Cell-free Stem Cell-Derived Extract Formulation for Regenerative Medicine Applications
 ,
, 


Abstract
1. Introduction
2. Results
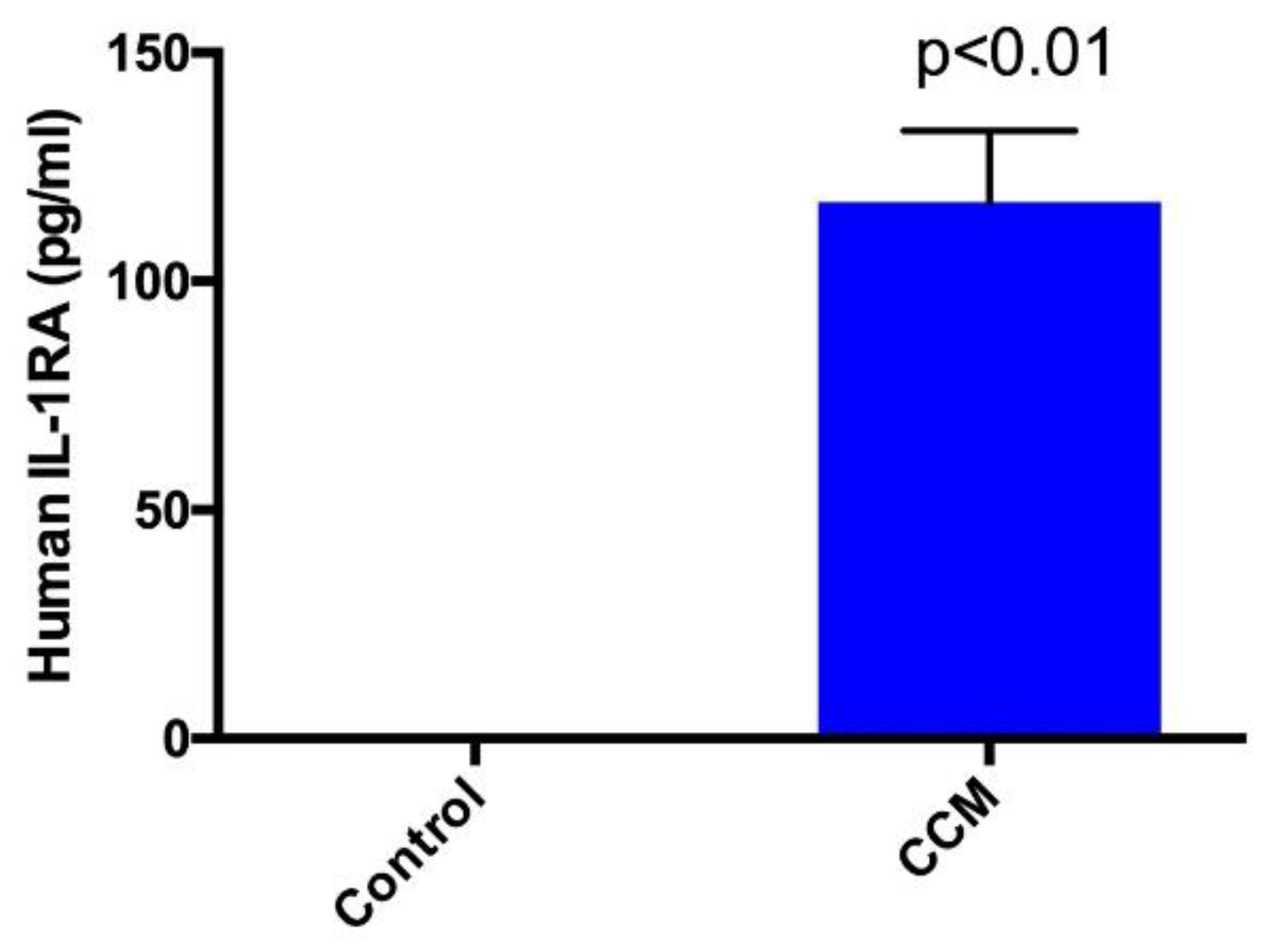
2.1. Enzyme-Linked Immunosorbent Assay (ELISA)
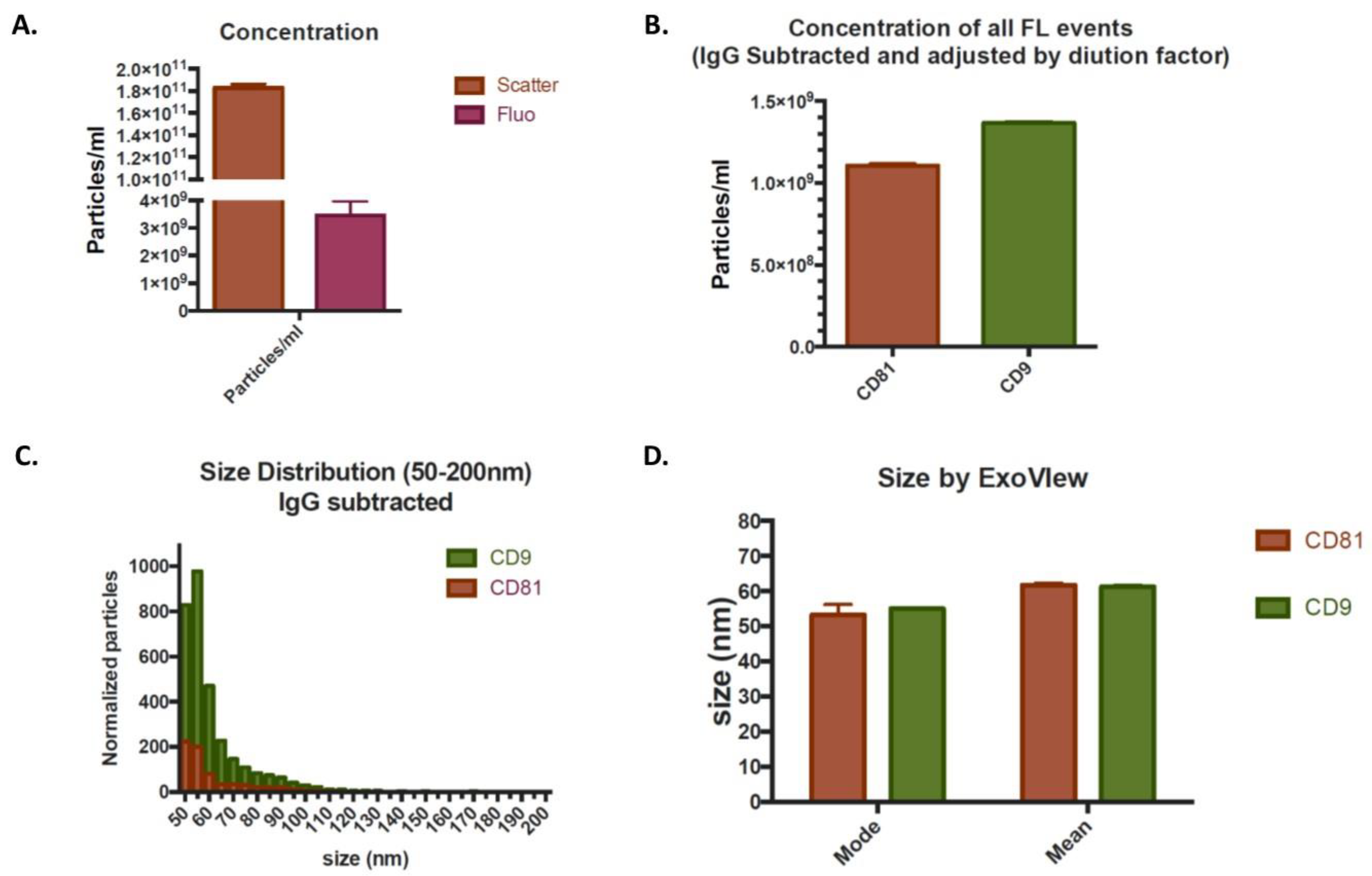
2.2. Exosome Analysis
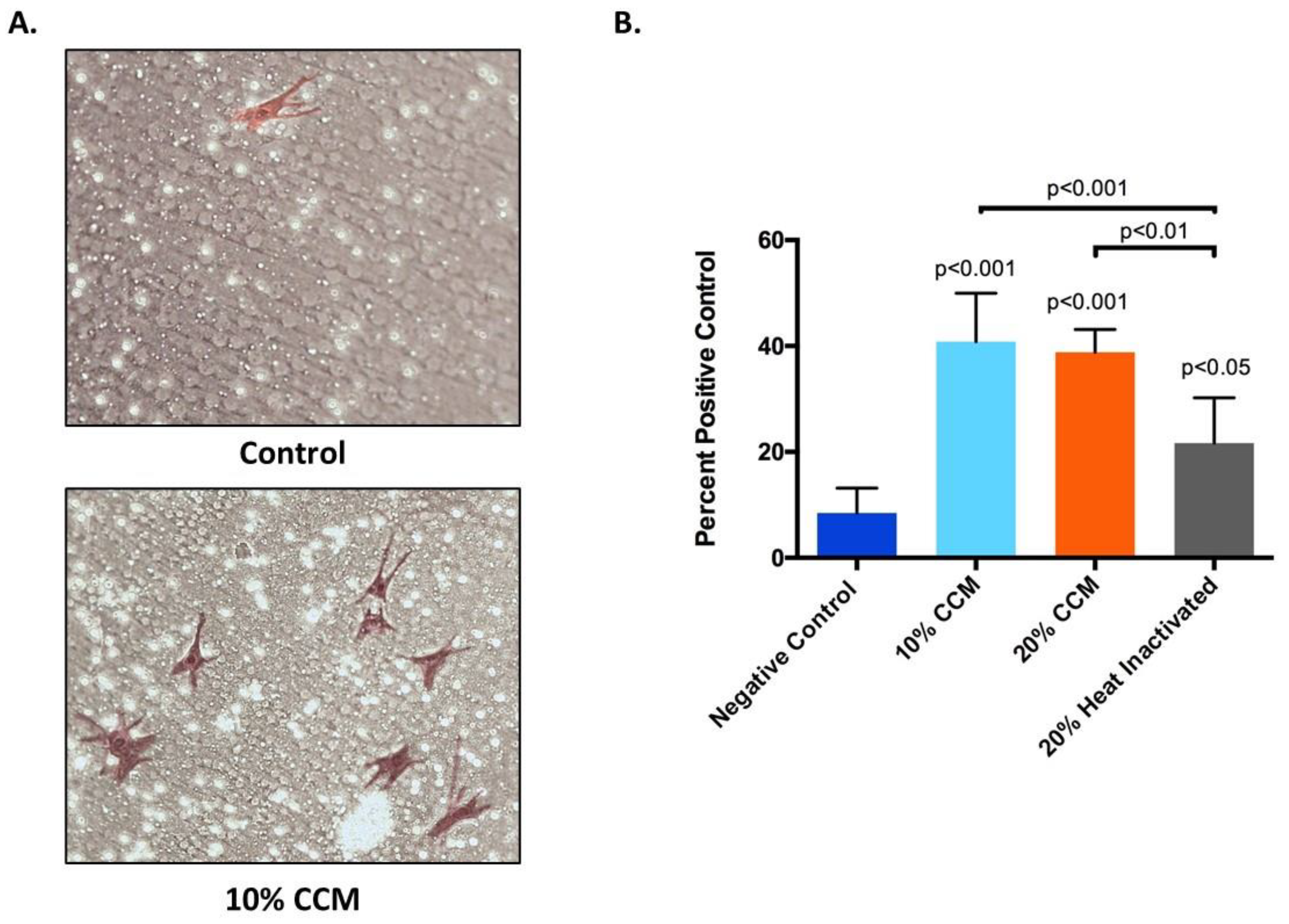
2.3. Cell Proliferation
2.4. Cell Migration
3. Discussion
4. Materials and Methods
4.1. Cells, Reagents, and Supplies
4.2. Cell Culture and Formulation of Novel Cell-Free Stem Cell-Derived Extract
4.3. Enzyme-Linked Immunosorbent Assay (ELISA)
4.4. Exosome Analysis
4.5. Cell Proliferation Assay
4.6. Cell Migration Assay
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| BMSC | Bone marrow mesenchymal stem cells |
| CCM | Cell-free stem cell-derived extract |
| CKs | Cytokines |
| ECM | Extracellular matrix |
| ELISA | Enzyme linked immunosorbent assay |
| EGFR | Epidermal growth factor receptor |
| EVs | Extracellular vesicles |
| GFs | Growth factors |
| GH | Growth hormone |
| IGF-1 | Insulin-like growth factor-1 |
| IGFBP | Insulin-like growth factor binding protein |
| IL-1RA | Interleukin 1 receptor antagonist |
| MSCs | Mesenchymal stem cells |
| OS | Osteoarthritis |
| PDGF-AA | Platelet derived growth factor-AA |
| SP-IRIS | Single particle interferometric reflectance imaging sensing |
| SEM | Standard error of mean |
| TGFα | Transforming growth factor alpha |
| TGFβ1 | Transforming growth factor beta-1 |
| VEGF | Vascular endothelial growth factor |
References
- Navani, A.; Manchikanti, L.; Albers, S.L.; Latchaw, R.E.; Sanapati, J.; Kaye, A.D.; Atluri, S.; Jordan, S.; Gupta, A.; Cedeno, D.L.; et al. Responsible, Safe, and Effective Use of Biologics in the Management of Low Back Pain: American Society of Interventional Pain Physicians (ASIPP) Guidelines. Pain Physician 2019, 22, S1–S74. [Google Scholar] [PubMed]
- Lamplot, J.D.; Rodeo, S.A.; Brophy, R.H. A Practical Guide for the Current Use of Biologic Therapies in Sports Medicine. Am. J. Sports Med. 2020, 48, 488–503. [Google Scholar] [CrossRef] [PubMed]
