DNA Methylation Profiling for Diagnosing Undifferentiated Sarcoma with Capicua Transcriptional Receptor (CIC) Alterations

 ,
,  , ,
, ,
Abstract
1. Introduction
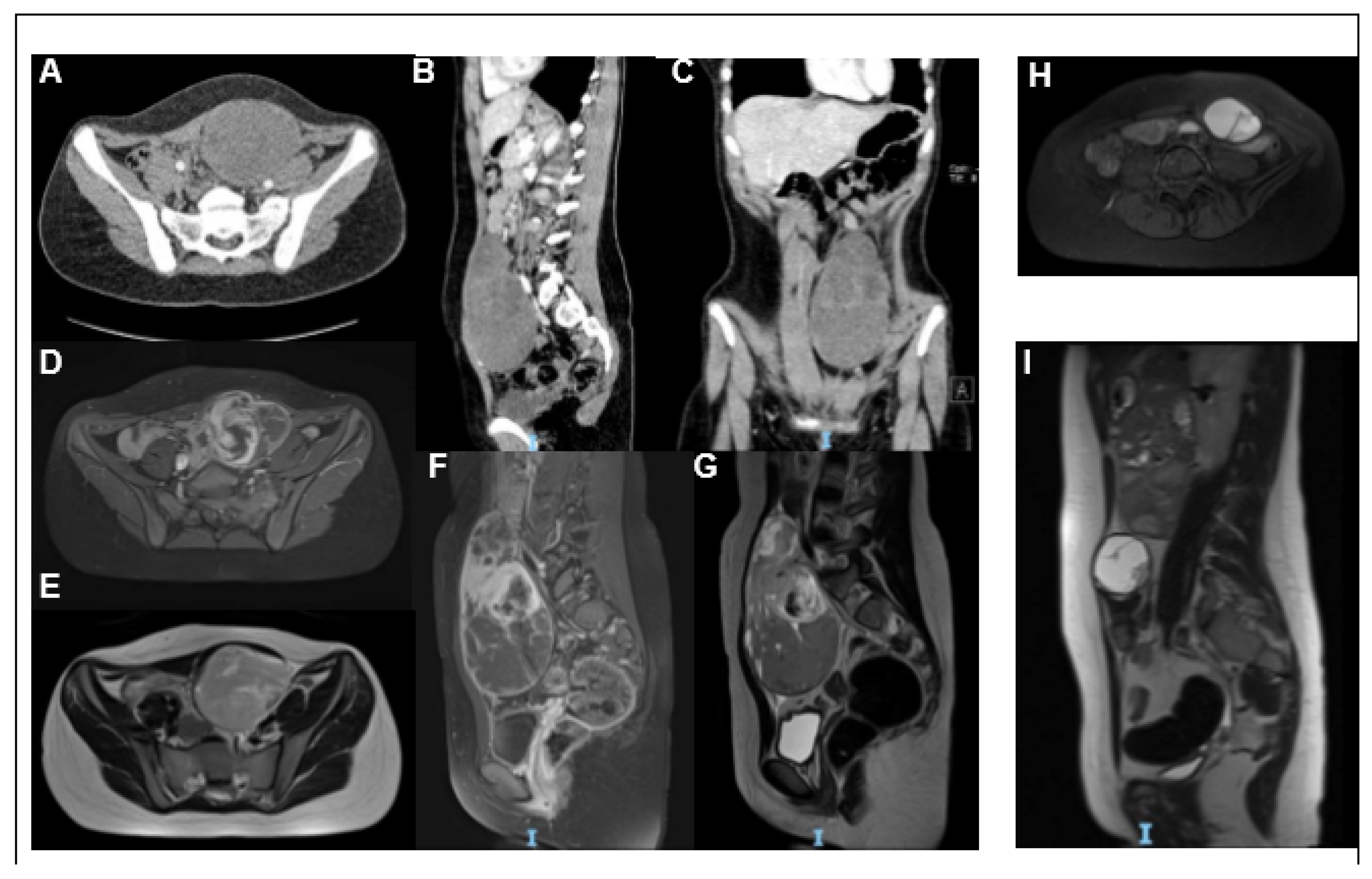
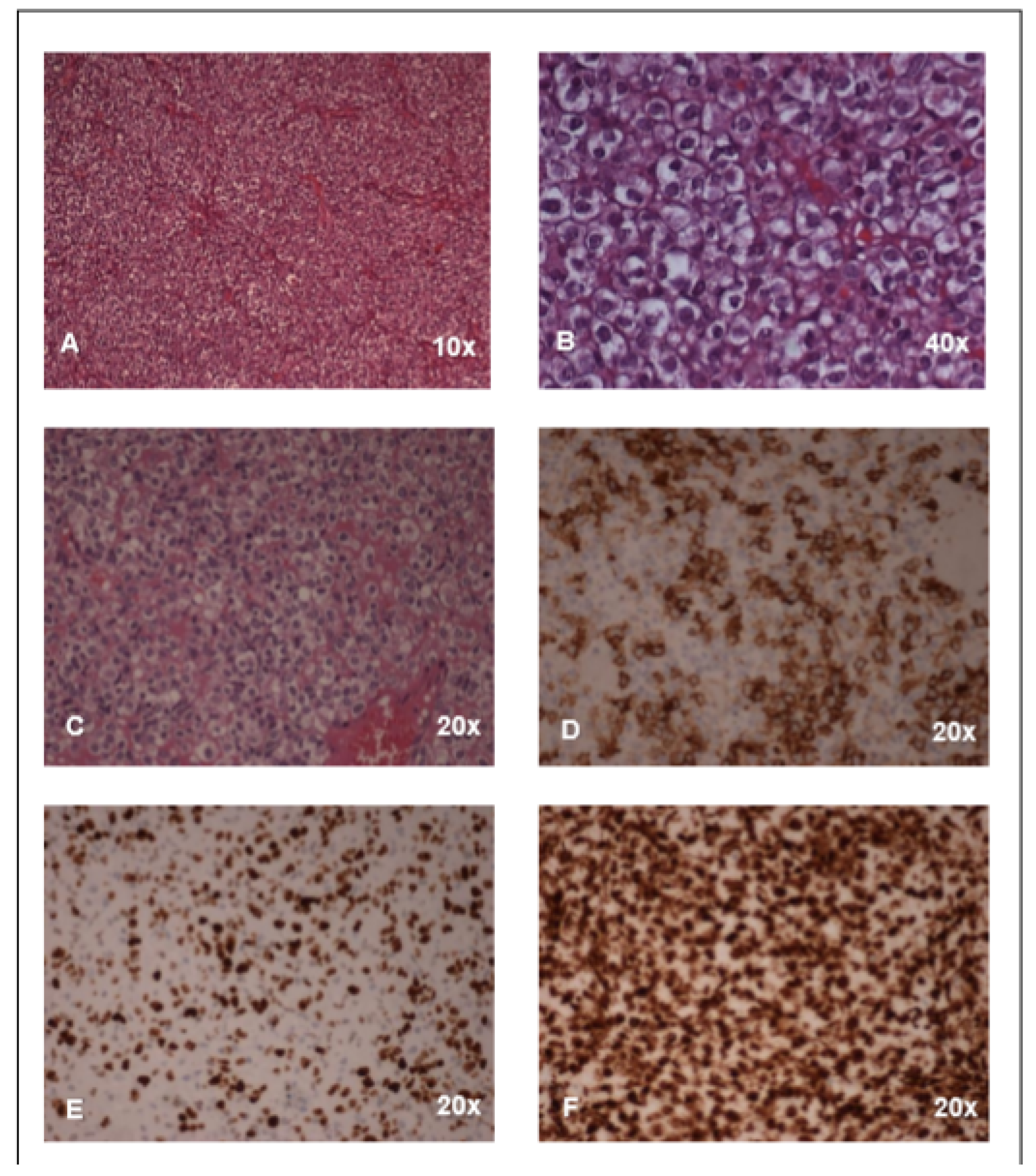
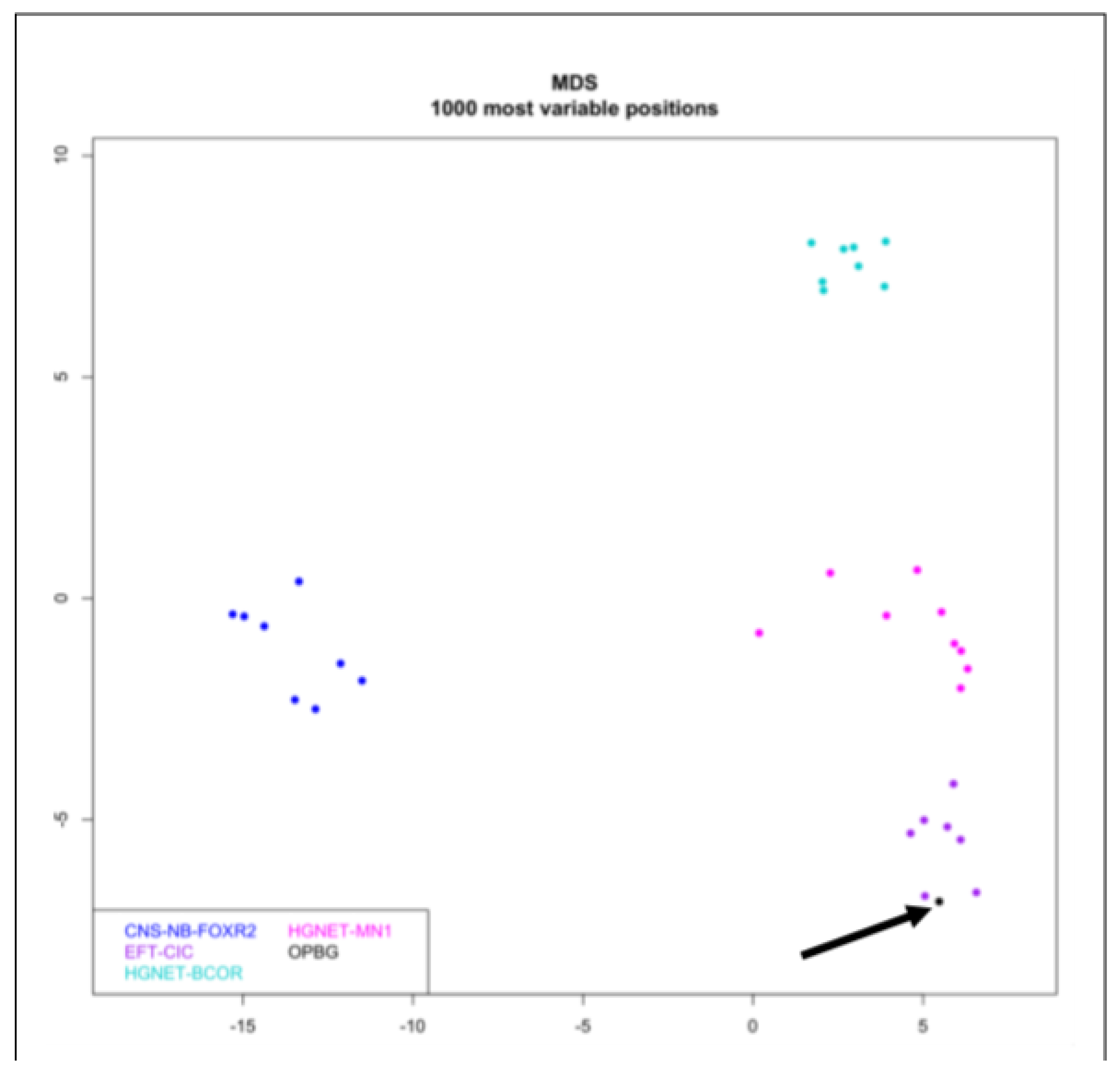
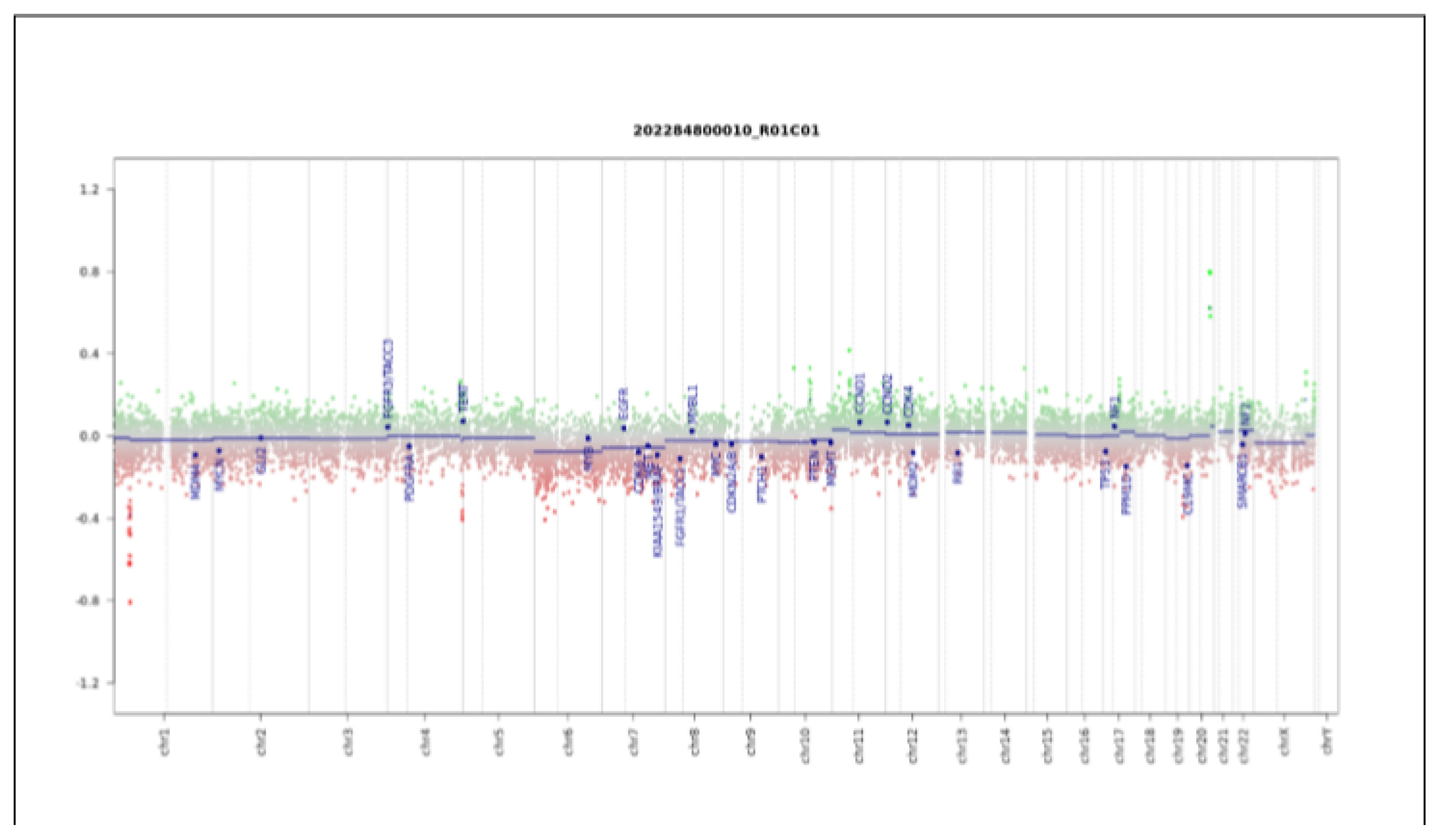
2. Case Report
3. Discussion
4. Conclusion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Ethics Statement
Informed Consent
Conflicts of Interest
References
- Renzi, S.; Anderson, N.D.; Light, N.; Gupta, A. Ewing-like sarcoma: An emerging family of round cell sarcomas. J. Cell Physiol. 2019, 234, 7999–8007. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.S.; Patel, R.M. Important Recently Characterized Non-Ewing Small Round Cell Tumors. Surg. Pathol. Clin. 2019, 12, 191–215. [Google Scholar] [CrossRef] [PubMed]
- Delattre, O.; Zucman, J.; Plougastel, B.; Desmaze, C.; Melot, T.; Peter, M.; Kovar, H.; Joubert, I.; de Jong, P.; Rouleau, G.; et al. Gene fusion with an ETS DNA-binding domain caused by chromosome translocation in human tumours. Nature 1992, 359, 162–165. [Google Scholar] [CrossRef]
- Sorensen, P.H.B.; Lessnick, S.L.; Lopez-Terrada, D.; Liu, X.F.; Triche, T.J.; Denny, C.T. A second Ewing’s sarcoma translocation, t(21;22), fuses the EWS gene to another ETS–family transcription factor, ERG. Nat. Genet. 1994, 6, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Szuhai, K.; IJszenga, M.; de Jong, D.; Karseladze, A.; Tanke, H.J.; Hogendoorn, P.C.W. The NFATc2 Gene Is Involved in a Novel Cloned Translocation in a Ewing Sarcoma Variant That Couples Its Function in Immunology to Oncology. Clin. Cancer Res. 2009, 15, 2259–2268. [Google Scholar] [CrossRef] [PubMed]
- Sumegi, J.; Nishio, J.; Nelson, M.; Frayer, R.W.; Perry, D.; Bridge, J.A. A novel t(4;22)(q31;q12) produces an EWSR1–SMARCA5 fusion in extraskeletal Ewing sarcoma/primitive neuroectodermal tumor. Mod. Pathol. 2011, 24, 333–342. [Google Scholar] [CrossRef]
- Llombart-Bosch, A.; Machado, I.; Navarro, S.; Bertoni, F.; Bacchini, P.; Alberghini, M.; Karzeladze, A.; Savelov, N.; Petrov, S.; Alvarado-Cabrero, I.; et al. Histological heterogeneity of Ewing’s sarcoma/PNET: An immunohistochemical analysis of 415 genetically confirmed cases with clinical support. Virchows Arch. 2009, 455, 397–411. [Google Scholar] [CrossRef]
- Mastrangelo, T.; Modena, P.; Tornielli, S.; Bullrich, F.; Testi, M.A.; Mezzelani, A.; Radice, P.; Azzarelli, A.; Pilotti, S.; Croce, C.M.; et al. A novel zinc finger gene is fused to EWS in small round cell tumor. Oncogene 2000, 19, 3799–3804. [Google Scholar] [CrossRef]
