Differential Expression of DUB Genes in Ovarian Cells Treated with Di-2-Ethylhexyl Phthalate



Abstract
1. Introduction
2. Results
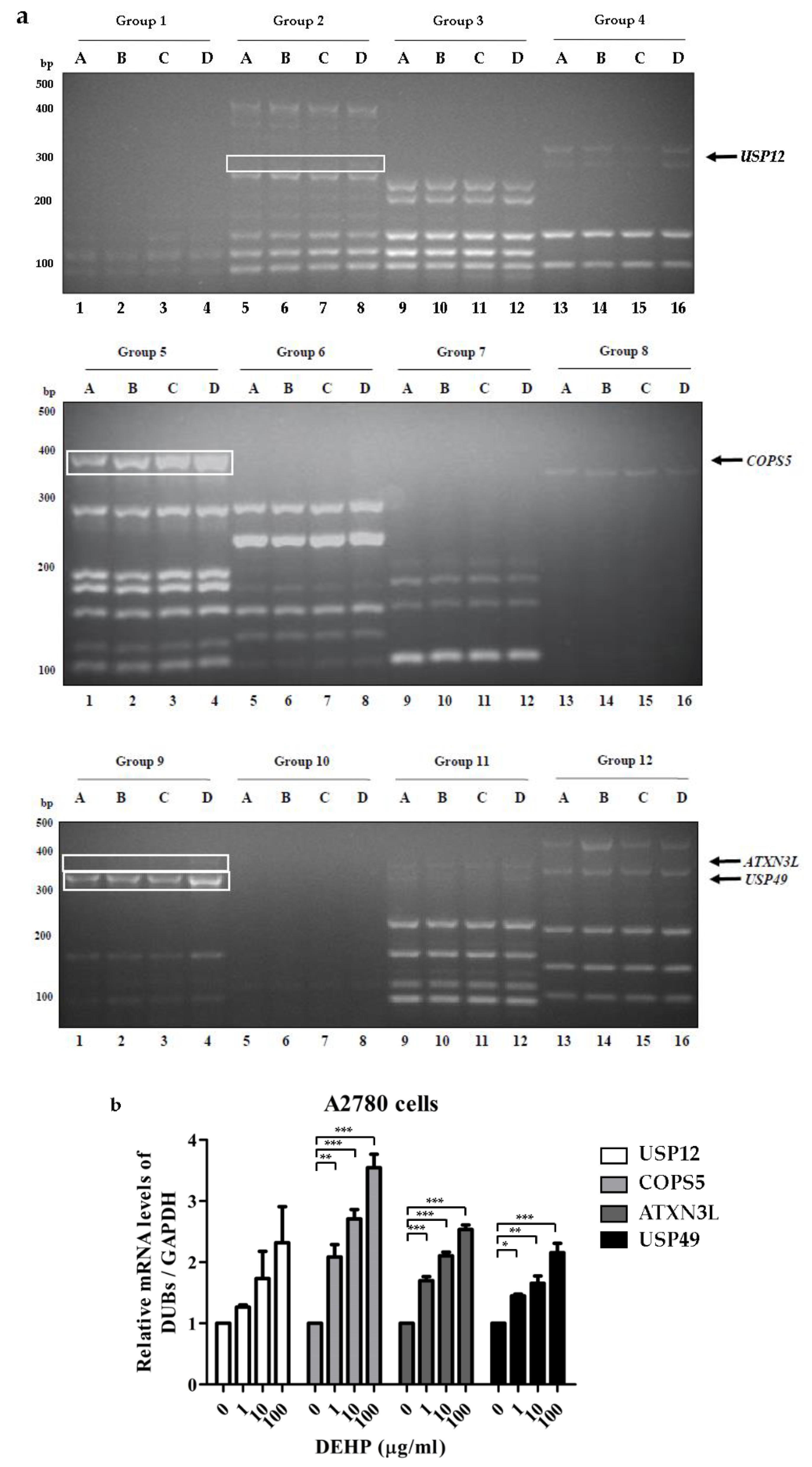
2.1. Differential Expressions of DUBs by DEHP in Ovarian Cells
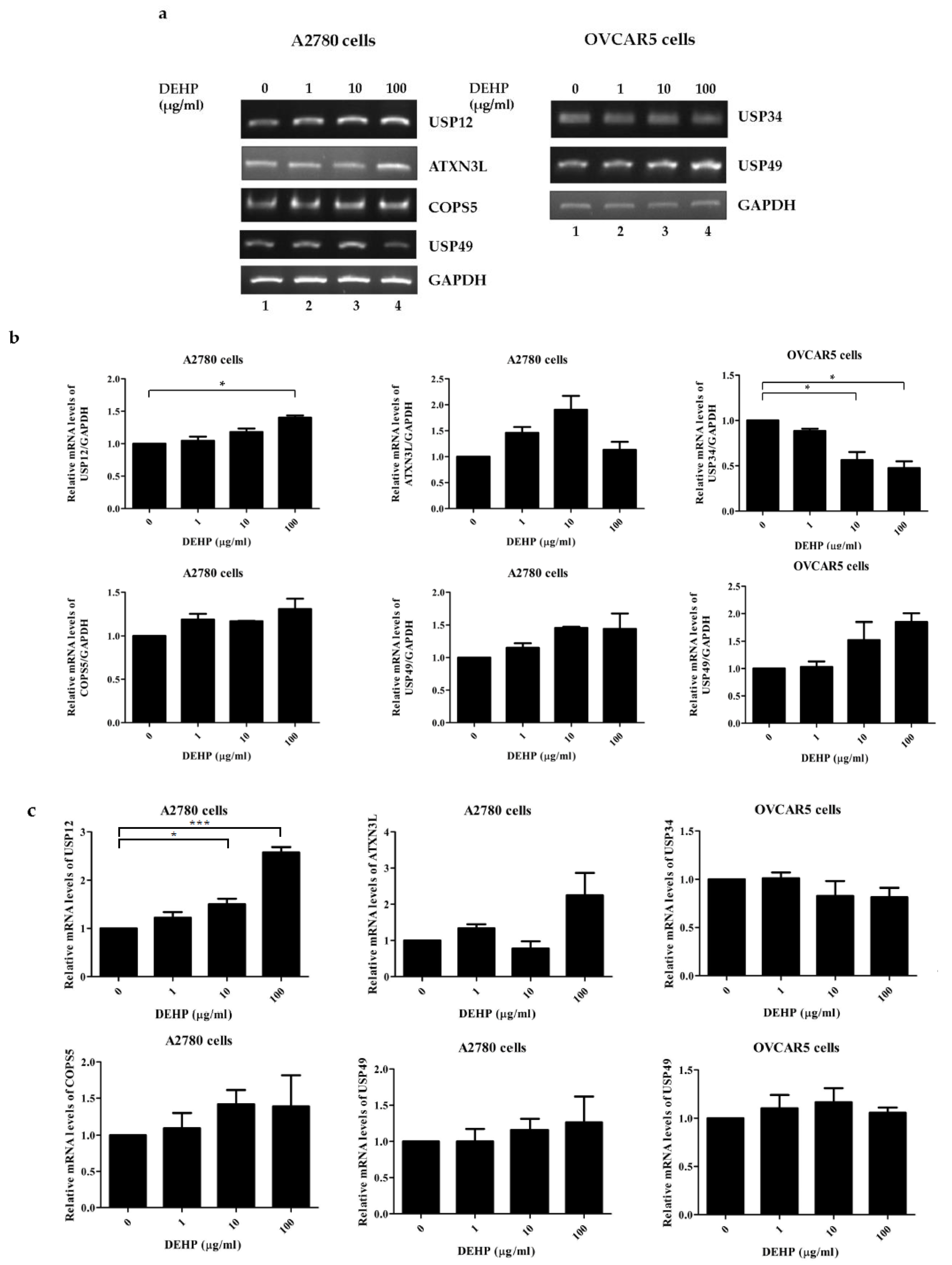
2.2. DEHP Exposure Influenced the mRNA Levels of Several DUB Genes in Ovarian Cells
2.3. Up-Regulation of USP49 Protein by DEHP in Ovarian Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. RNA Extraction and cDNA Synthesis
4.3. Multiplex RT-PCR and qRT-PCR
4.4. Western Blot Analysis
4.5. Antibodies
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kwon, S.K.; Saindane, M.; Baek, K.H. p53 stability is regulated by diverse deubiquitinating enzymes. Biochim. Biophys Acta Rev. Cancer 2017, 1868, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Pickart, C.M. Mechanisms underlying ubiquitination. Annu. Rev. Biochem. 2001, 70, 503–533. [Google Scholar] [CrossRef] [PubMed]
- Woelk, T.; Sigismund, S.; Penengo, L.; Polo, S. The ubiquitination code: A signalling problem. Cell Div. 2007, 2, 11. [Google Scholar] [CrossRef]
- Todi, S.V.; Paulson, H.L. Balancing act: Deubiquitinating enzymes in the nervous system. Trends Neurosci. 2011, 34, 370–382. [Google Scholar] [CrossRef] [PubMed]
- Abdul Rehman, S.A.; Kristariyanto, Y.A.; Choi, S.Y.; Nkosi, P.J.; Weidlich, S.; Labib, K.; Hofmann, K.; Kulathu, Y. MINDY-1 Is a Member of an Evolutionarily Conserved and Structurally Distinct New Family of Deubiquitinating Enzymes. Mol. Cell 2016, 63, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Beck-Peccoz, P.; Persani, L. Premature ovarian failure. Orphanet J. Rare. Dis. 2006, 1, 9. [Google Scholar] [CrossRef]
- Jankowska, K. Premature ovarian failure. Prz Menopauzalny 2017, 16, 51–56. [Google Scholar] [CrossRef]
- Ayesha, V.J.; Goswami, D. Premature Ovarian Failure: An Association with Autoimmune Diseases. J. Clin. Diagn. Res. 2016, 10, QC10–QC12. [Google Scholar] [CrossRef]
- Vabre, P.; Gatimel, N.; Moreau, J.; Gayrard, V.; Picard-Hagen, N.; Parinaud, J.; Leandri, R.D. Environmental pollutants, a possible etiology for premature ovarian insufficiency: A narrative review of animal and human data. Environ. Health 2017, 16, 37. [Google Scholar] [CrossRef]
