Food-Derived Opioid Peptides in Human Health: A Review





Abstract
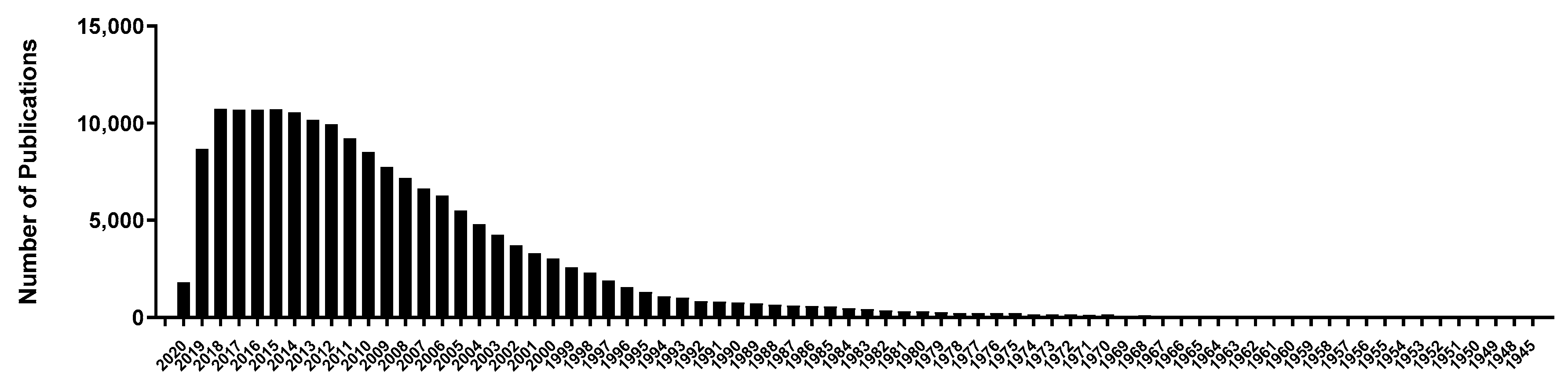
1. Introduction
Opioid Peptides
2. Classification of Opioid Peptides
2.1. Endogenous Opioid Peptides
2.2. Food Derived Exogenous Opioid Peptides
3. Production of Opioid Peptides from Food Proteins
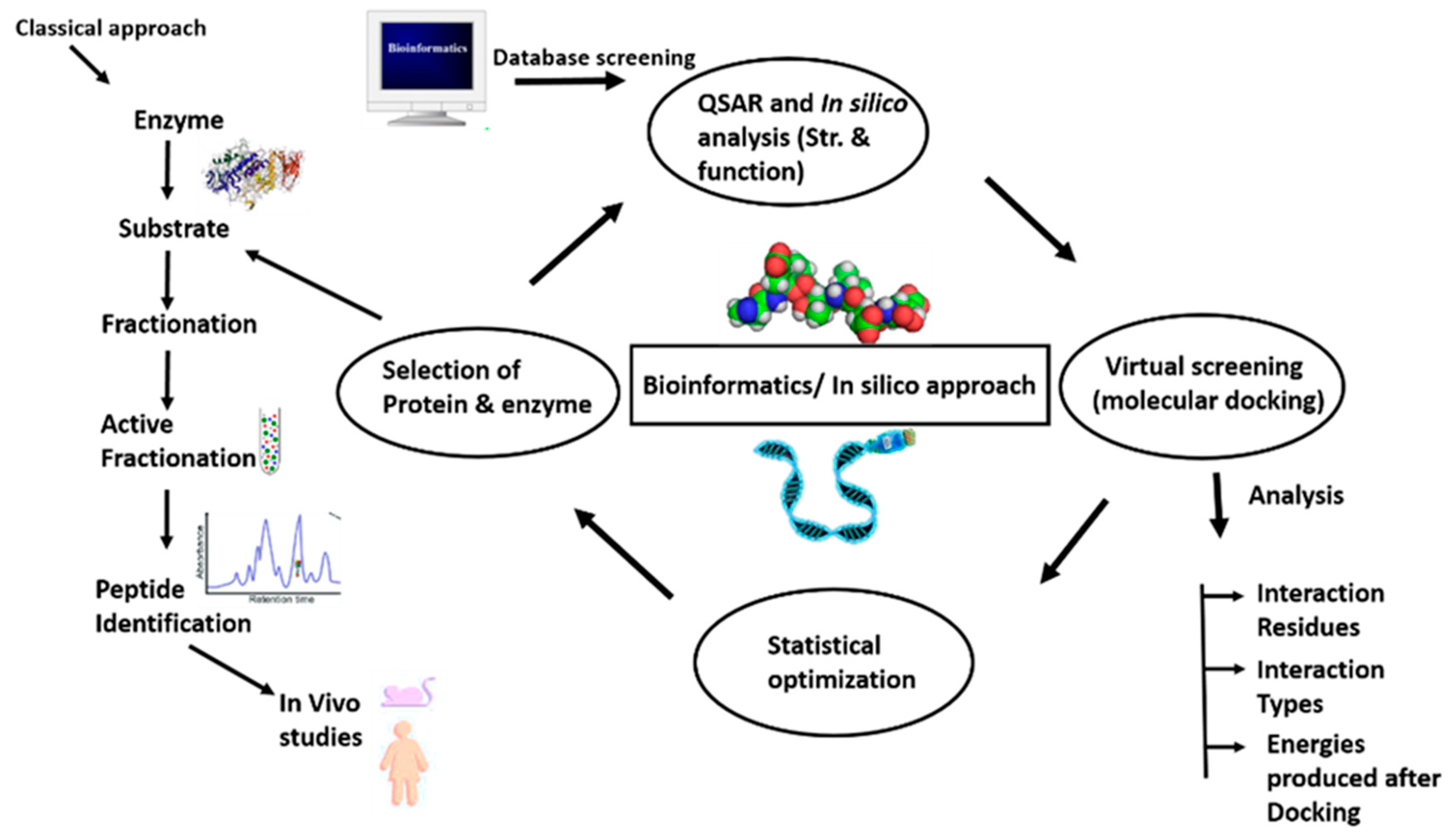
3.1. The Traditional Approach
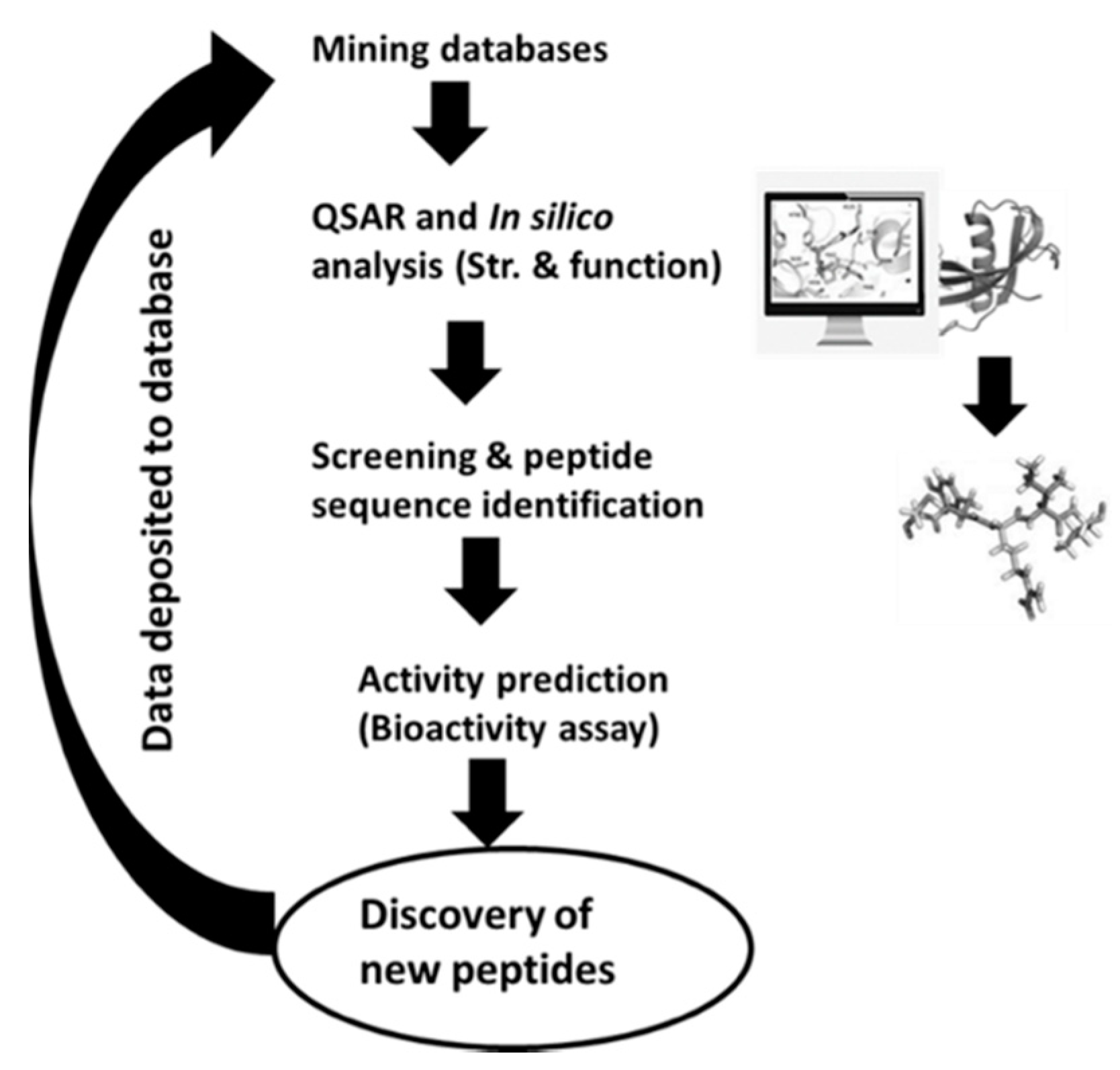
