Monocarboxylate Transporter 1 (MCT1) in Liver Pathology

 ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. mRNA Quantification
2.2. Protein Abundance
2.3. mRNA/Protein Correlation
2.4. Correlation between miRNAs and MCT1
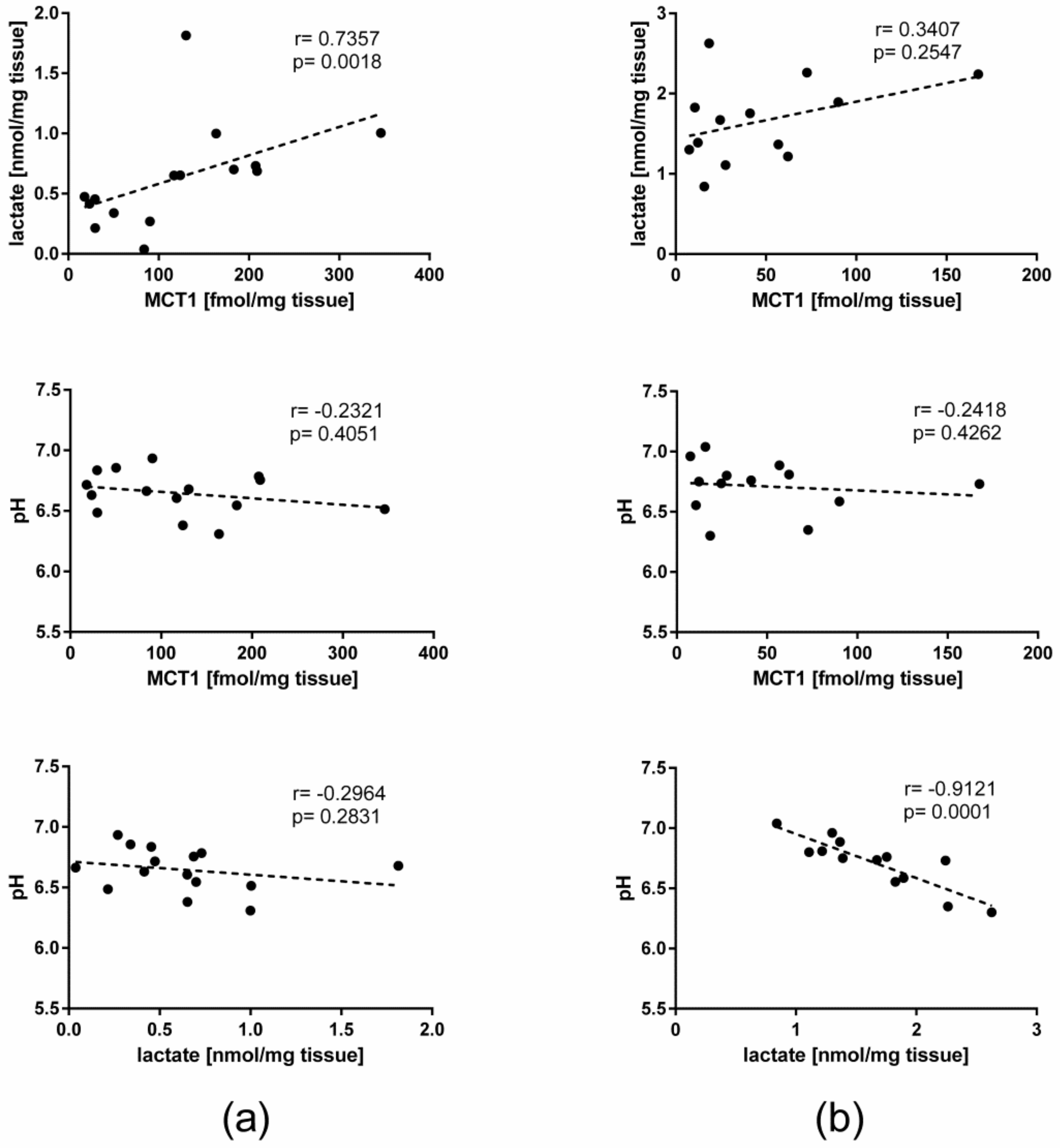
2.5. Lactate Concentration, pH, and Their Correlation with MCT1 in the Liver
2.6. Immunohistochemistry
3. Discussion
4. Materials and Methods
4.1. Liver Samples
4.2. mRNA Quantification
4.3. Protein Quantification by LC−MS/MS
4.4. miRNA Quantification and Analysis
4.5. Measurement of L-Lactate Content in Tissue Samples
4.6. pH Measurement
4.7. Immunohistochemistry
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MCT1 | monocarboxylate transporter 1 |
| SLC16A1 | solute carrier family 16 member 1 |
| ALD | alcoholic liver disease |
| qRT-PCR | quantitative reverse transcription–polymerase chain reaction |
| LC−MS/MS | liquid chromatography-tandem mass spectrometry |
References
- Philp, N.J.; Wang, D.; Yoon, H.; Hjelmeland, L.M. Polarized expression of monocarboxylate transporters in human retinal pigment epithelium and ARPE-19 cells. Invest. Ophthalmol. Vis. Sci. 2003, 44, 1716–1721. [Google Scholar] [CrossRef]
- Halestrap, A.P.; Wilson, M.C. The monocarboxylate transporter family—Role and regulation. IUBMB Life 2012, 64, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Fisel, P.; Schaeffeler, E.; Schwab, M. Clinical and Functional Relevance of the Monocarboxylate Transporter Family in Disease Pathophysiology and Drug Therapy. Clin. Transl. Sci. 2018, 11, 352–364. [Google Scholar] [CrossRef] [PubMed]
- Drozdzik, M.; Busch, D.; Lapczuk, J.; Muller, J.; Ostrowski, M.; Kurzawski, M.; Oswald, S. Protein Abundance of Clinically Relevant Drug Transporters in the Human Liver and Intestine: A Comparative Analysis in Paired Tissue Specimens. Clin. Pharmacol. Ther. 2019, 105, 1204–1212. [Google Scholar] [CrossRef] [PubMed]
- Marchiq, I.; Pouyssegur, J. Hypoxia, cancer metabolism and the therapeutic benefit of targeting lactate/H(+) symporters. J. Mol. Med. (Berl.) 2016, 94, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Pellerin, L.; Magistretti, P.J. Sweet sixteen for ANLS. J. Cereb. Blood Flow Metab. 2012, 32, 1152–1166. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.L.; Jacobs, S.B.; Moreno-Macias, H.; Huerta-Chagoya, A.; Churchhouse, C.; Marquez-Luna, C.; Garcia-Ortiz, H.; Gomez-Vazquez, M.J.; Burtt, N.P.; Aguilar-Salinas, C.A.; et al. Sequence variants in SLC16A11 are a common risk factor for type 2 diabetes in Mexico. Nature 2014, 506, 97–101. [Google Scholar] [CrossRef]
