Metabolomic Analysis of Morus Cultivar Root Extracts and Their Ameliorative Effect on Testosterone-Induced Prostate Enlargement in Sprague-Dawley Rats


Abstract
1. Introduction
2. Results
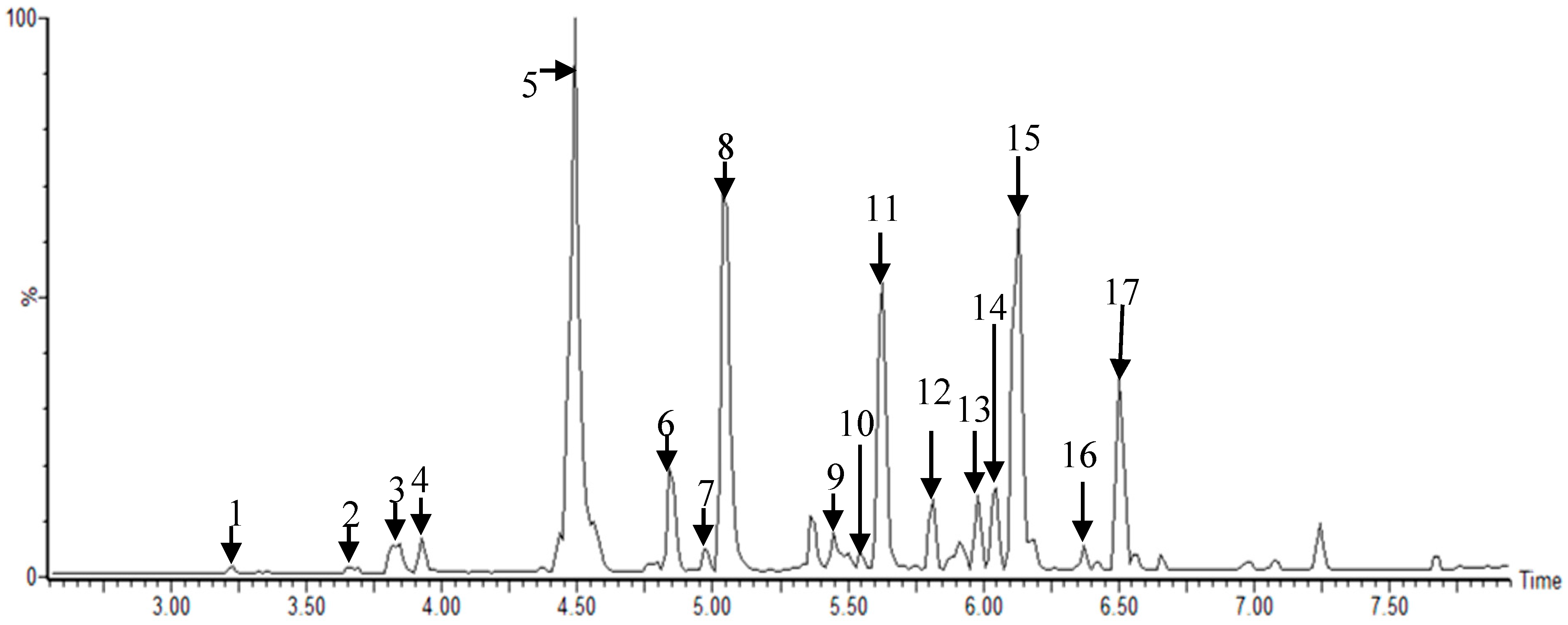
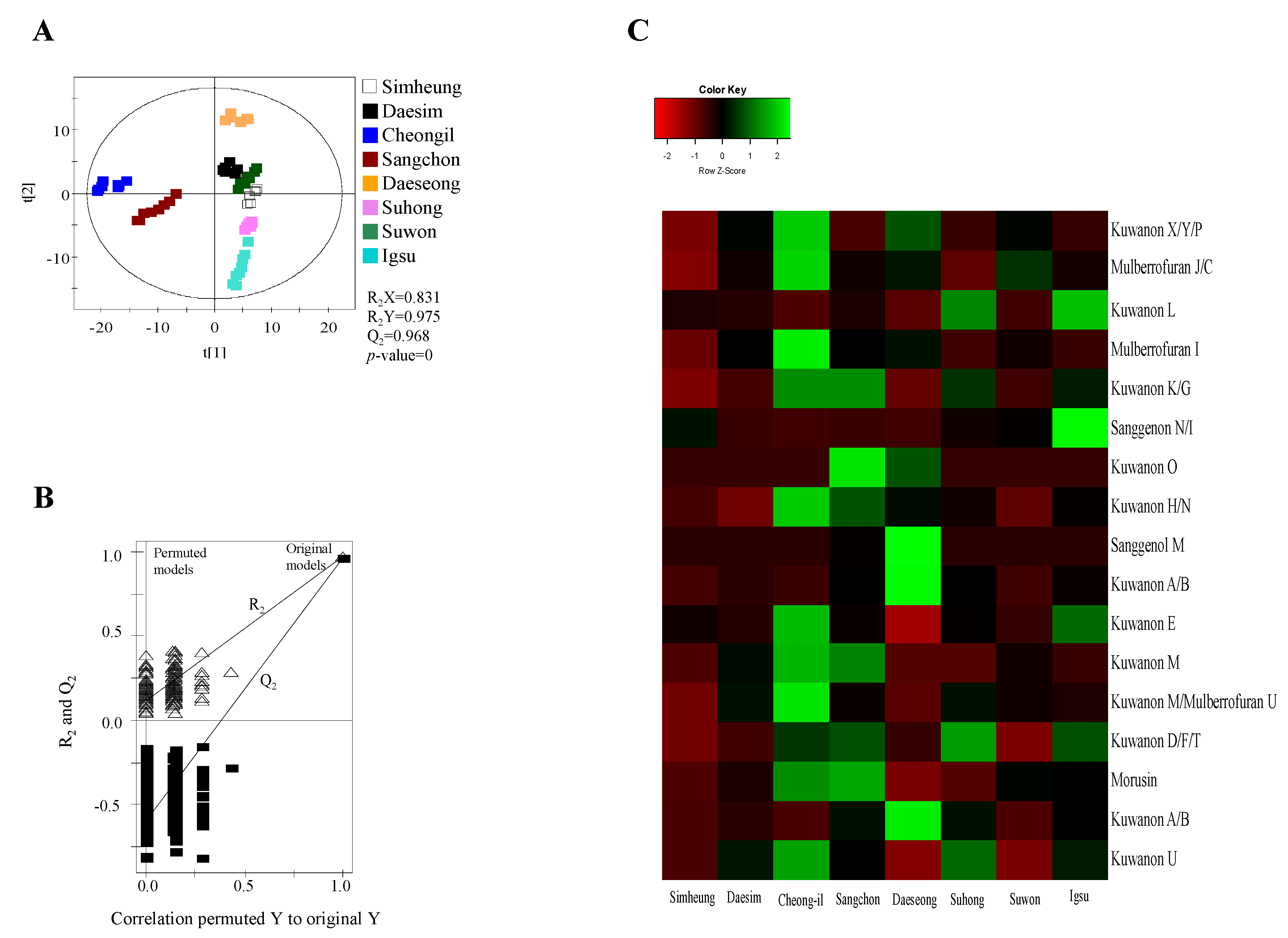
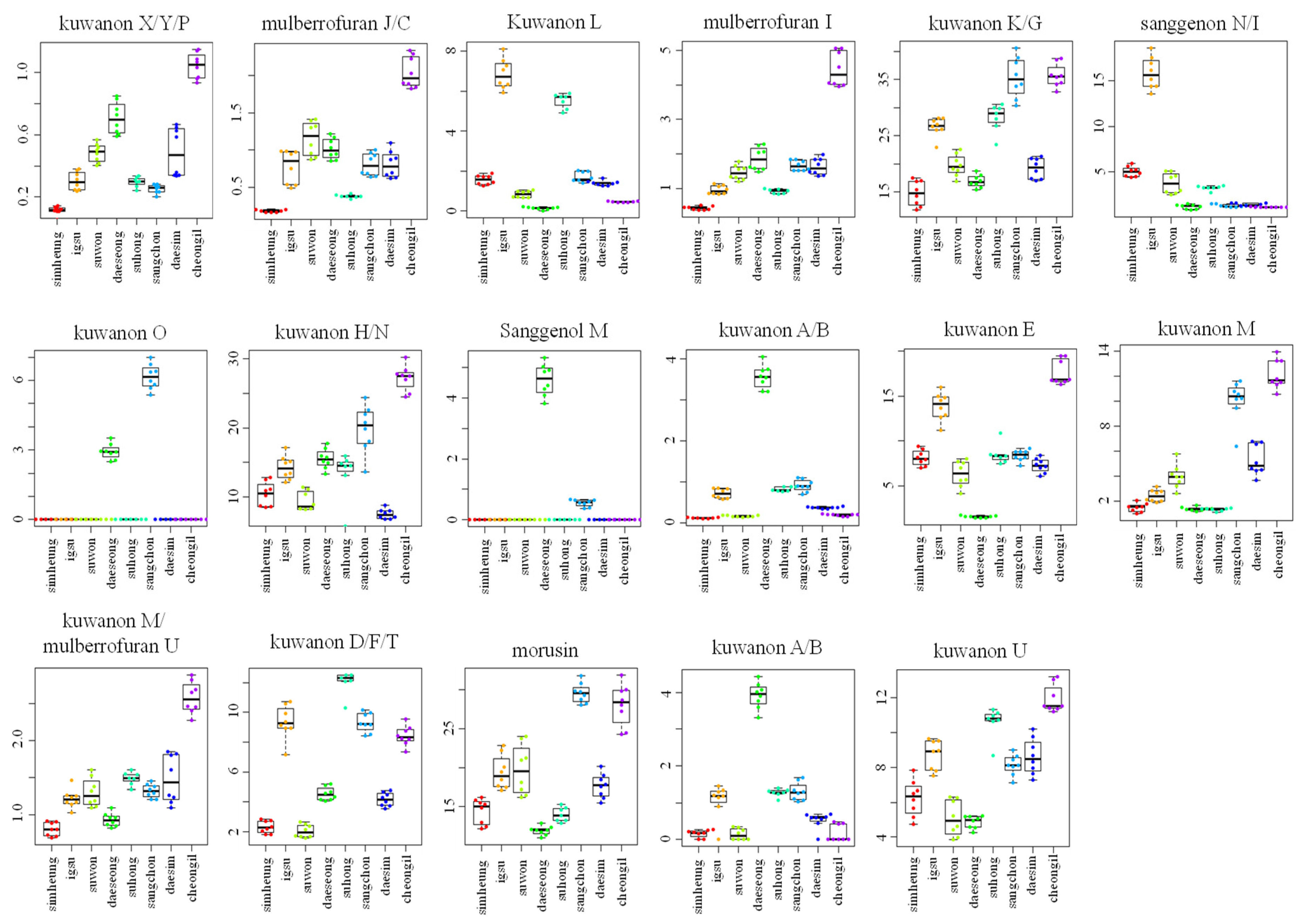
2.1. Metabolomic Analysis of MRs According to Different Cultivars
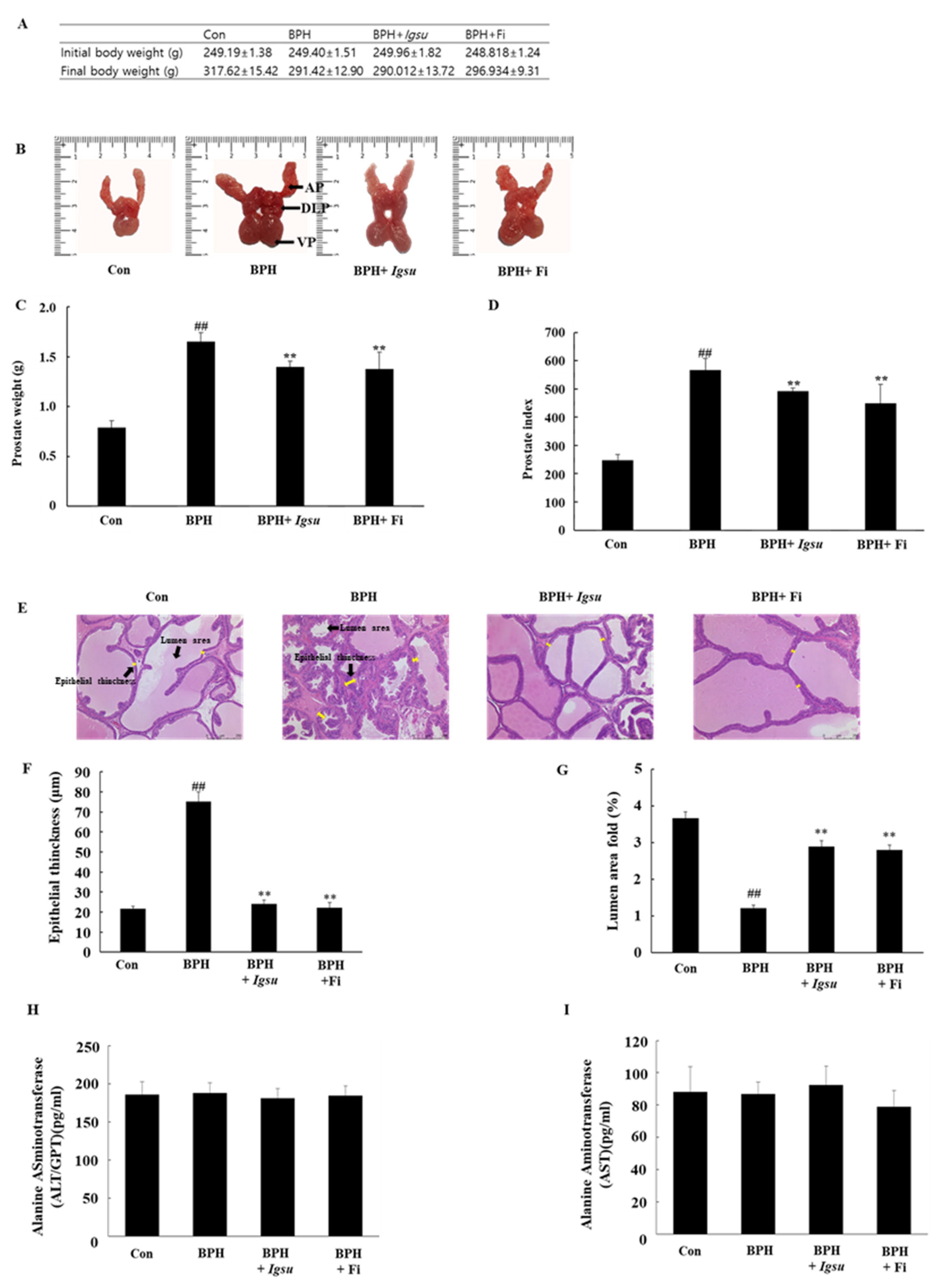
2.2. Effect of Igsu Administration on Prostate Tissue Weight in Rats
2.3. Effects of Igsu Administration on Prostate Histology in Rats
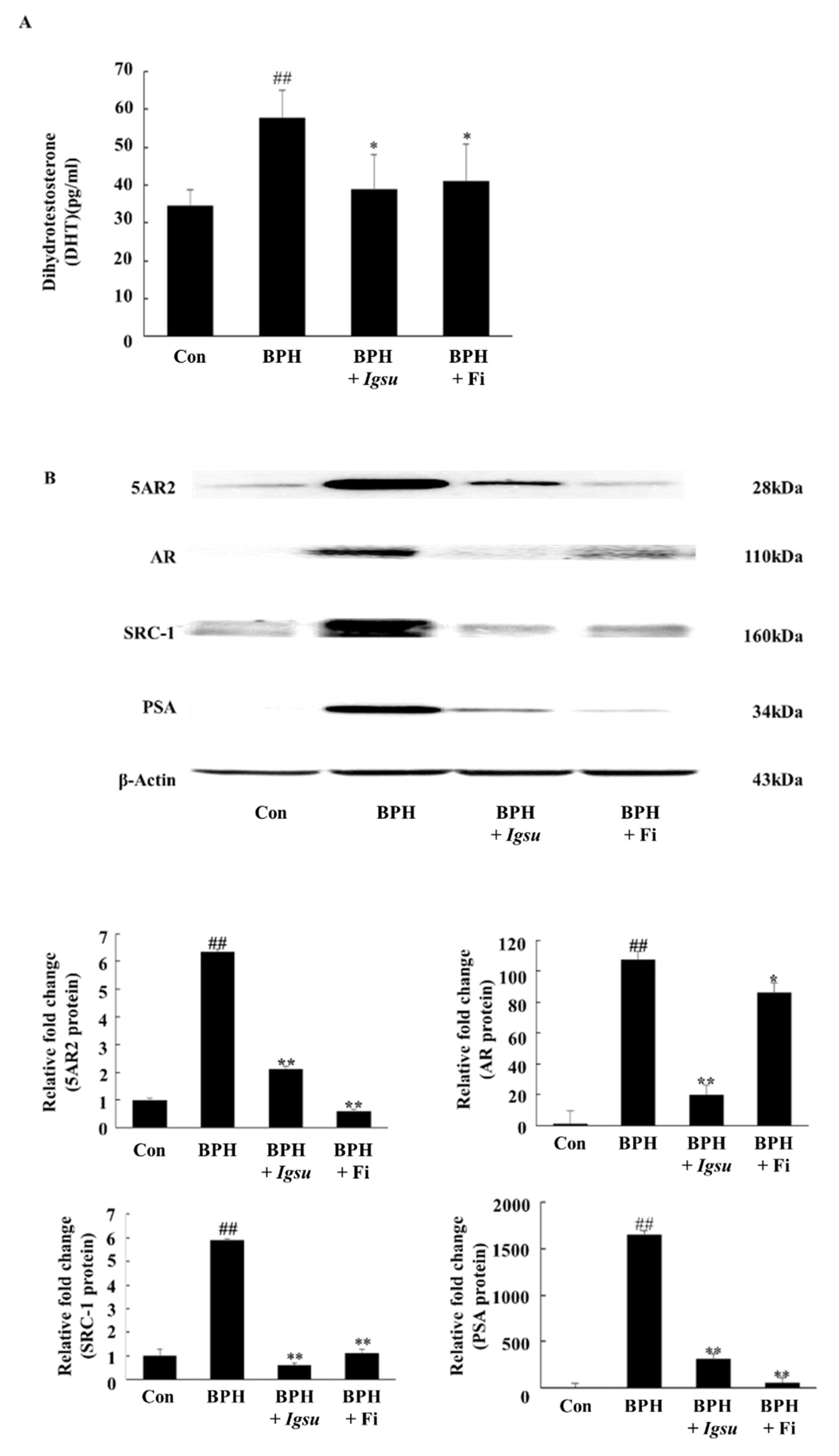
2.4. Effect of Igsu Administration on the Levels of DHT in Prostate Tissue of Rats
2.5. Effect of Igsu on Protein Expressions of 5AR2, AR, SRC-1, and PSA in Prostate Tissues of Rats
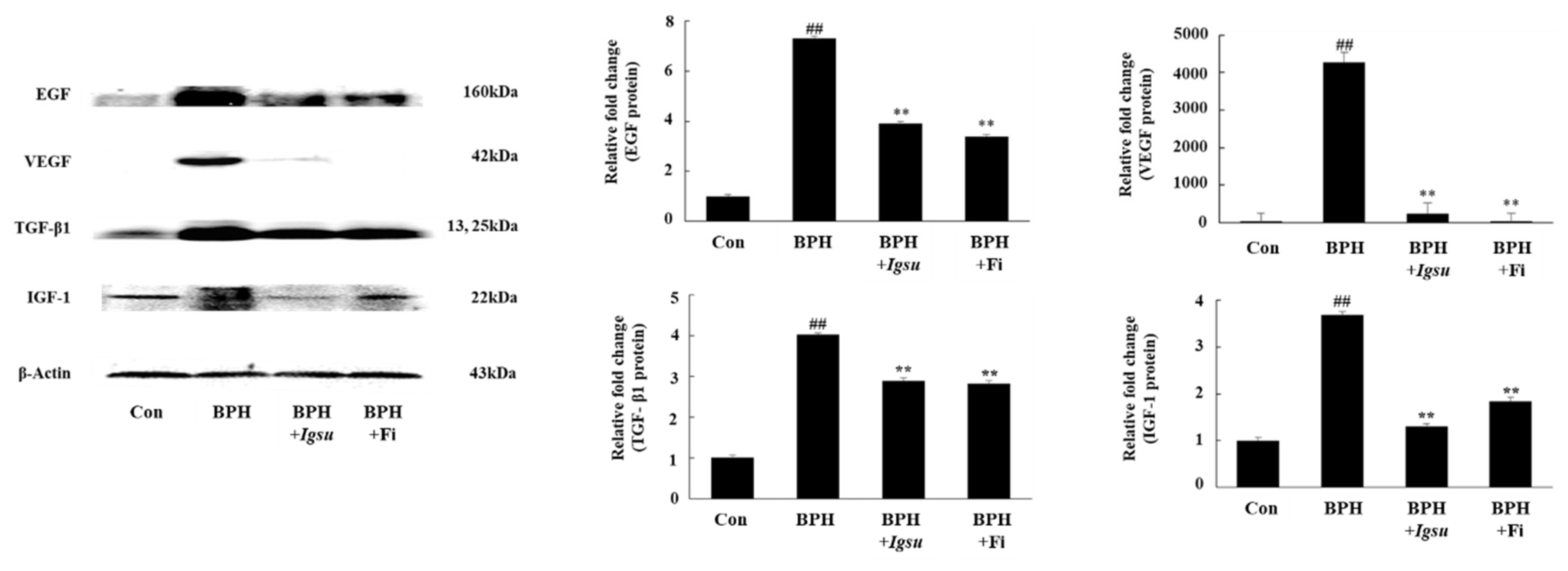
2.6. Effect of Igsu Administration on Expression of Growth Factor-Related Protein in Prostate Tissue of Rats
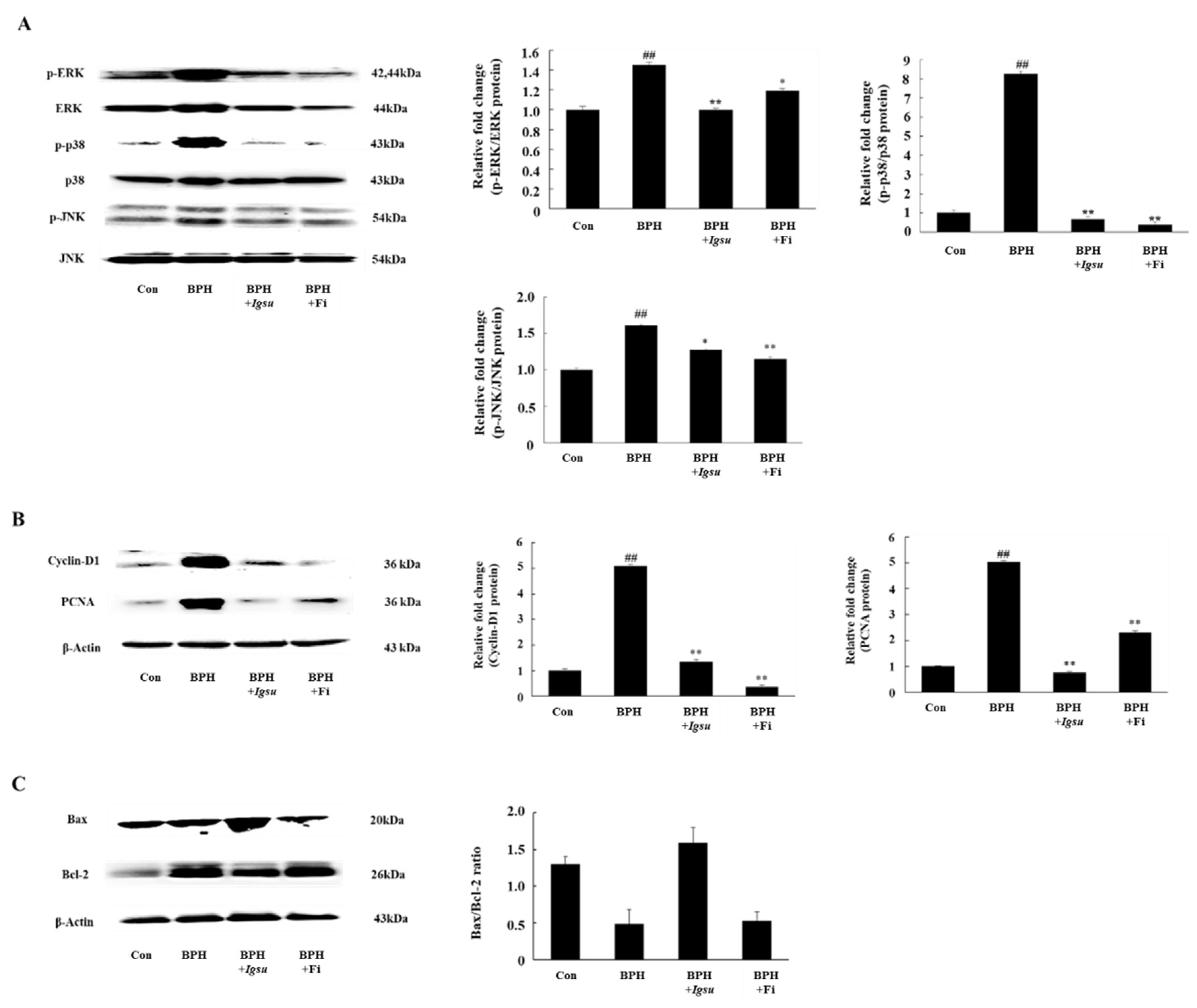
2.7. Effect of Igsu on Phosphorylation of MAPK Signaling in Prostate Tissue of Rats
2.8. Effect of Igsu Administration on Prostate Proliferation-Related Protein Expression in Prostate Tissue of Rats
2.9. Effect of Igsu Administration on Expression of Apoptosis Related Proteins in Prostate Tissue of Rats
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Sample Preparation
4.3. Ultra-Performance Liquid Chromatography–Quadrupole Time-of-Flight (UPLC-QTOF) MS Analysis
4.4. Data Processing
4.5. Treatment of the LNCaP Cells
4.6. Analysis of Messenger RNA (mRNA) Expression
4.7. Animal Experiments for Testosterone-Induced Benign Prostatic Hyperplasia
4.7.1. Animals
4.7.2. Animal Treatments and Experimental Design
4.7.3. Hematoxylin and Eosin (H&E) Staining
4.7.4. Western Blotting Assay
4.7.5. Serum Concentrations of DHT
