Differential RNA Editing and Intron Splicing in Soybean Mitochondria during Nodulation

 ,
,  and
and 
Abstract
1. Introduction
2. Results
2.1. RNA Sequencing and Differentially Expressed Mitochondrial Genes
2.2. Differential RNA Editing in Root Nodules
2.3. matR Transcripts Underwent Extensive RNA Editing in Root Nodules
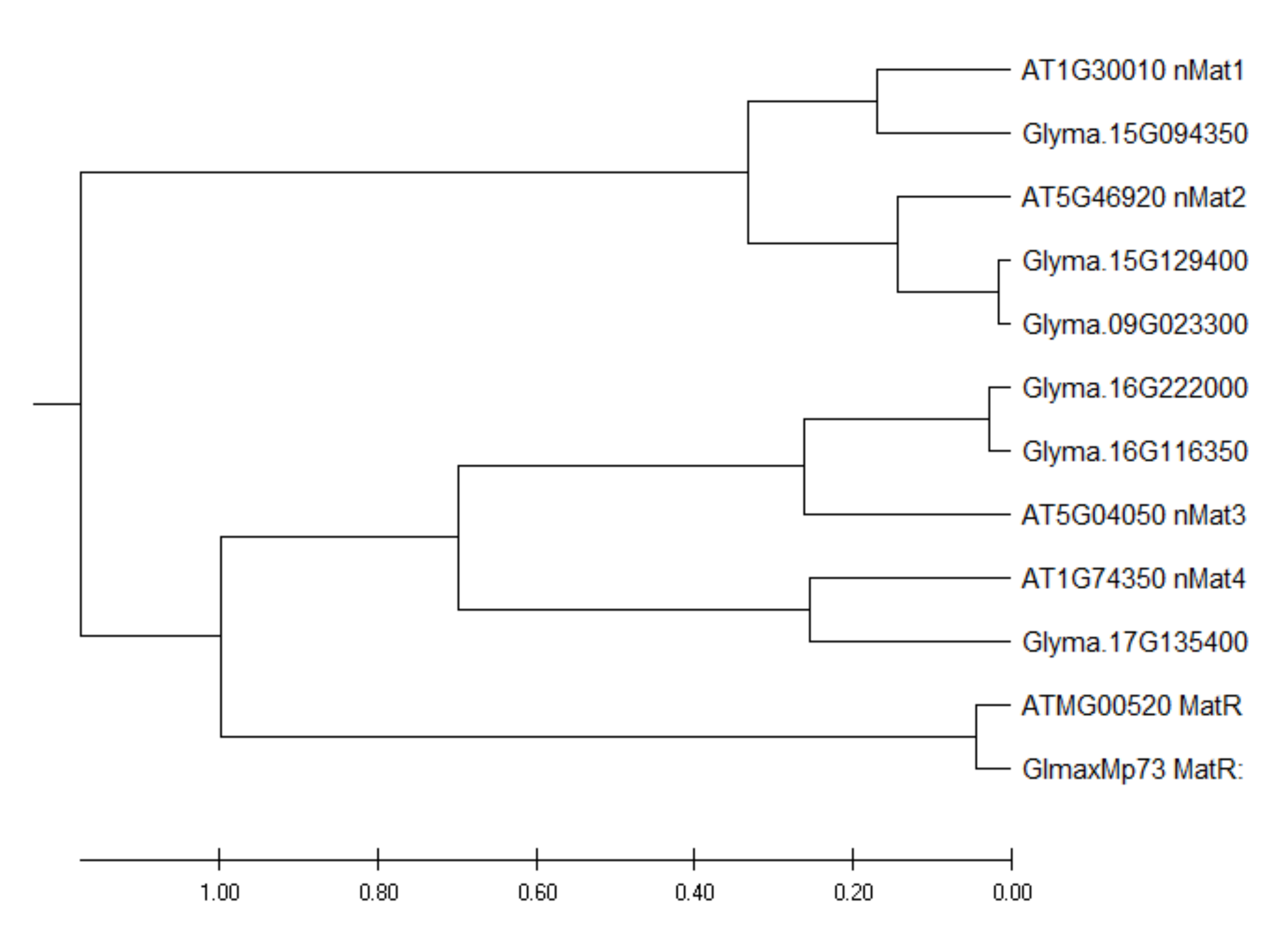
2.4. Identification of Nuclear-Encoded Mitochondrial Intron Maturases in the Soybean Genome
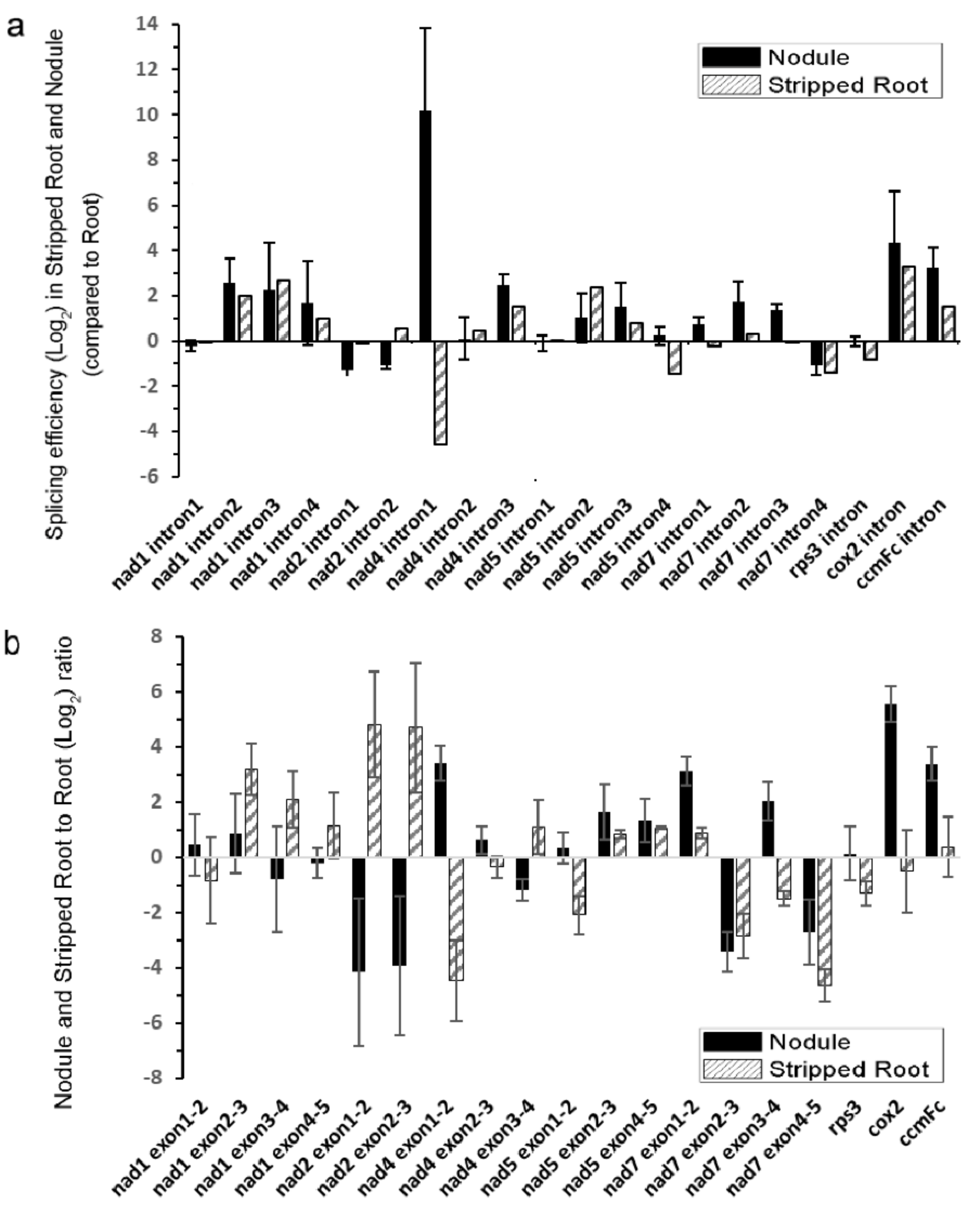
2.5. Intron Splicing of Mitochondrial Transcripts
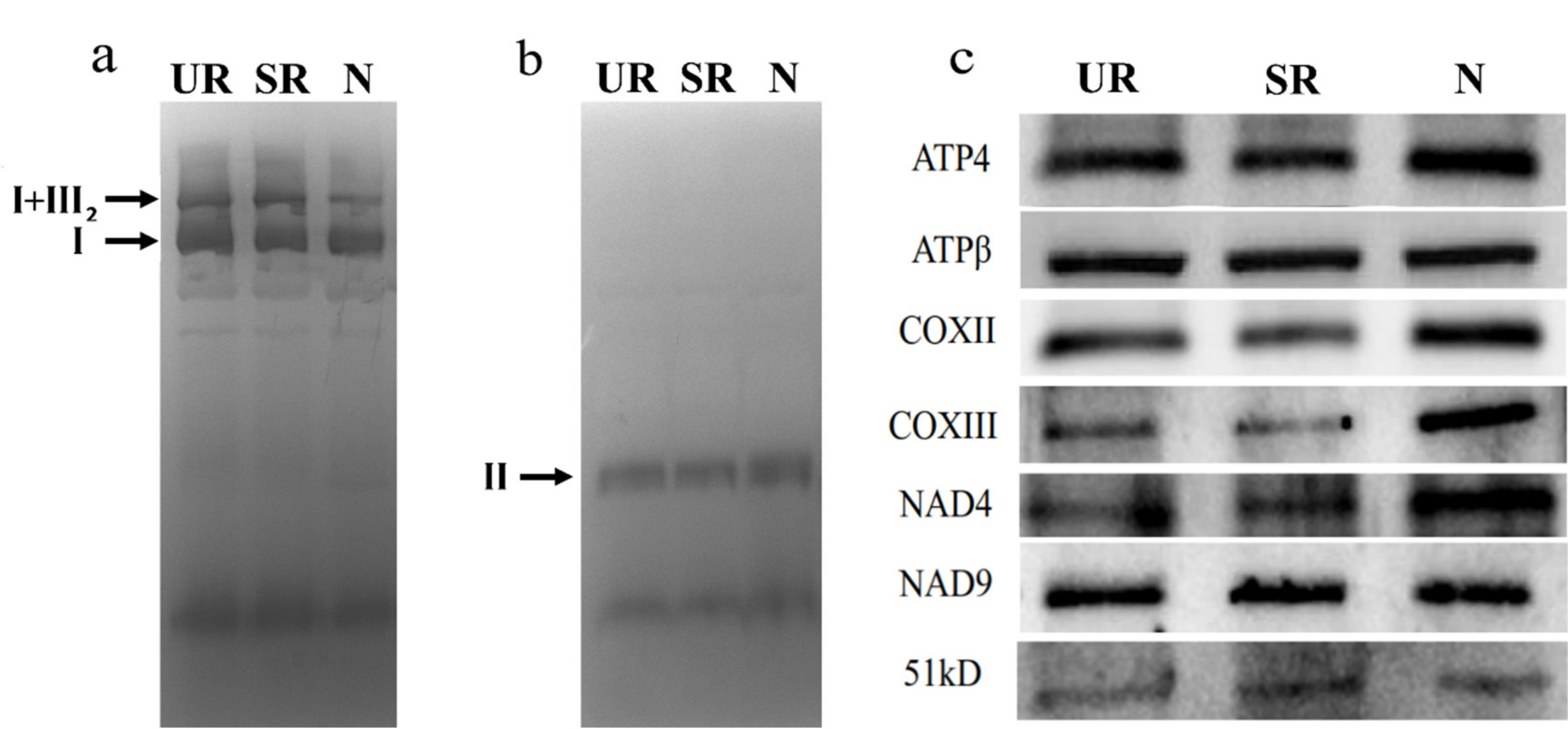
2.6. In-Gel Activity Assay and Western Blotting
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Sinorhizobium Fredii Inoculation
4.2. RNA Extraction and Sequencing
4.3. Bioinformatics Analysis
4.4. RT-PCR and Quantitative RT-PCR
4.5. Isolation of Soybean Mitochondria
4.6. Blue Native-Polyacrylamide Gel Electrophoresis (BN-PAGE) and In-Gel Enzyme Activity Staining
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, A.; Contador, C.; Fan, K.; Lam, H.-M. Interaction and regulation of carbon, nitrogen, and phosphorus metabolisms in root nodules of legumes. Front. Plant Sci. 2018, 9, 1860. [Google Scholar] [CrossRef] [PubMed]
- Copeland, L.; Vella, J.; Hong, Z. Enzymes of carbohydrate metabolism in soybean nodules. Phytochemistry 1989, 28, 57–61. [Google Scholar] [CrossRef]
- Dunn, M.F. Tricarboxylic acid cycle and anaplerotic enzymes in rhizobia. FEMS Microbiol. Rev. 1998, 22, 105–123. [Google Scholar] [CrossRef] [PubMed]
- Bryce, J.H.; Day, D.A. Tricarboxylic acid cycle activity in mitochondria from soybean nodules and cotyledons. J. Exp. Bot. 1990, 41, 961–967. [Google Scholar] [CrossRef]
- Law, Y.S.; Zhang, R.; Guan, X.; Cheng, S.; Sun, F.; Duncan, O.; Murcha, M.; Whelan, J.; Lim, B.L. Phosphorylation and dephosphorylation of the presequence of pMORF3 during import into mitochondria from Arabidopsis thaliana. Plant Physiol. 2015, 169, 1–12. [Google Scholar] [CrossRef]
- Law, Y.S.; Ngan, L.; Yan, J.; Kwok, L.Y.; Sun, Y.; Cheng, S.; Schwenkert, S.; Lim, B.L. Multiple kinases can phosphorylate the N-terminal sequences of mitochondrial proteins in Arabidopsis thaliana. Front. Plant Sci. 2018, 9, 982. [Google Scholar] [CrossRef]
- Mower, J.P. Variation in protein gene and intron content among land plant mitogenomes. Mitochondrion 2020, 53, 203. [Google Scholar] [CrossRef]
