A Reverse Genetics System for the Israeli Acute Paralysis Virus and Chronic Bee Paralysis Virus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
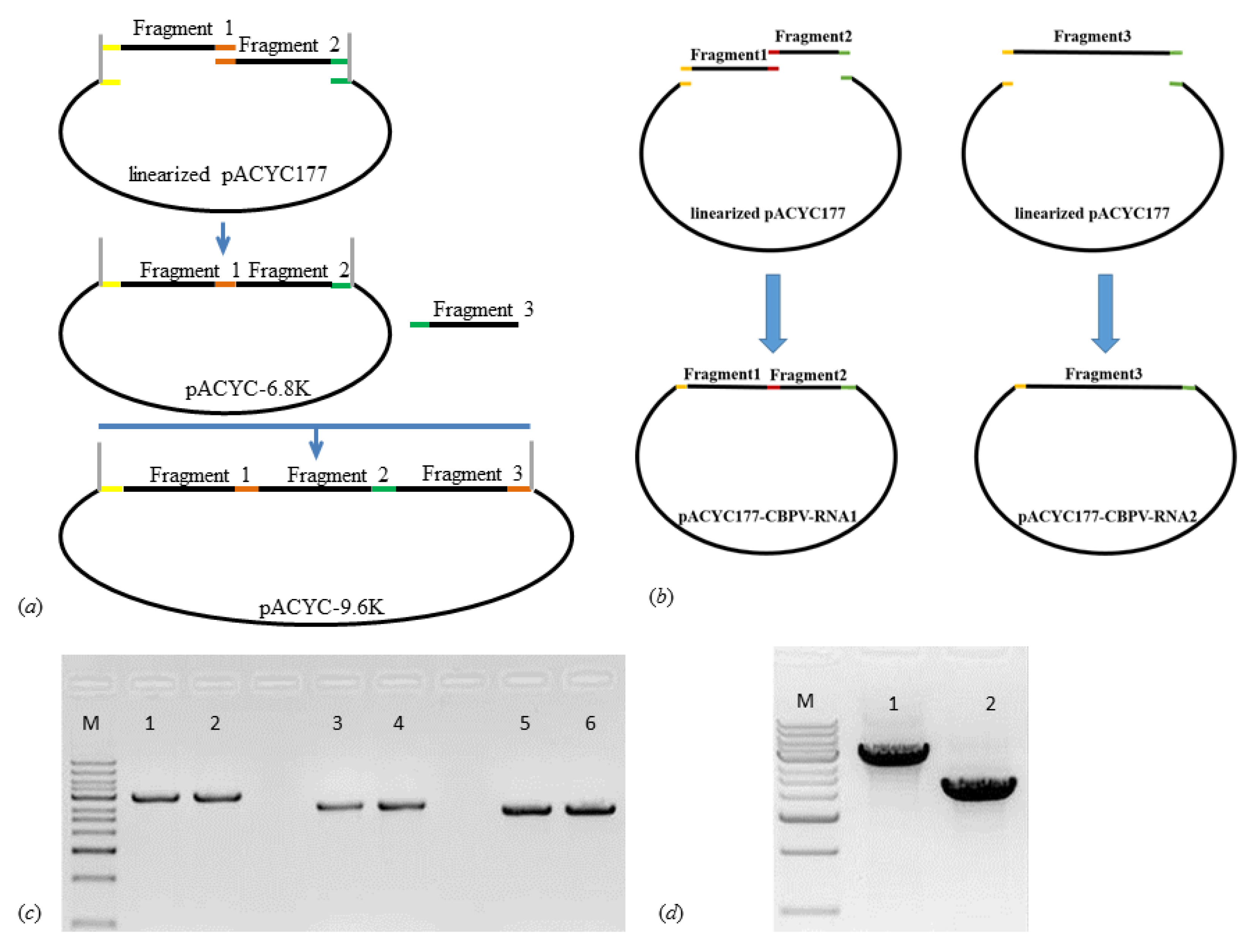
2.1. Construction and Characterization of Infectious Clones
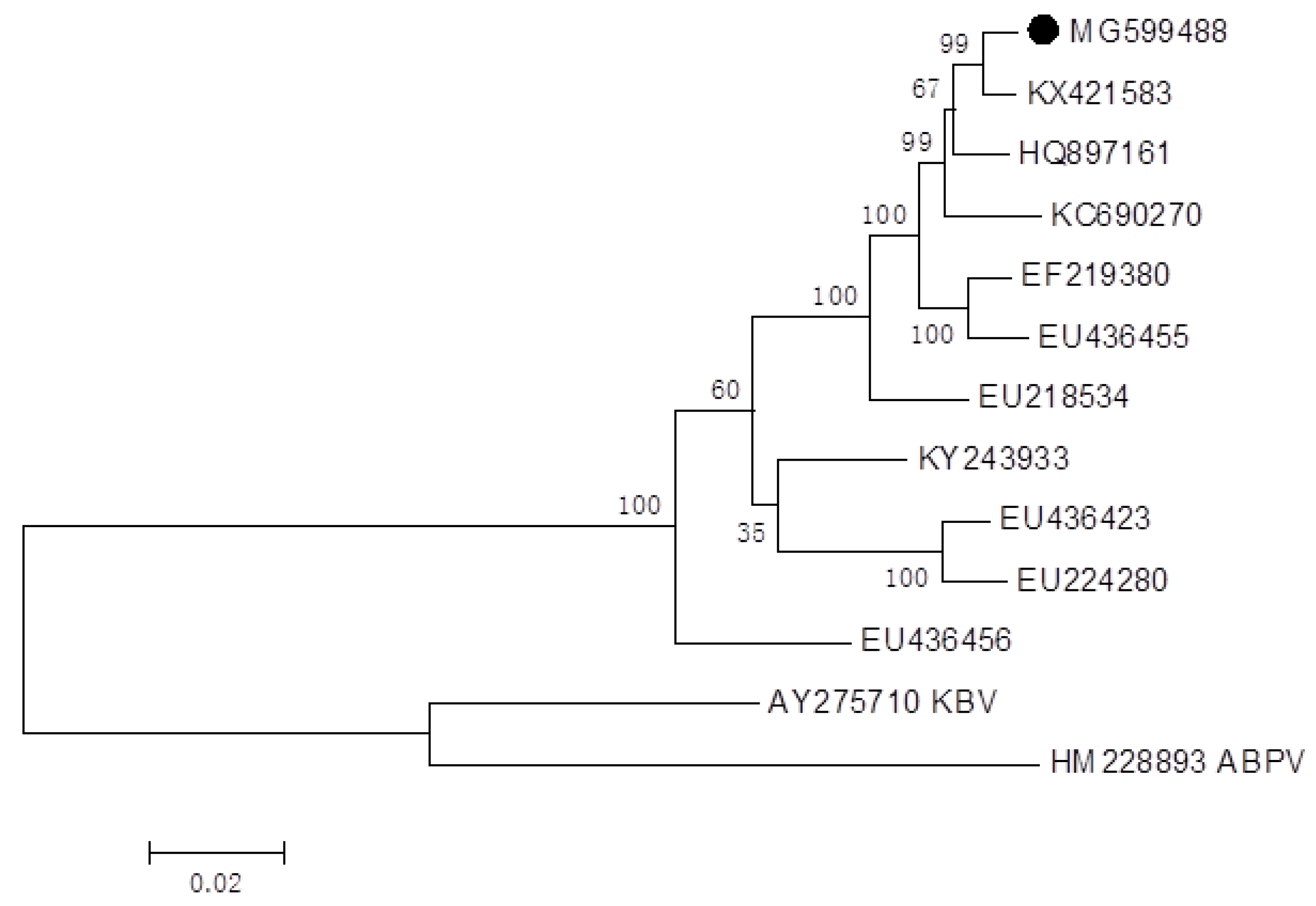
2.2. Genome Sequences of Infectious IAPV and CBPV
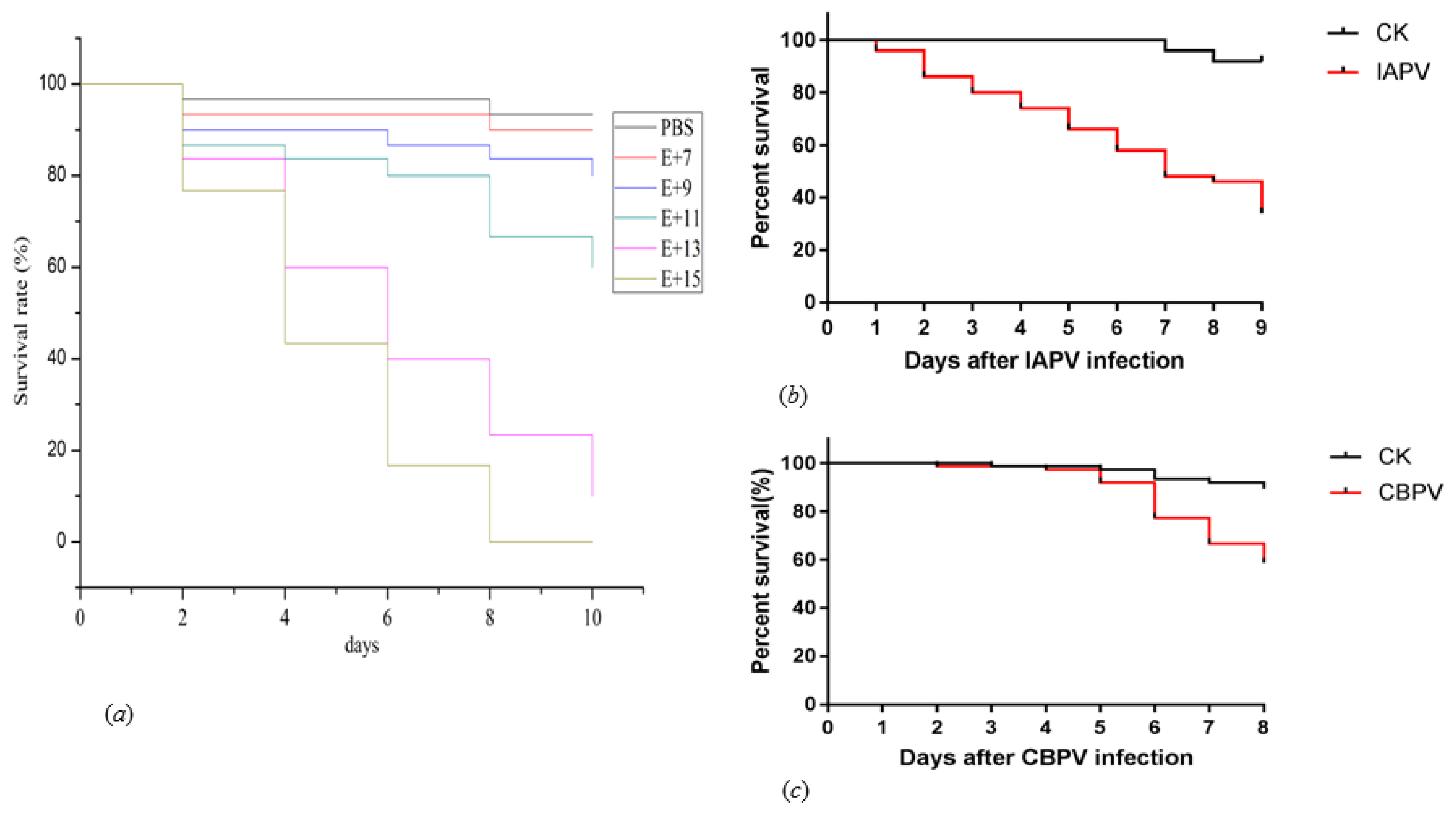
2.3. Assess the Infectivity of Infectious Clones
2.4. Detection the Pathogenesis of Infectious Clones
3. Discussion
4. Materials and Methods
4.1. Honey Bee Samples
4.2. Screening for the Presence of Common Viruses
4.3. Virus Purification
4.4. RNA Extraction and cDNA Synthesis
4.5. Construction of the Infectious Clone and RNA Synthesis
4.6. Assessment the Viral Growth
4.7. Quantification of the Replication Level of Rescue Virus
4.8. Protein Extraction and Western Blot
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Klein, A.M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tschamtke, T. Importance of pollinators in changing landscapes for world crops. Proc. Biol. Sci. 2007, 1608, 303–313. [Google Scholar] [CrossRef]
- Sun, C.Q.; Zhao, Z.J. Chinese agriculture’s dependence on bee pollination: Plantation status of crops depending on bee pollination. Chin. Agri. Sci. Bull. 2016, 8, 13–21. [Google Scholar]
- Cornman, R.S.; Tarpy, D.R.; Chen, Y.P.; Jeffreys, L.; Lopez, D.; Pettis, J.S.; van Engelsdorp, D.; Evans, J.D. Pathogen webs in collapsing honey bee colonies. PLoS ONE 2012, 8, e43562. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 6229, 1255957. [Google Scholar] [CrossRef]
- Shi, M.; Lin, X.D.; Tian, J.H.; Chen, L.J.; Chen, X.; Li, C.X.; Qin, X.C.; Li, J.; Cao, J.P.; Eden, J.S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A metagenomic survey of microbes in honey bee colony collapse disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Maori, E.; Lavi, S.; Mozes-Koch, R.; Gantman, Y.; Peretz, Y.; Edelbaum, O.; Tanne, E.; Sela, I. Isolation and characterization of Israeli acute paralysis virus, a dicistrovirus affecting honeybees in Israel: Evidence for diversity due to intra- and inter-species recombination. J. Gen. Virol. 