- Wang, L.; Bhargava, R.; Zheng, T.; Wexler, L.; Collins, M.H.; Roulston, D.; Ladanyi, M. Undifferentiated Small Round Cell Sarcomas with Rare EWS Gene Fusions. J. Mol. Diagn. 2007, 9, 498–509. [Google Scholar] [CrossRef]
- Watson, S.; Perrin, V.; Guillemot, D.; Reynaud, S.; Coindre, J.-M.; Karanian, M.; Guinebretière, J.-M.; Freneaux, P.; Le Loarer, F.; Bouvet, M.; et al. Transcriptomic definition of molecular subgroups of small round cell sarcomas: Molecular classification of sarcoma subtypes. J. Pathol. 2018, 245, 29–40. [Google Scholar] [CrossRef]
- Jo, V.Y.; Fletcher, C.D.M. WHO classification of soft tissue tumours: An update based on the 2013 (4th) edition. Pathology 2014, 46, 95–104. [Google Scholar] [CrossRef]
- Koelsche, C.; Hartmann, W.; Schrimpf, D.; Stichel, D.; Jabar, S.; Ranft, A.; Reuss, D.E.; Sahm, F.; Jones, D.T.W.; Bewerunge-Hudler, M.; et al. Array-based DNA-methylation profiling in sarcomas with small blue round cell histology provides valuable diagnostic information. Mod. Pathol. 2018, 31, 1246–1256. [Google Scholar] [CrossRef] [PubMed]
- Alaggio, R.; Coffin, C.M. The Evolution of Pediatric Soft Tissue Sarcoma Classification in the Last 50 Years. Pediatr. Dev. Pathol. 2015, 18, 481–494. [Google Scholar] [CrossRef] [PubMed]
- Kawamura-Saito, M.; Yamazaki, Y.; Kaneko, K.; Kawaguchi, N.; Kanda, H.; Mukai, H.; Gotoh, T.; Motoi, T.; Fukayama, M.; Aburatani, H.; et al. Fusion between CIC and DUX4 up-regulates PEA3 family genes in Ewing-like sarcomas with t(4;19)(q35;q13) translocation. Human Mol. Genet. 2006, 15, 2125–2137. [Google Scholar] [CrossRef] [PubMed]
- Pierron, G.; Tirode, F.; Lucchesi, C.; Reynaud, S.; Ballet, S.; Cohen-Gogo, S.; Perrin, V.; Coindre, J.-M.; Delattre, O. A new subtype of bone sarcoma defined by BCOR-CCNB3 gene fusion. Nat. Genet. 2012, 44, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Italiano, A.; Sung, Y.S.; Zhang, L.; Singer, S.; Maki, R.G.; Coindre, J.-M.; Antonescu, C.R. High prevalence of CIC fusion with double-homeobox (DUX4) transcription factors in EWSR1-negative undifferentiated small blue round cell sarcomas. Genes Chromosom. Cancer 2012, 51, 207–218. [Google Scholar] [CrossRef]