- Tran, D.N.; Jung, E.M.; Yoo, Y.M.; Ahn, C.; Kang, H.Y.; Choi, K.C.; Hyun, S.H.; Dang, V.H.; Pham, T.N.; Jeung, E.B. Depletion of follicles accelerated by combined exposure to phthalates and 4-vinylcyclohexene diepoxide, leading to premature ovarian failure in rats. Reprod. Toxicol. 2018, 80, 60–67. [Google Scholar] [CrossRef]
- Yin, J.; Liu, R.; Jian, Z.; Yang, D.; Pu, Y.; Yin, L.; Wang, D. Di (2-ethylhexyl) phthalate-induced reproductive toxicity involved in dna damage-dependent oocyte apoptosis and oxidative stress in Caenorhabditis elegans. Ecotoxicol. Environ. Saf. 2018, 163, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.C.; Lai, F.N.; Li, L.; Sun, X.F.; Cheng, S.F.; Ge, W.; Wang, Y.F.; Li, L.; Zhang, X.F.; De Felici, M.; et al. Di (2-ethylhexyl) phthalate exposure impairs meiotic progression and DNA damage repair in fetal mouse oocytes in vitro. Cell Death Dis. 2017, 8, e2966. [Google Scholar] [CrossRef]
- Absalan, F.; Saremy, S.; Mansori, E.; Taheri Moghadam, M.; Eftekhari Moghadam, A.R.; Ghanavati, R. Effects of Mono-(2-Ethylhexyl) Phthalate and Di-(2-Ethylhexyl) Phthalate Administrations on Oocyte Meiotic Maturation, Apoptosis and Gene Quantification in Mouse Model. Cell J. 2017, 18, 503–513. [Google Scholar] [PubMed]
- Hannon, P.R.; Peretz, J.; Flaws, J.A. Daily exposure to Di(2-ethylhexyl) phthalate alters estrous cyclicity and accelerates primordial follicle recruitment potentially via dysregulation of the phosphatidylinositol 3-kinase signaling pathway in adult mice. Biol. Reprod. 2014, 90, 136. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Kwon, S.K.; Lee, S.Y.; Baek, K.H. Ubiquitin-specific peptidase 5 and ovarian tumor deubiquitinase 6A are differentially expressed in p53+/+ and p53-/- HCT116 cells. Int. J. Oncol. 2018. [Google Scholar] [CrossRef]
- Hannon, P.R.; Brannick, K.E.; Wang, W.; Gupta, R.K.; Flaws, J.A. Di(2-ethylhexyl) phthalate inhibits antral follicle growth, induces atresia, and inhibits steroid hormone production in cultured mouse antral follicles. Toxicol. Appl. Pharmacol. 2015, 284, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Tzvetkov, N.; Breuer, P. Josephin domain-containing proteins from a variety of species are active de-ubiquitination enzymes. Biol. Chem. 2007, 388, 973–978. [Google Scholar] [CrossRef]
- Weeks, S.D.; Grasty, K.C.; Hernandez-Cuebas, L.; Loll, P.J. Crystal structure of a Josephin-ubiquitin complex: Evolutionary restraints on ataxin-3 deubiquitinating activity. J. Biol. Chem. 2011, 286, 4555–4565. [Google Scholar] [CrossRef]
