Combined Effects of Diatom-Derived Oxylipins on the Sea Urchin Paracentrotus lividus





 and
and 
Abstract
1. Introduction
2. Results
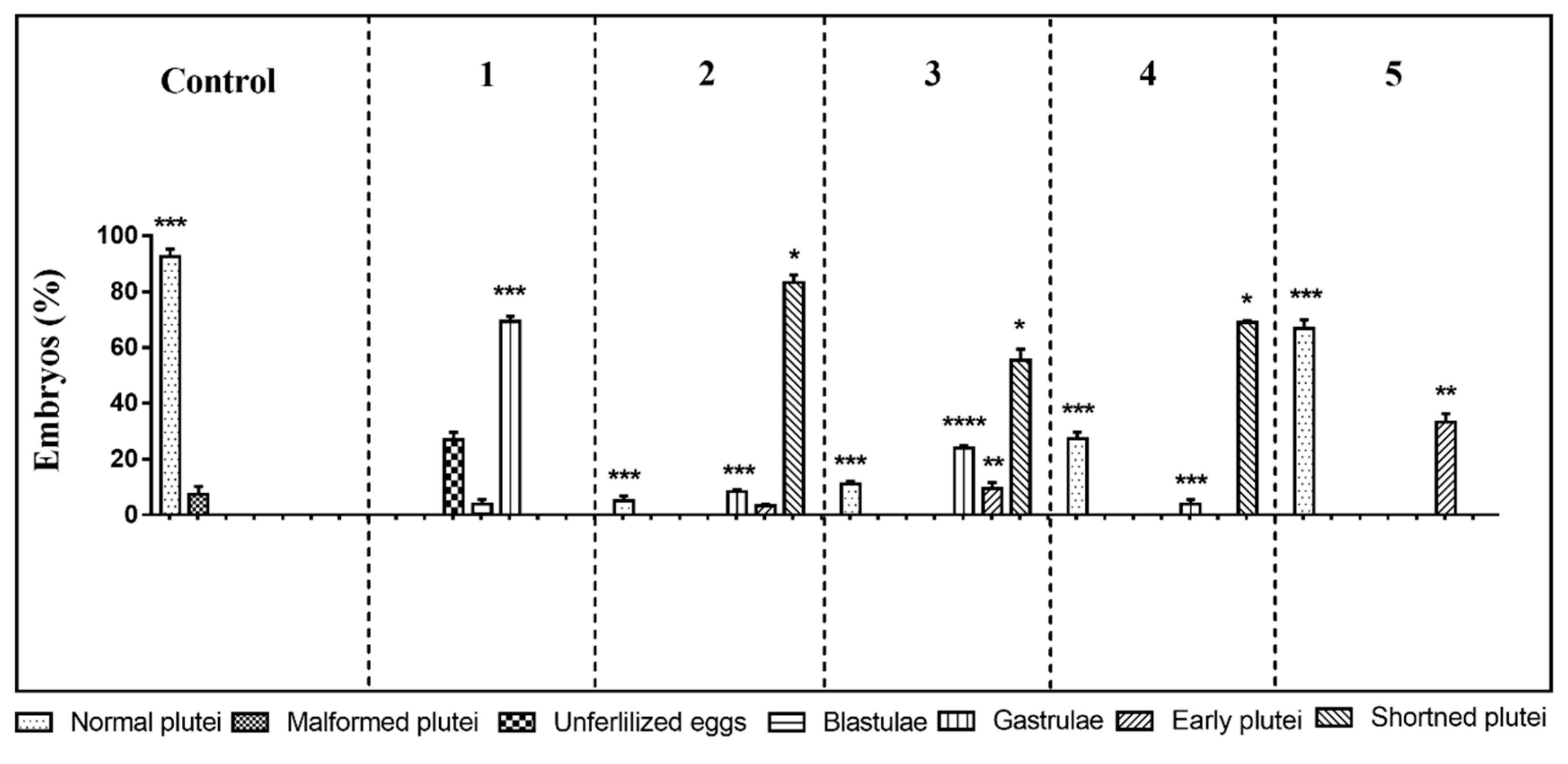
2.1. Morphological Effects of PUA/HEPE Mixtures
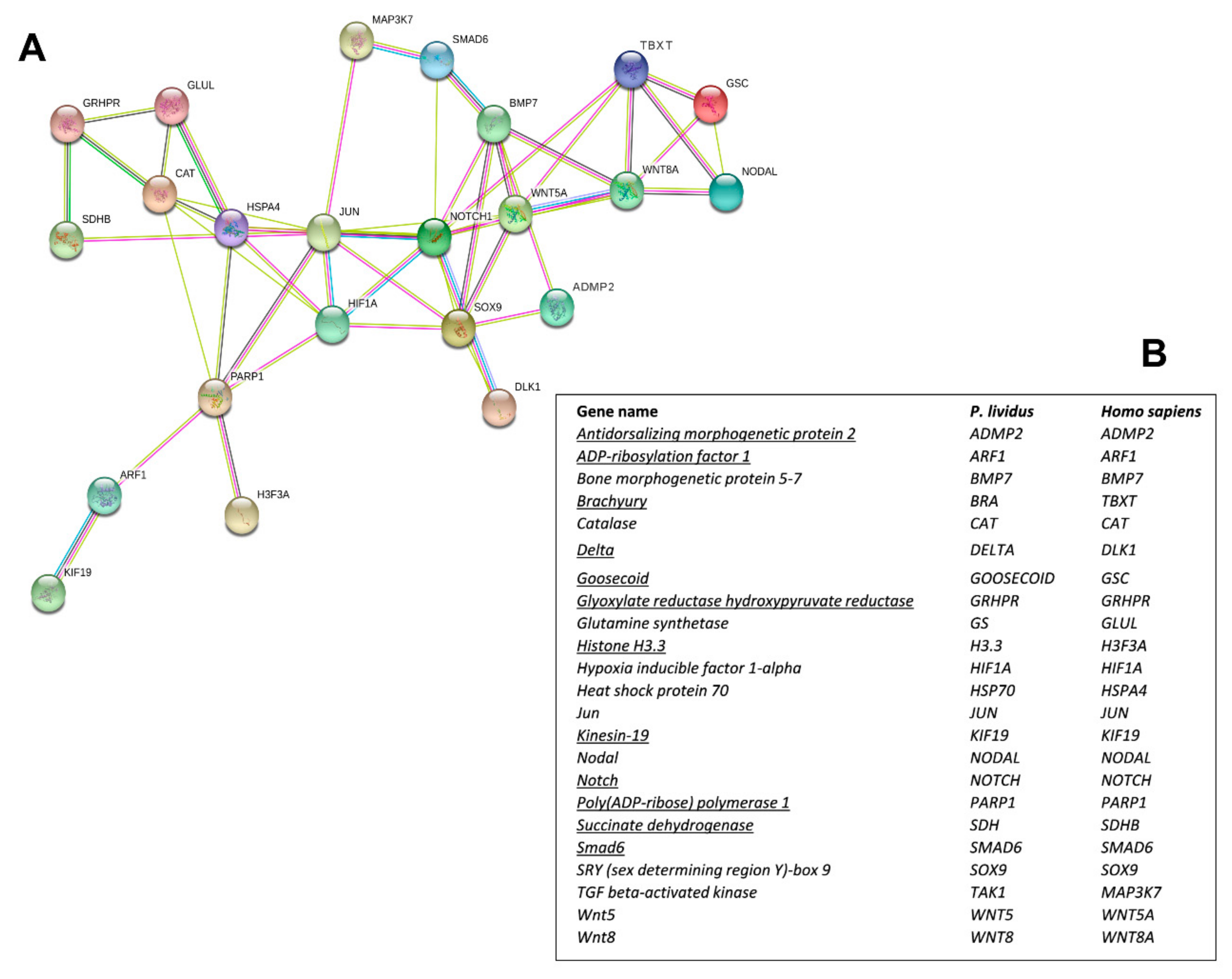
2.2. Network Analysis
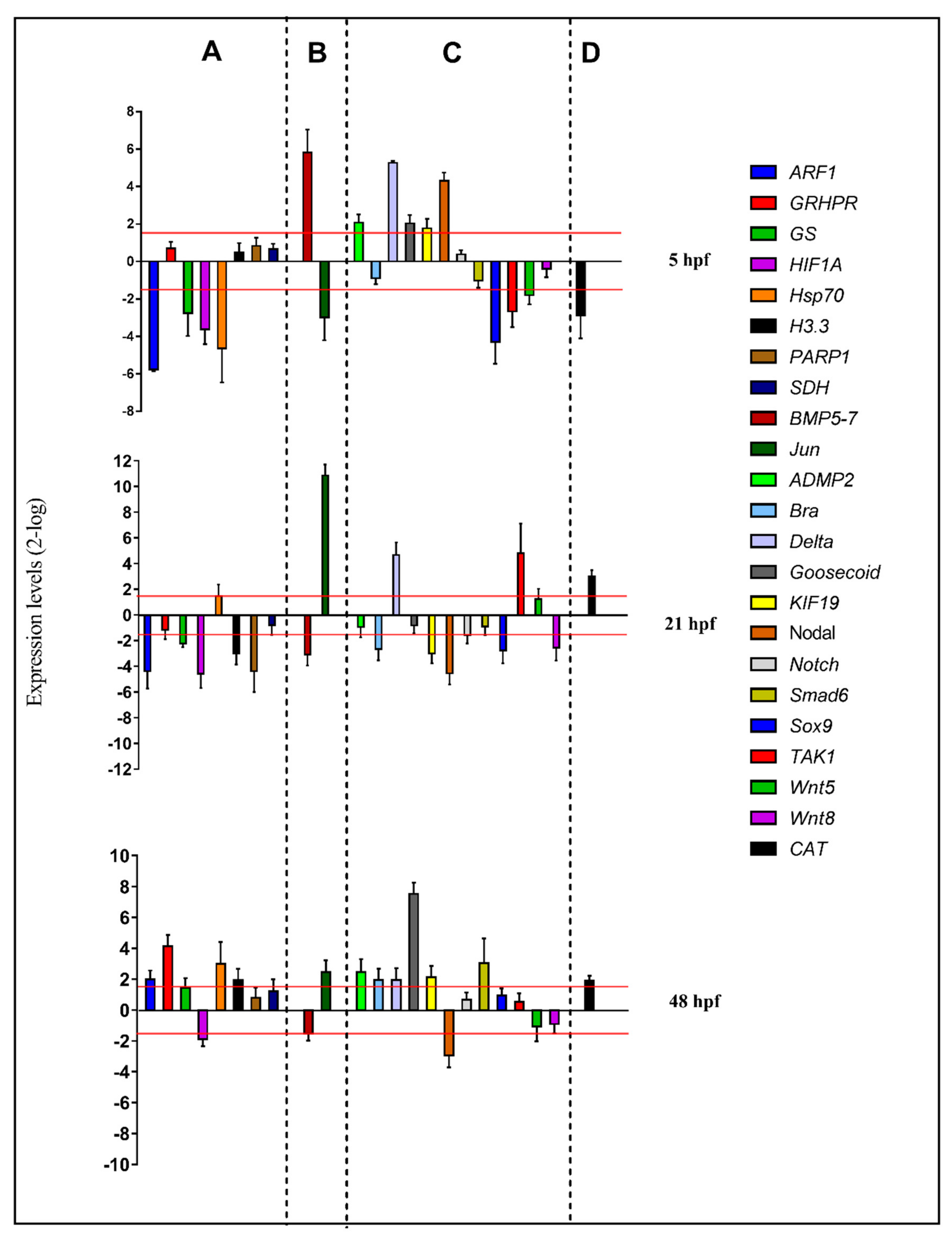
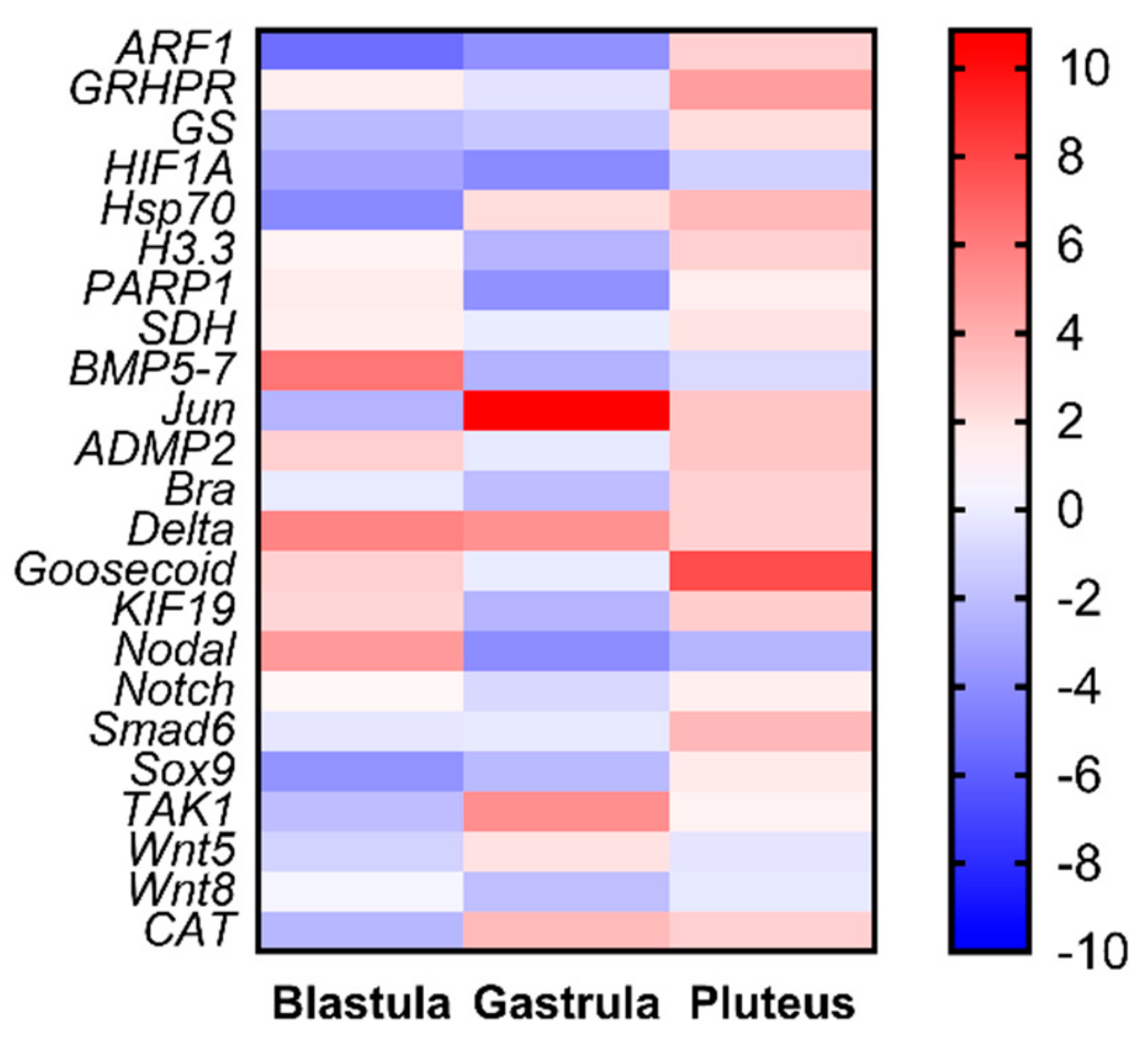
2.3. Molecular Analyses by Real-Time qPCR
2.3.1. Stress Response Genes
2.3.2. Skeletogenic Genes
2.3.3. Development/Differentiation Genes
2.3.4. Detoxification Genes
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Gamete Collection, Egg Incubation with Mixtures of PUAs and HEPEs, Embryo Cultures, and Morphological Analyses
4.3. RNA Extraction and cDNA Synthesis
4.4. Gene Isolation and Interactomic Analysis
4.5. Real-Time qPCR
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Singh, B.R.; Steinnes, E. Soil and water contamination by heavy metals. In Soil Processes and Water Quality; Lal, R., Stewart, B.A., Eds.; Advances in Soil Science; Lewis Publishers: Boca Raton, FL, USA, 1994; pp. 233–271. [Google Scholar]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melill, J.M. Human Domination of Earth’s Ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef]
- Cunningham, V.L.; Buzby, M.; Hutchinson, T.; Mastrocco, F.; Parke, N.; Roden, N. Effects of human pharmaceuticals on aquatic life: Next steps. Environ. Sci. Technol. 2006, 40, 3456–3562. [Google Scholar] [CrossRef] [PubMed]
- Lein, N.P.H.; Fujii, S.; Tanaka, S.; Nozoe, M.; Tanaka, H. Contamination of perfluorooctane sulfonate (PFOS) and perfluorooctanoate (PFOA) in surface water of the Yodo River basin (Japan). Desalination 2008, 226, 338–347. [Google Scholar] [CrossRef]
- Orr, J.C.; Fabry, V.J.; Aumont, O.; Bopp, L.; Doney, S.C.; Feely, R.A.; Gnanadesikan, A.; Gruber, N.; Ishida, A.; Joos, F.; et al. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 2005, 437, 681–686. [Google Scholar] [CrossRef]
- Guinotte, J.M.; Fabry, V.J. Ocean acidification and its potential effects on marine ecosystems. Ann. N. Y. Acad. Sci. 2008, 1134, 320–342. [Google Scholar] [CrossRef] [PubMed]
- Fabry, V.J.; Seibel, B.A.; Feely, R.A.; Orr, J.C. Impacts of ocean acidification on marine fauna and ecosystem processes. ICES J. Mar. Sci. 2008, 65, 414–432. [Google Scholar] [CrossRef]