3.2. The In Silico Approach
3.3. Chemical Synthesis Approach
3.4. The Integrated Approach
3.5. Screening for Bioactive Peptides
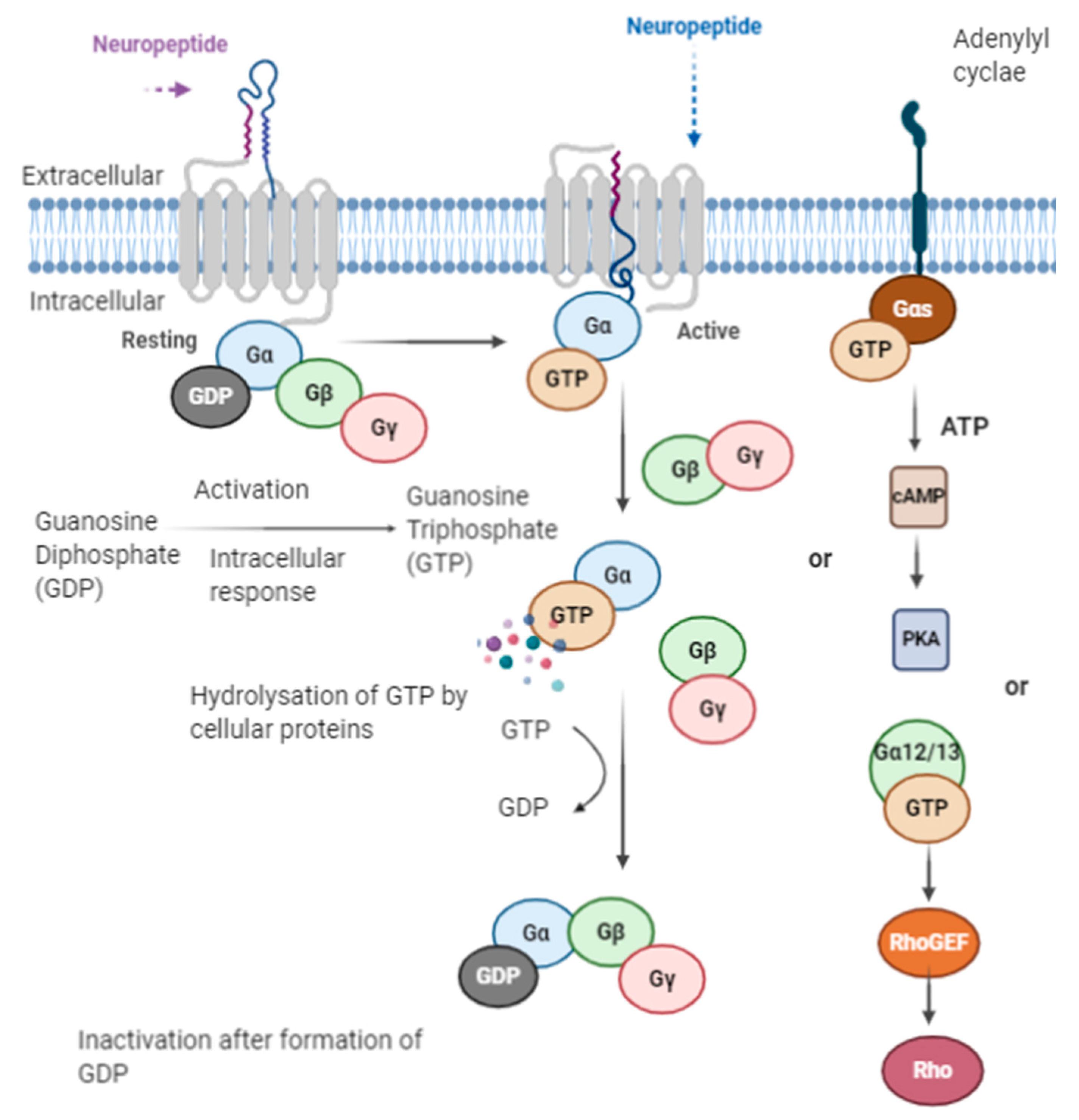
4. Mechanism of Action
Transport of Opioid Peptides in the Body
5. Clinical and Animal Studies Related to Exogenous Opioid Peptides in Stress-Related Conditions
Stress, Anxiety, and Depression
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization. World Mental Health Day, 10 October 2012. Available online: http://www.who.int/mental_health/management (accessed on 30 September 2020).
- Mills, K.T.; Stefanescu, A.; He, J. The global epidemiology of hypertension. Nat. Rev. Nephrol. 2020, 16, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Szelenberger, W.; Soldatos, C. Sleep disorders in psychiatric practice. World Psychiatry 2005, 4, 186. [Google Scholar] [PubMed]
- Substance Abuse and Mental Health Services Administration. Mental Health Services Administration. Key Substance Use and Mental Health Indicators in the United States: Results from the 2016 National Survey on Drug Use and Health (HHS Publication No. SMA 17-5044, NSDUH Series H-52); Center for Behavioral Health Statistics and Quality: Rockville, MD, USA, 2017.