- Ge, F.; Chen, W.; Qin, J.; Zhou, Z.; Liu, R.; Liu, L.; Tan, J.; Zou, T.; Li, H.; Ren, G.; et al. Ataxin-3 like (ATXN3L), a member of the Josephin family of deubiquitinating enzymes, promotes breast cancer proliferation by deubiquitinating Kruppel-like factor 5 (KLF5). Oncotarget 2015, 6, 21369–21378. [Google Scholar] [CrossRef]
- Cohn, M.A.; Kee, Y.; Haas, W.; Gygi, S.P.; D’Andrea, A.D. UAF1 is a subunit of multiple deubiquitinating enzyme complexes. J. Biol. Chem. 2009, 284, 5343–5351. [Google Scholar] [CrossRef]
- Kee, Y.; Yang, K.; Cohn, M.A.; Haas, W.; Gygi, S.P.; D’Andrea, A.D. WDR20 regulates activity of the USP12 x UAF1 deubiquitinating enzyme complex. J. Biol. Chem. 2010, 285, 11252–11257. [Google Scholar] [CrossRef] [PubMed]
- McClurg, U.L.; Harle, V.J.; Nabbi, A.; Batalha-Pereira, A.; Walker, S.; Coffey, K.; Gaughan, L.; McCracken, S.R.; Robson, C.N. Ubiquitin-specific protease 12 interacting partners Uaf-1 and WDR20 are potential therapeutic targets in prostate cancer. Oncotarget 2015, 6, 37724–37736. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.Y.; Jones, A.; Yang, C.; Zhai, L.; Smith, A.D.t.; Zhang, Z.; Chandrasekharan, M.B.; Sun, Z.W.; Renfrow, M.B.; Wang, Y.; et al. Regulation of histone H2A and H2B deubiquitination and Xenopus development by USP12 and USP46. J. Biol. Chem. 2011, 286, 7190–7201. [Google Scholar] [CrossRef] [PubMed]
- Burska, U.L.; Harle, V.J.; Coffey, K.; Darby, S.; Ramsey, H.; O’Neill, D.; Logan, I.R.; Gaughan, L.; Robson, C.N. Deubiquitinating enzyme Usp12 is a novel co-activator of the androgen receptor. J. Biol. Chem. 2013, 288, 32641–32650. [Google Scholar] [CrossRef]
- McClurg, U.L.; Chit, N.; Azizyan, M.; Edwards, J.; Nabbi, A.; Riabowol, K.T.; Nakjang, S.; McCracken, S.R.; Robson, C.N. Molecular mechanism of the TP53-MDM2-AR-AKT signalling network regulation by USP12. Oncogene 2018, 37, 4679–4691. [Google Scholar] [CrossRef]
- McClurg, U.L.; Azizyan, M.; Dransfield, D.T.; Namdev, N.; Chit, N.; Nakjang, S.; Robson, C.N. The novel anti-androgen candidate galeterone targets deubiquitinating enzymes, USP12 and USP46, to control prostate cancer growth and survival. Oncotarget 2018, 9, 24992–25007. [Google Scholar] [CrossRef]
- Tang, L.J.; Li, Y.; Liu, Y.L.; Wang, J.M.; Liu, D.W.; Tian, Q.B. USP12 regulates cell cycle progression by involving c-Myc, cyclin D2 and BMI-1. Gene 2016, 578, 92–99. [Google Scholar] [CrossRef]