- Chang, S.; Wang, Y.; Lu, J.; Gai, J.; Li, J.; Chu, P.; Guan, R.; Zhao, T. The mitochondrial genome of soybean reveals complex genome structures and gene evolution at intercellular and phylogenetic levels. PLoS ONE 2013, 8, e56502. [Google Scholar]
- Takenaka, M.; Verbitskiy, D.; Merwe, J.A.V.D.; Zehrmann, A.; Brennicke, A. The process of RNA editing in plant mitochondria. Mitochondrion 2008, 8, 35–46. [Google Scholar] [CrossRef]
- Giegé, P.; Sweetlove, L.J.; Cognat, V.; Leaver, C.J. Coordination of nuclear and mitochondrial genome expression during mitochondrial biogenesis in Arabidopsis. Plant Cell 2005, 17, 1497–1512. [Google Scholar] [CrossRef]
- Ichinose, M.; Sugita, M. RNA editing and its molecular mechanism in plant organelles. Genes 2016, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Shi, X.; Friso, G.; Wijk, K.V.; Bentolila, S.; Hanson, M.R. A zinc ginger motif-containing protein is essential for chloroplast RNA editing. PLoS Genet. 2015, 11, e1005028. [Google Scholar] [CrossRef] [PubMed]
- Shikanai, T. RNA editing in plants: Machinery and flexibility of site recognition. Biochim. Biophys. Acta BBA Bioenerg. 2015, 1847, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Castandet, B.; Araya, A. RNA editing in plant organelles. Why make it easy? Biochem. Mosc. 2011, 76, 924–931. [Google Scholar] [CrossRef]
- Sun, Y.; Law, Y.S.; Cheng, S.; Lim, B.L. RNA editing of cytochrome c maturation transcripts is responsive to the energy status of leaf cells in Arabidopsis thaliana. Mitochondrion 2017, 35, 23. [Google Scholar] [CrossRef]
- Tseng, C.-C.; Lee, C.-J.; Chung, Y.-T.; Sung, T.-Y.; Hsieh, M.-H. Differential regulation of Arabidopsis plastid gene expression and RNA editing in non-photosynthetic tissues. Plant Mol. Biol. 2013, 82, 375–392. [Google Scholar] [CrossRef]
- Miyata, Y.; Sugita, M. Tissue- and stage-specific RNA editing of rps14 transcripts in moss (Physcomitrella patens) chloroplasts. J. Plant Physiol. 2004, 161, 113–115. [Google Scholar] [CrossRef]
- B#xF6;rner, G.V.; Mörl, M.; Wissinger, B.; Brennicke, A.; Schmelzer, C. RNA editing of a group II intron in Oenothera as a prerequisite for splicing. Mol. Gen. Genet. 1995, 246, 739–744. [Google Scholar] [CrossRef]
- Xu, C.; Song, S.; Yang, Y.; Lu, F.; Zhang, M.; Sun, F.; Jia, R.; Song, R.; Tan, B.-C. DEK46 performs C-to-U editing of a specific site in mitochondrial nad7 introns that is critical for intron splicing and seed development in maize. Plant J. 2020, 5, 1767. [Google Scholar] [CrossRef]