- Specht, K.; Zhang, L.; Sung, Y.-S.; Nucci, M.; Dry, S.; Vaiyapuri, S.; Richter, G.H.S.; Fletcher, C.D.M.; Antonescu, C.R. Novel BCOR-MAML3 and ZC3H7B-BCOR Gene Fusions in Undifferentiated Small Blue Round Cell Sarcomas. Am. J. Surg. Pathol. 2016, 40, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Petruzzellis, G.; Alessi, I.; Colafati, G.S.; Diomedi-Camassei, F.; Ciolfi, A.; Pedace, L.; Cacchione, A.; Carai, A.; Tartaglia, M.; Mastronuzzi, A.; et al. Role of DNA Methylation Profile in Diagnosing Astroblastoma: A Case Report and Literature Review. Front. Genet. 2019, 10, 391. [Google Scholar] [CrossRef]
- Flex, E.; Martinelli, S.; Van Dijck, A.; Ciolfi, A.; Cecchetti, S.; Coluzzi, E.; Pannone, L.; Andreoli, C.; Radio, F.C.; Pizzi, S.; et al. Aberrant Function of the C-Terminal Tail of HIST1H1E Accelerates Cellular Senescence and Causes Premature Aging. Am. J. Hum. Genet. 2019, 105, 493–508. [Google Scholar] [CrossRef]
- Ballabio, C.; Anderle, M.; Gianesello, M.; Lago, C.; Miele, E.; Cardano, M.; Aiello, G.; Piazza, S.; Caron, D.; Gianno, F.; et al. Modeling Medulloblastoma in-vivo and with human cerebellar organoids. Nature Commun. 2019. [Google Scholar] [CrossRef]
- Capper, D.; Jones, D.T.W.; Sill, M.; Hovestadt, V.; Schrimpf, D.; Sturm, D.; Koelsche, C.; Sahm, F.; Chavez, L.; Reuss, D.E.; et al. DNA methylation-based classification of central nervous system tumours. Nature 2018, 555, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Le Loarer, F.; Pissaloux, D.; Coindre, J.M.; Tirode, F.; Vince, D.R. Update on Families of Round Cell Sarcomas Other than Classical Ewing Sarcomas. Surg. Pathol. Clin. 2017, 10, 587–620. [Google Scholar] [CrossRef] [PubMed]
- Machado, I.; Navarro, S.; Llombart-Bosch, A. Ewing sarcoma and the new emerging Ewing-like sarcomas: (CIC and BCOR-rearranged-sarcomas). A systematic review. Histol. Histopathol. 2016, 31, 1169–1181. [Google Scholar] [PubMed]
- Graham, C.; Chilton-MacNeill, S.; Zielenska, M.; Somers, G.R. The CIC-DUX4 fusion transcript is present in a subgroup of pediatric primitive round cell sarcomas. Hum. Pathol. 2012, 43, 180–189. [Google Scholar] [CrossRef]
- Yoshida, A.; Goto, K.; Kodaira, M.; Kobayashi, E.; Kawamoto, H.; Mori, T.; Yoshimoto, S.; Endo, O.; Kodama, N.; Kushima, R.; et al. CIC-rearranged Sarcomas: A Study of 20 Cases and Comparisons With Ewing Sarcomas. Am. J. Surg. Pathol. 2016, 40, 313–323. [Google Scholar] [CrossRef]
- Kilpatrick, S.E.; Reith, J.D.; Rubin, B. Ewing Sarcoma and the History of Similar and Possibly Related Small Round Cell Tumors: From Whence Have We Come and Where are We Going? Adv. Anat. Pathol. 2018, 25, 314–326. [Google Scholar] [CrossRef]