- Zhang, Z.; Jones, A.; Joo, H.Y.; Zhou, D.; Cao, Y.; Chen, S.; Erdjument-Bromage, H.; Renfrow, M.; He, H.; Tempst, P.; et al. USP49 deubiquitinates histone H2B and regulates cotranscriptional pre-mRNA splicing. Genes Dev. 2013, 27, 1581–1595. [Google Scholar] [CrossRef]
- Luo, K.; Li, Y.; Yin, Y.; Li, L.; Wu, C.; Chen, Y.; Nowsheen, S.; Hu, Q.; Zhang, L.; Lou, Z.; et al. USP49 negatively regulates tumorigenesis and chemoresistance through FKBP51-AKT signaling. EMBO J. 2017, 36, 1434–1446. [Google Scholar] [CrossRef]
- Tu, R.; Kang, W.; Yang, X.; Zhang, Q.; Xie, X.; Liu, W.; Zhang, J.; Zhang, X.D.; Wang, H.; Du, R.L. USP49 participates in the DNA damage response by forming a positive feedback loop with p53. Cell Death Dis. 2018, 9, 553. [Google Scholar] [CrossRef]
- Kim, E.; Yoon, S.J.; Kim, E.Y.; Kim, Y.; Lee, H.S.; Kim, K.H.; Lee, K.A. Function of COP9 signalosome in regulation of mouse oocytes meiosis by regulating MPF activity and securing degradation. PLoS ONE 2011, 6, e25870. [Google Scholar] [CrossRef] [PubMed]
- Robker, R.L.; Akison, L.K.; Russell, D.L. Control of oocyte release by progesterone receptor-regulated gene expression. Nucl. Recept. Signal. 2009, 7, e012. [Google Scholar] [CrossRef] [PubMed]
- Khan, D.R.; Landry, D.A.; Fournier, E.; Vigneault, C.; Blondin, P.; Sirard, M.A. Transcriptome meta-analysis of three follicular compartments and its correlation with ovarian follicle maturity and oocyte developmental competence in cows. Physiol. Genomics 2016, 48, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhong, A.; Sun, J.; Chen, M.; Xie, S.; Zheng, H.; Wang, Y.; Yu, Y.; Guo, L.; Lu, R. COPS5 inhibition arrests the proliferation and growth of serous ovarian cancer cells via the elevation of p27 level. Biochem. Biophys Res. Commun. 2017, 493, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Lui, T.T.; Lacroix, C.; Ahmed, S.M.; Goldenberg, S.J.; Leach, C.A.; Daulat, A.M.; Angers, S. The ubiquitin-specific protease USP34 regulates axin stability and Wnt/beta-catenin signaling. Mol. Cell Biol. 2011, 31, 2053–2065. [Google Scholar] [CrossRef] [PubMed]
- Sy, S.M.; Jiang, J.; O, W.S.; Deng, Y.; Huen, M.S. The ubiquitin specific protease USP34 promotes ubiquitin signaling at DNA double-strand breaks. Nucleic Acids Res. 2013, 41, 8572–8580. [Google Scholar] [CrossRef] [PubMed]