- Le, A.D.K.; Enweze, L.; DeBaun, M.R.; Dragoo, J.L. Platelet-Rich Plasma. Clin. Sports Med. 2019, 38, 17–44. [Google Scholar] [CrossRef]
- Duerr, R.A.; Ackermann, J.; Gomoll, A.H. Amniotic-Derived Treatments and Formulations. Clin. Sports Med. 2019, 38, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.M.; Saleh, K.S.; Burdick, J.A.; Mauck, R.L. Bioactive factors for cartilage repair and regeneration: Improving delivery, retention, and activity. Acta Biomater. 2019, 93, 222–238. [Google Scholar] [CrossRef]
- Ioannidou, E. Therapeutic modulation of growth factors and cytokines in regenerative medicine. Curr. Pharm. Des. 2006, 12, 2397–2408. [Google Scholar] [CrossRef]
- Caplan, A.I. Mesenchymal stem cells. J. Orthop. Res. 1991, 9, 641–650. [Google Scholar] [CrossRef]
- Andia, I.; Maffulli, N. New biotechnologies for musculoskeletal injuries. Surgeon 2019, 17, 244–255. [Google Scholar] [CrossRef]
- Minguell, J.J.; Erices, A.; Conget, P. Mesenchymal stem cells. Exp. Biol. Med. 2001, 226, 507–520. [Google Scholar] [CrossRef]
- Potty, A.G.R.; Gupta, A.; Rodriguez, H.C.; Stone, I.W.; Maffulli, N. Intraosseous Bioplasty for a Subchondral Cyst in the Lateral Condyle of Femur. J. Clin. Med. 2020, 9, 1358. [Google Scholar] [CrossRef]
- Wang, S.; Guo, L.; Ge, J.; Yu, L.; Cai, T.; Tian, R.; Jiang, Y.; Zhao, R.C.H.; Wu, Y. Excess Integrins Cause Lung Entrapment of Mesenchymal Stem Cells. Stem. Cells 2015, 33, 3315–3326. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.O.; Han, J.W.; Kim, J.M.; Cho, H.J.; Park, C.; Lee, N.; Kim, D.W.; Yoon, Y.S. Malignant tumor formation after transplantation of short-term cultured bone marrow mesenchymal stem cells in experimental myocardial infarction and diabetic neuropathy. Circ. Res. 2011, 108, 1340–1347. [Google Scholar] [CrossRef] [PubMed]