- Keren, I.; Bezawork-Geleta, A.; Kolton, M.; Maayan, I.; Belausov, E.; Levy, M.; Mett, A.; Gidoni, D.; Shaya, F.; Ostersetzer-Biran, O. AtnMat2, a nuclear-encoded maturase required for splicing of group-II introns in Arabidopsis mitochondria. RNA 2009, 15, 2299–2311. [Google Scholar] [CrossRef]
- Guo, W.; Mower, J.P. Evolution of plant mitochondrial intron-encoded maturases: Frequent lineage-specific loss and recurrent intracellular transfer to the nucleus. J. Mol. Evol. 2013, 77, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.G.; Francs-Small, C.C.D.; Ostersetzer-Biran, O. Group II intron splicing factors in plant mitochondria. Front. Plant Sci. 2014, 5, 35. [Google Scholar] [CrossRef] [PubMed]
- Keren, I.; Tal, L.; Francs-Small, C.C.D.; Araújo, W.L.; Shevtsov, S.; Shaya, F.; Fernie, A.R.; Small, I.; Ostersetzer-Biran, O. nMAT1, a nuclear-encoded maturase involved in the trans-splicing of nad1 intron 1, is essential for mitochondrial complex I assembly and function. Plant J. 2012, 71, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Bonen, L. Cis- and trans-splicing of group II introns in plant mitochondria. Mitochondrion 2008, 8, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Verbitskiy, D.; Härtel, B.; Zehrmann, A.; Brennicke, A.; Takenaka, M. The DYW-E-PPR protein MEF14 is required for RNA editing at site matR -1895 in mitochondria of Arabidopsis thaliana. FEBS Lett. 2011, 585, 700–704. [Google Scholar] [CrossRef]
- Cohen, S.; Zmudjak, M.; Francs-Small, C.C.d.; Malik, S.; Shaya, F.; Keren, I.; Belausov, E.; Many, Y.; Brown, G.G.; Small, I.; et al. nMAT4, a maturase factor required for nad1 pre-mRNA processing and maturation, is essential for holocomplex I biogenesis in Arabidopsis mitochondria. Plant J. 2014, 78, 253–268. [Google Scholar] [CrossRef]
- Sultan, L.D.; Mileshina, D.; Grewe, F.; Rolle, K.; Abudraham, S.; G#x142;odowicz, P.; Niazi, A.K.; Keren, I.; Shevtsov, S.; Klipcan, L.; et al. The reverse transcriptase/RNA maturase protein MatR is required for the splicing of various Group II introns in Brassicaceae mitochondria. Plant Cell 2016, 28, 2805. [Google Scholar] [CrossRef]
- Thomson, M.C.; Macfarlane, J.L.; Beagley, C.T.; Wolstenholme, D.R. RNA editing of matR transcripts in maize and soybean increases similarity of the encoded protein to fungal and bryophyte group II intron maturases: Evidence that matR encodes a functional protein. Nucleic Acids Res. 1994, 22, 5745–5752. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Small, I.; Schallenberg-Rüdinger, M.; Takenaka, M.; Mireau, H.; Ostersetzer-Biran, O. Plant organellar RNA editing: What 30 years of research has revealed. Plant J. 2019, 101, 1040. [Google Scholar] [CrossRef]
