In Vitro Vascular Network Modified to Function as Culture Platform and Angiogenic Induction Potential Test for Cancer Cells

 ,
,
Abstract
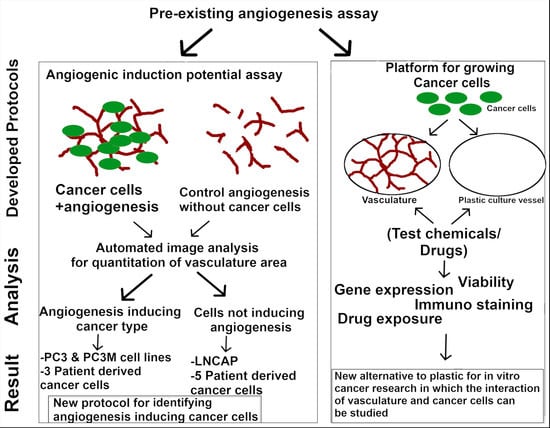
1. Introduction
2. Results
2.1. Scaling of Vascular Structures
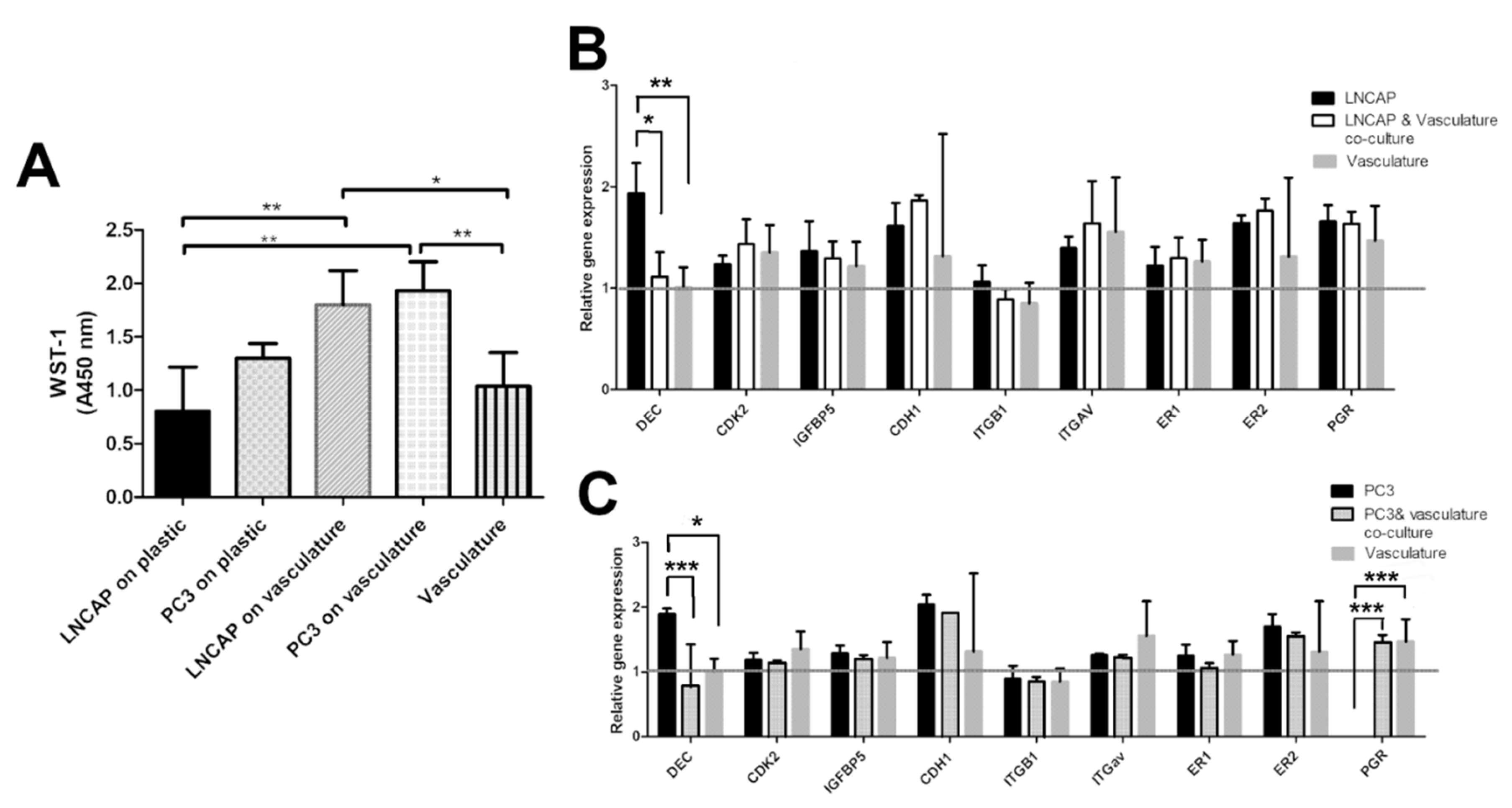
2.2. Characterization of Vascularized Cancer Assay
2.3. Responses of Cancer Cells Grown on Plastic and Those Grown on Vascular Structures
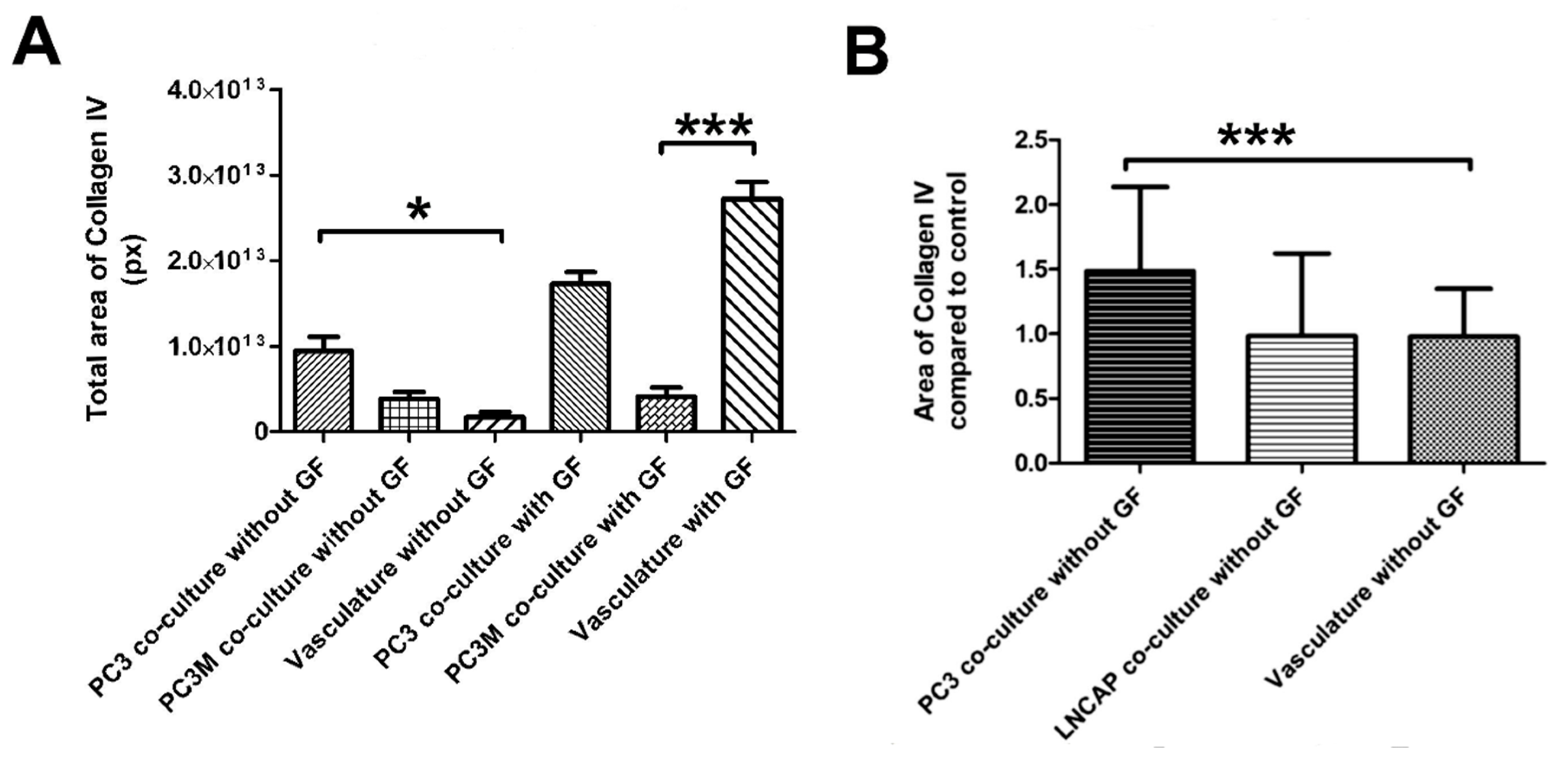
2.4. Angiogenic Induction Potential Assay for Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Ethical Considerations
4.2. The Isolation, Culture and Quality Control of Human Primary Cells hASC and HUVEC
4.3. Isolation of Primary Cancer Cells
4.4. Expansion of Cancer Cell Lines
4.5. Establishment of the Co-Culture of Cancer Cells and Vascular Network for Drug Exposure
4.6. Modifying the Co-Culture System for Testing the Angiogenic Potential of Cancer Cells
4.7. Testing the Angiogenic Potential of Patient Derived Cancer Cells
4.8. Immunofluorescence Staining
4.9. Microscopy and Quantification of Vascular Structures
4.10. PCR Analysis
4.11. Statistical Analyzes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ASM | Angiogenesis stimulating medium |
| GCM | General cancer medium |
| LCM | Liquid cancer sample medium |
| BSA | Bovine serum albumin |
| VEGF | Vascular endothelial growth factor |
| FGF-b | Fibroblast growth factor |
| hASC | human adipose stromal cell |
| HUVEC | human umbilical vein endothelial cell |
| DEC | Decorin |
| ITGB1 | Integrin B1 |
| ITGAV | Integrin av |
| ER1 | Estrogen receptor 1 |
| ER2 | Estrogen receptor 2 |
| CDH1 | E-cadherin |
| PGR | Progesterone receptor |
| 5-fu | 5-fluorouracil |
| CP | Cyclophosphamide |
| DX | Doxorubicin |
| DC | Docetaxel |
| LA | Lapatinib |
| CK19 | Cytokeratin 19 |
| EGM-2 | Endothelial Cell Growth Medium-2 BulletKit |
| VWF | Von Willebrand factor |
References
- Oehler, M.K.; Bicknell, R. The Promise of Anti-Angiogenic Cancer Therapy. Br. J. Cancer 2000, 82, 749–752. [Google Scholar] [CrossRef] [PubMed]
- Bhadriraju, K.; Chen, C.S. Engineering Cellular Microenvironments to Improve Cell-Based Drug Testing. Drug Discov. Today 2002, 7, 612–620. [Google Scholar] [CrossRef]