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MR | Morus root |
| TP | testosterone propionate |
| DHT | dihydrotestosterone |
| Fi | finasteride |
| BPH | benign prostatic hyperplasia |
| 5AR2 | 5-alpha-reductase-type 2 |
| EGF | epidermal growth factor |
| VEGF | vascular endothelial growth factor |
| IGF-1 | insulin-like growth factor-1 |
References
- Kato, T.; Ishibe, T.; Hirayama, M.; Fukushige, M.; Takenaka, I.; Kazuta, M. Basic studies on the prostate of rat under various hormonal environment. Endocrinol. Jpn. 1965, 12, 1–8. [Google Scholar] [CrossRef]
- Roehrborn, C.G. Pathology of benign prostatic hyperplasia. Int. J. Impot. Res. 2008, 20, 11. [Google Scholar] [CrossRef] [PubMed]
- McVary, K.T. BPH: Epidemiology and comorbidities. Am. J. Manag. Care 2006, 12, 122–128. [Google Scholar]
- Barkin, J. Benign prostatic hyperplasia and lower urinary tract symptoms: Evidence and approaches for best case management. Can. J. Urol. 2011, 18, 14–19. [Google Scholar] [PubMed]
- Barry, M.J.; Fowler, F.J.; O’leary, M.P.; Bruskewitz, R.C.; Holtgrewe, H.L.; Mebust, W.K.; Cockett, A.T. The American Urological Association symptom index for benign prostatic hyperplasia. The Measurement Committee of the American Urological Association. J. Urol. 1992, 148, 1549–1557. [Google Scholar] [CrossRef]
- Choi, H.M.; Jung, Y.; Park, J.; Kim, H.L.; Youn, D.H.; Kang, J.; Jeong, M.Y.; Lee, J.H.; Yang, W.M.; Lee, S.G.; et al. Cinnamomi cortex (Cinnamomum verum) suppresses testosterone-induced benign prostatic hyperplasia by regulating 5α-reductase. Sci. Rep. 2016, 6, 31906. [Google Scholar] [CrossRef]
- Patel, N.D.; Parsons, J.K. Epidemiology and etiology of benign prostatic hyperplasia and bladder outlet obstruction. Indian J. Urol. 2014, 30, 170–176. [Google Scholar]