- Barkholt, L.; Flory, E.; Jekerle, V.; Lucas-Samuel, S.; Ahnert, P.; Bisset, L.; Buscher, D.; Fibbe, W.; Foussat, A.; Kwa, M.; et al. Risk of tumorigenicity in mesenchymal stromal cell-based therapies--bridging scientific observations and regulatory viewpoints. Cytotherapy 2013, 15, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Huang, J.; Geng, Y.; Qian, H.; Wang, F.; Liu, X.; Shang, M.; Nie, S.; Liu, N.; Du, X.; et al. Paracrine action of mesenchymal stem cells revealed by single cell gene profiling in infarcted murine hearts. PLoS ONE 2015, 10, e0129164. [Google Scholar] [CrossRef]
- Deng, K.; Lin, D.L.; Hanzlicek, B.; Balog, B.; Penn, M.S.; Kiedrowski, M.J.; Hu, Z.; Ye, Z.; Zhu, H.; Damaser, M.S. Mesenchymal stem cells and their secretome partially restore nerve and urethral function in a dual muscle and nerve injury stress urinary incontinence model. Am. J. Physiol. Ren. Physiol. 2015, 308, F92–F100. [Google Scholar] [CrossRef]
- Chang, Y.H.; Wu, K.C.; Harn, H.J.; Lin, S.Z.; Ding, D.C. Exosomes and Stem Cells in Degenerative Disease Diagnosis and Therapy. Cell Transpl. 2018, 27, 349–363. [Google Scholar] [CrossRef]
- Mitchell, A.C.; Briquez, P.S.; Hubbell, J.A.; Cochran, J.R. Engineering growth factors for regenerative medicine applications. Acta Biomater. 2016, 30, 1–12. [Google Scholar] [CrossRef]
- Onishi, M.; Nosaka, T.; Kitamura, T. Cytokine receptors: Structures and signal transduction. Int. Rev. Immunol. 1998, 16, 617–634. [Google Scholar] [CrossRef]
- Barrientos, S.; Brem, H.; Stojadinovic, O.; Tomic-Canic, M. Clinical application of growth factors and cytokines in wound healing. Wound. Repair. Regen. 2014, 22, 569–578. [Google Scholar] [CrossRef]
- Heldring, N.; Mäger, I.; Wood, M.J.; Le Blanc, K.; Andaloussi, S.E. Therapeutic Potential of Multipotent Mesenchymal Stromal Cells and Their Extracellular Vesicles. Hum. Gene. Ther. 2015, 26, 506–517. [Google Scholar] [CrossRef]