- Weaver, V.M.; Petersen, O.W.; Wang, F.; Larabell, C.A.; Briand, P.; Damsky, C.; Bissell, M.J. Reversion of the Malignant Phenotype of Human Breast Cells in Three-Dimensional Culture and in Vivo by Integrin Blocking Antibodies. J. Cell Biol. 1997, 137, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Edmondson, R.; Broglie, J.J.; Adcock, A.F.; Yang, L. Three-Dimensional Cell Culture Systems and their Applications in Drug Discovery and Cell-Based Biosensors. Assay Drug Dev. Technol. 2014, 12, 207–218. [Google Scholar] [CrossRef]
- Breslin, S.; O’Driscoll, L. Three-Dimensional Cell Culture: The Missing Link in Drug Discovery. Drug Discov. Today 2013, 18, 240–249. [Google Scholar] [CrossRef]
- Birgersdotter, A.; Sandberg, R.; Ernberg, I. Gene Expression Perturbation in Vitro—A Growing Case for Three-Dimensional (3D) Culture Systems. Semin. Cancer Biol. 2005, 15, 405–412. [Google Scholar] [CrossRef]
- Heinonen, T. Better Science with Human Cell-Based Organ and Tissue Models. Altern. Lab. Anim. 2015, 43, 29–38. [Google Scholar] [CrossRef]
- Shield, K.; Ackland, M.L.; Ahmed, N.; Rice, G.E. Multicellular Spheroids in Ovarian Cancer Metastases: Biology and Pathology. Gynecol. Oncol. 2009, 113, 143–148. [Google Scholar] [CrossRef]
- Zietarska, M.; Maugard, C.M.; Filali-Mouhim, A.; Alam-Fahmy, M.; Tonin, P.N.; Provencher, D.M.; Mes-Masson, A.M. Molecular Description of a 3D in Vitro Model for the Study of Epithelial Ovarian Cancer (EOC). Mol. Carcinog. 2007, 46, 872–885. [Google Scholar] [CrossRef]
- Lee, J.; Cuddihy, M.J.; Kotov, N.A. Three-Dimensional Cell Culture Matrices: State of the Art. Tissue Eng. Part B Rev. 2008, 14, 61–86. [Google Scholar] [CrossRef]
- Huttala, O.; Vuorenpaa, H.; Toimela, T.; Uotila, J.; Kuokkanen, H.; Ylikomi, T.; Sarkanen, J.R.; Heinonen, T. Human Vascular Model with Defined Stimulation Medium—A Characterization Study. ALTEX 2015, 32, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Vuorenpaa, H.; Penttinen, K.; Heinonen, T.; Pekkanen-Mattila, M.; Sarkanen, J.R.; Ylikomi, T.; Aalto-Setala, K. Maturation of Human Pluripotent Stem Cell Derived Cardiomyocytes is Improved in Cardiovascular Construct. Cytotechnology 2017, 69, 785–800. [Google Scholar] [CrossRef] [PubMed]
