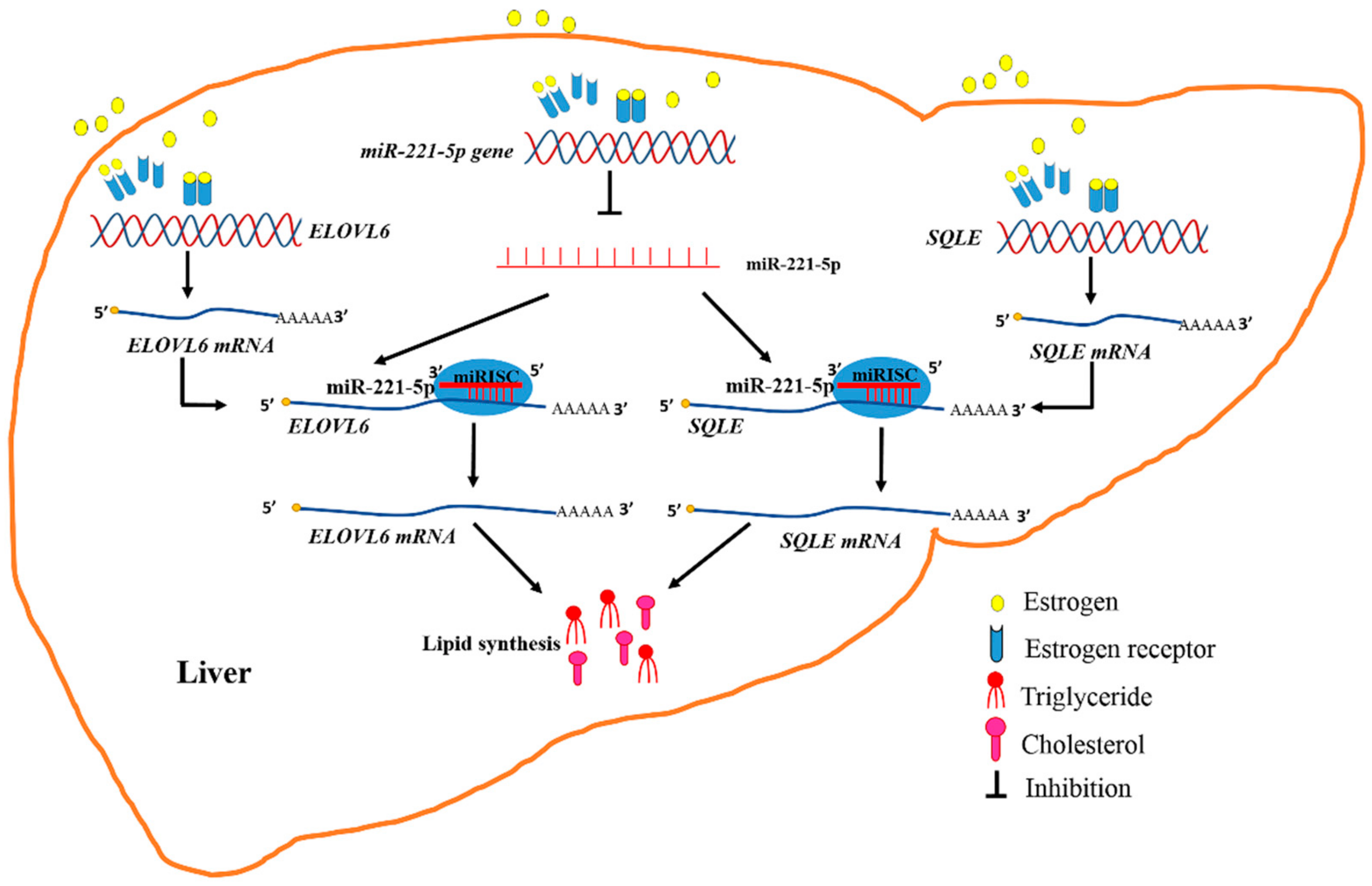
Estrogen Abolishes the Repression Role of gga-miR-221-5p Targeting ELOVL6 and SQLE to Promote Lipid Synthesis in Chicken Liver

 , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Conservative Analysis of miR-221-5p Among Species
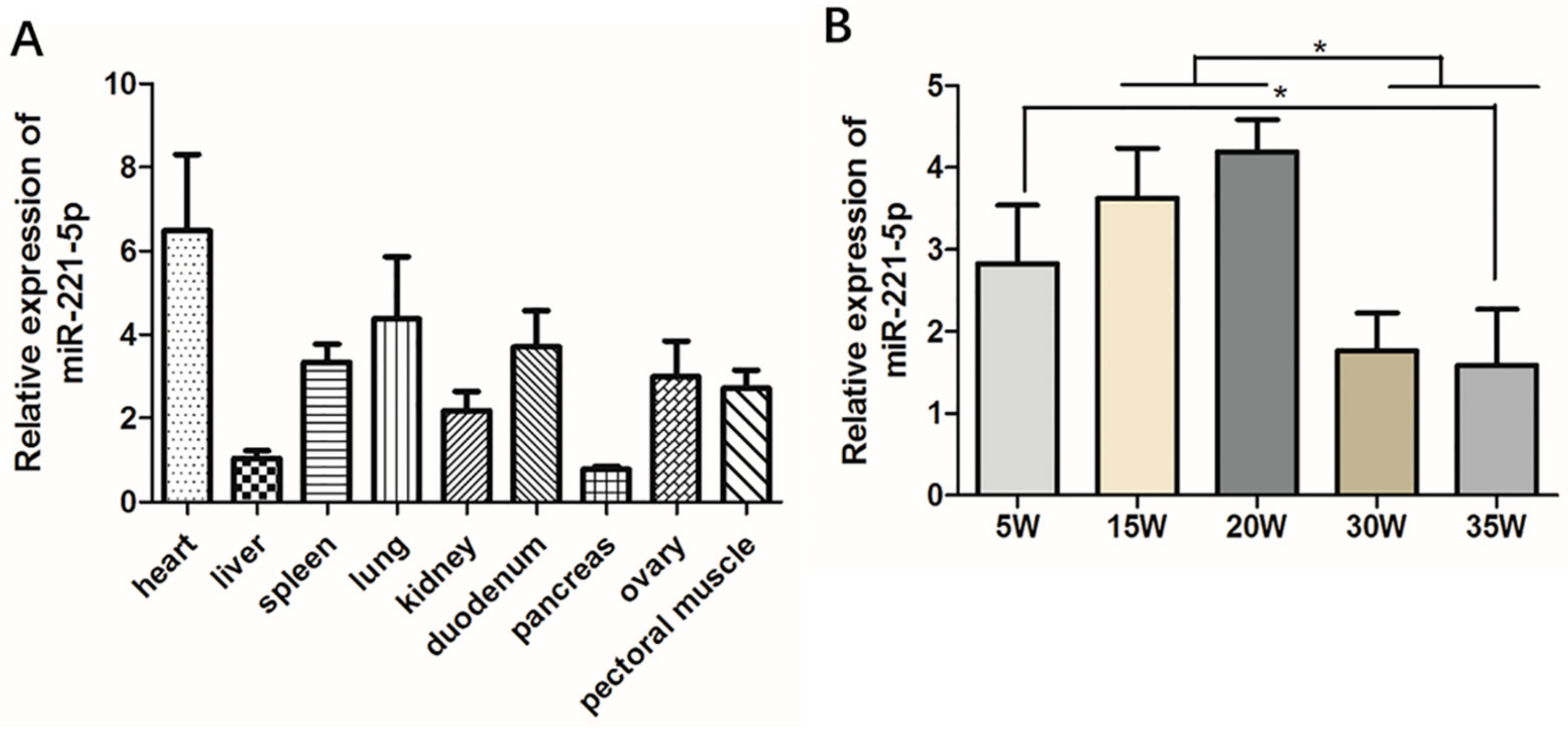
2.2. Spatiotemporal Expression Characteristics of gga-miR-221-5p
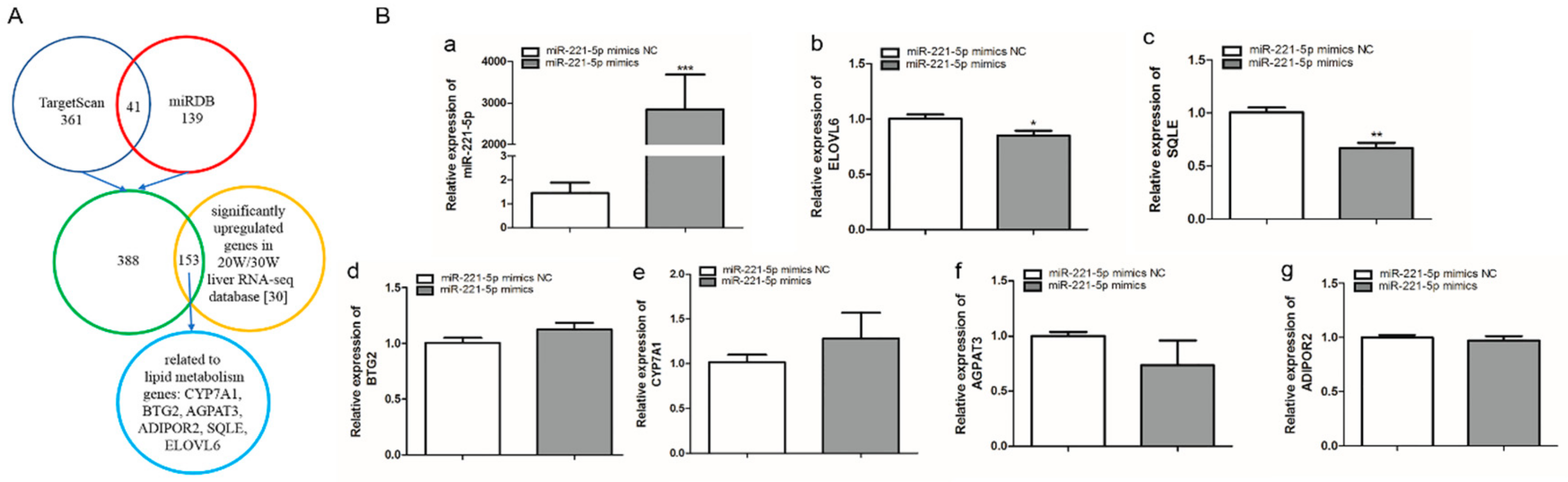
2.3. Screening of gga-miR-221-5p Candidate Target Genes
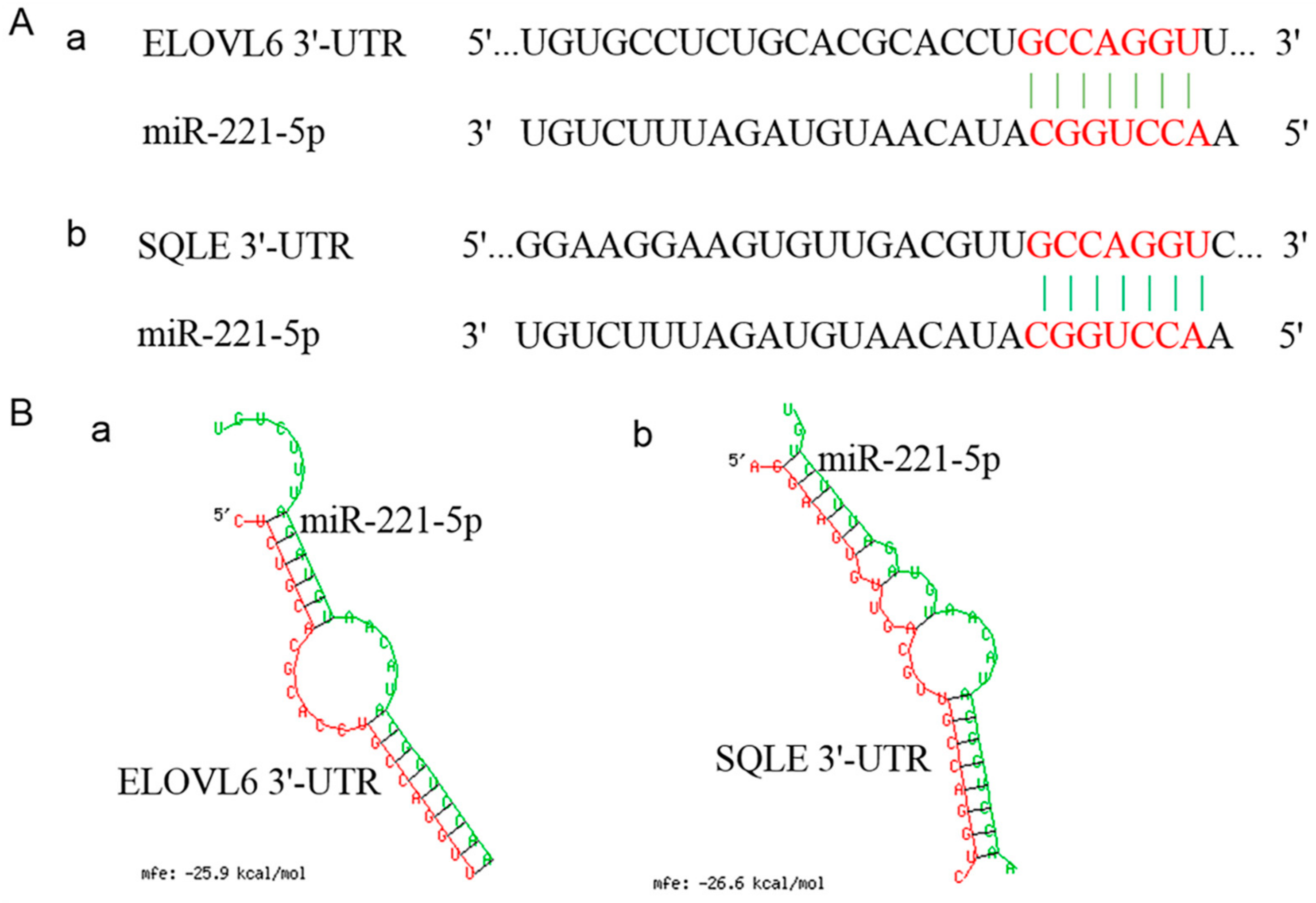
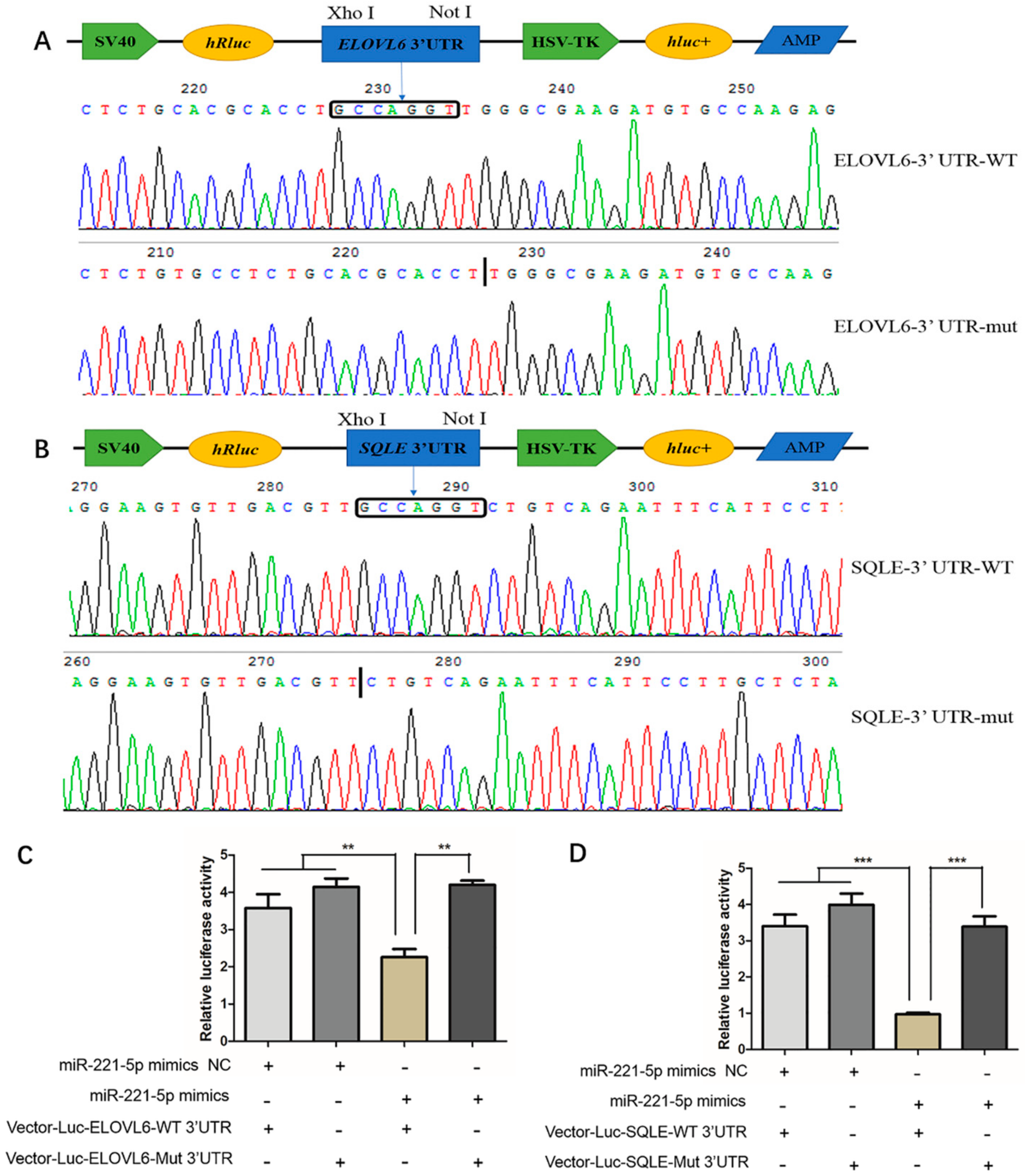
2.4. Verification of the Interaction Relationship between gga-miR-221-5p and Potential Target Genes
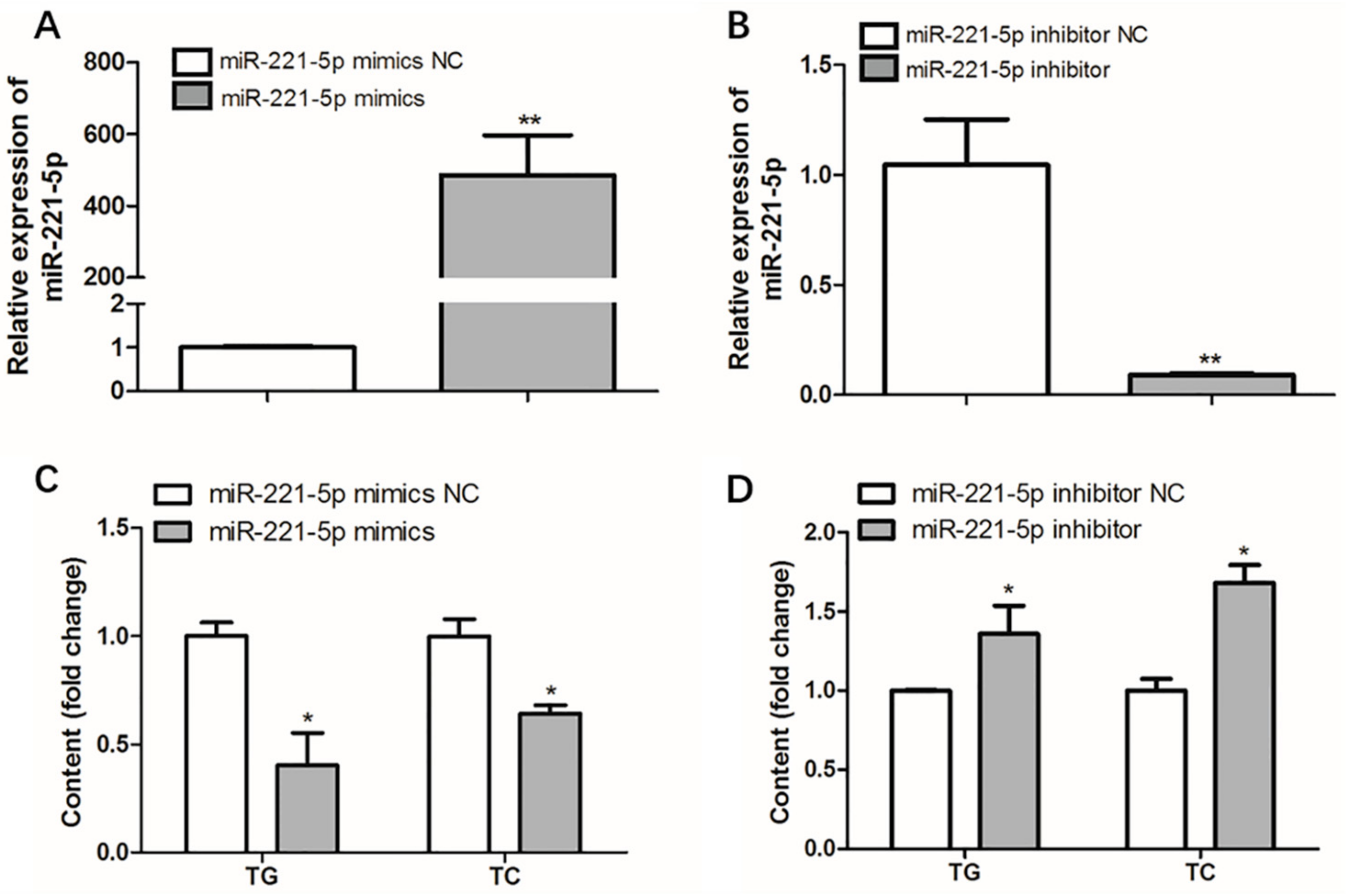
2.5. Effect of gga-miR-221-5p on Intracellular Triglyceride and Total Cholesterol Content
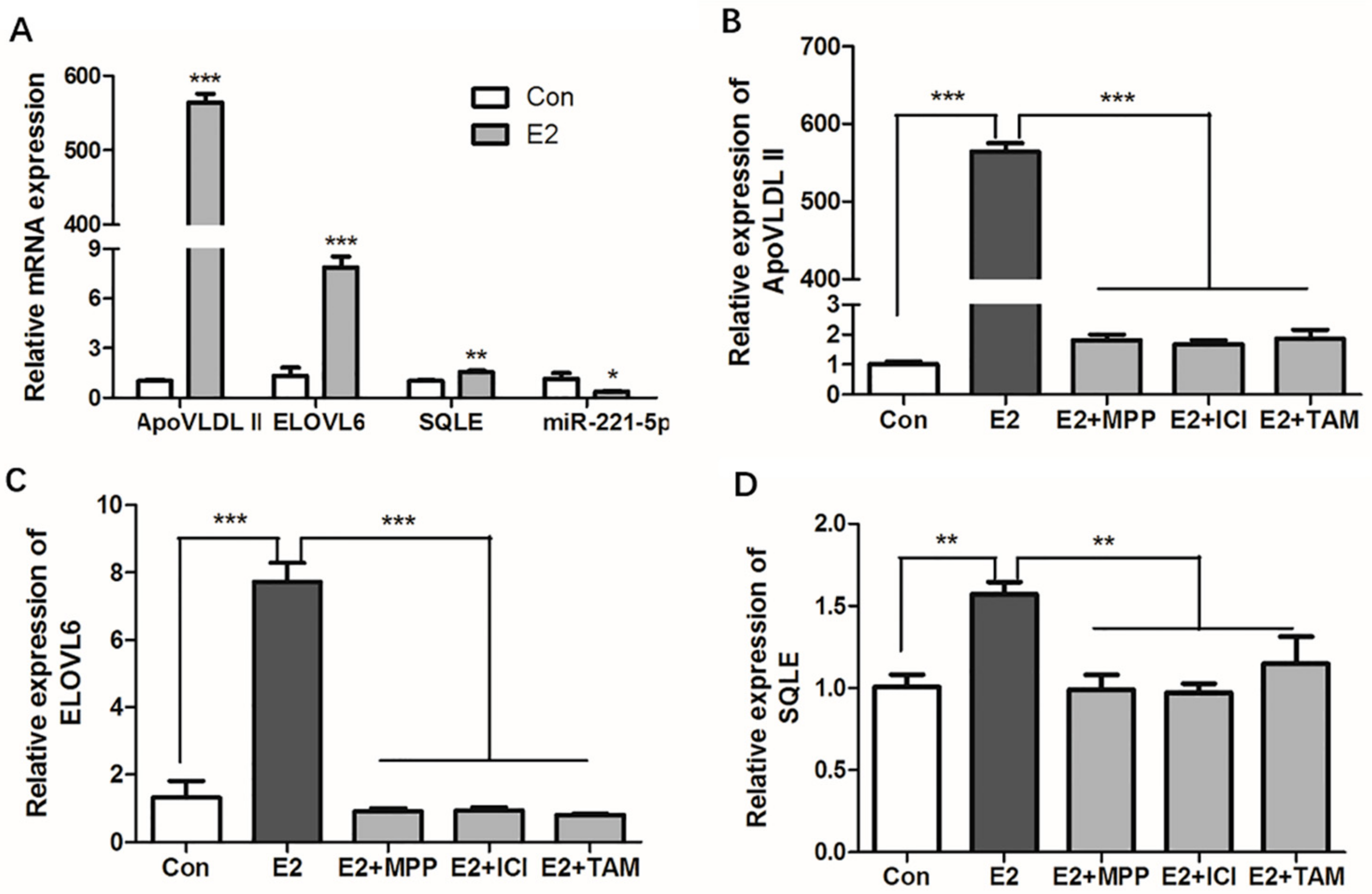
2.6. 17β-Estradiol Repressed the Expression of gga-miR-221-5p and Promoted the Expression of Target Genes in Chicken Primary Hepatocytes
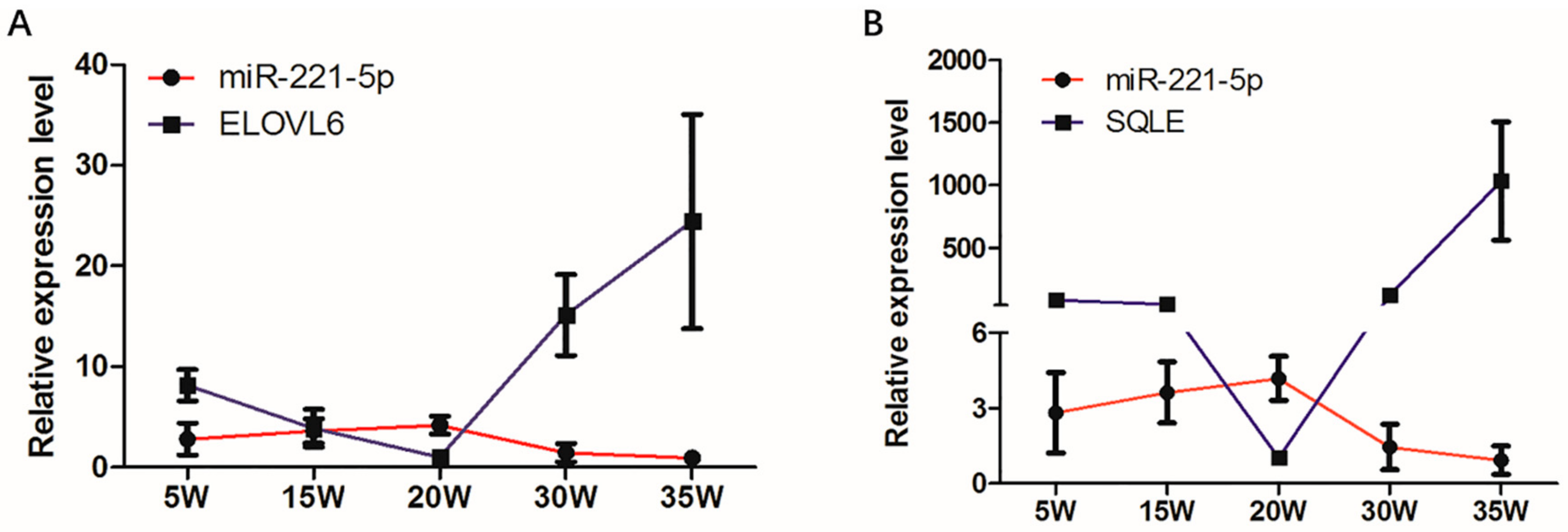
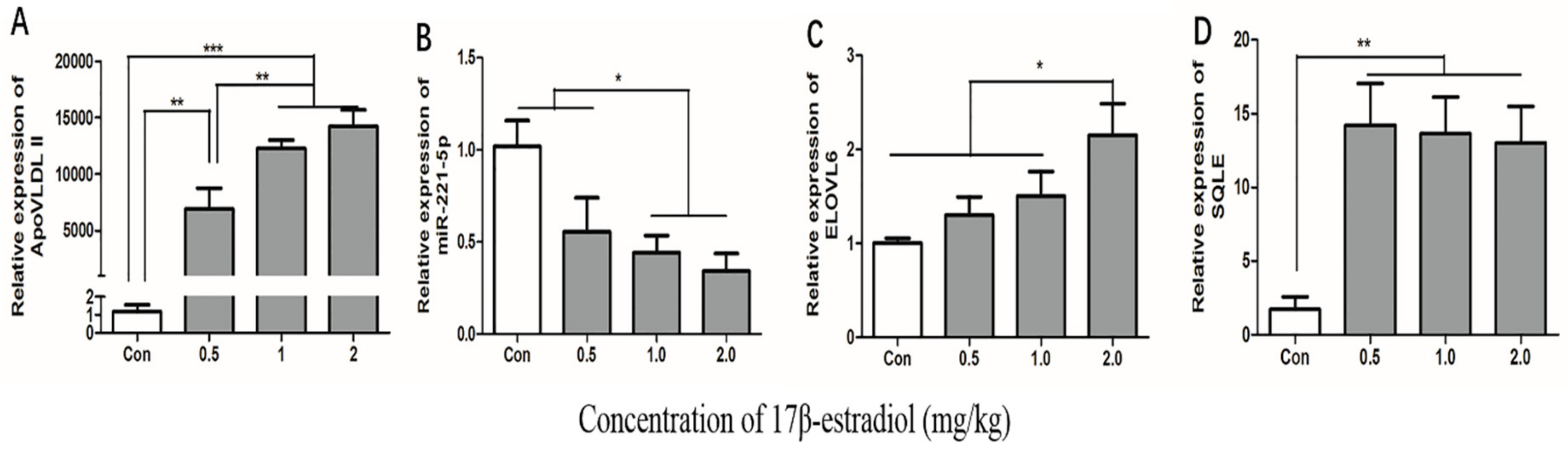
2.7. Effect of 17β-Estradiol on gga-miR-221-5p and the Target Genes in Chicken Liver
3. Discussion
4. Materials and Methods
4.1. Animal Ethics
4.2. Treatment and Sample Collection
4.3. Target Gene Prediction of gga-miR-221-5p and Bioinformatics Analysis
4.4. Vector Construction
4.5. Cell Culture and Treatment
4.6. Dual Luciferase Reporter Assay
4.7. Intracellular Triglyceride and Cholesterol Detection
4.8. cDNA Synthesis and Quantitative Real-Time PCR (qRT-PCR)
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ELOVL6 | elongase of very long chain fatty acids 6 |
| SQLE | squalene epoxidase |
| TG | Triglyceride |
| TC | Total cholesterol |
| VLDL | Very low density lipoprotein |
| MicroRNAs | miRNAs |
| WT | Wild-type |
| Mut | Mutation |
| ApoVLDL II | apovitellenin very low density lipoprotein Ⅱ |
| ERE | estrogen responsive elements |
| ER | estrogen receptor |
| miR-221-5p mimics NC | miR-221-5p mimics negative control |
| AP1 | activator protein-1 |
References
- Shao, F.; Wang, X.; Yu, J.; Jiang, H.; Zhu, B.; Gu, Z. Expression of miR-33 from an SREBF2 Intron targets the FTO gene in the chicken. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Kang, L.; Cui, X.; Zhang, Y.; Yang, C.; Jiang, Y. Identification of miRNAs associated with sexual maturity in chicken ovary by Illumina small RNA deep sequencing. BMC Genom. 2013, 14. [Google Scholar] [CrossRef]
- Brennecke, J.; Stark, A.; Russell, R.B.; Cohen, S.M. Principles of MicroRNA-target recognition. PLoS Biol. 2005, 3, e85. [Google Scholar] [CrossRef]