- Kroeker, K.J.; Kordas, R.L.; Crim, R.; Hendriks, I.E.; Ramajo, L.; Singh, G.S.; Duarte, C.M.; Gattuso, J.P. Impacts of ocean acidification on marine organisms: Quantifying sensitivities and interaction with warming. Glob. Chang. Biol. 2013, 19, 1884–1896. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Bruno, J.F. The Impact of Climate Change on the World’s Marine Ecosystems. Science 2010, 328, 1523–1528. [Google Scholar] [CrossRef]
- Miralto, A.; Barone, G.; Romano, G.; Poulet, S.A.; Ianora, A.; Russo, G.L.; Buttino, I.; Mazzarella, G.; Laabir, M.; Cabrini, M.; et al. The insidious effect of diatoms on copepod reproduction. Nature 1999, 402, 173–176. [Google Scholar] [CrossRef]
- Fontana, A.; d’Ippolito, G.; Cutignano, A.; Miralto, A.; Ianora, A.; Romano, G.; Cimino, G. Chemistry of oxylipin pathways in marine diatoms. Pure Appl. Chem. 2007, 79, 481–490. [Google Scholar] [CrossRef]
- Fontana, A.; d’Ippolito, G.; Cutignano, A.; Romano, G.; Lamari, N.; Massa Gallucci, A.; Cimino, G.; Miralto, A.; Ianora, A. LOX-induced lipid peroxidation mechanism responsible for the detrimental effect of marine diatoms on zooplankton grazers. Chembiochem 2007, 8, 1810–1818. [Google Scholar] [CrossRef] [PubMed]
- d’Ippolito, G.; Lamari, N.; Montresor, M.; Romano, G.; Cutignano, A.; Gerecht, A.; Cimino, G.; Fontana, A. 15S-lipoxygenase metabolism in the marine diatom Pseudo-nitzschia delicatissima. New Phytol. 2009, 183, 1064–1071. [Google Scholar] [CrossRef] [PubMed]
- Cutignano, A.; Lamari, N.; D’ippolito, G.; Manzo, E.; Cimino, G.; Fontana, A. Lipoxygenase products in marine diatoms: A concise analytical method to explore the functional potential of oxylipins. J. Physiol. 2011, 47, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, Y.; Wang, Y.; Wang, Y. Assessment of toxic interactions of heavy metals in multi-component mixtures using sea urchin embryo-larval bioassay. Toxicol. Vitr. 2011, 25, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Bonaventura, P.; Benedetti, G.; Albarède, F.; Miossec, P. Zinc and its role in immunity and inflammation. Autoimm. Rev. 2015, 14, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Ragusa, M.A.; Costa, S.; Cuttitta, A.; Gianguzza, F.; Nicosia, A. Coexposure to sulfamethoxazole and cadmium impairs development and attenuates transcriptional response in sea urchin embryo. Chemosphere 2017, 180, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Padilla-Gamiño, J.L.; Kelly, M.W.; Evans, T.G.; Hofmann, G.E. Temperature and CO2 additively regulate physiology, morphology and genomic responses of larval sea urchins, Strongylocentrotus purpuratus. Proc. R. Soc. B 2013, 280, 20130155. [Google Scholar] [CrossRef]