- Papatsoris, A.G.; Papavassiliou, A.G. Molecular ‘palpation’of BPH: A tale of MAPK signalling? Trends Mol. Med. 2001, 7, 288–292. [Google Scholar] [CrossRef]
- Park, E.; Lee, M.Y.; Jeon, W.Y.; Seo, C.S.; You, S.; Shin, H.K. Paljung-San, a traditional herbal medicine, attenuates benign prostatic hyperplasia in vitro and in vivo. J. Ethnopharmacol. 2018, 218, 109–115. [Google Scholar] [CrossRef]
- Gormley, G.J.; Stoner, E.; Bruskewitz, R.C.; Imperato-McGinley, J.; Walsh, P.C.; McConnell, J.D.; Andriole, G.L.; Geller, J.; Bracken, B.R.; Tenover, J.S.; et al. The Finasteride Study Group. The effect of finasteride in men with benign prostatic hyperplasia. N. Engl. J. Med. 1992, 327, 1185–1191. [Google Scholar] [CrossRef]
- Ma, Y.F.; Guo, N.N.; Chu, J.; Jin, S.; Yang, B.; Li, J.; Zhang, T.; Guo, J.T.; Chen, L.; Liang, C.Y.; et al. Glycyrrhizin treatment inhibits proliferation and invasive potential of lung cancer cells. Int. J. Clin. Exp. Med. 2016, 9, 10592–10596. [Google Scholar]
- Awasthi, A.K.; Nagaraja, G.M.; Naik, G.V.; Kanginakudru, S.; Thangavelu, K.; Nagaraju, J. Genetic diversity and relationships in mulberry (genus Morus) as revealed by RAPD and ISSR marker assays. BMC Genet. 2004, 5, 1–9. [Google Scholar] [CrossRef]
- Zerega, N.J.; Clement, W.L.; Datwyler, S.L.; Weiblen, G.D. Biogeography and divergence times in the mulberry family (Moraceae). Mol. Phylogenet. Evol. 2005, 37, 402–416. [Google Scholar] [CrossRef]
- Geng, C.; Yao, S.; Xue, D.; Zuo, A.; Zhang, X.; Jiang, Z.; Ma, Y.; Chen, J. New isoprenylated flavonoid from Morus alba. Zhongguo Zhong Yao Za Zhi 2010, 35, 1560–1565. [Google Scholar] [PubMed]
- Hano, Y.; Suzuki, S.; Nomura, T.; Iitaka, Y. Absolute configuration of natural Diels-Alder type adducts from the Morus root bark. Heterocycles 1988, 27, 2315–2325. [Google Scholar]
