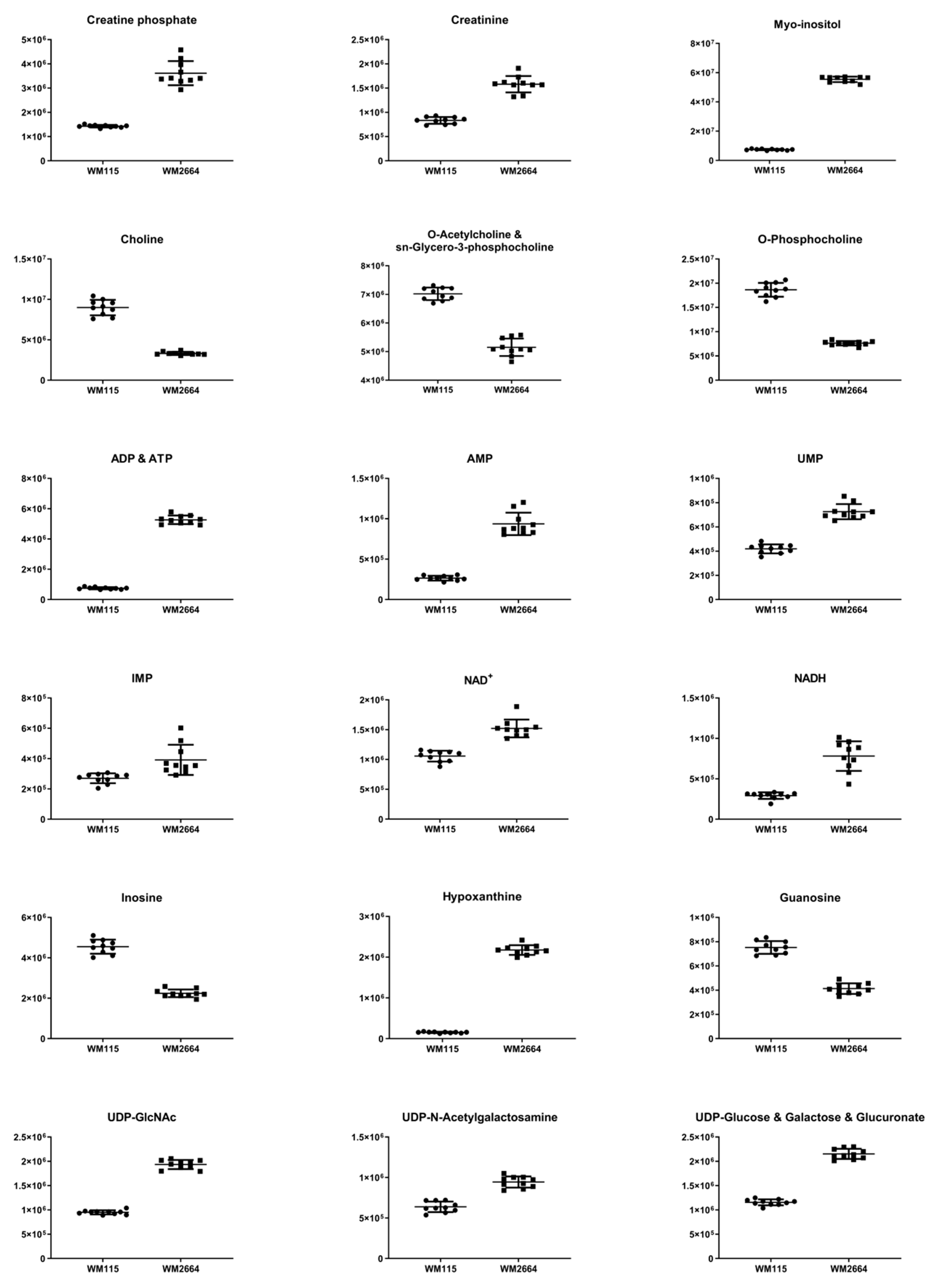
| Metabolite (bold: AUC ≥ 0.9 and adjusted p < 0.05) | Outliers (Grubbs) | Normality test (D’ Agostino and Pearson) | F-Test Significantly Different Variance (p < 0.05) | Welch’s t-Test (p Value) | t-Test (p Value) | Bonferroni–Dunn (Adjusted p Value) | z-Score | Fold Change (x) | AUC | Tendency |
|---|---|---|---|---|---|---|---|---|---|---|
| Alanine | 0 | √ | no | 4.77E−16 | 2.16E−14 | 11.09 | 1.80 | 1 | ↑WM2664 | |
| β-Alanine | 0 | √ | no | 2.38E−14 | 1.08E−12 | 9.52 | 1.95 | 1 | ↑WM2664 | |
| Aspartic acid | 0 | √ | yes | 1.12E−07 | 5.00E−06 | 11.50 | 1.29 | 1 | ↑WM2664 | |
| Glutamic acid | 0 | √ | yes | 6.68E−15 | 3.02E−13 | 52.93 | 4.84 | 1 | ↑WM2664 | |
| Glutathione | 0 | √ | no | 3.98E−15 | 1.80E−13 | 13.12 | 1.64 | 1 | ↑WM2664 | |
| Glycine | 0 | √ | no | 2.74E−12 | 1.21E−10 | 6.43 | 1.49 | 1 | ↑WM2664 | |
| Histidine | 0 | √ | no | 9.08E−09 | 4.09E−07 | 4.81 | 1.25 | 1 | ↑WM2664 | |
| Isoleucine | 0 | √ | no | 0.000911 | 0.0409 | −1.43 | 0.91 | 0.9 | ↑WM115 | |
| Leucine | 0 | √ | no | 4.22E−05 | 0.00189 | −1.94 | 0.87 | 0.96 | ↑WM115 | |
| Phenylalanine | 0 | √ | no | 8.58E−08 | 4.00E−06 | 3.45 | 1.20 | 1 | ↑WM2664 | |
| Proline | 0 | √ | no | 1.96E−19 | 9.00E−18 | 18.52 | 2.01 | 1 | ↑WM2664 | |
| Serine | 0 | √ | no | 0.394 | 1 | 0.39 | 1.02 | 0.6 | ||
| Taurine | 0 | √ | no | 9.89E−10 | 4.45E−08 | 7.08 | 1.52 | 1 | ↑WM2664 | |
| Threonine | 0 | √ | no | 0.0585 | 1 | −0.73 | 0.94 | 0.73 | ||
| Tryptophan | 0 | √ | no | 0.00159 | 0.0720 | 1.34 | 1.13 | 0.88 | ||
| Tyrosine | 0 | √ | no | 6.85E−10 | 3.11E−08 | 4.74 | 1.29 | 1 | ↑WM2664 | |
| Valine | 0 | √ | no | 0.0554 | 1 | −0.74 | 0.95 | 0.74 | ↑WM115 | |
| Creatine | 0 | √ | no | 1.59E−24 | 7.20E−23 | 54.01 | 4.91 | 1 | ↑WM2664 | |
| Creatine phosphate | 0 | √ | yes | 1.8E−07 | 8.00E−06 | 41.57 | 2.53 | 1 | ↑WM2664 | |
| Creatinine | 0 | √ | yes | 2.19E−08 | 9.90E−07 | 10.70 | 1.90 | 1 | ↑WM2664 | |
| Acetic acid | 0 | √ | no | 0.00870 | 0.392 | 1.47 | 1.10 | 0.83 | ||
| Formic acid | 0 | √ | yes | 0.00180 | 0.0810 | 1.37 | 1.24 | 0.85 | ||