- Richkind, K.E.; Romansky, S.G.; Finklestein, J.Z. t(4;19)(q35;q13.1): A recurrent change in primitive mesenchymal tumors? Cancer Genet. Cytogenet. 1996, 87, 71–74. [Google Scholar] [CrossRef]
- Van Geel, M.; van Deutekom, J.C.T.; van Staalduinen, A.; Lemmers, R.J.L.F.; Dickson, M.C.; Hofker, M.H.; Padberg, G.W.; Hewitt, J.E.; de Jong, P.J.; Frants, R.R. Identification of a novel β-tubulin subfamily with one member (TUBB4Q) located near the telomere of chromosome region 4q35. Cytogenet. Genome Res. 2000, 88, 316–321. [Google Scholar] [CrossRef]
- Van der Maarel, S.M.; Tawil, R.; Tapscott, S.J. Facioscapulohumeral muscular dystrophy and DUX4: Breaking the silence. Trends Mol. Med. 2011, 17, 252–258. [Google Scholar] [CrossRef]
- Krom, Y.D.; Thijssen, P.E.; Young, J.M.; den Hamer, B.; Balog, J.; Yao, Z.; Maves, L.; Snider, L.; Knopp, P.; Zammit, P.S.; et al. Intrinsic Epigenetic Regulation of the D4Z4 Macrosatellite Repeat in a Transgenic Mouse Model for FSHD. PLoS Genet. 2013, 9, e1003415. [Google Scholar] [CrossRef]
- Antonescu, C.R.; Owosho, A.A.; Zhang, L.; Chen, S.; Deniz, K.; Huryn, J.M.; Kao, Y.-C.; Huang, S.-C.; Singer, S.; Tap, W.; et al. Sarcomas with CIC-rearrangements Are a Distinct Pathologic Entity with Aggressive Outcome: A Clinicopathologic and Molecular Study of 115 Cases. Am. J. Surg. Pathol. 2017, 41, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.-Y.K.; Thomas, D.G.; McHugh, J.B.; Patel, R.M.; Roulston, D.; Schuetze, S.M.; Chugh, R.; Biermann, J.S.; Lucas, D.R. Undifferentiated Small Round Cell Sarcoma with t(4;19)(q35;q13.1) CIC-DUX4 Fusion: A Novel Highly Aggressive Soft Tissue Tumor with Distinctive Histopathology. Am. J. Surg. Pathol. 2013, 37, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, T.; Tanaka, M.; Homme, M.; Yamazaki, Y.; Takazawa, Y.; Antonescu, C.R.; Nakamura, T. CIC-DUX4 Induces Small Round Cell Sarcomas Distinct from Ewing Sarcoma. Cancer Res. 2017, 77, 2927–2937. [Google Scholar] [CrossRef] [PubMed]
- Jeon, I.S.; Davis, J.N.; Braun, B.S.; Sublett, J.E.; Roussel, M.F.; Denny, C.T.; Shapiro, D.N. A variant Ewing’s sarcoma translocation (7;22) fuses the EWS gene to the ETS gene ETV1. Oncogene 1995, 10, 1229–1234. [Google Scholar]
- Kaneko, Y.; Yoshida, K.; Handa, M.; Toyoda, Y.; Nishihira, H.; Tanaka, Y.; Sasaki, Y.; Ishida, S.; Higashino, F.; Fujinaga, K. Fusion of an ETS-family gene, EIAF, to EWS by t(17;22)(q12;q12) chromosome translocation in an undifferentiated sarcoma of infancy. Genes Chromosomes Cancer 1996, 15, 115–121. [Google Scholar] [CrossRef]