- Huttala, O.; Sarkanen, J.R.; Heinonen, T.; Ylikomi, T. Presence of Vasculature Results in Faster Insulin Response in Adipocytes in Vascularized Adipose Tissue Model. ALTEX 2019, 36, 419–434. [Google Scholar] [CrossRef] [PubMed]
- Toimela, T.; Huttala, O.; Sabell, E.; Mannerstrom, M.; Sarkanen, J.R.; Ylikomi, T.; Heinonen, T. Intra-Laboratory Validated Human Cell-Based in Vitro vasculogenesis/angiogenesis Test with Serum-Free Medium. Reprod. Toxicol. 2017, 70, 116–125. [Google Scholar] [CrossRef]
- Keyvani, S.; Karimi, N.; Orafa, Z.; Bouzari, S.; Oloomi, M. Assessment of Cytokeratin-19 Gene Expression in Peripheral Blood of Breast Cancer Patients and Breast Cancer Cell Lines. Biomark. Cancer 2016, 8, 57–63. [Google Scholar] [CrossRef]
- Spring, F.A.; Dalchau, R.; Daniels, G.L.; Mallinson, G.; Judson, P.A.; Parsons, S.F.; Fabre, J.W.; Anstee, D.J. The Ina and Inb Blood Group Antigens are Located on a Glycoprotein of 80,000 MW (the CDw44 Glycoprotein) Whose Expression is Influenced by the in(Lu) Gene. Immunology 1988, 64, 37–43. [Google Scholar]
- Aonuma, M.; Saeki, Y.; Akimoto, T.; Nakayama, Y.; Hattori, C.; Yoshitake, Y.; Nishikawa, K.; Shibuya, M.; Tanaka, N.G. Vascular Endothelial Growth Factor Overproduced by Tumour Cells Acts Predominantly as a Potent Angiogenic Factor Contributing to Malignant Progression. Int. J. Exp. Pathol. 1999, 80, 271–281. [Google Scholar] [CrossRef]
- Comsa, S.; Cimpean, A.M.; Raica, M. The Story of MCF-7 Breast Cancer Cell Line: 40 Years of Experience in Research. Anticancer Res. 2015, 35, 3147–3154. [Google Scholar]
- Bonaccorsi, L.; Carloni, V.; Muratori, M.; Salvadori, A.; Giannini, A.; Carini, M.; Serio, M.; Forti, G.; Baldi, E. Androgen Receptor Expression in Prostate Carcinoma Cells Suppresses alpha6beta4 Integrin-Mediated Invasive Phenotype. Endocrinology 2000, 141, 3172–3182. [Google Scholar] [CrossRef]
- Kaighn, M.E.; Narayan, K.S.; Ohnuki, Y.; Lechner, J.F.; Jones, L.W. Establishment and Characterization of a Human Prostatic Carcinoma Cell Line (PC-3). Investig. Urol. 1979, 17, 16–23. [Google Scholar]
- Harper, M.E.; Glynne-Jones, E.; Goddard, L.; Thurston, V.J.; Griffiths, K. Vascular Endothelial Growth Factor (VEGF) Expression in Prostatic Tumours and its Relationship to Neuroendocrine Cells. Br. J. Cancer 1996, 74, 910–916. [Google Scholar] [CrossRef]