| Fumaric acid | 0 | √ | no | 6.03E−10 | 2.70E−08 | 8.10 | 2.01 | 1 | ↑WM2664 | |
| Lactic acid | 0 | √ | yes | 6.79E−12 | 3.06E−10 | 32.51 | 3.27 | 1 | ↑WM2664 | |
| Succinic acid | 0 | √ | yes | 5.63E−11 | 2.52E−09 | 49.00 | 4.82 | 1 | ↑WM2664 | |
| Choline | 0 | √ | yes | 7.02E−09 | 3.15E−07 | −5.93 | 0.37 | 1 | ↑WM115 | |
| sn-Glycero-3-phosphocholine & O-Acetylcholine | 0 | √ | no | 6.01E−12 | 2.70E−10 | −8.45 | 0.73 | ↑WM115 | ||
| O-Phosphocholine | 0 | √ | yes | 1.52E−10 | 8.55E−17 | −7.68 | 0.41 | 1 | ↑WM115 | |
| AMP | 0 | √ | yes | 4.52E−08 | 2.00E−06 | 23.14 | 3.54 | 1 | ↑WM2664 | |
| ADP & ATP | 0 | √ | yes | 2.36E−13 | 1.08E−11 | 64.99 | 7.04 | 1 | ↑WM2664 | |
| Guanosine | 0 | √ | no | 5.86E−12 | 2.66E−10 | −6.45 | 0.55 | 1 | ↑WM115 | |
| Hypoxanthine | 0 | √ | yes | 6.81E−13 | 3.06E−11 | 131.73 | 14.05 | 1 | ↑WM2664 | |
| Inosine | 0 | √ | no | 4.06E−13 | 1.85E−11 | −6.63 | 0.49 | 1 | ↑WM115 | |
| IMP | 0 | √ | yes | 0.00381 | 0.171 | 3.74 | 1.45 | 0.97 | ↑WM2664 | |
| NAD+ | 0 | √ | no | 5.17E−09 | 2.34E−07 | 4.63 | 1.40 | 1 | ↑WM2664 | |
| NADH | 0 | √ | yes | 7.67E−07 | 0.000035 | 21.87 | 4.46 | 1 | ↑WM2664 | |
| Uracil | 0 | √ | no | 0.0149 | 0.671 | −0.97 | 0.89 | 0.82 | ↑WM115 | |
| UDP-N-Acetylglucosamine | 0 | √ | yes | 3.66E−13 | 1.66E−11 | 23.04 | 2.04 | 1 | ↑WM2664 | |
| UDP-Galactose & Glucose & Glucuronic acid | 0 | √ | no | 9.18E−16 | 4.14E−14 | 16.21 | 1.86 | 1 | ↑WM2664 | |
| UDP-N-Acetylgalactosamine | 0 | √ | no | 6.31E−09 | 2.84E−07 | 4.73 | 1.48 | 1 | ↑WM2664 | |
| UMP | 0 | √ | no | 8.74E−11 | 3.91E−09 | 8.33 | 1.73 | 1 | ↑WM2664 | |
| Myo-inositol | 0 | √ | yes | 8.72E−15 | 3.91E−13 | 101.96 | 7.76 | 1 | ↑WM2664 | |
| Niacinamide | 0 | √ | no | 9.13E−15 | 4.10E−13 | 10.58 | 1.67 | 1 | ↑WM2664 |
| Name | Adducts | Theoretical m/z | Experimental m/z | ppm Error | tR (min) | Adducts | MS/MS | Isotopic Ratio RSD (%) | p Value | FC | Trust Level | ESI |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 8-Hydroxy-deoxyguanosine or Guanosine | M-H | 282.084 | 282.0839 | −1.86 | 1.14 | M+FA-H | 0 | <30% | 1.51E−13 | 0.42 | 4.5 | (-) |