- Solomon, D.A.; Brohl, A.S.; Khan, J.; Miettinen, M. Clinicopathologic Features of a Second Patient with Ewing-like Sarcoma Harboring CIC-FOXO4 Gene Fusion. Am. J. Surg. Pathol. 2014, 38, 1724–1725. [Google Scholar] [CrossRef]
- Sugita, S.; Arai, Y.; Tonooka, A.; Hama, N.; Totoki, Y.; Fujii, T.; Aoyama, T.; Asanuma, H.; Tsukahara, T.; Kaya, M.; et al. A Novel CIC-FOXO4 Gene Fusion in Undifferentiated Small Round Cell Sarcoma: A Genetically Distinct Variant of Ewing-like Sarcoma. Am. J. Surg. Pathol. 2014, 38, 1571–1576. [Google Scholar] [CrossRef]
- Sturm, D.; Orr, B.A.; Toprak, U.H.; Hovestadt, V.; Jones, D.T.W.; Capper, D.; Sill, M.; Buchhalter, I.; Northcott, P.A.; Leis, I.; et al. New Brain Tumor Entities Emerge from Molecular Classification of CNS-PNETs. Cell 2016, 164, 1060–1072. [Google Scholar] [CrossRef]
- Smith, S.C.; Palanisamy, N.; Martin, E.; Almenara, J.; McHugh, J.B.; Choi, E.-Y.K.; Lucas, D.R.; Betz, B.L.; Thomas, D.; Patel, R.M. The utility of ETV1, ETV4 and ETV5 RNA in-situ hybridization in the diagnosis of CIC-DUX sarcomas. Histopathology 2017, 70, 657–663. [Google Scholar] [CrossRef]
- Gambarotti, M.; Benini, S.; Gamberi, G.; Cocchi, S.; Palmerini, E.; Sbaraglia, M.; Donati, D.; Picci, P.; Vanel, D.; Ferrari, S.; et al. CIC-DUX4 fusion-positive round-cell sarcomas of soft tissue and bone: A single-institution morphological and molecular analysis of seven cases. Histopathology 2016, 69, 624–634. [Google Scholar] [CrossRef]
- Haidar, A.; Arekapudi, S.; DeMattia, F.; Abu-Isa, E.; Kraut, M. High-grade undifferentiated small round cell sarcoma with t(4;19)(q35;q13.1) CIC-DUX4 fusion: Emerging entities of soft tissue tumors with unique histopathologic features--a case report and literature review. Am. J. Case Rep. 2015, 16, 87–94. [Google Scholar] [PubMed]
- Pappo, A.S.; Dirksen, U. Rhabdomyosarcoma, Ewing Sarcoma, and Other Round Cell Sarcomas. JCO 2018, 36, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Specht, K.; Sung, Y.-S.; Zhang, L.; Richter, G.H.S.; Fletcher, C.D.; Antonescu, C.R. Distinct transcriptional signature and immunoprofile of CIC-DUX4 fusion-positive round cell tumors compared to EWSR1 -rearranged ewing sarcomas: Further evidence toward distinct pathologic entities: Gene Expression in CIC-DUX4 Sarcomas. Genes Chromosomes Cancer 2014, 53, 622–633. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Kuda, M.; Kohashi, K.; Yamamoto, H.; Takemoto, J.; Ishii, T.; Iura, K.; Maekawa, A.; Bekki, H.; Ito, T.; et al. Histological and immunohistochemical characteristics of undifferentiated small round cell sarcomas associated with CIC-DUX4 and BCOR-CCNB3 fusion genes. Virchows Arch. 2017, 470, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.C.; Buehler, D.; Choi, E.