- Connolly, J.M.; Rose, D.P. Angiogenesis in Two Human Prostate Cancer Cell Lines with Differing Metastatic Potential when Growing as Solid Tumors in Nude Mice. J. Urol. 1998, 160, 932–936. [Google Scholar] [CrossRef]
- Jain, R.; Fischer, S.; Serra, S.; Chetty, R. The use of Cytokeratin 19 (CK19) Immunohistochemistry in Lesions of the Pancreas, Gastrointestinal Tract, and Liver. Appl. Immunohistochem. Mol. Morphol. 2010, 18, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Velazquez, M.A.; Popov, V.M.; Lisanti, M.P.; Pestell, R.G. The Role of Breast Cancer Stem Cells in Metastasis and Therapeutic Implications. Am. J. Pathol. 2011, 179, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Hida, K.; Maishi, N.; Akiyama, K.; Ohmura-Kakutani, H.; Torii, C.; Ohga, N.; Osawa, T.; Kikuchi, H.; Morimoto, H.; Morimoto, M.; et al. Tumor Endothelial Cells with High Aldehyde Dehydrogenase Activity show Drug Resistance. Cancer Sci. 2017, 108, 2195–2203. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ma, H.; Zhang, J.; Zhu, L.; Wang, C.; Yang, Y. Unraveling the Roles of CD44/CD24 and ALDH1 as Cancer Stem Cell Markers in Tumorigenesis and Metastasis. Sci. Rep. 2017, 7, 13856. [Google Scholar] [CrossRef] [PubMed]
- Nazarenko, I.; Rana, S.; Baumann, A.; McAlear, J.; Hellwig, A.; Trendelenburg, M.; Lochnit, G.; Preissner, K.T.; Zoller, M. Cell Surface Tetraspanin Tspan8 Contributes to Molecular Pathways of Exosome-Induced Endothelial Cell Activation. Cancer Res. 2010, 70, 1668–1678. [Google Scholar] [CrossRef] [PubMed]
- Hromada, C.; Muhleder, S.; Grillari, J.; Redl, H.; Holnthoner, W. Endothelial Extracellular Vesicles-Promises and Challenges. Front. Physiol. 2017, 8, 275. [Google Scholar] [CrossRef]
- Ciardiello, C.; Cavallini, L.; Spinelli, C.; Yang, J.; Reis-Sobreiro, M.; de Candia, P.; Minciacchi, V.R.; Di Vizio, D. Focus on Extracellular Vesicles: New Frontiers of Cell-to-Cell Communication in Cancer. Int. J. Mol. Sci. 2016, 17, 175. [Google Scholar] [CrossRef]
- Minciacchi, V.R.; Freeman, M.R.; Di Vizio, D. Extracellular Vesicles in Cancer: Exosomes, Microvesicles and the Emerging Role of Large Oncosomes. Semin. Cell Dev. Biol. 2015, 40, 41–51. [Google Scholar] [CrossRef]