- Lee, H.J.; Lyu, D.H.; Koo, U.; Nam, K.W.; Hong, S.S.; Kim, K.O.; Kim, K.H.; Lee, D.; Mar, W. Protection of prenylated flavonoids from Mori Cortex Radicis (Moraceae) against nitric oxide-induced cell death in neuroblastoma SH-SY5Y cells. Arch. Pharm. Res. 2012, 35, 163–170. [Google Scholar] [CrossRef]
- Jee, S.O. Antioxidant activities and whitening effect of the mulberry (Morus alba L.) root bark extracts. Korean J. Plant Res. 2009, 22, 145–151. [Google Scholar]
- Zheng, Z.P.; Tan, H.Y.; Wang, M. Tyrosinase inhibition constituents from the roots of Morus australis. Fitoterapia 2012, 83, 1008–1013. [Google Scholar] [CrossRef]
- Yang, Z.G.; Matsuzaki, K.; Takamatsu, S.; Kitanaka, S. Inhibitory effects of constituents from Morus alba var. multicaulis on differentiation of 3T3-L1 cells and nitric oxide production in RAW264.7 cells. Molecules 2011, 16, 6010–6022. [Google Scholar] [CrossRef]
- Nade, V.S.; Kawale, L.A.; Naik, R.A.; Yadav, A.V. Adaptogenic effect of Morus alba on chronic footshock-induced stress in rats. Indian J. Pharmacol. 2009, 41, 246–251. [Google Scholar] [CrossRef]
- Ye, X.; Han, S.J.; Tsai, S.Y.; DeMayo, F.J.; Xu, J.; Tsai, M.J.; O’Malley, B.W. Roles of steroid receptor coactivator (SRC)-1 and transcriptional intermediary factor (TIF) 2 in androgen receptor activity in mice. Proc. Natl. Acad. Sci. USA 2005, 102, 9487–9492. [Google Scholar] [CrossRef]
- Ho, C.K.; Habib, F.K. Estrogen and androgen signaling in the pathogenesis of BPH. Nat. Rev. Urol. 2011, 8, 29–41. [Google Scholar] [CrossRef] [PubMed]
- McConnell, J.D.; Wilson, J.D.; George, F.W.; Geller, J.; Pappas, F.; Stoner, E. Finasteride, an inhibitor of 5 alpha-reductase, suppresses prostatic dihydrotestosterone in men with benign prostatic hyperplasia. J. Clin. Endocrinol. Metab. 1992, 74, 505–508. [Google Scholar] [PubMed]