2007, 88, 3428–3438. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J.; Highfield, A.C.; Laura, B.; Ethel, M.V.; Giles, E.; Budge, M.P.; Powell, M.; Nikaido, S.; Schroeder, D.C. Global honey bee viral landscape altered by a parasitic mite. Science 2012, 336, 1304–1306. [Google Scholar] [CrossRef]
- Diao, Q.Y.; Li, B.B.; Zhao, H.X.; Wu, Y.Y.; Guo, R.; Dai, P.L.; Chen, D.; Wang, Q.; Hou, C.S. Enhancement of chronic bee paralysis virus levels in honeybees acute exposed to imidacloprid: A Chinese case study. Sci. Total Environ. 2018, 630, 487–494. [Google Scholar] [CrossRef]
- Singh, R.; Abby, L.L.; Edwin, G.R.; Edward, C.H.; Ostiguy, N.; Vanengelsdorp, D.; Lipkin, W.I.; Depamphilis, C.W.; Toth, A.L.; Cox-Foster, D.L. RNA viruses in hymenopteran pollinators: Evidence of inter-Taxa virus transmission via pollen and potential impact on non-Apis hymenopteran species. PLoS ONE 2010, 12, e14357. [Google Scholar] [CrossRef]
- Di Prisco, G.; Pennacchio, F.; Caprio, E.; Boncristiani, H.F.; Evans, J.D.; Chen, Y.P. Varroa destructor is an effective vector of Israeli acute paralysis virus in the honeybee, Apis mellifera. J. Gen. Virol. 2011, 1, 151–155. [Google Scholar] [CrossRef]
- Francis, R.M.; Nielsen, S.L.; Kryger, P. Varroa-virus interaction in collapsing honey bee colonies. PLoS ONE 2013, 8, e57540. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Siede, R. Honey bee viruses. Adv. Virus Res. 2007, 1, 33–80. [Google Scholar]
- De Miranda, J.R.; Cordoni, G.; Budge, G. The acute bee paralysis virus–Kashmir bee virus–Israeli acute paralysis virus complex. J. Invertebr. Pathol. 2010, 103, S30–S47. [Google Scholar] [CrossRef]
- Hou, C.S.; Rivkin, H.; Slabezki, Y.; Chejanovsky, N. Dynamics of the presence of Israeli acute paralysis virus in honey bee colonies with colony collapse disorder. Viruses 2014, 6, 2012–2027. [Google Scholar] [CrossRef] [PubMed]