- Vogel, K.G.; Trotter, J.A. The Effect of Proteoglycans on the Morphology of Collagen Fibrils Formed in Vitro. Coll. Relat. Res. 1987, 7, 105–114. [Google Scholar] [CrossRef]
- Danielson, K.G.; Baribault, H.; Holmes, D.F.; Graham, H.; Kadler, K.E.; Iozzo, R.V. Targeted Disruption of Decorin Leads to Abnormal Collagen Fibril Morphology and Skin Fragility. J. Cell Biol. 1997, 136, 729–743. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Sun, H.; Owens, R.T.; Wu, J.; Chen, Y.Q.; Berquin, I.M.; Perry, D.; O’Flaherty, J.T.; Edwards, I.J. Decorin Suppresses Prostate Tumor Growth through Inhibition of Epidermal Growth Factor and Androgen Receptor Pathways. Neoplasia 2009, 11, 1042–1053. [Google Scholar] [CrossRef] [PubMed]
- Reed, C.C.; Waterhouse, A.; Kirby, S.; Kay, P.; Owens, R.T.; McQuillan, D.J.; Iozzo, R.V. Decorin Prevents Metastatic Spreading of Breast Cancer. Oncogene 2005, 24, 1104–1110. [Google Scholar] [CrossRef]
- Nash, M.A.; Deavers, M.T.; Freedman, R.S. The Expression of Decorin in Human Ovarian Tumors. Clin. Cancer Res. 2002, 8, 1754–1760. [Google Scholar]
- Koninger, J.; Giese, T.; di Mola, F.F.; Wente, M.N.; Esposito, I.; Bachem, M.G.; Giese, N.A.; Buchler, M.W.; Friess, H. Pancreatic Tumor Cells Influence the Composition of the Extracellular Matrix. Biochem. Biophys. Res. Commun. 2004, 322, 943–949. [Google Scholar] [CrossRef]
- Sofeu Feugaing, D.D.; Gotte, M.; Viola, M. More than Matrix: The Multifaceted Role of Decorin in Cancer. Eur. J. Cell Biol. 2013, 92, 1–11. [Google Scholar] [CrossRef]
- Bergqvist, A.; Bergqvist, D.; Ferno, M. Estrogen and Progesterone Receptors in Vessel Walls. Biochemical and Immunochemical Assays. Acta Obstet. Gynecol. Scand. 1993, 72, 10–16. [Google Scholar] [CrossRef]
- Yu, Y.; Lee, J.S.; Xie, N.; Li, E.; Hurtado-Coll, A.; Fazli, L.; Cox, M.; Plymate, S.; Gleave, M.; Dong, X. Prostate Stromal Cells Express the Progesterone Receptor to Control Cancer Cell Mobility. PLoS ONE 2014, 9, e92714. [Google Scholar] [CrossRef]
- DiMasi, J.A.; Grabowski, H.G. Economics of New Oncology Drug Development. J. Clin. Oncol. 2007, 25, 209–216. [Google Scholar] [CrossRef]
- Fornari, F.A.; Randolph, J.K.; Yalowich, J.C.; Ritke, M.K.; Gewirtz, D.A. Interference by Doxorubicin with DNA Unwinding in MCF-7 Breast Tumor Cells. Mol. Pharmacol. 1994, 45, 649–656. [Google Scholar] [PubMed]