| Malic acid | M-H | 133.014 | 133.0146 | 3.05 | 0.74 | - | 114.91 | <30% | 3.33E−11 | 10.74 | 4 | (-) |
| 7-Hydroxy-6- methyl-8-ribityl lumazine | M+K-2H | 365.051 | 365.0495 | −2.66 | 1.17 | - | 267.06 | <30% | 1.81E−13 | 0.53 | 4 | (-) |
| beta-Citryl-l- glutamic acid | M-H | 320.062 | 320.0621 | −0.54 | 0.85 | - | 0 | <30% | 1.29E−08 | 18.93 | 2.5 | (-) |
| S-pyruvyl glutathione | M+FA-H | 422.088 | 422.0868 | −1.66 | 0.99 | - | 0 | <30% | 3.92E−08 | 22.79 | 2.5 | (-) |
| Oxidized glutathione | M-H | 611.145 | 611.1432 | −2.40 | 0.99 | - | 0 | <30% | 3.43E−09 | 2.36 | 2.5 | (-) |
| 3-Phenylserine | M + H | 182.0812 | 182.0812 | 0.44 | 0.94 | - | 136, 165, 147 | <30% | 0.00153 | 2.07 | 7 | (+) |
| Methionine | M+H | 150.0582 | 150.0582 | −0.99 | 0.86 | - | 133, 104 | <30% | 0.00495 | 3.89 | 5.5 | (+) |
| Phenylalanine | M+H | 166.0862 | 166.0862 | −0.13 | 2.05 | - | 120, 149 | <30% | 0.000736 | 1.53 | 5.5 | (+) |
| Niacinamide | M+H | 123.055 | 123.0550 | −2.02 | 1.05 | - | 80 | <30% | 0.00448 | 4.56 | 4 | (+) |
| Name | Adducts | Theoretical m/z | Experimental m/z | ppm Error | tR (min) | Adducts | MS/MS | Isotopic Ratio RSD (%) | p Value | FC | Trust Level |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Inosine | M-H | 267.0730 | 267.0731 | 0.21 | 1.16 | M+Cl, M+FA-H, 2M+FA-H, 2M-H | 134.93, 134.98, 135.04 | <30% | 7.95E−15 | 0.46 | 13.5 |
| Succinic acid | M-H | 117.0187 | 117.0196 | 7.69 | 1.11 | 99, 73 | <30% | 0.000302 | 4.04 | 5.5 | |
| Allantoic acid | 2M-H | 351.1010 | 351.1008 | −0.51 | 4.54 | 307.11 (pseudo) | <30% | 0.000339 | 0.67 | 4.5 | |
| Xanthosine | M-H | 283.0678 | 283.0678 | 0.00 | 1.50 | M+FA-H | <30% | 8.77E−06 | 2.02 | 4.5 | |
| Dihydroxyacetone | 2M-H | 179.0550 | 179.0561 | 6.14 | 0.82 | M+Cl | ND2 | 0.000297 | 32.96 | 4 | |
| S-Adenosyl- homocysteine | M-H | 383.1138 | 383.1139 | 0.26 | 0.92 | <30% | 1.13E−08 | 36.60 | 2.5 | ||
| Xylulose | M+K-2H | 187.0003 | 187.0012 | 5.05 | 0.82 | <30% | 1.31E−05 | 2.39 | 2.5 | ||