- Mareéchal-Drouard, L.; Ramamonjisoa, D.; Cosset, A.; Weil, J.; Dietrich, A. Editing corrects mispairing in the acceptor stem of bean and potato mitochondrial phenylalanine transfer RNAs. Nucleic Acids Res. 1993, 21, 4909–4914. [Google Scholar] [CrossRef]
- Sun, F.; Carrie, C.; Law, S.; Murcha, M.W.; Zhang, R.; Law, Y.S.; Suen, P.K.; Whelan, J.; Lim, B.L. AtPAP2 is a tail-anchored protein in the outer membrane of chloroplasts and mitochondria. Plant Signal. Behav. 2012, 7, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Suen, P.K.; Zhang, Y.; Liang, C.; Carrie, C.; Whelan, J.; Ward, J.L.; Hawkins, N.D.; Jiang, L.; Lim, B.L. A dual-targeted purple acid phosphatase in Arabidopsis thaliana moderates carbon metabolism and its overexpression leads to faster plant growth and higher seed yield. New Phytol. 2012, 194, 206–219. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Aubé, F.; Quadrado, M.; Dargel-Graffin, C.; Mireau, H. Three new pentatricopeptide repeat proteins facilitate the splicing of mitochondrial transcripts and complex I biogenesis in Arabidopsis. J. Exp. Bot. 2018, 69, 5131. [Google Scholar] [CrossRef] [PubMed]
- Hsu-Liang, H.; Wang, H.-J.; Hsieh, M.-H.; Hsieh, H.-L.; Jauh, G.-Y. Arabidopsis mTERF15 is required for mitochondrial nad2 intron 3 splicing and functional complex I activity. PLoS ONE 2014, 9, e112360. [Google Scholar] [CrossRef]
- Zhao, P.; Wang, F.; Li, N.; Shi, D.-Q.; Yang, W.-C. Pentatricopeptide repeat protein MID1 modulates nad2 intron 1 splicing and Arabidopsis development. Sci. Rep. 2020, 10, 2008. [Google Scholar] [CrossRef] [PubMed]
- Braun, H.-P.; Binder, S.; Brennicke, A.; Eubel, H.; Fernie, A.R.; Finkemeier, I.; Klodmann, J.; König, A.-C.; Kühn, >K.; Meyer, E.; et al. The life of plant mitochondrial complex I. Mitochondrion 2014, 19, 295–313. [Google Scholar] [CrossRef]
- Long, R. Faculty Opinions recommendation of Splicing enhances translation in mammalian cells: An additional function of the exon junction complex. Genes Dev. 2004, 18, 210–222. [Google Scholar] [CrossRef]
- Machinaga, A.; Ishihara, S.; Shirai, A.; Takase-Yoden, S. Splicing of friend murine leukemia virus env-mRNA enhances its ability to form polysomes. Front. Microbiol. 2016, 7, 160. [Google Scholar] [CrossRef]