- Longley, D.B.; Harkin, D.P.; Johnston, P.G. 5-Fluorouracil: Mechanisms of Action and Clinical Strategies. Nat. Rev. Cancer 2003, 3, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Stewart, R.J.; Panigrahy, D.; Flynn, E.; Folkman, J. Vascular Endothelial Growth Factor Expression and Tumor Angiogenesis are Regulated by Androgens in Hormone Responsive Human Prostate Carcinoma: Evidence for Androgen Dependent Destabilization of Vascular Endothelial Growth Factor Transcripts. J. Urol. 2001, 165, 688–693. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, B.; Theodorescu, D. Angiogenesis and Prostate Cancer Tumor Growth. J. Cell. Biochem. 2004, 91, 125–150. [Google Scholar] [CrossRef] [PubMed]
- Pulukuri, S.M.; Gondi, C.S.; Lakka, S.S.; Jutla, A.; Estes, N.; Gujrati, M.; Rao, J.S. RNA Interference-Directed Knockdown of Urokinase Plasminogen Activator and Urokinase Plasminogen Activator Receptor Inhibits Prostate Cancer Cell Invasion, Survival, and Tumorigenicity in Vivo. J. Biol. Chem. 2005, 280, 36529–36540. [Google Scholar] [CrossRef] [PubMed]
- Balbay, M.D.; Pettaway, C.A.; Kuniyasu, H.; Inoue, K.; Ramirez, E.; Li, E.; Fidler, I.J.; Dinney, C.P. Highly Metastatic Human Prostate Cancer Growing within the Prostate of Athymic Mice Overexpresses Vascular Endothelial Growth Factor. Clin. Cancer Res. 1999, 5, 783–789. [Google Scholar]
- Revert, F.; Revert-Ros, F.; Blasco, R.; Artigot, A.; Lopez-Pascual, E.; Gozalbo-Rovira, R.; Ventura, I.; Gutierrez-Carbonell, E.; Roda, N.; Ruiz-Sanchis, D.; et al. Selective Targeting of Collagen IV in the Cancer Cell Microenvironment Reduces Tumor Burden. Oncotarget 2018, 9, 11020–11045. [Google Scholar] [CrossRef]
- Sarkanen, J.R.; Vuorenpaa, H.; Huttala, O.; Mannerstrom, B.; Kuokkanen, H.; Miettinen, S.; Heinonen, T.; Ylikomi, T. Adipose Stromal Cell Tubule Network Model Provides a Versatile Tool for Vascular Research and Tissue Engineering. Cells Tissues Organs 2012, 196, 385–397. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Origin | Other Info | Reference | Angiogenesis Induction in Our Assay |
|---|---|---|---|---|
| MCF7 | Invasive breast ductal carcinoma: pleural effusion | Low tumorigenicity, poor angiogenic potential | [17,18] | - |