-Y.K.; McHugh, J.B.; Rubin, B.P.; Billings, S.D.; Balzer, B.; Thomas, D.G.; Lucas, D.R.; Goldblum, J.R.; et al. CIC-DUX sarcomas demonstrate frequent MYC amplification and ETS-family transcription factor expression. Mod. Pathol. 2015, 28, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.P.; Fletcher, C.D.; Hornick, J.L. Evaluation of ETV4 and WT1 expression in CIC-rearranged sarcomas and histologic mimics. Mod. Pathol. 2016, 29, 1324–1334. [Google Scholar] [CrossRef] [PubMed]
- Le Guellec, S.; Velasco, V.; Pérot, G.; Watson, S.; Tirode, F.; Coindre, J.-M. ETV4 is a useful marker for the diagnosis of CIC-rearranged undifferentiated round-cell sarcomas: A study of 127 cases including mimicking lesions. Mod. Pathol. 2016, 29, 1523–1531. [Google Scholar] [CrossRef]
- Hung, Y.P.; Fletcher, C.D.M.; Hornick, J.L. Evaluation of NKX2-2 expression in round cell sarcomas and other tumors with EWSR1 rearrangement: Imperfect specificity for Ewing sarcoma. Mod. Pathol. 2016, 29, 370–380. [Google Scholar] [CrossRef]
- Siegele, B.; Roberts, J.; Black, J.O.; Rudzinski, E.; Vargas, S.O.; Galambos, C. DUX4 Immunohistochemistry Is a Highly Sensitive and Specific Marker for CIC-DUX4 Fusion-positive Round Cell Tumor. Am. J. Surg. Pathol. 2017, 41, 423–429. [Google Scholar] [CrossRef]
- Kao, Y.-C.; Sung, Y.-S.; Chen, C.-L.; Zhang, L.; Dickson, B.C.; Swanson, D.; Vaiyapuri, S.; Latif, F.; Alholle, A.; Huang, S.-C.; et al. ETV transcriptional upregulation is more reliable than RNA sequencing algorithms and FISH in diagnosing round cell sarcomas with CIC gene rearrangements: Kao et al. Genes Chromosomes Cancer 2017, 56, 501–510. [Google Scholar] [CrossRef]
- Kunju, L.P.; Carskadon, S.; Siddiqui, J.; Tomlins, S.A.; Chinnaiyan, A.M.; Palanisamy, N. Novel RNA Hybridization Method for the In Situ Detection of ETV1, ETV4, and ETV5 Gene Fusions in Prostate Cancer. Appl. Immunohistochem. Mol. Morphol. 2014, 22, e32–e40. [Google Scholar] [CrossRef] [PubMed]
- Charville, G.W.; Wang, W.-L.; Ingram, D.R.; Roy, A.; Thomas, D.; Patel, R.M.; Hornick, J.L.; van de Rijn, M.; Lazar, A.J. EWSR1 fusion proteins mediate PAX7 expression in Ewing sarcoma. Mod. Pathol. 2017, 30, 1312–1320. [Google Scholar] [CrossRef] [PubMed]