- Karpova, O.V.; Newton, K.J. A partially assembled complex I in NAD4-deficient mitochondria of maize. Plant J. 1999, 17, 511–521. [Google Scholar] [CrossRef]
- Davies, K.M.; Blum, T.B.; Kuhlbrandt, W. Conserved in situ arrangement of complex I and III2 in mitochondrial respiratory chain supercomplexes of mammals, yeast, and plants. Proc. Natl. Acad. Sci. USA 2018, 115, 3024. [Google Scholar] [CrossRef] [PubMed]
- Hunte, C. Faculty Opinions recommendation of structure of a mitochondrial supercomplex formed by respiratory-chain complexes I and III. Proc. Natl. Acad. Sci. USA 2005, 102, 3225–3229. [Google Scholar] [CrossRef]
- Seefeldt, L.C.; Hoffman, B.M.; Dean, D.R. Mechanism of Mo-dependent nitrogenase. Annu. Rev. Biochem. 2009, 78, 701–722. [Google Scholar] [CrossRef] [PubMed]
- Mitsch, M.J.; DiCenzo, G.C.; Cowie, A.; Finan, T.M. Succinate transport is not essential for symbiotic nitrogen fixation by Sinorhizobium meliloti or Rhizobium leguminosarum. Appl. Environ. Microbiol. 2017, 84, 1. [Google Scholar] [CrossRef] [PubMed]
- Gardeström, P.; Bergman, A.; Ericson, I. Oxidation of glycine via the respiratory chain in mitochondria prepared from different parts of Spinach. Plant Physiol. 1980, 65, 389–391. [Google Scholar] [CrossRef]
- Hoa, L.T.-P.; Nomura, M.; Kajiwara, H.; Day, D.A.; Tajima, S. Proteomic analysis on symbiotic differentiation of mitochondria in soybean nodules. Plant Cell Physiol. 2004, 45, 300–308. [Google Scholar] [CrossRef]
- Tajima, S. Ureide biosynthesis in legume nodules. Front. Biosci. 2004, 9, 1374–1381. [Google Scholar] [CrossRef]
- Qi, X.; Li, M.-W.; Xie, M.; Liu, X.; Ni, M.; Shao, G.; Song, C.; Yim, A.K.-Y.; Tao, Y.; Wong, F.-L.; et al. Identification of a novel salt tolerance gene in wild soybean by whole-genome sequencing. Nat. Commun. 2014, 5, 4340. [Google Scholar] [CrossRef]
- Callow, J.A.; Vincent, J.M. A manual for the practical study of root-nodule bacteria. J. Appl. Ecol. 1971, 8, 977. [Google Scholar] [CrossRef]
- Rehman, H.M.; Cheung, W.-L.; Wong, K.-S.; Xie, M.; Luk, C.-Y.; Wong, F.-L.; Li, M.-W.; Tsai, S.-N.; To, W.-T.; Chan, L.-Y.; et al. High-throughput mass spectrometric analysis of the whole proteome and secretome from Sinorhizobium fredii strains CCBAU25509 and CCBAU45436. Front. Microbiol. 2019, 10, 2569. [Google Scholar] [CrossRef]