- Palacios, G.; Hui, J.; Quan, P.L.; Kalkstein, A.; Honkavuori, K.S.; Bussetti, A.V.; Conlan, S.; Evans, J.; Chen, Y.P.; van Engelsdorp, D.; et al. Genetic analysis of Israel acute paralysis virus: Distinct clusters are circulating in the United States. J. Virol. 2008, 13, 6209–6217. [Google Scholar] [CrossRef] [PubMed]
- Maori, E.; Tanne, E.; Sela, I. Reciprocal sequence exchange between non-retro viruses and hosts leading to the appearance of new host phenotypes. Virology 2007, 2, 342–349. [Google Scholar] [CrossRef]
- Mullapudi, E.; Přidal, A.; Pálková, L.; de Miranda, J.R.; Plevka, P. Virion structure of Israeli acute bee paralysis virus. J. Virol. 2016, 18, 8150–8159. [Google Scholar] [CrossRef]
- Youssef, I.; Schurr, F.; Goulet, A.; Cougoule, N.; Ribière-Chabert, M.; Darbon, H.; Thiéry, R.; Dubois, E. RNA 1 and RNA 2 genomic segments of chronic bee paralysis virus are infectious and induce chronic bee paralysis disease. J. Immunol. Res. 2015, 2015, 423493. [Google Scholar] [CrossRef]
- Olivier, V.; Blanchard, P.; Chaouch, S.; Lallemand, P.; Schurr, F.; Celle, O.; Dubois, E.; Tordo, N.; Thiéry, R.; Houlgatte, R.; et al. Molecular characterization and phylogenetic analysis of chronic bee paralysis virus, a honey bee virus. Virus Res. 2008, 132, 59–68. [Google Scholar] [CrossRef]
- Fries, I. Nosema ceranae in European honey bees (Apis mellifera). J. Invertebr. Pathol. 2010, 103, S73–S79. [Google Scholar] [CrossRef] [PubMed]
- Coulon, M.; Schurr, F.; Martel, A.C.; Cougoule, N.; Bégaud, A.; Mangoni, P.; Dalmon, A.; Alaux, C.; Le Conte, Y.; Thiéry, R.; et al. Metabolisation of thiamethoxam (a neonicotinoid pesticide) and interaction with the Chronic bee paralysis virus in honeybees. Pestic. Biochem. Physiol. 2018, 144, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Toplak, I.; Jamnikar, C.U.; Aronstein, K.; Gregorc, A. Chronic bee paralysis virus and Nosema ceranae experimental co-infection of winter honey bee workers (Apis mellifera L.). Viruses 2013, 9, 2282–2297. [Google Scholar] [CrossRef] [PubMed]