| LNCAP | human prostate adenocarcinoma: lymph node metastasis | Noninvasive | [19] | No induction |
| PC3 | bone metastasis of grade IV of prostate cancer | High tumorigenicity, Secreting VEGF | [20,21] | Induction |
| PC3M | Highly metastatic cell line derived from PC3 | Highly tumorigenic, angiogenic but not secreting VEGF165 | [22] | Induction |
| Sample Number | Form of Sample | Cancer Type/Origin | Drugs Received by the Patient | Angiogenesis Inducer |
|---|---|---|---|---|
| 1 | Ascites | Progressive, originating from colon, adenocarcinoma | Regorafenib | No |
| 2 | Ascites | CA12s increasing ascites increasing, ovarian cancer | Bevacizumab, Paclitaxel, Carboplatin, Letrozol | No |
| 3 | Pleural effusion | Metastatic thyroid cancer, progressive | Sorafenib | (Yes) |
| 4 | Solid tumor | Ovarian cancer | (No) | |
| 5 | Pleural effusion | Breast cancer, progressive | Eribulin | (Yes) |
| 6 | Ascites | Sigma adenocarcinoma, progressive | Capecitabine, Oxaliplatin | Yes |
| 7 | Solid tumor | Ovarian cancer | No | |
| 8 | Pleural effusion | Unknown | No |
| Angiogenesis stimulating medium (ASM) | DMEM/F12 2.56 mM L-glutamine 0.1 nM 3,3′,5-Triiodo-L-thyronine sodium salt ITSTM Premix: 6.65 µg/mL insulin 6.65 µg/mL Transferrin 6.65 ng/mL selenious acid 1% Bovine serum albumin (BSA) 2.8 mM Sodium puryvate 200 µg/mL Ascorbic acid 0.5 µg/mL Heparin 2 µg/mL Hydrocortisone/ cortisol 10 ng/mL Vascular endothelial growth factor (VEGF) 1 ng/mL Fibroblast growth factor (FGF-b) | Gibco Gibco Sigma BD Roche Gibco Sigma Sigma Sigma R&D systems (Minneapolis, MN, USA) R&D Systems |
| General cancer cell medium (GCM) | DMEM/F12 2.56 mM L-glutamine 5% human serum | Gibco Gibco Lonza |
| Liquid cancer sample medium (LCM) | DMEM/F12 2.56 mM L-glutamine 10% Supernatant from the isolation of the cells | Gibco Gibco |
| MCF7 medium | DMEM/F12 supplemented 10% fetal bovine serum 1% L-glutamine 10 ng/mL insulin | Gibco Gibco Gibco Sigma |
| PC3, LNCAP, and PC3M medium | RPMI1640 (containing 1mM L-glutamine) 1% penicillin-streptomycin (pen/strep) 10% fetal bovine serum | Gibco Gibco Gibco |
| Well Plate | Growth Area | Cancer cells Per Well | hASC Per Well (20,000 Cells/ cm2) | HUVEC Per Well (4100 Cells/ cm2) |