- Beringer, J.E. R factor transfer in Rhizobium leguminosarum. J. Gen. Microbiol. 1974, 84, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Auwera, G.A.V.D.; Carneiro, M.O.; Hartl, C.; Poplin, R.; Del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. From FastQ data to high-confidence variant calls: The genome analysis toolkit best practices pipeline. Curr. Protoc. Bioinform. 2013, 43, 10–11. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. Feature Counts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2013, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef]
- Koboldt, D.C.; Zhang, Q.; Larson, D.E.; Shen, D.; McLellan, M.D.; Lin, L.; Miller, C.A.; Mardis, E.R.; Ding, L.; Wilson, R.K. VarScan 2: Somatic mutation and copy number alteration discovery in cancer by exome sequencing. Genome Res. 2012, 22, 568–576. [Google Scholar] [CrossRef]
- Liang, C.; Zhang, Y.; Cheng, S.; Osorio, S.; Sun, Y.; Fernie, A.R.; Cheung, C.Y.M.; Lim, B.L. Impacts of high ATP supply from chloroplasts and mitochondria on the leaf metabolism of Arabidopsis thaliana. Front. Plant Sci. 2015, 6, 922. [Google Scholar] [CrossRef]
- Day, D.; Neuburger, M.; Douce, R. Biochemical characterization of chlorophyll-free mitochondria from pea leaves. Funct. Plant Biol. 1985, 12, 219–228. [Google Scholar] [CrossRef]
- Lister, R.; Carrie, C.; Duncan, O.; Ho, L.H.; Howell, K.A.; Murcha, M.W.; Whelan, J. Functional definition of outer membrane proteins involved in preprotein import into mitochondria. Plant Cell 2007, 19, 3739–3759. [Google Scholar] [CrossRef]
- Eubel, H.; Braun, H.-P.; Millar, A.H. Blue-native PAGE in plants: A tool in analysis of protein-protein interactions. Plant Methods 2005, 1, 11. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Read Length (bp) | Read Count | Total Length (bp) | |
|---|---|---|---|---|
| Stripped root | D3 | 125 | 66,851,390 | 8,356,423,750 |
| E3 | 125 | 75,008,428 | 9,376,053,500 | |
| F3 | 125 | 70,072,218 | 8,759,027,250 | |
| Nodule | D4 | 125 | 68,175,300 | 8,521,912,500 |
| E4 | 125 | 63,858,336 | 7,982,292,000 | |
| F4 | 125 | 62,615,136 | 7,826,892,000 | |
| Control Root | C08-Root1 | 150 | 111,230,354 | 16,684,553,100 |
| C08-Root2 | 150 | 106,852,728 | 16,027,909,200 | |
| C08-Root3 | 150 | 115,102,338 | 17,265,350,700 | |
| Gene ID | Gene | Function Description | UR FPKM | SR FPKM | N FPKM | N/UR | p-Value | N/SR | p-Value | SR/UR | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|---|
| GlmaxMp01 | ccmC | cytochrome c biogenesis C | 15,863 | 20,620 | 10,842 | 0.68 | 0.000 | 0.53 | 0.000 | 1.30 | 0.001 |
| GlmaxMp51 | ccmFn | cytochrome c biogenesis FN | 4048 | 2982 | 1586 | 0.39 | 0.000 | 0.53 | 0.000 | 0.74 | 0.006 |