- Ned, H.; Kalin, M.D. The Critical Relationship Between Anxiety and Depression. Am. J. Psychiatry 2020, 177, 365–367. [Google Scholar] [CrossRef]
- Kessler, R.C.; Sampson, N.A.; Berglund, P.; Gruber, M.; Al-Hamzawi, A.; Andrade, L.; Bunting, B.; Demyttenaere, K.; Florescu, S.; De Girolamo, G. Anxious and non-anxious major depressive disorder in the World Health Organization World Mental Health Surveys. Epidemiol. Psychiatr. Sci. 2015, 24, 210–226. [Google Scholar] [CrossRef]
- Gee, D.G.; Kribakaran, S. Developmental Differences in Neural Responding to Threat and Safety: Implications for Treating Youths with Anxiety. Am. Psychiatr. Assoc. 2020. [Google Scholar] [CrossRef] [PubMed]
- Poppitt, S.D. Milk proteins and human health. In Milk Proteins; Elsevier: Amsterdam, The Netherlands, 2020; pp. 651–669. [Google Scholar]
- González, S. Dietary Bioactive Compounds and Human Health and Disease. Nutrients 2020, 12, 348. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Lee, B.H.; Oh, D.H. Current trends and perspectives of bioactive peptides. Crit. Rev. Food Sci. Nutr. 2018, 58, 2273–2284. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Oh, D.H.; Lee, B.H. Bioactive peptides. Foods 2017, 6, 32. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Lee, B.H.; Park, M.H.; Kim, J.-H.; Oh, D.-H. Novel angiotensin I-converting enzyme inhibitory peptides from soybean protein isolates fermented by Pediococcus pentosaceus SDL1409. LWT 2018, 93, 88–93. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Ofosu, F.K.; Chelliah, R.; Park, M.H.; Kim, J.-H.; Oh, D.-H. Development of a soy protein hydrolysate with an antihypertensive effect. Int. J. Mol. Sci. 2019, 20, 1496. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Lee, B.H.; Park, B.-J.; Kim, S.-H.; Oh, D.-H. Antihypertensive peptides from whey proteins fermented by lactic acid bacteria. Food Sci. Biotechnol. 2018, 27, 1781–1789. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Villaluenga, C.; Hernández-Ledesma, B. Peptides for Health Benefits 2019. Int. J. Mol. Sci. 2020, 21, 2543. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.; Smith, T.; Kosterlitz, H.; Fothergill, L.A.; Morgan, B.; Morris, H. Identification of two related pentapeptides from the brain with potent opiate agonist activity. Nature 1975, 258, 577–579. [Google Scholar] [CrossRef] [PubMed]
- Gulpinar, M.A.; Yegen, B.C. The Physiology of Learning and Memory: Role of Peptides and Stress. Curr. Protein Pept. Sci. 2004, 5, 457–473. [Google Scholar] [CrossRef]
- Wang, Y.; Van Bockstaele, E.J.; Liu-Chen, L.-Y. In vivo trafficking of endogenous opioid receptors. Life Sci. 2008, 83, 693–699. [Google Scholar] [CrossRef]
- Rutherfurd-Markwick, K.J. Food proteins as a source of bioactive peptides with diverse functions. Br. J. Nutr. 2012, 108, S149–S157. [Google Scholar] [CrossRef]
- Bodnar, R.J. Endogenous opiates and behavior: 2012. Peptides 2013, 50, 55–95. [Google Scholar] [CrossRef]
- Bagley, E.E.; Ingram, S.L. Endogenous opioid peptides in the descending pain modulatory circuit. Neuropharmacology 2020, 173, 108131. [Google Scholar] [CrossRef]
- Teschemacher, H.; Koch, G.; Brantl, V. Milk protein-derived opioid receptor ligands. Pept. Sci. 1997, 43, 99–117. [Google Scholar] [CrossRef]
- Meisel, H.; Fitzgerald, R.J. Opioid peptides encrypted in intact milk protein sequences. Br. J. Nutr. 2000, 84, 27–31. [Google Scholar] [CrossRef]
- Carrasco-Castilla, J.; Hernández-Álvarez, A.J.; Jiménez-Martínez, C.; Gutiérrez-López, G.F.; Dávila-Ortiz, G. Use of proteomics and peptidomics methods in food bioactive peptide science and engineering. Food Eng. Rev. 2012, 4, 224–243. [Google Scholar] [CrossRef]