- Dorey, N.; Maboloc, E.; Chana, K.Y.K. Development of the sea urchin Heliocidaris crassispina from Hong Kong is robust to ocean acidification and copper contamination. Aquat. Toxicol. 2018, 205, 1–10. [Google Scholar] [CrossRef]
- Mak, K.K.Y.; Chan, K.K.Y. Interactive effects of temperature and salinity on early life stages of the sea urchin Heliocidaris crassispina. Mar. Biol. 2018, 165, 57. [Google Scholar] [CrossRef]
- Feehan, C.J.; Grauman, B.C.; Strathmann, R.R.; Dethier, M.N.; Duggins, D.O. Kelp detritus provides high quality food for sea urchin larvae. Limnol. Oceanogr. 2018, 63, S299–S306. [Google Scholar] [CrossRef]
- Glockner-Fagetti, A.; Phillips, N.E. Low salinity and sediment stress on sea urchin Evechinus chloroticus larvae has latent effects on juvenile performance. Mar. Ecol. Prog. Ser. 2019, 619, 85–96. [Google Scholar] [CrossRef]
- Lenz, B.; Fogartya, N.D.; Figueiredo, J. Effects of ocean warming and acidification on fertilization success and early larval development in the green sea urchin Lytechinus variegatus. Mar. Pollut. Bull. 2019, 141, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, G.S. The influence of bioactive oxylipins from marine diatoms on invertebrate reproduction and development. Mar. Drugs 2009, 7, 367–400. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.A.; Bentley, M.G.; Cadwell, G.S. 2,4-Decadienal: Exploring a novel approach for the control of polychaete pests on cultured abalone. Aquaculture 2010, 310, 52–60. [Google Scholar] [CrossRef]
- Lettieri, A.; Esposito, R.; Ianora, A.; Spagnuolo, A. Ciona intestinalis as a marine model system to study some key developmental genes targeted by the diatom-derived aldehyde decadienal. Mar. Drugs 2015, 13, 1451–1465. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Romano, G.; Roncalli, V.; Amoresano, A.; Fontanarosa, C.; Bastianini, M.; Braga, F.; Carotenuto, Y.; Ianora, A. New oxylipins produced at the end of a diatom bloom and their effects on copepod reproductive success and gene expression levels. Harmful Algae 2016, 55, 221–229. [Google Scholar] [CrossRef]
- Romano, G.; Miralto, A.; Ianora, A. Teratogenic effects of diatom metabolites on sea urchin Paracentrotus lividus embryos. Mar. Drugs 2010, 8, 950–967. [Google Scholar] [CrossRef]