- Mondaini, N.; Gontero, P.; Giubilei, G.; Lombardi, G.; Cai, T.; Gavazzi, A.; Bartoletti, R. Finasteride 5 mg and sexual side effects: How many of these are related to a nocebo phenomenon? J. Sex. Med. 2007, 4, 1708–1712. [Google Scholar] [CrossRef]
- Choi, Y.J.; Fan, M.; Tang, Y.; Yang, H.P.; Hwang, J.Y.; Kim, E.K. In Vivo Effects of Polymerized Anthocyanin from Grape Skin on Benign Prostatic Hyperplasia. Nutrients 2019, 11, 2444–2458. [Google Scholar] [CrossRef]
- Joseph, I.B.; Nelson, J.B.; Denmeade, S.R.; Isaacs, J.T. Androgens regulate vascular endothelial growth factor content in normal and malignant prostatic tissue. Clin. Cancer Res. 1997, 3, 2507–2511. [Google Scholar]
- Untergasser, G.; Gander, R.; Lilg, C.; Lepperdinger, G.; Plas, E.; Berger, P. Profiling molecular targets of TGF-β1 in prostate fibroblast-to-myofibroblast. Mech. Ageing Dev. 2005, 126, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Ginty, D.D.; Greenberg, M.E. Coupling of the RAS-MAPK pathway to gene activation by RSK2, a growth factor-regulated CREB kinase. Science 1996, 273, 959–963. [Google Scholar] [CrossRef]
- Kim, E.K.; Choi, E.J. Pathological roles of MAPK signaling pathways in human diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2010, 1802, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Peng, J.; Wu, D.; Han, Z.; Bi, X.; Dai, Q. Ki-67 and PCNA expression in prostate cancer and benign prostatic hyperplasia. Clin. Investig. Med. 2008, 31, 8–15. [Google Scholar] [CrossRef]
- Stacey, D.W. Cyclin D1 serves as a cell cycle regulatory switch in actively proliferating cells. Curr. Opin. Cell Biol. 2003, 15, 158–163. [Google Scholar] [CrossRef]