| GlmaxMp04 | cox3 | cytochrome c oxidase subunit III | 31,643 | 31,486 | 60,520 | 1.91 | 0.000 | 1.92 | 0.000 | 1.00 | 0.943 |
| GlmaxMp20-2 | nad1 | NADH dehydrogenase subunit 1 | 12,430 | 21,952 | 27,133 | 2.18 | 0.000 | 1.24 | 0.002 | 1.77 | 0.000 |
| GlmaxMp31-1 | nad2 | NADH dehydrogenase subunit 2 | 12,565 | 19,324 | 16,626 | 1.32 | 0.048 | 0.86 | 0.001 | 1.54 | 0.009 |
| GlmaxMp32 | nad4L-1 | NADH dehydrogenase subunit 4L | 30,659 | 51,843 | 39,578 | 1.29 | 0.257 | 0.76 | 0.007 | 1.69 | 0.035 |
| GlmaxMp46 | nad4L-2 | NADH dehydrogenase subunit 4L | 29,685 | 47,502 | 36,863 | 1.24 | 0.353 | 0.78 | 0.009 | 1.60 | 0.057 |
| GlmaxMp05-1 | nad5 | NADH dehydrogenase subunit 5 | 17,739 | 26,994 | 25,170 | 1.42 | 0.025 | 0.93 | 0.036 | 1.52 | 0.014 |
| GlmaxMp05-2 | nad5 | NADH dehydrogenase subunit 5 | 26,303 | 42,023 | 55,654 | 2.12 | 0.000 | 1.32 | 0.003 | 1.60 | 0.004 |
| GlmaxMp07 | rp15 | ribosomal protein subunit L5 | 15,181 | 19,217 | 10,240 | 0.67 | 0.145 | 0.53 | 0.000 | 1.27 | 0.213 |
| GlmaxMp53 | rps1 | ribosomal protein S1 | 9695 | 21,173 | 17,874 | 1.84 | 0.013 | 0.84 | 0.005 | 2.18 | 0.004 |
| GlmaxMp16 | rps12 | ribosomal protein S12 | 10,421 | 11,110 | 19,813 | 1.90 | 0.001 | 1.78 | 0.000 | 1.07 | 0.572 |
| GlmaxMp08 | rps14 | ribosomal protein subunit S14 | 24,486 | 25,459 | 14,704 | 0.60 | 0.077 | 0.58 | 0.000 | 1.04 | 0.826 |
| GlmaxMp52 | rps4 | ribosomal protein S4 | 6737 | 15,174 | 16,296 | 2.42 | 0.004 | 1.07 | 0.012 | 2.25 | 0.006 |
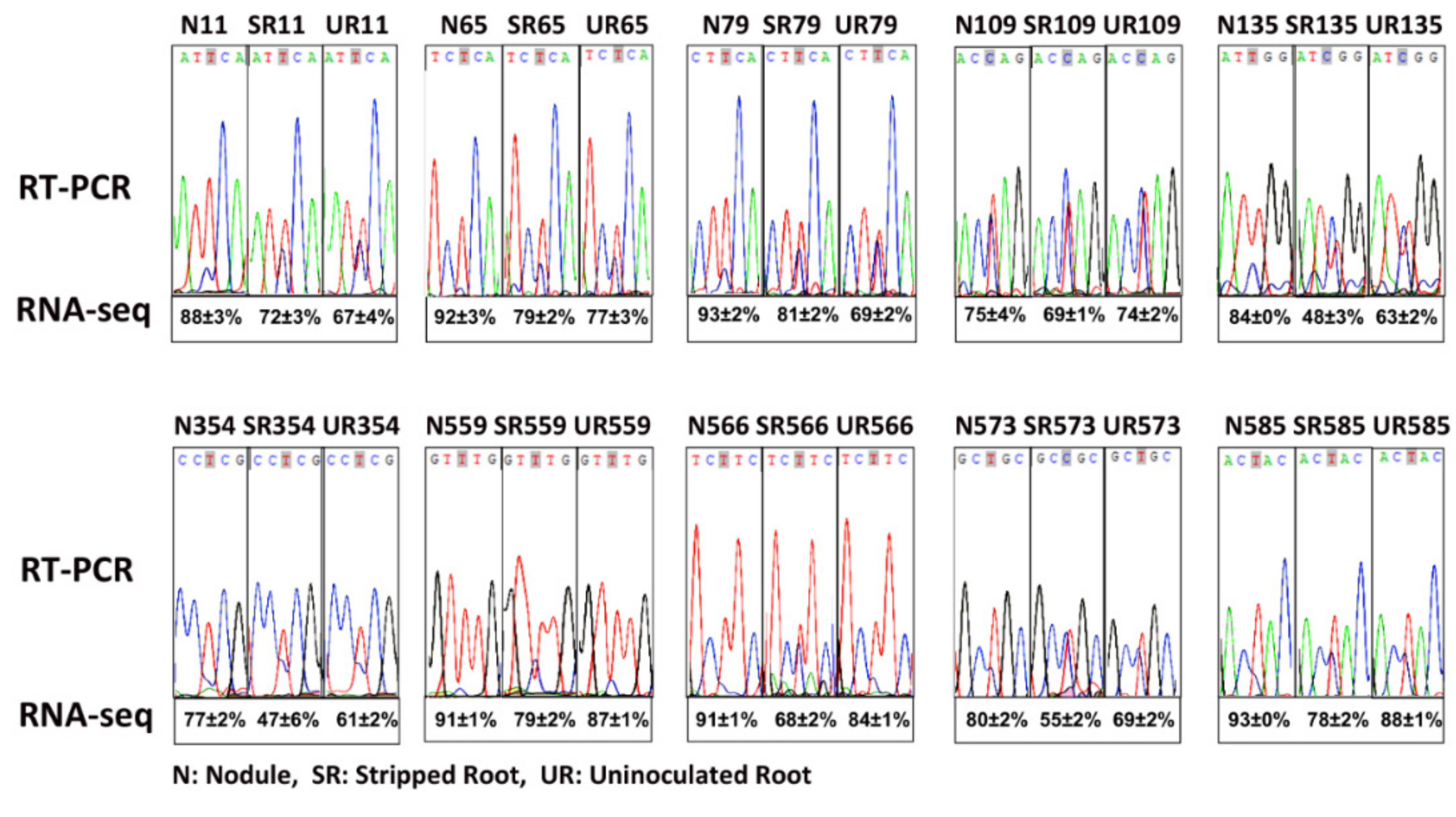
| Genome Position | Transcript Position | A.A. Position | Codon Change | A.A. Change | Nodule Average Editing Degree (%) | Stripped Root Average Editing Degree (%) | Root Average Editing Degree (%) | Changes (N-UR) (%) | Changes (N-SR) (%) | Changes (SR-UR) (%) | Editing Sites in Arabidopsis (Y/N) * | Editing Degree (%) * | Editing Sites in Soybean (Y/N) # |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 340,005 | 32 | 11 | TCC>-<TTC | S>-<F | 88 ± 3 a | 72 ± 3 b | 67 ± 4 b | 21 | 16 | 5 | No | - | Yes |
| 339,890 | 147 | 49 | TTC>-<TTT | No (F) | 26 ± 1 a | 12 ± 2 b | 13 ± 4 b | 13 | 14 | −1 | No | - | No |
| 339,844 | 193 | 65 | CCA>-<TCA | P>-<S | 92 ± 3 a | 79 ± 2 b | 77 ± 3 b | 15 | 13 | 2 | No | - | Yes |
| 339,801 | 236 | 79 | TCC>-<TTT | S>-<F | 93 ± 2 a | 81 ± 2 b | 69 ± 2 c | 24 | 12 | 12 | No | - | Yes |
| 339,711 | 326 | 109 | CCA>-<CTA | P>-<L | 75 ± 4 a | 69 ± 1 b | 74 ± 2 a | 0 | 6 | −6 | Yes | 87 | Yes |