|---|---|---|---|---|
| 48 | 1.1 cm2 | 10,000 | 22,000 | 4500 |
| 96 | 0.33 cm2 | 5000 | 6600 | 1350 |
| 384 | 0.13 cm2 | 1000 | 2600 | 533 |
| Gene | Abbreviation | Primer Sequence |
|---|---|---|
| Decorin | DEC | GGACCGTTTCAACAGAGAGG Reverse GAGTTGTGTCAGG GGGAAGA |
| IGFBP5 | GCTCAGGCTTGAGGGTTTCT Reverse primer AGAGCGAGAGTGCAGGGATA | |
| Integrin B1 | ITGB1 | GCCGCGCGGAAAAGATGAA Reverse primer TGCTGTTCCTTTGCTACGGT |
| Integrin av | ITGAV | CGGAGGGAAGCAAAGGACC Reverse primer GTAGCAGGAGTCCCGAGAGA |
| Estrogen receptor 1 | ER1 | CGTCGCCTCTAACCTCGG Reverse primer AGCTCGTTCCCTTGGATCTG |
| Estrogen receptor 2 | ER2 | GGCTGCGAGAAATAACTGCC Reverse AATGCGGACACGTGCTTTTC |
| E-cadherin | CDH1 | CAGTTCAGACTCCAGCCCG Reverse CACCGTGAACGTGTAGCTCT |
| Progesterone receptor | PGR | ACCCGCCCTATCTCAACTACC Reverse AGGACACCATAATGACAGCCT), |
| CDK2 | GACACGCTGCTGGATGTCA Reverse CAGAAAGCTAGGCCCTGGAG | |
| RPLP0 | AATCTCCAGGGGCACCATT Reverse CGCTGGCTCCCACTTTGT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huttala, O.; Staff, S.; Heinonen, T.; Mäenpää, J.; Tanner, M.; Ylikomi, T. In Vitro Vascular Network Modified to Function as Culture Platform and Angiogenic Induction Potential Test for Cancer Cells. Int. J. Mol. Sci. 2020, 21, 1833. https://doi.org/10.3390/ijms21051833
Huttala O, Staff S, Heinonen T, Mäenpää J, Tanner M, Ylikomi T. In Vitro Vascular Network Modified to Function as Culture Platform and Angiogenic Induction Potential Test for Cancer Cells. International Journal of Molecular Sciences. 2020; 21(5):1833. https://doi.org/10.3390/ijms21051833
Chicago/Turabian StyleHuttala, Outi, Synnöve Staff, Tuula Heinonen, Johanna Mäenpää, Minna Tanner, and Timo Ylikomi. 2020. "In Vitro Vascular Network Modified to Function as Culture Platform and Angiogenic Induction Potential Test for Cancer Cells" International Journal of Molecular Sciences 21, no. 5: 1833. https://doi.org/10.3390/ijms21051833
APA StyleHuttala, O., Staff, S., Heinonen, T., Mäenpää, J., Tanner, M., & Ylikomi, T. (2020). In Vitro Vascular Network Modified to Function as Culture Platform and Angiogenic Induction Potential Test for Cancer Cells. International Journal of Molecular Sciences, 21(5), 1833. https://doi.org/10.3390/ijms21051833