- Poalas, K.; Hatchi, E.M.; Cordeiro, N.; Dubois, S.M.; Leclair, H.M.; Leveau, C.; Alexia, C.; Gavard, J.; Vazquez, A.; Bidere, N. Negative regulation of NF-kappaB signaling in T lymphocytes by the ubiquitin-specific protease USP34. Cell Commun. Signal. 2013, 11, 25. [Google Scholar] [CrossRef]
- Zhao, S.; Tian, Y.; Zhang, W.; Xing, X.; Li, T.; Liu, H.; Huang, T.; Ning, Y.; Zhao, H.; Chen, Z.J. An association study between USP34 and polycystic ovary syndrome. J. Ovarian. Res. 2015, 8, 30. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ovarian Cells | DEHP (di-2-ethylhexyl phthalate) |
|---|---|
| A | 0 μg/mL (0 mM) |
| B | 1 μg/mL (2.77 mM) |
| C | 10 μg/mL (27.7 mM) |
| D | 100 μg/mL (277 mM) |
| DUB Gene | Primer Sequence | Group | DUB Gene | Primer Sequence | Group |
|---|---|---|---|---|---|
| Ataxin3 | F: GTC CAA CAG ATG CAT CGA CCA A | G4 | USP12 | F: GAA CTC TGA GTC TGG TTA CAT CCT | G2 |
| R: CGT CTA ACA TTC CTG AGC CAT C | R: GAG GAG CTG GTA TCT CTG ATT TCA | ||||
| ATXN3L | F: TCA GAA GAA AGT GAT GAG TCT GG | G9 | USP13 | F: ACC CAG CTG GAC AAT GGA GTC A | G2 |
| R: CTC TCA ATT GCT CTC GAA CTT G | R: CAG CTT GAT GTC ATT GTC CTG GA | ||||
| BAP1 | F: TCC GTG ATC TGG GTC CTG TC | G11 | USP14 | F: TCA GTG TAT TCG TTC TGT GCC TGA | G2 |
| R: TCC CCG TCT TCT CTC TGC TG | R: CTC GCA TCA TTT GTA TCC AAC ATT CA | ||||
| BRCC3 | F: GAG TTC AGA GTA TGA GAG AAT CG | G4 | USP15 | F: AAA CCT CGC TCC GGA AAG GGG A | G2 |
| R: CCT TTT CTT CTT GTT GTA ATT CCT G | R: CAG TTG GCA ACA GTA TGT AAT CCA A | ||||
| COPS5 | F: GCA GTG GTG ATT GAT CCA ACA A | G5 | USP16 | F: AAA CTT TAG AAC CTG TGT GCA G | G3 |
| R: AGA CCT GAC CAG TGG TAT AGT C | R: CCT GAG AAT TTC TGC CAC AGC C | ||||
| COPS6 | F: AGG TGT TCA AGG AGC TGG AGT T | G5 | USP17 | F: GAG CAA CGC AAG GAG AGC TCA AG | G8 |
| R: GGA AGA TCT GTG TGC TTG GTC A | R: AGG GTA CCT TCG ACT TTT CTG ACG | ||||
| CYLD | F: GCC AAG AAA AAG TCA CTT CAC CC | G11 | USP18 | F: ATT GGA CAG ACC TGC CTT A | G3 |
| R: TGC CTT TTT GCA GAA GGA ATC CT | R: AAG GAT TCC TTC ACC CGG ATC G | ||||
| EIF3S3 | F: GTC CAA ACT CTT CAA ACC ACC A | G5 | USP19 | F: GTT CTT TCC TTC ATC GTC AGG GTC | G3 |
| R: AGT GAA CTC CTT GAT GTT CTG G | R: AGT GGG AGT AGC CAA GAG ATC ATG | ||||
| EIF3S5 | F: TCT GCC TGG TCC TGC TCT TCC A | G4 | USP20 | F: TGG GCT CCT CTT CCA AGT TCT | G11 |
| R: TTG TCG ACA GTT CCC AAC AGG G | R: AGG TTT CAG GTC ATC GTC CTC T | ||||
| JOSD1 | F: GTG AAT GTC ATT ATG GCA GCA C | G4 | USP21 | F: TGA CAA AGC CGG AAG TCC TGT A | G3 |