| N2-Methyl guanosine | M+K-2H | 334.0548 | 334.0553 | 1.64 | 3.29 | <30% | 0.000287 | 434068.2 | 2.5 | ||
| Xanthurenic acid | M+Na-2H | 226.0116 | 226.0122 | 2.65 | 0.84 | ND | 0.000459 | 0.83 | 2 | ||
| Glutamic acid | M-H | 146.0453 | 146.0460 | 4.79 | 0.76 | ND | 9.3E−07 | 5.81 | 2 | ||
| Acetoacetic acid | M+FA-H | 147.0300 | 147.0299 | −0.80 | 0.80 | ND | 2.67E−07 | 188.88 | 2 | ||
| N-Formyl methionine | M-H | 176.0381 | 176.0385 | 2.14 | 3.14 | ND | 0.00212 | 1.21 | 2 | ||
| Pseudouridine | M+FA-H | 289.0671 | 289.0674 | 1.04 | 0.95 | ND | 1.71E−05 | 0.62 | 2 |
| Name | Adducts | Theoretical m/z | Experimental m/z | ppm Error | tR (min) | Adducts | MS/MS | Isotopic Ratio RSD (%) | p Value | FC | Trust Level |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Inosine | M+H | 269.0880 | 269.0884 | 1.31 | 1.20 | M+K, M+Na, 2M+H | 136 | < 30% | 8.77E−14 | 0.50 | 10 |
| Hypoxanthine | M+H | 137.0458 | 137.0456 | −1.53 | 0.86 | M+2K-H, M+K | 119, 109 | < 30% | 2.51E−08 | 94.26 | 9.5 |
| Creatinine | M+H | 114.0662 | 114.0659 | −2.03 | 0.78 | M+2K-H, M+K | 86 | < 30% | 0.000462 | 6.89 | 8 |
| Betaine | M+K | 156.0421 | 156.0420 | −0.41 | 0.69 | M+2K-H | ND 2 | 1.72E−12 | 3.50 | 4 | |
| Isovaleric acid | 2M+ACN+H | 246.1700 | 246.1701 | 0.41 | 3.75 | 85 | < 30% | 0.000229 | 0.74 | 4 | |
| Propionic acid | 2M+ACN+H | 190.1074 | 190.1074 | 0 | 3.68 | M+ACN+H | ND | 2.07E−05 | 0.46 | 4 | |
| S-Adenosyl-homocysteine | M+H | 385.1289 | 385.1293 | 0.98 | 1.02 | < 30% | 1.21E−09 | 3.38 | 2.5 | ||
| Allantoic acid | 2M+H | 353.1169 | 353.1166 | −0.68 | 4.41 | < 30% | 0.01502 | 0.83 | 2.5 | ||
| Methylglyoxal | 2M+K | 183.0054 | 183.0054 | 0 | 0.76 | < 30% | 2.12E−09 | 20.70 | 2.5 | ||
| Butyric acid | 2M+ACN+H | 218.1392 | 218.1388 | −1.83 | 1.48 | < 30% | 1.22E−12 | 3.75 | 2.5 | ||
| Formic acid | M+2K+H | 122.9250 | 122.9242 | −6.22 | 0.69 | < 30% | 1.06E−05 | 1.60 | 2.5 | ||
| Dihydroxy acetone | M+2K+H | 166.9512 | 166.9507 | −3.17 | 0.71 | ND | 1.56E−07 | 2.90 | 2 | ||