- Kurzawski, M.; Szelag-Pieniek, S.; Lapczuk-Romanska, J.; Wrzesinski, M.; Sienko, J.; Oswald, S.; Drozdzik, M. The reference liver—ABC and SLC drug transporters in healthy donor and metastatic livers. Pharmacol. Rep. 2019, 71, 738–745. [Google Scholar] [CrossRef]
- Halestrap, A.P.; Meredith, D. The SLC16 gene family-from monocarboxylate transporters (MCTs) to aromatic amino acid transporters and beyond. Pflugers Arch. 2004, 447, 619–628. [Google Scholar] [CrossRef]
- Halestrap, A.P. The monocarboxylate transporter family—Structure and functional characterization. IUBMB Life 2012, 64, 1–9. [Google Scholar] [CrossRef]
- Schofield, Z.; Reed, M.A.; Newsome, P.N.; Adams, D.H.; Gunther, U.L.; Lalor, P.F. Changes in human hepatic metabolism in steatosis and cirrhosis. World J. Gastroenterol. 2017, 23, 2685–2695. [Google Scholar] [CrossRef] [PubMed]
- Strazzabosco, M.; Boyer, J.L. Regulation of intracellular pH in the hepatocyte. Mechanisms and physiological implications. J. Hepatol. 1996, 24, 631–644. [Google Scholar] [CrossRef]
- Hou, L.; Zhao, Y.; Song, G.Q.; Ma, Y.H.; Jin, X.H.; Jin, S.L.; Fang, Y.H.; Chen, Y.C. Interfering cellular lactate homeostasis overcomes Taxol resistance of breast cancer cells through the microRNA-124-mediated lactate transporter (MCT1) inhibition. Cancer Cell Int. 2019, 19, 193. [Google Scholar] [CrossRef]
- Morse, B.L.; Felmlee, M.A.; Morris, M.E. γ-Hydroxybutyrate blood/plasma partitioning: Effect of physiologic pH on transport by monocarboxylate transporters. Drug Metab. Dispos. 2012, 40, 64–69. [Google Scholar] [CrossRef]
- Pullen, T.J.; da Silva Xavier, G.; Kelsey, G.; Rutter, G.A. miR-29a and miR-29b contribute to pancreatic beta-cell-specific silencing of monocarboxylate transporter 1 (Mct1). Mol. Cell Biol. 2011, 31, 3182–3194. [Google Scholar] [CrossRef] [PubMed]
- Romero-Cordoba, S.L.; Rodriguez-Cuevas, S.; Bautista-Pina, V.; Maffuz-Aziz, A.; D’Ippolito, E.; Cosentino, G.; Baroni, S.; Iorio, M.V.; Hidalgo-Miranda, A. Loss of function of miR-342-3p results in MCT1 over-expression and contributes to oncogenic metabolic reprogramming in triple negative breast cancer. Sci. Rep. 2018, 8, 12252. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cao, Z.; Yang, G.; You, L.; Zhang, T.; Zhao, Y. MicroRNA-27a (miR-27a) in Solid Tumors: A Review Based on Mechanisms and Clinical Observations. Front. Oncol. 2019, 9, 893. [Google Scholar] [CrossRef]
- Bruckmueller, H.; Martin, P.; Kahler, M.; Haenisch, S.; Ostrowski, M.; Drozdzik, M.; Siegmund, W.; Cascorbi, I.; Oswald, S. Clinically Relevant Multidrug Transporters Are Regulated by microRNAs along the Human Intestine. Mol. Pharm. 2017, 14, 2245–2253. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, C.; Cheng, J.; Chen, B.; Ke, Q.; Lv, Z.; Wu, J.; Zhou, Y. MicroRNA-145 suppresses hepatocellular carcinoma by targeting IRS1 and its downstream Akt signaling. Biochem. Biophys Res. Commun. 2014, 446, 1255–1260. [Google Scholar] [CrossRef]