- Li, H.; Sun, G.R.; Lv, S.J.; Wei, Y.; Han, R.L.; Tian, Y.D.; Kang, X.T. Association study of polymorphisms inside the miR-1657 seed region with chicken growth and meat traits. Br. Poult. Sci. 2012, 53, 770–776. [Google Scholar] [CrossRef]
- Pillai, R.S.; Bhattacharyya, S.N.; Biology, W.F. Repression of protein synthesis by miRNAs: How many mechanisms? Trends Cell Biol. 2007, 17, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Fabian, M.R.; Sonenberg, N. The mechanics of miRNA-mediated gene silencing: A look under the hood of miRISC. Nat. Struct. Mol. Biol. 2012, 19, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Heneghan, H.M.; Miller, N.; Kerin, M.J. Role of microRNAs in obesity and the metabolic syndrome. Obes. Rev. 2010, 11, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Kim, A.Y.; Lee, H.W.; Son, Y.H.; Lee, G.Y.; Lee, J.-W.; Lee, Y.S.; Kim, J.B. miR-27a is a negative regulator of adipocyte differentiation via suppressing PPAR gamma expression. Biochem. Biophy. Res. Commun. 2010, 392, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Xie, H.; Mori, M.A.; Alexander, R.; Yuan, B.; Hattangadi, S.M.; Liu, Q.; Kahn, C.R.; Lodish, H.F. Mir193b-365 is essential for brown fat differentiation. Nat. Cell Biol. 2011, 13, 958–965. [Google Scholar] [CrossRef]
- Moschenross, D.M. Characterization of a Liver Specific microRNA: miR-122. Ph.D. Thesis, University of Connecticut, Mansfield, UK, 2011. [Google Scholar]
- Esau, C.; Davis, S.; Murray, S.F.; Yu, X.X.; Pandey, S.K.; Pear, M.; Watts, L.; Booten, S.L.; Graham, M.; McKay, R.; et al. miR-122 regulation of lipid metabolism revealed by in vivo antisense targeting. Cell Metab. 2006, 3, 87–98. [Google Scholar] [CrossRef]
- Rayner, K.J.; Suarez, Y.; Davalos, A.; Parathath, S.; Fitzgerald, M.L.; Tamehiro, N.; Fisher, E.A.; Moore, K.J.; Fernandez-Hernando, C. MiR-33 contributes to the regulation of cholesterol homeostasis. Science 2010, 328, 1570–1573. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Li, H.; Zheng, H.; Jiang, K.; Yan, F.; Tian, Y.; Kang, X.; Wang, Y.; Liu, X. Hepatic ELOVL6 mRNA is regulated by the gga-miR-22-3p in egg-laying hen. Gene 2017, 623, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Li, H.; Zheng, H.; Jiang, K.; Jia, L.; Yan, F.; Tian, Y.; Kang, X.; Wang, Y.; Liu, X. MicroRNA-101-2-5p Targets the ApoB gene in the liver of chicken (Gallus gallus). Genome 2017, 60, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ma, Z.; Jia, L.; Li, Y.; Xu, C.; Wang, T.; Han, R.; Jiang, R.; Li, Z.; Sun, G.; et al. Systematic analysis of the regulatory functions of microRNAs in chicken hepatic lipid metabolism. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Fu, T.; Choi, S.-E.; Kim, D.-H.; Seok, S.; Suino-Powell, K.M.; Xu, H.E.; Kemper, J.K. Aberrantly elevated microRNA-34a in obesity attenuates hepatic responses to FGF19 by targeting a membrane coreceptor β-Klotho. Proc. Natl. Acad. Sci. USA. 2012, 109, 16137–16142. [Google Scholar] [CrossRef]
- Ortega, F.J.; Moreno-Navarrete, J.M.; Pardo, G.; Sabater, M.; Hummel, M.; Ferrer, A.; Rodriguez-Hermosa, J.I.; Ruiz, B.; Ricart, W.; Peral, B.; et al. MiRNA expression profile of human subcutaneous adipose and during adipocyte differentiation. PLoS ONE 2010, 5. [Google Scholar] [CrossRef]
- O’Hea, E.K.; Leveille, G.A. Lipid biosynthesis and transport in the domestic chick (Gallus domesticus). Comp. Bioche. Physi. 1969, 30, 149–159. [Google Scholar] [CrossRef]
- Walzem, R.L.; Hansen, R.J.; Williams, D.L.; Hamilton, R.L. Estrogen induction of VLDLy assembly in egg-laying hens. J. Nutr. 1999, 129, 467S–472S. [Google Scholar] [CrossRef]
- Ren, J.X.; Tian, W.H.; Yang, L.Y.; Zheng, H.; Li, Z.J.; Yan, F.B.; Wang, Y.B.; Tian, Y.D.; Wang, H.J.; Kang, X.T.; et al. Global investigation of estrogen-responsive genes regulating lipid metabolism in liver of chicken (Gallus gallus). Int. J. Biol. Sci. 2020, in press. [Google Scholar]
- Li, T.; Li, M.; Hu, S.; Cheng, X.; Gao, Y.; Jiang, S.; Yu, Q.; Zhang, C.; Sun, P.; Xian, W.; et al. MiR-221 mediates the epithelial-mesenchymal transition of hepatocellular carcinoma by targeting AdipoR1. Int. J. Biol. Macromol. 2017, 103, 1054–1061. [Google Scholar] [CrossRef]
- Celic, T.; Metzinger-Le Meuth, V.; Six, I.; Massy, Z.A.; Metzinger, L. The mir-221/222 cluster is a key player in vascular biology via the fine-tuning of endothelial cell physiology. Curr. Vasc. Pharmacol. 2017, 15, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Baxi, S.M.J.P.O. Retraction: miR-221 promotes tumorigenesis in human triple negative breast cancer cells. PLoS ONE 2017, 12, e0175869. [Google Scholar] [CrossRef]
- Jin, X.; Cai, L.; Wang, C.; Deng, X.; Yi, S.; Lei, Z.; Xiao, Q.; Xu, H.; Luo, H.; Sun, J. CASC2/miR-24/miR-221 modulates the TRAIL resistance of hepatocellular carcinoma cell through caspase-8/caspase-3. Cell Death. Dis. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-F.; Huang, J.; Li, H.; Zhang, C.; Huang, X.; Tong, G.; Xu, Y.-Z. MicroRNA-221 regulates endothelial nitric oxide production and inflammatory response by targeting adiponectin receptor 1. Gene 2015, 565, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Lustig, Y.; Barhod, E.; Ashwal-Fluss, R.; Gordin, R.; Shomron, N.; Baruch-Umansky, K.; Hemi, R.; Karasik, A.; Kanety, H. RNA-Binding protein PTB and MicroRNA-221 coregulate AdipoR1 translation and adiponectin signaling. Diabetes 2014, 63, 433–445. [Google Scholar] [CrossRef]