| Dimethyl arginine | M+H | 203.1508 | 203.1503 | −2.59 | 0.75 | ND | 2.23E−06 | 0.17 | 2 | ||
| Pyroglutamic acid | M+2K+H | 205.9616 | 205.9617 | 0.44 | 0.72 | ND | 0.002998 | 0.60 | 2 | ||
| Glycolic acid | 2M+NH4 | 170.0659 | 170.0655 | −1.96 | 1.02 | ND | 2.99E−12 | 5.37 | 2 | ||
| Hippuric acid | M+ACN+H | 221.0921 | 221.0922 | 0.42 | 1.42 | ND | 1.47E−08 | 0.05 | 2 | ||
| Urea | M+2Na-H | 105.0035 | 105.0032 | −2.31 | 0.62 | ND | 0.000128 | 2.63 | 2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kosmopoulou, M.; Giannopoulou, A.F.; Iliou, A.; Benaki, D.; Panagiotakis, A.; Velentzas, A.D.; Konstantakou, E.G.; Papassideri, I.S.; Mikros, E.; Stravopodis, D.J.; et al. Human Melanoma-Cell Metabolic Profiling: Identification of Novel Biomarkers Indicating Metastasis. Int. J. Mol. Sci. 2020, 21, 2436. https://doi.org/10.3390/ijms21072436
Kosmopoulou M, Giannopoulou AF, Iliou A, Benaki D, Panagiotakis A, Velentzas AD, Konstantakou EG, Papassideri IS, Mikros E, Stravopodis DJ, et al. Human Melanoma-Cell Metabolic Profiling: Identification of Novel Biomarkers Indicating Metastasis. International Journal of Molecular Sciences. 2020; 21(7):2436. https://doi.org/10.3390/ijms21072436
Chicago/Turabian StyleKosmopoulou, Mariangela, Aikaterini F. Giannopoulou, Aikaterini Iliou, Dimitra Benaki, Aristeidis Panagiotakis, Athanassios D. Velentzas, Eumorphia G. Konstantakou, Issidora S. Papassideri, Emmanuel Mikros, Dimitrios J. Stravopodis, and et al. 2020. "Human Melanoma-Cell Metabolic Profiling: Identification of Novel Biomarkers Indicating Metastasis" International Journal of Molecular Sciences 21, no. 7: 2436. https://doi.org/10.3390/ijms21072436
APA StyleKosmopoulou, M., Giannopoulou, A. F., Iliou, A., Benaki, D., Panagiotakis, A., Velentzas, A. D., Konstantakou, E. G., Papassideri, I. S., Mikros, E., Stravopodis, D. J., & Gikas, E. (2020). Human Melanoma-Cell Metabolic Profiling: Identification of Novel Biomarkers Indicating Metastasis. International Journal of Molecular Sciences, 21(7), 2436. https://doi.org/10.3390/ijms21072436