- Chai, J.; Cai, S.Y.; Liu, X.; Lian, W.; Chen, S.; Zhang, L.; Feng, X.; Cheng, Y.; He, X.; He, Y.; et al. Canalicular membrane MRP2/ABCC2 internalization is determined by Ezrin Thr567 phosphorylation in human obstructive cholestasis. J. Hepatol. 2015, 63, 1440–1448. [Google Scholar] [CrossRef]
- Wilson, M.C.; Meredith, D.; Fox, J.E.; Manoharan, C.; Davies, A.J.; Halestrap, A.P. Basigin (CD147) is the target for organomercurial inhibition of monocarboxylate transporter isoforms 1 and 4: The ancillary protein for the insensitive MCT2 is EMBIGIN (gp70). J. Biol. Chem. 2005, 280, 27213–27221. [Google Scholar] [CrossRef] [PubMed]
- Polanski, R.; Hodgkinson, C.L.; Fusi, A.; Nonaka, D.; Priest, L.; Kelly, P.; Trapani, F.; Bishop, P.W.; White, A.; Critchlow, S.E.; et al. Activity of the monocarboxylate transporter 1 inhibitor AZD3965 in small cell lung cancer. Clin. Cancer Res. 2014, 20, 926–937. [Google Scholar] [CrossRef] [PubMed]
- Mehibel, M.; Ortiz-Martinez, F.; Voelxen, N.; Boyers, A.; Chadwick, A.; Telfer, B.A.; Mueller-Klieser, W.; West, C.M.; Critchlow, S.E.; Williams, K.J.; et al. Statin-induced metabolic reprogramming in head and neck cancer: A biomarker for targeting monocarboxylate transporters. Sci. Rep. 2018, 8, 16804. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Groer, C.; Bruck, S.; Lai, Y.; Paulick, A.; Busemann, A.; Heidecke, C.D.; Siegmund, W.; Oswald, S. LC-MS/MS-based quantification of clinically relevant intestinal uptake and efflux transporter proteins. J. Pharm. Biomed Anal. 2013, 85, 253–261. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | CTRL (n = 20) | Liver Pathologies (n = 77) | HCV (n = 21) | PBC (n = 10) | PSC (n = 6) | ALD (n = 20) | AIH (n = 20) | All Samples (n = 97) |
|---|---|---|---|---|---|---|---|---|
| hsa-miR-27a-3p | −0.591 | −0.251 | −0.156 | −0.417 | −0.829 | 0.174 | −0.335 | −0.301 * |
| hsa-miR-27b-3p | −0.107 | 0.003 | 0.339 | 0.150 | −0.200 | −0.039 | 0.102 | 0.092 |
| hsa-miR-29a-3p | −0.128 | −0.095 | 0.042 | −0.533 | −0.543 | 0.223 | −0.137 | 0.020 |
| hsa-miR-29b-3p | −0.152 | −0.031 | −0.027 | −0.100 | −0.486 | 0.167 | 0.135 | −0.163 |
| hsa-miR-29c-3p | −0.099 | −0.184 | −0.129 | −0.733 | −0.543 | 0.062 | −0.480 | −0.186 |
| hsa-miR-145-5p | −0.475 | −0.166 | −0.005 | 0.533 | −0.543 | 0.069 | −0.346 | −0.253 |
| hsa-miR-320a-3p | −0.209 | −0.053 | −0.048 | 0.333 | −0.714 | 0.463 | −0.368 | 0.049 |
| hsa-miR-324-5p | −0.412 | −0.123 | −0.223 | 0.583 | −0.486 | 0.089 | −0.348 | −0.160 |