- Yang, E.; Zha, J.; Jockel, J.; Boise, L.H.; Thompson, C.B.; Korsmeyer, S.J. Bad, a heterodimeric partner for Bcl-XL and Bcl-2, displaces Bax and promotes cell death. Cell 1995, 80, 285–291. [Google Scholar] [CrossRef]
- Adams, J.M.; Cory, S. The Bcl-2 protein family: Arbiters of cell survival. Science 1998, 281, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Fang, T.; Li, M.; Song, Y.; Li, J.; Xue, Z.; Li, J.; Bu, D.; Liu, W.; Zeng, Q.; et al. Pao pereira extract Attenuates testosterone-induced Benign prostatic Hyperplasia in Rats by inhibiting 5α-Reductase. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Ub Wijerathne, C.; Park, H.S.; Jeong, H.Y.; Song, J.W.; Moon, O.S.; Seo, Y.W.; Won, Y.S.; Son, H.Y.; Lim, J.H.; Yeon, S.H.; et al. Quisqualis indica improves benign prostatic hyperplasia by regulating prostate cell proliferation and apoptosis. Biol. Pharm. Bull. 2017, 40, 2015–2133. [Google Scholar] [CrossRef]
- Jeon, W.Y.; Kim, O.S.; Seo, C.S.; Jin, S.E.; Kim, J.A.; Shin, H.K.; Kim, Y.U.; Lee, M.Y. Inhibitory effects of Ponciri Fructus on testosterone-induced benign prostatic hyperplasia in rats. BMC Complement. Altern. Med. 2017, 17, 384–393. [Google Scholar] [CrossRef]
- Jena, A.K.; Vasisht, K.; Sharma, N.; Kaur, R.; Dhingra, M.S.; Karan, M. Amelioration of testosterone induced benign prostatic hyperplasia by Prunus species. J. Ethnopharmacol. 2016, 190, 33–45. [Google Scholar] [CrossRef]
- Ohtake, A.; Ukai, M.; Saitoh, C.; Sonoda, R.; Noguchi, Y.; Okutsu, H.; Yuyama, H.; Sato, S.; Sasamata, M.; Miyata, K. Effect of tamsulosin on spontaneous bladder contraction in conscious rats with bladder outlet obstruction: Comparison with effect on intraurethral pressure. Eur. J. Pharmacol. 2006, 545, 185–191. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | RT (min) a | Identification | Exact Mass (m/z) | Fragment Ions (m/z) | p-Value b | VIP c | |