- Ciolfi, A.; Aref-Eshghi, E.; Pizzi, S.; Pedace, L.; Miele, E.; Kerkhof, J.; Flex, E.; Martinelli, S.; Radio, F.C.; Ruivenkamp, C.A.L.; et al. Frameshift mutations at the C-terminus of HIST1H1E result in a specific DNA hypomethylation signature. Clin. Epigenet. 2020, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Renner, M.; Wolf, T.; Meyer, H.; Hartmann, W.; Penzel, R.; Ulrich, A.; Lehner, B.; Hovestadt, V.; Czwan, E.; Egerer, G.; et al. Integrative DNA methylation and gene expression analysis in high-grade soft tissue sarcomas. Genome. Biol. 2013, 14, r137. [Google Scholar] [CrossRef] [PubMed]
- Heyn, H.; Esteller, M. DNA methylation profiling in the clinic: Applications and challenges. Nat. Rev. Genet. 2012, 13, 679–692. [Google Scholar] [CrossRef]
- Kumar, R.; Liu, A.P.Y.; Orr, B.A.; Northcott, P.A.; Robinson, G.W. Advances in the classification of pediatric brain tumors through DNA methylation profiling: From research tool to frontline diagnostic: Classification of Pediatric Brain Tumors. Cancer 2018, 124, 4168–4180. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CIC-Rearranged Sarcoma | BCOR-Rearranged Sarcoma | Ewing Sarcoma | Poorly Differentiated Synovial Sarcoma | Desmoplastic Small Round Cell Tumor | Alveolar Rhabdomyosarcoma | Mesenchymal Chondrosarcoma | High-Grade Myxoid Liposarcoma | |
|---|---|---|---|---|---|---|---|---|
| Peak incidence | 3rd–4th decade | 2nd decade | 2nd decade | 4th decade | 3rd decade | 2nd decade | 3rd decade | 4th decade |
| Most common anatomic site | Soft tissues of trunk/pelvis and extremities | Pelvic bones and metadiaphysis of long bones of lower extremities | Metadiaphysis of long bones, ribs, pelvis | Soft tissues of extremities | Soft tissues and viscera of abdomen | Soft tissues of extremities | Facial bones, long bones of lower extremities, and pelvic bones | Soft tissues of lower limbs, especially thigh |
| Cytomorphology | Round cells with moderate eosinophilic cytoplasm | Monotonous round cells | At least focal spindling | Monotonous round cells, minimal cytoplasm | Monotonous round cells, rare multinucleated giant cells or striated cells | Round cells and spindled cells | Round cells with rare univacuolated lipoblasts | |
| Nuclear features | Vesicular chromatin, prominent nucleoli, moderate pleomorphism | Fine chromatin, inconspicuous nucleoli | Fine chromatin, inconspicuous nucleoli | Vesicular chromatin, prominent nucleoli | Uniform hyperchromasia, inconspicuous nucleoli, and minimal eosinophilic cytoplasm | Large, fine chromatin, prominent nucleoli | Small, fine chromatin, inconspicuous nucleoli | Vesicular chromatin, prominent nucleoli, moderate pleomorphism |