| hsa-miR-342-3p | 0.015 | −0.269 | −0.252 | 0.250 | −0.314 | −0.323 | −0.280 | −0.310 * |
| hsa-miR-374a-5p | −0.379 | −0.234 | −0.323 | −0.617 | −0.486 | 0.107 | −0.437 | −0.312 * |
| hsa-miR-374b-5p | −0.436 | −0.195 | −0.075 | −0.100 | −0.600 | 0.045 | −0.628 | −0.173 |
| hsa-miR-376a-3p | −0.131 | −0.182 | −0.319 | 0.317 | −0.486 | −0.003 | −0.391 | −0.145 |
| hsa-miR-425-5p | −0.286 | −0.139 | 0.000 | 0.200 | −0.600 | −0.018 | −0.234 | −0.045 |
| hsa-miR-495-3p | 0.083 | −0.219 | −0.229 | 0.350 | −0.257 | −0.077 | −0.503 | −0.193 |
| hsa-miR-590-3p | −0.189 | −0.126 | −0.209 | −0.833 | −0.086 | 0.045 | −0.292 | −0.124 |
| hsa-miR-605-5p | −0.042 | −0.198 | 0.065 | −0.167 | −0.371 | −0.531 | −0.257 | −0.090 |
| Parameter/Disease | Controls n = 20 | HCV n = 21 | PBC n = 10 | PSC n = 6 | ALD n = 20 | AIH n = 20 |
|---|---|---|---|---|---|---|
| Sex (male/female) | 11/9 | 10/11 | 1/9 | 4/2 | 16/4 | 8/12 |
| Age (years) | 63 ± 10 | 52 ± 5 | 59 ± 4 | 43 ± 10 | 51 ± 6 | 47 ± 16 |
| Child–Pugh (A/B/C) | - | 7/10/4 | 2/4/4 | 3/3/0 | 0/8/12 | 6/6/8 |
| Total bilirubin (mg/dl) | 0.59 ± 0.25 | 2.38 ± 1.37 | 6.42 ± 6.72 | 8.14 ± 8.14 | 4.4 ± 4.02 | 3.54 ± 3.53 |
| Albumin (g/dl) | 3.89 ± 0.38 | 3.31 ± 0.45 | 3.13 ± 0.65 | 3.7 ± 0.44 | 3.03 ± 0.50 | 3.29 ± 0.39 |
| PT (s) | 12.7 ± 2.3 | 14.4 ± 2.0 | 12.5 ± 1.2 | 13.2 ± 2.8 | 16.0 ± 2.2 | 14.6 ± 2.5 |
| INR | 1.14 ± 0.21 | 1.39 ± 0.27 | 1.19 ± 0.21 | 1.4 ± 0.52 | 1.47 ± 0.23 | 1.42 ± 0.41 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Droździk, M.; Szeląg-Pieniek, S.; Grzegółkowska, J.; Łapczuk-Romańska, J.; Post, M.; Domagała, P.; Miętkiewski, J.; Oswald, S.; Kurzawski, M. Monocarboxylate Transporter 1 (MCT1) in Liver Pathology. Int. J. Mol. Sci. 2020, 21, 1606. https://doi.org/10.3390/ijms21051606
Droździk M, Szeląg-Pieniek S, Grzegółkowska J, Łapczuk-Romańska J, Post M, Domagała P, Miętkiewski J, Oswald S, Kurzawski M. Monocarboxylate Transporter 1 (MCT1) in Liver Pathology. International Journal of Molecular Sciences. 2020; 21(5):1606. https://doi.org/10.3390/ijms21051606
Chicago/Turabian StyleDroździk, Marek, Sylwia Szeląg-Pieniek, Justyna Grzegółkowska, Joanna Łapczuk-Romańska, Mariola Post, Pawel Domagała, Janusz Miętkiewski, Stefan Oswald, and Mateusz Kurzawski. 2020. "Monocarboxylate Transporter 1 (MCT1) in Liver Pathology" International Journal of Molecular Sciences 21, no. 5: 1606. https://doi.org/10.3390/ijms21051606
APA StyleDroździk, M., Szeląg-Pieniek, S., Grzegółkowska, J., Łapczuk-Romańska, J., Post, M., Domagała, P., Miętkiewski, J., Oswald, S., & Kurzawski, M. (2020). Monocarboxylate Transporter 1 (MCT1) in Liver Pathology. International Journal of Molecular Sciences, 21(5), 1606. https://doi.org/10.3390/ijms21051606