- Lian, L.; Zhang, D.; Wang, Q.; Yang, N.; Qu, L.J. The inhibitory effects of gga-miR-199-3p, gga-miR-140-3p, and gga-miR-221-5p in Marek’s disease tumorigenesis. Poul. Sci. 2015, 94, 2131–2135. [Google Scholar] [CrossRef]
- Li, H.; Ji, J.; Xie, Q.; Shang, H.; Zhang, H.; Xin, X.; Chen, F.; Sun, B.; Xue, C.; Ma, J.; et al. Aberrant expression of liver microRNA in chickens infected with subgroup J avian leukosis virus. Virus Res. 2012, 169, 268–271. [Google Scholar] [CrossRef]
- Dai, Z.; Ji, J.; Yan, Y.; Lin, W.; Li, H.; Chen, F.; Liu, Y.; Chen, W.; Bi, Y.; Xie, Q. Role of gga-miR-221 and gga-miR-222 during tumour formation in chickens infected by subgroup J avian leukosis virus. Viruses 2015, 7, 6538–6551. [Google Scholar] [CrossRef]
- Li, H.; Wang, T.A.; Xu, C.L.; Wang, D.D.; Ren, J.X.; Li, Y.M.; Tian, Y.D.; Wang, Y.B.; Jiao, Y.P.; Kang, X.T.; et al. Transcriptome profile of liver at different physiological stages reveals potential mode for lipid metabolism in laying hens. BMC Genom. 2015, 16, 763. [Google Scholar] [CrossRef]
- Klein-Hitpass, L.; Ryffel, G.U.; Heitlinger, E.; Cato, A.C. A 13 bp palindrome is a functional estrogen responsive element and interacts specifically with estrogen receptor. Nucleic Acids Res. 1988, 16, 647–663. [Google Scholar] [CrossRef]
- Pollock, A.; Bian, S.; Zhang, C.; Chen, Z.; Sun, T. Growth of the developing cerebral cortex is controlled by MicroRNA-7 through the p53 Pathway. Cell Rep. 2014, 8, 1184–1196. [Google Scholar] [CrossRef]
- Ng, R.; Wu, H.; Xiao, H.; Chen, X.; Willenbring, H.; Steer, C.J.; Song, G. Inhibition of MicroRNA-24 expression in liver prevents hepatic lipid accumulation and hyperlipidemia. Hepatology 2014, 60, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Shao, N.; Ma, G.; Zhang, J.; Zhu, W. miR-221-5p enhances cell proliferation and metastasis through post-transcriptional regulation of SOCS1 in human prostate cancer. BMC Urol. 2018, 18. [Google Scholar] [CrossRef] [PubMed]
- Krek, A.; Grun, D.; Poy, M.N.; Wolf, R.; Rosenberg, L.; Epstein, E.J.; MacMenamin, P.; da Piedade, I.; Gunsalus, K.C.; Stoffel, M.; et al. Combinatorial microRNA target predictions. Nat. Genet. 2005, 37, 495–500. [Google Scholar] [CrossRef]