- Romano, G.; Costantini, M.; Buttino, I.; Ianora, A.; Palumbo, A. Nitric oxide mediates the stress response induced by diatom aldehydes in the sea urchin Paracentrotus lividus. PLoS ONE 2011, 6, e25980. [Google Scholar] [CrossRef]
- Varrella, S.; Romano, G.; Ianora, A.; Bentley, M.G.; Ruocco, N.; Costantini, M. Molecular response to toxic diatom-derived aldehydes in the sea urchin Paracentrotus lividus. Mar. Drugs 2014, 12, 2089–2113. [Google Scholar] [CrossRef]
- Varrella, S.; Romano, G.; Ruocco, N.; Ianora, A.; Bentley, M.G.; Costantini, M. First morphological and molecular evidence of the negative impact of diatom-derived hydroxyacids on the sea urchin Paracentrotus lividus. Toxicol. Sci. 2016, 151, 419–433. [Google Scholar] [CrossRef]
- Ruocco, N.; Varrella, S.; Romano, G.; Ianora, A.; Bentley, M.G.; Somma, D.; Leonardi, A.; Mellone, S.; Zuppa, A.; Costantini, M. Diatom-derived oxylipins induce cell death in sea urchin embryos activating caspase-8 and caspase 3/7. Aquat. Toxicol. 2016, 176, 128–140. [Google Scholar] [CrossRef] [PubMed]
- Pohnert, G.; Lumineau, O.; Cueff, A.; Adolph, S.; Cordevant, C.; Lange, M.; Poulet, S. Are volatile unsaturated aldehydes from diatoms the main line of chemical defence against copepods? Mar. Ecol. Prog. Ser. 2002, 245, 33–45. [Google Scholar] [CrossRef]
- Taylor, R.L.; Abrahamsson, K.; Godhe, A.; Wangberg, S.A. Seasonal variability in polyunsaturated aldehyde production potential among strains of Skeletonema marinoi (bacillariophyceae). J. Phycol. 2009, 45, 46–53. [Google Scholar] [CrossRef] [PubMed]
- D’Ippolito, G.; Romano, G.; Caruso, T.; Spinella, A.; Cimino, G.; Fontana, A. Production of Octadienal in the Marine Diatom Skeletonema costatum. Org. Lett. 2003, 6, 885–887. [Google Scholar] [CrossRef]
- d’Ippolito, G.; Tucci, S.; Cutignano, A.; Romano, G.; Cimino, G.; Miralto, A.; Fontana, A. The role of complex lipids in the synthesis of bioactive aldehydes of the marine diatom Skeletonema costatum. Biochim. Biophys. Acta 2004, 1686, 100–107. [Google Scholar] [CrossRef]
- Pohnert, G. Wound-activated chemical defense in unicellular planktonic algae. Angew. Chem. Int. Ed. 2000, 39, 4352–4354. [Google Scholar] [CrossRef]
- Pohnert, G. Phospholipase A(2) activity triggers the wound-activated chemical defense in the diatom Thalassiosira rotula. Plant Physiol. 2002, 129, 103–111. [Google Scholar] [CrossRef]
- d’Ippolito, G.; Iadicicco, I.; Romano, G.; Fontana, A.; Iadicicco, O. Detection of short-chain aldehydes in marine organisms: The diatom Thalassiosira rotula. Tetrahedron Lett. 2002, 43, 6137–6140. [Google Scholar] [CrossRef]