- Gupta, A.; Kashte, S.; Gupta, M.; Rodriguez, H.C.; Gautam, S.S.; Kadam, S. Mesenchymal stem cells and exosome therapy for COVID-19: Current status and future perspective. Hum. Cell 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Matei, A.C.; Antounians, L.; Zani, A. Extracellular Vesicles as a Potential Therapy for Neonatal Conditions: State of the Art and Challenges in Clinical Translation. Pharmaceutics 2019, 11, 404. [Google Scholar] [CrossRef] [PubMed]
- Bagno, L.; Hatzistergos, K.E.; Balkan, W.; Hare, J.M. Mesenchymal Stem Cell-Based Therapy for Cardiovascular Disease: Progress and Challenges. Mol. Ther. 2018, 26, 1610–1623. [Google Scholar] [CrossRef]
- Lou, G.; Chen, Z.; Zheng, M.; Liu, Y. Mesenchymal stem cell-derived exosomes as a new therapeutic strategy for liver diseases. Exp. Mol. Med. 2017, 49, e346. [Google Scholar] [CrossRef]
- Liew, L.C.; Katsuda, T.; Gailhouste, L.; Nakagama, H.; Ochiya, T. Mesenchymal stem cell-derived extracellular vesicles: A glimmer of hope in treating Alzheimer’s disease. Int. Immunol. 2017, 29, 11–19. [Google Scholar] [CrossRef]
- Börger, V.; Bremer, M.; Ferrer-Tur, R.; Gockeln, L.; Stambouli, O.; Becic, A.; Giebel, B. Mesenchymal Stem/Stromal Cell-Derived Extracellular Vesicles and Their Potential as Novel Immunomodulatory Therapeutic Agents. Int. J. Mol. Sci. 2017, 18, 1450. [Google Scholar] [CrossRef]
- Gupta, A.; El-Amin, S.F.; Levy, H.J.; Sze-Tu, R.; Ibim, S.E.; Maffulli, N. Umbilical cord-derived Wharton’s jelly for regenerative medicine applications. J. Orthop. Surg. Res. 2020, 15, 49. [Google Scholar] [CrossRef]
- Thrailkill, K.M.; Lumpkin, C.K.; Bunn, R.C.; Kemp, S.F.; Fowlkes, J.L. Is insulin an anabolic agent in bone? Dissecting the diabetic bone for clues. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E735–E745. [Google Scholar] [CrossRef]
- Zhang, X.; Tamasi, J.; Lu, X.; Zhu, J.; Chen, H.; Tian, X.; Lee, T.C.; Threadgill, D.W.; Kream, B.E.; Kang, Y.; et al. Epidermal growth factor receptor plays an anabolic role in bone metabolism in vivo. J. Bone Miner. Res. 2011, 26, 1022–1034. [Google Scholar] [CrossRef]