| 339,633 | 404 | 135 | TCG>-<TTG | S>-<L | 84 ± 0 a | 48 ± 3 b | 63 ± 2 c | 20 | 35 | −15 | Yes | 90 | No |
| 339,126 | 911 | 304 | GCC>-<GTC | A>-<V | 17 ± 2 a | 13 ± 0 b | 19 ± 5 ab | −2 | 4 | −6 | No | - | No |
| 338,976 | 1061 | 354 | CCC>-<CTC | P>-< | 77 ± 2 a | 47 ± 6 b | 61 ± 2 c | 15 | 30 | −15 | No | - | Yes |
| 338,495 | 1542 | 514 | CCC>-<CCT | No (P) | 42 ± 1 a | 22 ± 2 b | 20 ± 4 b | 22 | 20 | 2 | Yes | 43 | No |
| 338,361 | 1676 | 559 | TCT>-<TTT | S>-<F | 91 ± 1 a | 79 ± 2 b | 87 ± 1 c | 4 | 13 | −9 | Yes | 88 | Yes |
| 338,340 | 1697 | 566 | CCT>-<CTT | P>-<L | 91 ± 1 a | 68 ± 2 b | 84 ± 1 c | 7 | 23 | −16 | Yes | 84 | Yes |
| 338,320 | 1717 | 573 | CGC>-<TGT | R>-<C | 80 ± 2 a | 55 ± 2 b | 69 ± 2 c | 11 | 25 | −14 | Yes | 83 | Yes |
| 338,306 | 1731 | 577 | TAC>-<TAT | No (Y) | 97 ± 1 a | 93 ± 1 b | 95 ± 0 c | 2 | 4 | −2 | No | - | No |
| 338,284 | 1753 | 585 | CAC>-<TAC | H>-<Y | 93 ± 0 a | 78 ± 2 b | 88 ± 1 c | 5 | 15 | −10 | Yes | 79 | Yes |
| 338,214 | 1823 | 608 | CCC>-<CTC | P>-<L | 83 ± 1 a | 72 ± 1 b | 71 ± 1 b | 12 | 11 | 2 | No | - | Yes |
| 338,196 | 1841 | 614 | TCA>-<TTA | S>-<L | 97 ± 1 a | 91 ± 1 b | 96 ± 1 a | 1 | 7 | −5 | Yes | 93 | Yes |
| 338,186 | 1851 | 617 | GTC>-<GTT | No (V) | 42 ± 2 a | 33 ± 1 b | 39 ± 2a | 3 | 9 | −6 | No | - | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Xie, M.; Xu, Z.; Chan, K.C.; Zhong, J.Y.; Fan, K.; Wong-Bajracharya, J.; Lam, H.-M.; Lim, B.L. Differential RNA Editing and Intron Splicing in Soybean Mitochondria during Nodulation. Int. J. Mol. Sci. 2020, 21, 9378. https://doi.org/10.3390/ijms21249378
Sun Y, Xie M, Xu Z, Chan KC, Zhong JY, Fan K, Wong-Bajracharya J, Lam H-M, Lim BL. Differential RNA Editing and Intron Splicing in Soybean Mitochondria during Nodulation. International Journal of Molecular Sciences. 2020; 21(24):9378. https://doi.org/10.3390/ijms21249378
Chicago/Turabian StyleSun, Yuzhe, Min Xie, Zhou Xu, Koon Chuen Chan, Jia Yi Zhong, Kejing Fan, Johanna Wong-Bajracharya, Hon-Ming Lam, and Boon Leong Lim. 2020. "Differential RNA Editing and Intron Splicing in Soybean Mitochondria during Nodulation" International Journal of Molecular Sciences 21, no. 24: 9378. https://doi.org/10.3390/ijms21249378
APA StyleSun, Y., Xie, M., Xu, Z., Chan, K. C., Zhong, J. Y., Fan, K., Wong-Bajracharya, J., Lam, H.-M., & Lim, B. L. (2020). Differential RNA Editing and Intron Splicing in Soybean Mitochondria during Nodulation. International Journal of Molecular Sciences, 21(24), 9378. https://doi.org/10.3390/ijms21249378