- Shimano, H. Novel qualitative aspects of tissue fatty acids related to metabolic regulation: Lessons from Elovl6 knockout. Prog. Lipid Res. 2012, 51, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Corominas, J.; Marchesi, J.A.P.; Puig-Oliveras, A.; Revilla, M.; Estelle, J.; Alves, E.; Folch, J.M.; Ballester, M. Epigenetic regulation of the ELOVL6 gene is associated with a major QTL effect on fatty acid composition in pigs. Genet. Sel. Evol. 2015, 47. [Google Scholar] [CrossRef]
- D’Andre, H.C.; Paul, W.; Shen, X.; Jia, X.; Zhang, R.; Sun, L.; Zhang, X. Identification and characterization of genes that control fat deposition in chickens. J. Anim. Sci. Biotechnol. 2013, 4. [Google Scholar] [CrossRef]
- Lorbek, G.; Perse, M.; Horvat, S.; Bjorkhem, I.; Rozman, D. Sex Differences in the Hepatic Cholesterol Sensing Mechanisms in Mice. Molecules 2013, 18, 11067–11085. [Google Scholar] [CrossRef]
- Stopsack, K.H.; Gerke, T.A.; Andren, O.; Andersson, S.-O.; Giovannucci, E.L.; Mucci, L.A.; Rider, J.R. Cholesterol uptake and regulation in high-grade and lethal prostate cancers. Carcinogenesis 2017, 38, 806–811. [Google Scholar] [CrossRef]
- Helms, M.W.; Kemming, D.; Pospisil, H.; Vogt, U.; Buerger, H.; Korsching, E.; Liedtke, C.; Schlotter, C.M.; Wang, A.; Chan, S.Y.; et al. Squalene epoxidase, located on chromosome 8q24.1, is upregulated in 8q+ breast cancer and indicates poor clinical outcome in stage I and II disease. Br. J. Cancer 2008, 99, 774–780. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, W.; Zhang, K.; Zheng, H.; Ma, Y.; Lin, D.; Zhang, X.; Feng, L.; Lei, W.; Zhang, Z.; et al. Identification of genes differentially expressed in human primary lung squamous cell carcinoma. Lung Cancer 2007, 56, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, Y.; Nakamura, T.; Tanase, H.; Doi, O. Comparisons of plasma LH, progesterone, testosterone and estradiol concentrations in male and female chickens (Gallus domesticus) from 28 to 1141 days of age. Endocrinol. Jpn. 1981, 28, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.; Harvey, S. Plasma concentrations of luteinizing hormone growth hormone, oestradiol, testosterone and androstenedione in the domestic hen from 6 weeks of age to sexual maturity. Reprod. Nutr. Dev. 1986, 26, 515–522. [Google Scholar] [CrossRef]
- Palmisano, B.T.; Zhu, L.; Stafford, J.M. Role of Estrogens in the regulation of liver lipid metabolism. Adv. Exp. Med. Biol. 2017, 1043, 227–256. [Google Scholar] [CrossRef]