- Edwards, D.R.; Murphy, G.; Reynolds, J.J.; Whitham, S.E.; Docherty, A.J.; Angel, P.; Heath, J.K. Transforming growth factor beta modulates the expression of collagenase and metalloproteinase inhibitor. EMBO J. 1987, 6, 1899–1904. [Google Scholar] [CrossRef] [PubMed]
- van Beuningen, H.M.; van der Kraan, P.M.; Arntz, O.J.; van den Berg, W.B. Transforming growth factor-beta 1 stimulates articular chondrocyte proteoglycan synthesis and induces osteophyte formation in the murine knee joint. Lab. Investig. 1994, 71, 279–290. [Google Scholar] [PubMed]
- Blaney Davidson, E.N.; Vitters, E.L.; van den Berg, W.B.; van der Kraan, P.M. TGF beta-induced cartilage repair is maintained but fibrosis is blocked in the presence of Smad7. Arthr. Res. Ther. 2006, 8, R65. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Olsen, B.R. The roles of vascular endothelial growth factor in bone repair and regeneration. Bone 2016, 91, 30–38. [Google Scholar] [CrossRef]
- Frizziero, A.; Giannotti, E.; Oliva, F.; Masiero, S.; Maffulli, N. Autologous conditioned serum for the treatment of osteoarthritis and other possible applications in musculoskeletal disorders. Br. Med. Bull. 2013, 105, 169–184. [Google Scholar] [CrossRef]
- Amable, P.R.; Teixeira, M.V.; Carias, R.B.; Granjeiro, J.M.; Borojevic, R. Protein synthesis and secretion in human mesenchymal cells derived from bone marrow, adipose tissue and Wharton’s jelly. Stem Cell Res. Ther. 2014, 5, 53. [Google Scholar] [CrossRef]
- Wei, F.; Li, Z.; Crawford, R.; Xiao, Y.; Zhou, Y. Immunoregulatory role of exosomes derived from differentiating mesenchymal stromal cells on inflammation and osteogenesis. J. Tissue Eng. Regen. Med. 2019, 13, 1978–1991. [Google Scholar] [CrossRef]
- Connor, D.E.; Paulus, J.A.; Dabestani, P.J.; Thankam, F.K.; Dilisio, M.F.; Gross, R.M.; Agrawal, D.K. Therapeutic potential of exosomes in rotator cuff tendon healing. J. Bone Miner. Metab. 2019, 37, 759–767. [Google Scholar] [CrossRef]