| R: TCC TCC AAC TCT GAT GAG CCT C | R: AAA GGG CTT CAC AGG TGC CAG A | ||||
| JOSD2 | F: GTG TCT ACT ACA ACC TGG ACT C | G4 | USP22 | F: ACC AAC CAA ACG GGA GCT TG | G11 |
| R: ATG AAG TGC TGG CCT TTC CCA G | R: CCC AAG GTT GAT CAG CCC AC | ||||
| MPND | F: CGG GCA GAC CTT CAA CTC AC | G12 | USP24 | F: CCG ACA GTT GTC CGT GTC TG | G12 |
| R: CCC AGT GGT CTC CGA CTC TT | R: TCC GAA GCT GTA GGC ACG TA | ||||
| OTUB1 | F: AGG AAC CTC AGC AGC AGA AGC A | G6 | USP26 | F: CAG CCA CCT GTG AGA CCT GGT AA | G8 |
| R: GTC TTG CGG ATG TAC GAG TAC T | R: CTG ATA ACT CTC CGC AAG TAA G | ||||
| OTUB2 | F: CAT TCT TCG GGA CCA TCC TGA A | G6 | USP27 | F: CTC CAG CTT TAC GAT CGG TTT AAG | G1 |
| R: GTT CCC ATC CCC TTT GGT CTT | R: CCG AAA CAG CGA CGA CAT CTC AC | ||||
| OTUD1 | F: ATG GGG CAG ATG CTG AAT GTG A | G6 | USP28 | F: GAG GCA GCC CCA ACT GAA TC | G11 |
| R: TGC ACC AGT TGT CGT ACT CTG | R: TGC TCA GAT GAC AAG CAG CG | ||||
| OTUD3 | F: GAA GAC GAC CTG AGA GAT GAA G | G7 | USP29 | F: GGG ATG ACT AAG CTG AAA GAA GCT | G10 |
| R: CTG GGC TCA AGA TTC TCT TCT G | R: TTT CAA AGT TAA ACG CAG GTG ACT | ||||
| OTUD4 | F: GCT CTG CTA TGT GTC AGT CTC T | G7 | USP31 | F: TGA GGA TTG GTG TGG CCG TA | G11 |
| R: TTA CTT GCA ACT GTC ATC CTC TG | R: AAT CTT GTC GCT GCC TGC TC | ||||
| OTUD5 | F: ATC GGA GGA GTC ATG GAT TGA A | G6 | USP33 | F: CCC TTG GTA CTT GTCA GGA TTG TA | G3 |
| R: ACC TGG CGA GCC TGT TTC TCC T | R: AAG CAT AAC ACC ATA CTC GAA GAG | ||||
| OTUD6A | F: TGG ATG ATC CGA AGA GTG AAC | G10 | USP34 | F: CAG CCA TAG TGC TGA AGT TCA AGT | G3 |
| R: TCT TGG AAC TTC TCC AGC TCC T | R: GAC TGA CAT CAC CAG ATT GTG CT | ||||
| OTUD6B | F: AAG AAT GCT GTT CCC AAG AAT G | G7 | USP35 | F: AAG TAC ATG CTC CTG ACC TTC CA | G8 |
| R: CCA TAT GTC TGG CTC CTG TTA A | R: CCC AGG TTG ATG AGA CCA ATC TT | ||||
| OTUD7A | F: GCA GCA CTT CTA CAT GAT CCT A | G8 | USP36 | F: TCC CAG ACA CCC ACA CAC AT | G12 |
| R: TGT GTA GAT TGG CAT CTC CAG G | R: GTG GTG TTG TCC GTG TCT G | ||||
| OTUD7B | F: ACT TCA CAG GGG TGC CTT GTT | G7 | USP37 | F: CAG AAG GAA ACC AGC AGG CA | G12 |
| R: GTT CTT CCC TGT AAC AAC AGG A | R: CGT CCG AGC TAT TCC ACT TCC | ||||
| PARP11 | F: CAG CTA CAA GAT AGA CTT TGC AG | G7 | USP38 | F: CGT GTT GGG CCT CCT TCA TC | G11 |
| R: GAT GGC CTC GTT TTC ACA GAT G | R: TGC AGG GAA GGC AGT AGT GT | ||||
| PRPF8 | F: TCT ATG ACG ACT GGC TCA AGA C | G5 | USP39 | F: GGA GTC TCG CGG TTC CAC T | G12 |