- Pellegrini, M.; Pallottini, V.; Marin, R.; Marino, M. Role of the sex hormone estrogen in the prevention of lipid disorder. Curr. Med. Chem. 2014, 21, 2734–2742. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, Z.; Ou, K.; Wang, T.; Li, Z.; Tian, Y.; Wang, Y.; Kang, X.; Li, H.; Liu, X. Evolution, dynamic expression changes and regulatory characteristics of gene families involved in the glycerophosphate pathway of triglyceride synthesis in chicken (Gallus gallus). Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Ivessa, N.E.; Rehberg, E.; Kienzle, B.; Seif, F.; Hermann, R.; Hermann, M.; Schneider, W.J.; Gordon, D.A. Molecular cloning, expression, and hormonal regulation of the chicken microsomal triglyceride transfer protein. Gene 2013, 523, 1–9. [Google Scholar] [CrossRef]
- Lu, T.T.; Repa, J.J.; Mangelsdorf, D.J. Orphan nuclear receptors as eLiXiRs and FiXeRs of sterol metabolism. J. Biol. Chem. 2001, 276, 37735–37738. [Google Scholar] [CrossRef]
- Peet, D.J.; Turley, S.D.; Ma, W.; Janowski, B.; Lobaccare, J.; Hammer, R.; Mangelsdorf, D. Cholesterol and bile acid metabolism are impaired in mice lacking the nuclear oxysterol receptor LXRα. Cell 1998, 93, 693–704. [Google Scholar] [CrossRef]
- Mcfadden, J.W.; Corl, B.A. Activation of liver X receptor (LXR) enhances de novo fatty acid synthesis in bovine mammary epithelial cells. J. Dairy Sci. 2010, 93, 4651–4658. [Google Scholar] [CrossRef]
- Darimont, C.; Avanti, O.; Zbinden, I.; Leonevautravers, P.; Mansourian, R.; Giusti, V.; Mace, K. Liver X receptor preferentially activates de novo lipogenesis in human preadipocytes. Biochimie 2006, 88, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Breevoort, S.R.; Angdisen, J.; Mingui, F.; Daniel, R.S.; Sam, R.H.; Steven, A.K.; David, J.M.; Ira, G.S. Liver LXRα expression is crucial for whole body cholesterol homeostasis and reverse cholesterol transport in mice. J. Clin. Investig. 2012, 122, 1688–1699. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Zheng, H.; Yang, L.; Li, H.; Tian, Y.; Wang, Y.; Lyu, S.; Brockmann, G.A.; Kang, X.; Liu, X. Dynamic expression profile, regulatory mechanism and correlation with egg-laying performance of ACSF gene family in chicken (Gallus gallus). Sci. Rep. 2018, 8, 8457–8467. [Google Scholar] [CrossRef] [PubMed]
- Paech, K.; Webb, P.; Kuiper, G.J.M.; Nilsson, S.; Gustafsssn, J.A.; Kushiser, P.J.; Scanlan, T.S. Differential ligand activation of estrogen receptors ER and ER at AP1 sites. Science 1997, 277, 1508–1510. [Google Scholar] [CrossRef]
- Petz, L.N.; Ziegler, Y.S.; Schultz, J.R.; Nardulli, A.M. Fos and Jun inhibit estrogen-induced transcription of the human progesterone receptor gene through an activator protein-1 site. Mol. Endocrinol. 2004, 18, 521–532. [Google Scholar] [CrossRef]