- Liu, Y.; Zou, R.; Wang, Z.; Wen, C.; Zhang, F.; Lin, F. Exosomal KLF3-AS1 from hMSCs promoted cartilage repair and chondrocyte proliferation in osteoarthritis. Biochem. J. 2018, 475, 3629–3638. [Google Scholar] [CrossRef]
- Fu, X.; Liu, G.; Halim, A.; Ju, Y.; Luo, Q.; Song, A.G. Mesenchymal Stem Cell Migration and Tissue Repair. Cells 2019, 8, 784. [Google Scholar] [CrossRef]
- Frizziero, A.; Vittadini, F.; Oliva, F.; Abatangelo, G.; Bacciu, S.; Berardi, A.; Maffulli, N.; Vertuccio, A.; Pintus, E.; Romiti, D.; et al. I.S. Mu. L.T. Hyaluronic acid injections in musculoskeletal disorders guidelines. MLTJ 2018, 8, 364–398. [Google Scholar] [CrossRef]
- Sekiya, I.; Larson, B.; Smith, J.; Pochampally, R.; Cui, J.; Prockop, D. Expansion of human adult stem cells from bone marrow stroma: Conditions that maximize the yields of early progenitors and evaluate their quality. Stem Cells 2002, 20, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Colter, D.; Class, R.; DiGirolamo, C.; Prockop, D. Rapid expansion of recycling stem cells in cultures of plastic-adherent cells from bone marrow. Proc. Natl. Acad. Sci. USA 2000, 97, 3213–3218. [Google Scholar] [CrossRef] [PubMed]
- Bachurski, D.; Schuldner, M.; Nguyen, P.H.; Malz, A.; Reiners, K.S.; Grenzi, P.C.; Babatz, F.; Schauss, A.C.; Hansen, H.P.; Hallek, M.; et al. Extracellular vesicle measurements with nanoparticle tracking analysis—An accuracy and repeatability comparison between NanoSight NS300 and ZetaView. J. Extracell. Vesicles 2019, 8, 1596016. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Factors | Average Amount (pg/mL) | |
|---|---|---|
| IGFBP-1 | Insulin-like growth factor-binding protein-1 | 70 |
| IGFBP-2 | Insulin-like growth factor-binding protein-2 | 18.1 |
| IGFBP-3 | Insulin-like growth factor-binding protein-3 | 552.6 |
| IGFBP-6 | Insulin-like growth factor-binding protein-6 | 20.3 |
| Insulin | Insulin | 518.5 |
| GH | Growth hormone | 10.5 |