- Giauffret, A.; Quiot, J.M.; Vago, C.; Poutier, F. In vitro culture of cells of the bee. C. R. Acad. Sci. Hebd. Seances Acad. Sci. D 1967, 11, 800–803. [Google Scholar]
- Monica, B.; Kari, N.; Aamodt, R.M. Long-term maintenance of in vitro cultured honeybee (Apis mellifera) embryonic cells. BMC Dev. Biol. 2006, 1, 17–27. [Google Scholar]
- Chan, M.M.; Choi, S.Y.; Chan, Q.W.; Li, P.; Guarna, M.M.; Foster, L.J. Proteome profile and lentiviral transduction of cultured honey bee (Apis mellifera L.) cells. Insect Mol. Biol. 2010, 5, 653–658. [Google Scholar] [CrossRef]
- Gascuel, J.; Masson, C.; Bermudez, I.; Beadle, D.J. Morphological analysis of honeybee antennal cells growing in primary cultures. Tissue Cell 1994, 4, 551–558. [Google Scholar] [CrossRef]
- Kreissl, S.; Bicker, G. Dissociated neurons of the pupal honeybee brain in cell culture. J. Neurocytol. 1992, 8, 545–556. [Google Scholar] [CrossRef]
- Hunter, W.; Ellis, J.; Vanengelsdorp, D.; Hayes, J.; Westervelt, D.; Glick, E.; Williams, M.; Sela, I.; Maori, E.; Pettis, J.; et al. Large-scale field application of RNAi technology reducing Israeli acute paralysis virus disease in honey bees (Apis mellifera, Hymenoptera: Apidae). PLoS Pathog. 2010, 12, e1001160. [Google Scholar] [CrossRef]
- Xia, X.; Mao, Q.; Wang, H.; Zhou, B.; Wei, T. Replication of Chinese sacbrood virus in primary cell cultures of Asian honeybee (Apis cerana). Arch. Virol. 2014, 12, 3435–3438. [Google Scholar] [CrossRef]
- Goblirsch, M.J.; Spivak, M.S.; Kurtti, T.J. A cell line resource derived from honey bee (Apis mellifera) embryonic tissues. PLoS ONE 2013, 7, e69831. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Tripp, J.; Dolezal, A.G.; Goblirsch, M.J.; Miller, W.A.; Toth, A.L.; Bonning, B.C. In vivo and in vitro infection dynamics of honey bee viruses. Sci. Rep. 2016, 6, 22265. [Google Scholar] [CrossRef]