|---|---|---|---|---|---|---|---|
| Positive [M + H] | 1 | 3.36 | Mulberrofuran G | 563.17 | 453 | 3.56 × 10−38 | 1.28 |
| 2 | 3.78 | Mulberrofuran I | 561.15 | 451 | 1.38 × 10−46 | 1.40 | |
| 3 | 4.31 | Kuwanon G | 693.23 | 137 | 5.01 × 10−22 | 1.12 | |
| 4 | 4.69 | Kuwanon D | 423.18 | 153 | 1.88 × 10−41 | 1.12 | |
| 5 | 4.89 | Kuwanon H | 761.29 | 205 | 8.09 × 10−27 | 1.21 | |
| 6 | 5.23 | Kuwanon F | 423.18 | 153 | 2.79 × 10−28 | 0.60 | |
| 7 | 5.44 | Luteolin-methyl ester-glycoside fragment | 301.07 | 283, 177, 153 | 7.22 × 10−29 | 1.35 | |
| 8 | 5.64 | Kuwanon A | 421.16 | 365, 153 | 8.38 × 10−13 | 0.94 | |
| 9 | 5.88 | Kuwanon T | 423.18 | 153 | 3.59 × 10−31 | 0.76 | |
| 10 | 5.96 | Morusin | 421.16 | 365 | 1.92 × 10−22 | 1.11 | |
| Negative [M − H] | 1 | 3.22 | Kuwanon X | 581.18 | 471, 361, 183, 125 | 1.97 × 10−19 | 0.77 |
| 2 | 3.66 | Mulberrofuran J | 579.16 | 469, 359, 183, 125 | 1.58 × 10−27 | 1.04 | |
| 3 | 3.84 | Kuwanon L | 625.17 | 499, 389, 311, 183, 125 | 1.14 × 10−45 | 1.03 | |
| 4 | 3.92 | Mulberrofuran I | 559.13 | 449, 183, 125 | 1.09 × 10−35 | 0.84 | |
| 5 | 4.50 | Kuwanon K | 691.21 | 581, 539, 419, 379, 353 | 1.12 × 10−29 | 0.90 | |
| 6 | 4.84 | Sanggenon N | 421.00 | 299, 309, 183, 109 | 3.32 × 10−42 | 1.19 | |
| 7 | 4.97 | Kuwanon O | 693.00 | 531, 421, 287, 259, 183, 125 | 8.52 × 10−53 | 1.04 | |
| 8 | 5.05 | Kuwanon H | 759.28 | 581, 539, 379, 353 | 1.43 × 10−24 | 0.82 | |
| 9 | 5.44 | Sanggenol M | 761.00 | 311, 183, 125 | 6.24 × 10−49 | 1.09 | |
| 10 | 5.54 | Kuwanon A | 419.14 | 311, 231, 183, 125 | 1.01 × 10−50 | 1.11 | |
| 11 | 5.67 | Kuwanon E | 423.17 | 297, 125 | 2.88 × 10−34 | 0.91 | |
| 12 | 5.81 | Kuwanon M | 839.31 | 419, 365, 183 | 3.36 × 10−34 | 0.77 | |