| Stroma | Myxoid changes common, subset with dense hyaline bands | Subset with myxoid changes | Scant | Scant | Abundant desmoplastic stroma encasing round cell nests | Fibrotic septae between anastomosing round cells | Geographically variable chondroid matrix | Myxoid changes |
| Typical positive immunohistochemical profile | Patchy CD99, ETV4, WT1, can be ERG and FLI1 | Patchy CD99, TLE1, BCOR, CCNB3 | Diffuse membranous CD99, NKX2-2 | TLE1, patchy EMA and keratin | WT1, keratin, EMA, desmin (perinuclear) | Desmin, myogenin, MyoD1 | Diffuse membranous CD99, NKX2-2, S100 in chondroid areas | S100 |
| Genetics (in order of incidence) | CIC-DUX4, CIC-DUX4L, CIC-FOX04 | BCOR-CCNB3, BCOR-MAML3, ZC3H7B-BCOR | EWSR1-FLI1, EWSR1-ERG, EWSR1-PATZ1, also FUS-ERG | SS18-SSX1, SS18-SSX2 | EWSR1-WT1 | PAX3-FOX01, PAX7-FOXO1 | HEY1-NCOA2 | FUS-DDIT3 EWSR1-DDIT3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miele, E.; De Vito, R.; Ciolfi, A.; Pedace, L.; Russo, I.; De Pasquale, M.D.; Di Giannatale, A.; Crocoli, A.; Angelis, B.D.; Tartaglia, M.; et al. DNA Methylation Profiling for Diagnosing Undifferentiated Sarcoma with Capicua Transcriptional Receptor (CIC) Alterations. Int. J. Mol. Sci. 2020, 21, 1818. https://doi.org/10.3390/ijms21051818
Miele E, De Vito R, Ciolfi A, Pedace L, Russo I, De Pasquale MD, Di Giannatale A, Crocoli A, Angelis BD, Tartaglia M, et al. DNA Methylation Profiling for Diagnosing Undifferentiated Sarcoma with Capicua Transcriptional Receptor (CIC) Alterations. International Journal of Molecular Sciences. 2020; 21(5):1818. https://doi.org/10.3390/ijms21051818
Chicago/Turabian StyleMiele, Evelina, Rita De Vito, Andrea Ciolfi, Lucia Pedace, Ida Russo, Maria Debora De Pasquale, Angela Di Giannatale, Alessandro Crocoli, Biagio De Angelis, Marco Tartaglia, and et al. 2020. "DNA Methylation Profiling for Diagnosing Undifferentiated Sarcoma with Capicua Transcriptional Receptor (CIC) Alterations" International Journal of Molecular Sciences 21, no. 5: 1818. https://doi.org/10.3390/ijms21051818
APA StyleMiele, E., De Vito, R., Ciolfi, A., Pedace, L., Russo, I., De Pasquale, M. D., Di Giannatale, A., Crocoli, A., Angelis, B. D., Tartaglia, M., Alaggio, R., & Milano, G. M. (2020). DNA Methylation Profiling for Diagnosing Undifferentiated Sarcoma with Capicua Transcriptional Receptor (CIC) Alterations. International Journal of Molecular Sciences, 21(5), 1818. https://doi.org/10.3390/ijms21051818