- d’Ippolito, G.; Romano, G.; Iadicicco, O.; Miralto, A.; Ianora, A.; Cimino, G.; Fontana, A. New birth-control aldehydes from the marine diatom Skeletonema costatum: Characterization and biogenesis. Tetrahedron Lett. 2002, 43, 6133–6136. [Google Scholar] [CrossRef]
- Wichard, T.; Poulet, S.A.; Pohnert, G. Determination and quantification of α,β,γ,δ-unsaturated aldehydes as pentafluorobenzyl-oxime derivates in diatom cultures and natural phytoplankton populations: Application in marine field studies. J. Chromatogr. B Biomed. Sci. Appl. 2005, 814, 155–161. [Google Scholar] [CrossRef]
- Wichard, T.; Poulet, S.A.; Halsband-Lenk, C.; Albaina, A.; Harris, R.; Liu, D.; Pohnert, G. Survey of the chemical defense potential of diatoms: Screening of fifty species for α,β,γ,δ-unsaturated aldehydes. J. Chem. Ecol. 2005, 31, 949–958. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, A.; Carotenuto, Y.; Lamari, N.; Esposito, F.; D’Ippolito, G.; Fontana, A.; Romano, G.; Ianora, A.; Miralto, A.; Guisande, C. Diatom induction of reproductive failure in copepods: The effect of PUAs versus non volatile oxylipins. J. Exp. Mar. Biol. Ecol. 2011, 401, 13–19. [Google Scholar] [CrossRef]
- Nanjappa, D.; D’Ippolito, G.; Gallo, C.; Zingone, A.; Fontana, A. Oxylipin diversity in the diatom family Leptocylindraceae reveals DHA derivatives in marine diatoms. Mar. Drugs 2014, 12, 368–384. [Google Scholar] [CrossRef] [PubMed]
- Ianora, A.; Miralto, A. Toxigenic effects of diatoms on grazers, phytoplankton and other microbes: A review. Ecotoxicology 2010, 19, 493–511. [Google Scholar] [CrossRef] [PubMed]
- Albarano, L.; Ruocco, N.; Ianora, A.; Libralato, G.; Manfra, L.; Costantini, M. Molecular and morphological toxicity of diatom-derived hydroxyacid mixtures to sea urchin Paracentrotus lividus embryos. Mar. Drugs 2019, 17. [Google Scholar] [CrossRef] [PubMed]
- Ruocco, N.; Annunziata, C.; Ianora, A.; Libralato, G.; Manfra, L.; Costantini, S.; Costantini, M. Toxicity of diatom-derived polyunsaturated aldehyde mixtures on sea urchin Paracentrotus lividus development. Sci. Rep. 2019, 9, 517. [Google Scholar] [CrossRef]
- Varrella, S.; Romano, G.; Costantini, S.; Ruocco, N.; Ianora, A.; Bentley, M.G.; Costantini, M. Toxic diatom aldehydes affect defence gene networks in sea urchins. PLoS ONE 2016, 11, e0149734. [Google Scholar] [CrossRef]