| R: ATC GCC ATG CTT GTT GAC AGT G | R: CGC ACA AAC GGG ACA ACA GA | ||||
| PSMD14 | F: GGT TTG ACA CTT CAG GAC TAC A | G5 | USP41 | F: GGT TCT GCT TCA ATG ACT CCA ATA | G10 |
| R: GAG GTC ATA AGT ACA TCC ACAT G | R: AGC CAT CTC ACG ATT GAC CGG CT | ||||
| PSMD7 | F: ACG TCT TCA ACC TGC TGC CAG A | G5 | USP42 | F: TTA CTC ATC CCA CCC ATA GCC | G1 |
| R: TCC TGC CCT TCT TTC TTC TCT G | R: TCA TGT GAG AGG GAA GCT GTG GT | ||||
| STAMBP | F: GAA GCC CTC CTT AGA TGT GTT | G4 | USP43 | F: GAC AGA GCT GTT TCC TGG GC | G11 |
| R: TGT CCA CCA CAG GTG GCT TAG CT | R: ATA GCT GCA GGC CAC AGA GA | ||||
| STAMBPL1 | F: TTC GAA GAT CAA CTC AAG AAG CA | G5 | USP45 | F: TGG GCT GTT CAG ATC CAG TAG T | G1 |
| R: TCT GGT GTG TGG AAA AGC AGG A | R: ACT GTC AGT CTC CTT GGT GTA CAG | ||||
| TNFAIP3 | F: CCG AGC TGT TCC ACT TGT TAA CA | G6 | USP46 | F: CCA ATC CTG CTG ATG TGG CAG TC | G2 |
| R: CAA CTT TGC GGC ATT GAT GAG A | R: GCT GAT GGC TGG AAA GAT GTA GTA | ||||
| USP1 | F: GAC CAA ATG TGT GAA ATA GGT AAG C | G2 | USP47 | F: CAA TGA TCA ACA TGT CAG CAG GA | G1 |
| R: GCA AGT AAG GAG TAG AAG TAG GAG | R: TTT CTG GCT GGA TCC TTC AGT CT | ||||
| USP2 | F: TAT GGT GCC TAC ACC CCG TCC T | G9 | USP48 | F: GCT GGT AGA TCG GGA TAA TTC CA | G2 |
| R: TGA GGA AGC TGC TGG TGG GGA C | R: AAC TCA TAG GGC TCA GCT CCA G | ||||
| USP3 | F: CCT TGG GTC TGT TTG ACT TGT TCA | G3 | USP49 | F: AGG ACT ACG TGC TCA ATG ATA ACC | G9 |
| R: CCA GTC CCA GCT TGG TGT CAT TA | R: GCA GGA GCA GCC GTG CAC TCT | ||||
| USP4 | F: GTA GAA GGC CAG CAA CCC ATC G | G1 | USP50 | F: CTA TGA TAC CCT TCC AGT TAA GG | G8 |
| R: ACT AGC ACC TGA CCC TGG TAT AG | R: TGG CAT TCA CGC AGC ATG TGT TG | ||||
| USP5 | F: GTC CAC AAA GAC GAG TGC GCC T | G1 | USP51 | F: GGA CCC CAG AGA CTA GGA AAC G | G1 |
| R: AGG CTG AGT CGG CCG ACA GTA | R: CAT AAT CCT TAC ACA TGA AGC A | ||||
| USP6 | F: CGT TGG AAT CAA CAG CAG CAT TGA | G10 | USP52 | F: TCT GGC AAG GTT TCC CTG AGA GA | G2 |
| R: CCA TCC ACT TGC TCG TTC GTG TCA | R: GGT TGC CAT GCA CAT CAA AGT CT | ||||
| USP7 | F: CTC TCA GAC CAT GGG ATT TCC AC | G9 | USP53 | F: GAC ATT TCC AGA GAA TGT GCT CTG | G3 |
| R: ATT GGT GTG TAG ATA TGC CCA CAG | R: GAT CCA GAT TGG AAA TGT GAA AGG | ||||
| USP8 | F: GAC GCC ACC TGC ATC TAT AGA AG | G1 | USP54 | F: CGT GGT AGT GTA CAA GGG ATG TTT | G2 |
| R: GGA AAG TAA AAC TGT CCT GCG CAA | R: CTC CCA TGC ACT TGT GAG TTG TAA | ||||