- Xiao, B.; Zhang, W.; Chen, L.; Hang, J.; Wang, L.; Zhang, R.; Liao, Y.; Chen, J.; Ma, Q.; Sun, Z.; et al. Analysis of the miRNA–mRNA–lncRNA network in human estrogen receptor-positive and estrogen receptor-negative breast cancer based on TCGA data. Gene 2018, 658, 28–35. [Google Scholar] [CrossRef]
- Zhao, X.-Y.; Li, S.; Wang, G.-X.; Yu, Q.; Lin, J.D. A long noncoding RNA transcriptional regulatory circuit drives thermogenic adipocyte differentiation. Mol. Cell 2014, 55, 372–382. [Google Scholar] [CrossRef]
- Wagschal, A.; Najafi-Shoushtari, S.H.; Wang, L.; Goedeke, L.; Sinha, S.; deLemos, A.S.; Black, J.C.; Ramirez, C.M.; Li, Y.; Tewhey, R.; et al. Genome-wide identification of microRNAs regulating cholesterol and triglyceride homeostasis. Nat. Med. 2015, 21, 1290–1297. [Google Scholar] [CrossRef]
- Li, Y.; Song, Y.; Zhao, M.; Guo, Y.; Yu, C.; Chen, W.; Shao, S.; Xu, C.; Zhou, X.; Zhao, L.J.H. A novel role for CRTC2 in hepatic cholesterol synthesis via SREBP-2. Hepatology 2017, 66, 481–497. [Google Scholar] [CrossRef]
- Cheng, Y.; Meng, Q.; Wang, C.; Li, H.; Huang, Z.; Chen, S.; Xiao, F.; Guo, F. Leucine deprivation decreases fat mass by stimulation of lipolysis in white adipose tissue and upregulation of uncoupling protein 1 (UCP1) in brown adipose tissue. Diabetes 2010, 59, 17–25. [Google Scholar] [CrossRef]
- Marquart, T.J.; Allen, R.M.; Ory, D.S.; Baldan, A. miR-33 links SREBP-2 induction to repression of sterol transporters. Proc. Natl. Acad. Sci. USA 2010, 107, 12228–12232. [Google Scholar] [CrossRef] [PubMed]
- Gerin, I.; Clerbaux, L.A.; Haumont, O.; Lanthier, N.; Das, A.K.; Burant, C.F.; Leclercq, I.A.; MacDougald, O.A.; Bommer, G.T. Expression of miR-33 from an SREBP2 intron inhibits cholesterol export and fatty acid oxidation. J. Biol. Chem. 2010, 285, 33652–33661. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zalzala, M.; Xu, J.; Li, Y.; Yin, L.; Zhang, Y. A metabolic stress-inducible miR-34a-HNF4α pathway regulates lipid and lipoprotein metabolism. Nat. Commun. 2015, 6, 7466. [Google Scholar] [CrossRef]
- Tian, W.H.; Wang, Z.; Yue, Y.X.; Li, H.; Li, Z.J.; Han, R.L.; Tian, Y.D.; Kang, X.T.; Liu, X.J. miR-34a-5p increases hepatic triglycerides and total cholesterol levels by regulating ACSL1 protein expression in laying hens. Int. J. Mol. Sci. 2019, 20, 4420. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5’~3’) |
|---|---|
| ELOVL6-WT | F: ccgCTCGAGACTCTTGCCAGCGAGCGGCCC |
| R: atttGCGGCCGCAGCTAAGCCATTGCA GCAAAATC | |
| ELOVL6-overlap | F:CTGTGCCTCTGCACGCACCTGCCAGGTTGGGCGAAGATGTGCCAAGA |