- Ruocco, N.; Fedele, A.M.; Costantini, S.; Romano, G.; Ianora, A.; Costantini, M. New inter-correlated genes targeted by diatom-derived polyunsaturated aldehydes in the sea urchin Paracentrotus lividus. Ecotox. Environ. Safe 2017, 142, 355–362. [Google Scholar] [CrossRef]
- Marrone, V.; Piscopo, M.; Romano, G.; Ianora, A.; Palumbo, A.; Costantini, M. Defensome against toxic diatom aldehydes in the sea urchin Paracentrotus lividus. PLoS ONE 2012, 7, e31750. [Google Scholar] [CrossRef]
- D’Ippolito, G.; Cutignano, A.; Briante, R.; Febbraio, F.; Cimino, G.; Fontana, A. New C16 fatty-acid-based oxylipin pathway in the marine diatom Thalassiosira rotula. Org. Biomol. Chem. 2005, 3, 4065–4070. [Google Scholar] [CrossRef]
- D’Ippolito, G.; Cutignano, A.; Tucci, S.; Romano, G.; Cimino, G.; Fontana, A. Biosynthetic intermediates and stereochemical aspects of aldehyde biosynthesis in the marine diatom Thalassiosira Rotula. Phytochemistry 2006, 67, 314–322. [Google Scholar] [CrossRef]
- Vanaverbeke, J.; Franco, M.A.; van Oevelen, D.; Moodley, L.; Provoost, P.; Steyaert, M.; Soetaert, K.; Vincx, M. Benthic responses to sedimentation of phytoplankton on the Belgian Continental Shelf. In Current Status of Eutrophication in the Belgian Coastal Zone; Presses Universitaires de Bruxelles: Bruxelles, Belgium, 2006; pp. 73–90. [Google Scholar]
- Lignell, R.; Heiskanen, A.S.; Kuosa, H.; Gundersen, K.; Kuuppo-Leinikki, P.; Pajuniemi, R.; Vitto, A. Fate of a phytoplankton spring bloom: Sedimentation and carbon flow in the planktonic food web in the northern baltic. Mar. Ecol. Prog. Ser. 1993, 94, 239–252. [Google Scholar] [CrossRef]
- Wetzel, R.G. Death, detritus and energy flow in aquatic ecosystems. Freshw. Biol. 1995, 33, 83–89. [Google Scholar] [CrossRef]
- Russo, E.; Ianora, A.; Carotenuto, Y. Re-shaping marine plankton communities: Effects of diatoms oxylipins on copepods and beyond. Mar. Biol. 2019, 116, 9. [Google Scholar] [CrossRef]
- Raniello, R.; Iannicielli, M.M.; Nappo, M.; Avilla, C.; Zupo, V. Production of Cocconeis neothumensis (Bacillariophyceae) biomass in batch cultures and bioreactors for biotechnological applications: Light and nutrient requirements. J. Appl. Phycol. 2007, 4, 383–391. [Google Scholar] [CrossRef]
- Nappo, M.; Berkov, S.; Codina, C.; Avila, C.; Messina, P.; Zupo, V.; Bastida, J. Metabolite profiling of the benthic diatom Cocconeis scutellum by GC-MS (2009). J. Appl. Physiol. 2009, 21, 295. [Google Scholar]