| USP9X | F: AGC TTC AAG GGT TCC AGG ACA AG | G1 | USPL1 | F: TCC CAA GTG ACA GAT AAA GAA GCT G | G12 |
| R: GAA GAC TAT CTC GCA ACA CTA TGG | R: ACC CAC AGA ACA CGA TGT TAA AGA | ||||
| USP9Y | F: GAG GCT GTG AGT GGC TGG AAG T | G9 | VCPIP1 | F: GCT CGC TAT GGA ATG GAC AAA C | G6 |
| R: CGG ACG TGT ACC ATT GTAAGA TAT G | R: ACA TGC TCT GGT TCT ATG AGG | ||||
| USP10 | F: CCT CCA CAG CCC GCA GTA TAT TT | G3 | YOD1 | F: ACT TGC CCA TCC AAT CTG GTG A | G7 |
| R: GAG ATA GGA TCA TCG CCA CCA TCT | R: ACG TAA CTA GAA GCA CCA CGT T | ||||
| USP11 | F: TGG TGG AAG GCG AGG ATT ATG TG | G2 | ZRANB1 | F: CTA GTG CAA GAC CAA GGG TG | G6 |
| R: GCT GGG CCA AGT GCC ATC TTT C | R: ACA CAT CTT TTA GCC TTG GCC C |
| DUB Gene | Primer Sequence |
|---|---|
| ATXN3L | F: 5′-TCA GAA GAA AGT GAT GAG TCT GG-3′ |
| R: 5′-CTC TCA ATT GCT CTC GAA CTT G-3′ | |
| USP12 | F: 5′-GAA CTC TGA GTC TGG TTA CAT CCT-3′ |
| R: 5′-GAG GAG CTG GTA TCT CTG ATT TCA-3′ | |
| USP49 | F: 5′-CCC TGA ACG CTA TCA CTG CA-3′ |
| R: 5′-TTG GCC AGC ATC TCA GTG AG-3′ | |
| COPS5 | F: 5′-GCA GTG GTG ATT GAT CCA ACA A-3′ |
| R: 5′-AGA CCT GAC CAG TGG TAT AGT C-3′ | |
| USP34 | F: 5′-CAG CCA TAG TGC TGA AGT TCA AGT-3′ |
| R: 5′-GAC TGA CAT CAC CAG ATT GTG CT-3′ | |
| GAPDH | F: 5′-CCC TGA ACG CTA TCA CTG CA-3′ |
| R: 5′-CCA TCA CGC CAC AGT TTC-3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.-H.; Park, J.-H.; Choi, J.; Lee, K.-J.; Yun, B.-S.; Baek, K.-H. Differential Expression of DUB Genes in Ovarian Cells Treated with Di-2-Ethylhexyl Phthalate. Int. J. Mol. Sci. 2020, 21, 1755. https://doi.org/10.3390/ijms21051755
Lee D-H, Park J-H, Choi J, Lee K-J, Yun B-S, Baek K-H. Differential Expression of DUB Genes in Ovarian Cells Treated with Di-2-Ethylhexyl Phthalate. International Journal of Molecular Sciences. 2020; 21(5):1755. https://doi.org/10.3390/ijms21051755
Chicago/Turabian StyleLee, Da-Hye, Jun-Hyeok Park, Jihye Choi, Kyung-Ju Lee, Bo-Seong Yun, and Kwang-Hyun Baek. 2020. "Differential Expression of DUB Genes in Ovarian Cells Treated with Di-2-Ethylhexyl Phthalate" International Journal of Molecular Sciences 21, no. 5: 1755. https://doi.org/10.3390/ijms21051755
APA StyleLee, D.-H., Park, J.-H., Choi, J., Lee, K.-J., Yun, B.-S., & Baek, K.-H. (2020). Differential Expression of DUB Genes in Ovarian Cells Treated with Di-2-Ethylhexyl Phthalate. International Journal of Molecular Sciences, 21(5), 1755. https://doi.org/10.3390/ijms21051755