| FGF-7 | Fibroblast growth factor-7 | 45.5 |
| HB-EGF | Heparin-binding EGF-like growth factor | 42.2 |
| PDGF-AA | Platelet derived growth factor-AA | 58.1 |
| TGF-α | Transforming growth factor alpha | 2.4 |
| TGFβ1 | Transforming growth factor beta 1 | 1036.90 |
| EG-VEGF | Endocrine-gland-derived vascular endothelial growth factor | 86.6 |
| VEGF | Vascular endothelial growth factor | 30.8 |
| VEGF R2 | Vascular endothelial growth factor-receptor 2 | 20.6 |
| VEGF R3 | Vascular endothelial growth factor-receptor 3 | 300 |
| VEGF-D | Vascular endothelial growth factor D | 16.6 |
| BDNF | Brain-derived neurotrophic factor | 68.6 |
| b-NGF | Beta-nerve Growth factor | 9.5 |
| NGF-R | Nerve growth factor receptor | 21 |
| NT-4 | Neurotrophin-4 | 110.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupta, A.; Cady, C.; Fauser, A.-M.; Rodriguez, H.C.; Mistovich, R.J.; Potty, A.G.R.; Maffulli, N. Cell-free Stem Cell-Derived Extract Formulation for Regenerative Medicine Applications. Int. J. Mol. Sci. 2020, 21, 9364. https://doi.org/10.3390/ijms21249364
Gupta A, Cady C, Fauser A-M, Rodriguez HC, Mistovich RJ, Potty AGR, Maffulli N. Cell-free Stem Cell-Derived Extract Formulation for Regenerative Medicine Applications. International Journal of Molecular Sciences. 2020; 21(24):9364. https://doi.org/10.3390/ijms21249364
Chicago/Turabian StyleGupta, Ashim, Craig Cady, Anne-Marie Fauser, Hugo C. Rodriguez, R. Justin Mistovich, Anish G. R. Potty, and Nicola Maffulli. 2020. "Cell-free Stem Cell-Derived Extract Formulation for Regenerative Medicine Applications" International Journal of Molecular Sciences 21, no. 24: 9364. https://doi.org/10.3390/ijms21249364
APA StyleGupta, A., Cady, C., Fauser, A.-M., Rodriguez, H. C., Mistovich, R. J., Potty, A. G. R., & Maffulli, N. (2020). Cell-free Stem Cell-Derived Extract Formulation for Regenerative Medicine Applications. International Journal of Molecular Sciences, 21(24), 9364. https://doi.org/10.3390/ijms21249364