- Ruocco, N.; Costantini, S.; Zupo, V.; Lauritano, C.; Caramiello, D.; Ianora, A.; Budillon, A.; Romano, G.; Nuzzo, G.; D’Ippolito, G.; et al. Toxigenic effects of two benthic diatoms upon grazing activity of the sea urchin: Morphological, metabolomic and de novo transcriptomic analysis. Sci. Rep. 2018, 8, 5622. [Google Scholar] [CrossRef] [PubMed]
- Ruocco, N.; Cavaccini, V.; Caramiello, D.; Ianora, A.; Fontana, A.; Zupo, V.; Costantini, M. Noxious effects of the benthic diatoms Cocconeis scutellum and Diploneis sp. on sea urchin development: Morphological and de novo transcriptomic analysis. Harmful Algae 2019, 86, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Ruocco, N.; Costantini, S.; Zupo, V.; Romano, G.; Ianora, A.; Fontana, A.; Costantini, M. High-quality RNA extraction from the sea urchin Paracentrotus lividus embryos. PLoS ONE 2017, 12, e0172171. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2018, 47, 607–613. [Google Scholar] [CrossRef]
- Ragusa, M.A.; Costa, S.; Gianguzza, M.; Roccheri, M.C.; Gianguzza, F. Effects of cadmium exposure on sea urchin development assessed by SSH and RT-qPCR: Metallothionein genes and their differential induction. Mol. Biol. Rep. 2013, 40, 2157–2167. [Google Scholar] [CrossRef] [PubMed]
- Pinsino, A.; Bergami, E.; Della Torre, C.; Vannuccini, M.L. Amino modified polystyrene nanoparticles affect signalling pathways of the sea urchin (Paracentrotus lividus) embryos. Nanotoxicology 2017, 2, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in realtime RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Concentration (μM) | ||||||
|---|---|---|---|---|---|---|---|
| Decadienal | Heptadienal | Octadienal | 5-HEPE | 9-HEPE | 11-HEPE | 15-HEPE | |
| 1 | 0.5 | 1.0 | 1.5 | 2.8 | 2.8 | 2.8 | 2.8 |
| 2 | 0.4 | 0.8 | 1.3 | 2.1 | 2.1 | 2.1 | 2.1 |
| 3 | 0.3 | 0.7 | 1.0 | 1.8 | 1.8 | 1.8 | 1.8 |
| 4 | 0.3 | 0.7 | 1.0 | 1.6 | 1.6 | 1.6 | 1.6 |
| 5 | 0.25 | 0.5 | 0.75 | 1.4 | 1.4 | 1.4 | 1.4 |
| Gene Names | Acronym | Accession Number | Primer | Sequence(5′→3′) | Amplicon Length (bp) | Efficiency (%) |
|---|---|---|---|---|---|---|
| Antidorsalizing morphogenetic | ADMP2 | KT276376 | ADMP2_F1 | CGAGCAAGAGGCTCCTCC | 161 | 96 |
| protein 2 | ADMP2_R1 | CTTTGGCAACGCCGACAAAC | ||||
| ADP-ribosylation factor 1 | ARF1 | SUB2817153 * | ADP_F2 | CTGGTGCATCAGTAAACTAC | 256 | 100 |
| ADP_R2 | CGTCGTAAGAGCAGGAACG | |||||
| Brachyury | Bra | AJ419790 | BRA_F1 | CCGCGAACTCGACCGAAC | 178 | 94 |