- Benjeddou, M.; Leat, N.; Allsopp, M.; Davison, S. Development of infectious transcripts and genome manipulation of black queen-cell virus of honey bees. J. Gen. Virol. 2002, 12, 3139–3146. [Google Scholar] [CrossRef] [PubMed]
- Lamp, B.; Url, A.; Seitz, K.; Eichhorn, J.; Riedel, C.; Sinn, L.J.; Indik, S.; Koè, H.; Rümenapf, T. Construction and rescue of a molecular clone of deformed wing virus (DWV). PLoS ONE 2016, 11, e0164639. [Google Scholar] [CrossRef] [PubMed]
- Seitz, K.; Buczolich, K.; Dikunová, A.; Plevka, P.; Power, K.; Rümenapf, T.; Lamp, B. A molecular clone of chronic bee paralysis virus (CBPV) causes mortality in honey bee pupae (Apis mellifera). Sci. Rep. 2019, 1, 16274. [Google Scholar] [CrossRef] [PubMed]
- Ai, H.X.; Yan, X.; Han, R. Occurrence and prevalence of seven bee viruses in Apis mellifera and Apis cerana apiaries in China. J. Invertebr. Pathol. 2012, 1, 160–164. [Google Scholar] [CrossRef]
- Maori, E.; Paldi, N.; Shafir, S.; Kalev, H.; Tsur, E.; Glick, E.; Sela, I. IAPV, a bee-affecting virus associated with Colony Collapse Disorder can be silenced by dsRNA ingestion. Insect Mol. Biol. 2009, 1, 55–60. [Google Scholar] [CrossRef]
- Boncristiani, H.F.; Evans, J.D.; Chen, Y.P.; Pettis, J.; Murphy, C.; Lopez, D.L.; Simone-Finstrom, M.; Strand, M.; Tarpy, D.R.; Rueppell, O. In vitro infection of pupae with Israeli acute paralysis virus suggests disturbance of transcriptional homeostasis in honey bees (Apis mellifera). PLoS ONE 2013, 9, e73429. [Google Scholar] [CrossRef]
- Hunter, W.B. Medium for development of bee cell cultures (Apis mellifera: Hymenoptera: Apidae). In Vitro Cell. Dev. Biol. Anim. 2010, 2, 83–86. [Google Scholar] [CrossRef]
- Nakashima, N.; Uchiumi, T. Functional analysis of structural motifs in dicistroviruses. Virus Res. 2009, 139, 137–147. [Google Scholar] [CrossRef]
- Topisirovic, I.; Svitkin, Y.V.; Sonenberg, N.; Shatkin, A.J. Cap and cap-binding proteins in the control of gene expression. Wiley Interdiscip. Rev. RNA 2011, 2, 277–298. [Google Scholar] [CrossRef] [PubMed]