| 13 | 5.91 | Mulberrofuran U | 647.22 | 419, 183 | 2.33 × 10−26 | 0.95 | |
| 14 | 6.05 | Kuwanon D | 421.16 | 183, 125 | 1.37 × 10−38 | 1.28 | |
| 15 | 6.12 | Morusin | 419.14 | 297, 217, 191, 173, 109 | 1.84 × 10−27 | 0.85 | |
| 16 | 6.37 | Kuwanon B | 419.14 | 311, 231, 183, 125 | 1.25 × 10−35 | 1.12 | |
| 17 | 6.50 | Kuwanon U | 437.19 | 325, 183, 125 | 5.50 × 10−25 | 1.20 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, Y.-J.; Lee, J.I.; Fan, M.; Tang, Y.; Yoon, E.-J.; Ryu, Y.B.; Kim, E.-K. Metabolomic Analysis of Morus Cultivar Root Extracts and Their Ameliorative Effect on Testosterone-Induced Prostate Enlargement in Sprague-Dawley Rats. Int. J. Mol. Sci. 2020, 21, 1435. https://doi.org/10.3390/ijms21041435
Choi Y-J, Lee JI, Fan M, Tang Y, Yoon E-J, Ryu YB, Kim E-K. Metabolomic Analysis of Morus Cultivar Root Extracts and Their Ameliorative Effect on Testosterone-Induced Prostate Enlargement in Sprague-Dawley Rats. International Journal of Molecular Sciences. 2020; 21(4):1435. https://doi.org/10.3390/ijms21041435
Chicago/Turabian StyleChoi, Young-Jin, Jae In Lee, Meiqi Fan, Yujiao Tang, Eun-Jung Yoon, Young Bae Ryu, and Eun-Kyung Kim. 2020. "Metabolomic Analysis of Morus Cultivar Root Extracts and Their Ameliorative Effect on Testosterone-Induced Prostate Enlargement in Sprague-Dawley Rats" International Journal of Molecular Sciences 21, no. 4: 1435. https://doi.org/10.3390/ijms21041435
APA StyleChoi, Y.-J., Lee, J. I., Fan, M., Tang, Y., Yoon, E.-J., Ryu, Y. B., & Kim, E.-K. (2020). Metabolomic Analysis of Morus Cultivar Root Extracts and Their Ameliorative Effect on Testosterone-Induced Prostate Enlargement in Sprague-Dawley Rats. International Journal of Molecular Sciences, 21(4), 1435. https://doi.org/10.3390/ijms21041435