| BRA_R1 | CAACCCGACAGAGCTCCC | |||||
| Delta | Delta | DQ536193 | DELTA_F1 | CTGCATGCGAGATGTCATG | 178 | 101 |
| DELTA_R1 | GCAACGTAGATGACCAGCTG | |||||
| Goosecoid | Goosecoid | HM449798 | Goosecoid_F1 | GCGCGCATGTCCAGACTG | 161 | 98 |
| Goosecoid_R1 | GTCCATCTCAAAGATCAGAGC | |||||
| Glyoxylate reductase | GRHPR | SUB2817153 * | GLR_F1 | GGCTCACAACAGATGAAGTAG | 171 | 100 |
| hydroxypyruvate reductase | GLR_R1 | CTTGGCGTATCTTCGTTCTC | ||||
| Histone H3.3 | H3.3 | X76081 | H3.3_F1 | GCTGCTATTGGTGCTCTG | 156 | 93 |
| H3.3_R1 | GATTGATCAGGATGGTCACG | |||||
| Kinesin-19 | KIF19 | SUB2817153 * | KIF19_F1 | CGCAGACTCATCACAGAGC | 176 | 106 |
| KIF19_R1 | GCCTTGGCCACCTTCATGC | |||||
| Notch | Notch | JQ861276 | Notch_F1 | CGAGAGGGAGGTGGATAGC | 188 | 94 |
| Notch_R1 | GGGTTGCTCACACAGTGG | |||||
| Poly(ADP-ribose) | PARP-1 | SUB2817153 * | pADP_F1 | GGCAATGATGCTGTCTGTAG | 184 | 101 |
| polymerase 2 | pADP_R1 | CAGTTACAGCACTGGTTTCATC | ||||
| Succinate dehydrogenase | SDH | SUB2817153 * | SDH_F2 | GGTGAGTAGGAAATTCACAGTG | 210 | 102 |
| assembly factor mitochondrial | SDH_R2 | CTCACGTCGGAATGTCTTCG | ||||
| Smad6 | Smad6 | HM449802 | Smad6_F1 | CGAGACCGACCTCAAAGC | 134 | 105 |
| Smad6_R1 | GTTCCCCTTTCGAGATGG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esposito, R.; Ruocco, N.; Albarano, L.; Ianora, A.; Manfra, L.; Libralato, G.; Costantini, M. Combined Effects of Diatom-Derived Oxylipins on the Sea Urchin Paracentrotus lividus. Int. J. Mol. Sci. 2020, 21, 719. https://doi.org/10.3390/ijms21030719
Esposito R, Ruocco N, Albarano L, Ianora A, Manfra L, Libralato G, Costantini M. Combined Effects of Diatom-Derived Oxylipins on the Sea Urchin Paracentrotus lividus. International Journal of Molecular Sciences. 2020; 21(3):719. https://doi.org/10.3390/ijms21030719
Chicago/Turabian StyleEsposito, Roberta, Nadia Ruocco, Luisa Albarano, Adrianna Ianora, Loredana Manfra, Giovanni Libralato, and Maria Costantini. 2020. "Combined Effects of Diatom-Derived Oxylipins on the Sea Urchin Paracentrotus lividus" International Journal of Molecular Sciences 21, no. 3: 719. https://doi.org/10.3390/ijms21030719
APA StyleEsposito, R., Ruocco, N., Albarano, L., Ianora, A., Manfra, L., Libralato, G., & Costantini, M. (2020). Combined Effects of Diatom-Derived Oxylipins on the Sea Urchin Paracentrotus lividus. International Journal of Molecular Sciences, 21(3), 719. https://doi.org/10.3390/ijms21030719