| R:TCTTGGCACATCTTCGCCCAACCTGGCAGGTGCGTGCAGAGGCACAG | |
| SQLE-WT | F: ccgCTCGAGCCAAGGGGATGTGACTGGAC |
| R: atttGCGGCCGCAGTGATGCACTCTGCAATGGAT | |
| SQLE-overlap | F:CGGAAGGAAGTGTTGACGTTGCCAGGTCTGTCAGAATTTCATTCCTT |
| R:AAGGAATGAAATTCTGACAGACCTGGCAACGTCAACACTTCCTTCC |
| Gene | Primer Sequence (5′~3′) | Annealing Temperature (°C) | Product Length (bp) | Accession Number |
|---|---|---|---|---|
| BTG2 | F: GCTCGCAGAGCACTACAAACA | 60 | 127 | XM_418053.6 |
| R: GAGTCCGATCTGGCTAGCTG | ||||
| CYP27A1 | F: GTGGACACGACCTCCAACAC | 60 | 148 | XM_422056.6 |
| R: GCATCGGCATCTTGGGGATA | ||||
| ADIPOR2 | F: GGGAGGCGGTAGCGATTG | 61 | 121 | NM_001007854.1 |
| R: GATTATGCGTTGGCTCTGGC | ||||
| AGPAT3 | F: GTCTCTCCATGACCGCTGAG | 59 | 237 | XM_004934600.3 |
| R: AACTGTCCTTTCTTTGTGCCG | ||||
| SQLE | F: GAATTGTTGCAGCCTGGTGG | 60 | 241 | NM_001194927.1 |
| R: ATTTTGCATTGGGCTCTGCC | ||||
| ELOVL6 | F: GTGGATGCAGGAGAACTGGAAG | 60 | 80 | NM_001031539.1 |
| R: TTAGGTGCCGACCACCAAAT | ||||
| β-actin | F: GAGAGAAGATGACACAGATC | 60 | 116 | NM_205518.1 |
| R: GTCCATCACAATACCAGTGG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, D.-D.; Wang, D.-D.; Wang, Z.; Wang, Y.-B.; Li, G.-X.; Sun, G.-R.; Tian, Y.-D.; Han, R.-L.; Li, Z.-J.; Jiang, R.-R.; et al. Estrogen Abolishes the Repression Role of gga-miR-221-5p Targeting ELOVL6 and SQLE to Promote Lipid Synthesis in Chicken Liver. Int. J. Mol. Sci. 2020, 21, 1624. https://doi.org/10.3390/ijms21051624
Zhang D-D, Wang D-D, Wang Z, Wang Y-B, Li G-X, Sun G-R, Tian Y-D, Han R-L, Li Z-J, Jiang R-R, et al. Estrogen Abolishes the Repression Role of gga-miR-221-5p Targeting ELOVL6 and SQLE to Promote Lipid Synthesis in Chicken Liver. International Journal of Molecular Sciences. 2020; 21(5):1624. https://doi.org/10.3390/ijms21051624
Chicago/Turabian StyleZhang, Ding-Ding, Dan-Dan Wang, Zhang Wang, Yang-Bin Wang, Guo-Xi Li, Gui-Rong Sun, Ya-Dong Tian, Rui-Li Han, Zhuan-Jian Li, Rui-Rui Jiang, and et al. 2020. "Estrogen Abolishes the Repression Role of gga-miR-221-5p Targeting ELOVL6 and SQLE to Promote Lipid Synthesis in Chicken Liver" International Journal of Molecular Sciences 21, no. 5: 1624. https://doi.org/10.3390/ijms21051624
APA StyleZhang, D.-D., Wang, D.-D., Wang, Z., Wang, Y.-B., Li, G.-X., Sun, G.-R., Tian, Y.-D., Han, R.-L., Li, Z.-J., Jiang, R.-R., Liu, X.-J., Kang, X.-T., & Li, H. (2020). Estrogen Abolishes the Repression Role of gga-miR-221-5p Targeting ELOVL6 and SQLE to Promote Lipid Synthesis in Chicken Liver. International Journal of Molecular Sciences, 21(5), 1624. https://doi.org/10.3390/ijms21051624