- Magali, R.; Violaine, O.; Philippe, B. Chronic bee paralysis: A disease and a virus like no other? J. Invertebr. Pathol. 2010, 103, S120–S131. [Google Scholar]
- Chevin, A.; Schurr, F.; Blanchard, P.; Thiéry, R.; Ribière, M. Experimental infection of the honeybee (Apis mellifera L.) with the chronic bee paralysis virus (CBPV): Infectivity of naked CBPV RNAs. Virus Res. 2012, 2, 173–178. [Google Scholar] [CrossRef]
- Yang, S.A.; Gayral, P.; Zhao, H.X.; Wu, Y.J.; Jiang, X.J.; Wu, Y.Y.; Bigot, D.; Wang, X.L.; Yang, D.H.; Herniou, E.A.; et al. Occurrence and molecular phylogeny of honey bee viruses in Vespids. Viruses 2020, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Michaud, S.; Boncristiani, H.F.; Gouw, J.W.; Strand, M.K.; Pettis, J.; Rueppell, O. Response of the honey bee (Apis mellifera) proteome to Israeli acute paralysis virus. Can. J. Zool. 2015, 9, 711–721. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Zhao, H.; Deng, Y.; Deng, S.; Wang, X.; Diao, Q.; Hou, C. A Reverse Genetics System for the Israeli Acute Paralysis Virus and Chronic Bee Paralysis Virus. Int. J. Mol. Sci. 2020, 21, 1742. https://doi.org/10.3390/ijms21051742
Yang S, Zhao H, Deng Y, Deng S, Wang X, Diao Q, Hou C. A Reverse Genetics System for the Israeli Acute Paralysis Virus and Chronic Bee Paralysis Virus. International Journal of Molecular Sciences. 2020; 21(5):1742. https://doi.org/10.3390/ijms21051742
Chicago/Turabian StyleYang, Sa, Hongxia Zhao, Yanchun Deng, Shuai Deng, Xinling Wang, Qingyun Diao, and Chunsheng Hou. 2020. "A Reverse Genetics System for the Israeli Acute Paralysis Virus and Chronic Bee Paralysis Virus" International Journal of Molecular Sciences 21, no. 5: 1742. https://doi.org/10.3390/ijms21051742
APA StyleYang, S., Zhao, H., Deng, Y., Deng, S., Wang, X., Diao, Q., & Hou, C. (2020). A Reverse Genetics System for the Israeli Acute Paralysis Virus and Chronic Bee Paralysis Virus. International Journal of Molecular Sciences, 21(5), 1742. https://doi.org/10.3390/ijms21051742