- Trivedi, M.S.; Shah, J.S.; Al-Mughairy, S.; Hodgson, N.W.; Simms, B.; Trooskens, G.A.; Van Criekinge, W.; Deth, R.C. Food-derived opioid peptides inhibit cysteine uptake with redox and epigenetic consequences. J. Nutr. Biochem. 2014, 25, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Bhat, Z.; Kumar, S.; Bhat, H.F. Bioactive peptides of animal origin: A review. J. Food Sci. Technol. 2015, 52, 5377–5392. [Google Scholar] [CrossRef] [PubMed]
- Koyama, M.; Hattori, S.; Amano, Y.; Watanabe, M.; Nakamura, K. Blood pressure-lowering peptides from neo-fermented buckwheat sprouts: A new approach to estimating ACE-inhibitory activity. PLoS ONE 2014, 9, e105802. [Google Scholar] [CrossRef]
- Popik, P.; Bobula, B.; Janusz, M.; Lisowski, J.; Vetulani, J. Colostrinin, a polypeptide isolated from early milk, facilitates learning and memory in rats. Pharmacol. Biochem. Behav. 1999, 64, 183–189. [Google Scholar] [CrossRef]
- Gülpınar, M.A.; Özbeyli, D.; Arbak, S.; Yeğen, B.Ç. Anti-inflammatory effect of acute stress on experimental colitis is mediated by cholecystokinin-B receptors. Life Sci. 2004, 75, 77–91. [Google Scholar] [CrossRef]
- Bracci-Laudiero, L.; Aloe, L.; Lundeberg, T.; Theodorsson, E.; Stenfors, C. Altered levels of neuropeptides characterize the brain of lupus prone mice. Neurosci. Lett. 1999, 275, 57–60. [Google Scholar] [CrossRef]
- Sprick, U.; Hasenöhrl, R.; Krauth, J.; Klapdor, K.; Huston, J. Effects of chronic substance P treatment and intracranial fetal grafts on learning after hippocampal kainic acid lesions. Peptides 1996, 17, 275–285. [Google Scholar] [CrossRef]
- Sandin, J.; Georgieva, J.; Schött, P.A.; Ögren, S.O.; Terenius, L. Nociceptin/orphanin FQ microinjected into hippocampus impairs spatial learning in rats. Eur. J. Neurosci. 1997, 9, 194–197. [Google Scholar] [CrossRef]
- Manabe, T.; Noda, Y.; Mamiya, T.; Katagiri, H.; Houtani, T.; Nishi, M.; Noda, T.; Takahashi, T.; Sugimoto, T.; Nabeshima, T. Facilitation of long-term potentiation and memory in mice lacking nociceptin receptors. Nature 1998, 394, 577–581. [Google Scholar] [CrossRef]
- Hiramatsu, M.; Inoue, K. Improvement by low doses of nociceptin on scopolamine-induced impairment of learning and/or memory. Eur. J. Pharmacol. 2000, 395, 149–156. [Google Scholar] [CrossRef]
- Lynch, G.; Larson, J.; Staubli, U.; Granger, R. Variants of Synaptic Potentiation and Different Types of Memory Operations in Hippocampus and Related Structures; Oxford University Press: Washington, DC, USA, 1991. [Google Scholar]
- Chen, Q.; Patel, R.; Sales, A.; Oji, G.; Kim, J.; Monreal, A.; Brinton, R. Vasopressin-induced neurotrophism in cultured neurons of the cerebral cortex: Dependency on calcium signaling and protein kinase C activity. Neuroscience 2000, 101, 19–26. [Google Scholar] [CrossRef]
- Crawley, J.N.; Corwin, R.L. Biological actions of cholecystokinin. Peptides 1994, 15, 731–755. [Google Scholar] [CrossRef]
- Dauge, V.; Pophillat, M.; Crete, D.; Melik-Parsadaniantz, S.; Roques, B. Involvement of brain endogenous cholecystokinin in stress-induced impairment of spatial recognition memory. Neuroscience 2003, 118, 19–23. [Google Scholar] [CrossRef]
- Telegdy, G.; Adamik, A.; Glover, V. The action of isatin (2, 3-dioxoindole) an endogenous indole on brain natriuretic and C-type natriuretic peptide-induced facilitation of memory consolidation in passive-avoidance learning in rats. Brain Res. Bull. 2000, 53, 367–370. [Google Scholar] [CrossRef]
- Telegdy, G.; Kokavszky, K.; Nyerges, A. Action of C-type natriuretic peptide (CNP) on passive avoidance learning in rats: Involvement of transmitters. Eur. J. Neurosci. 1999, 11, 3302–3306. [Google Scholar] [CrossRef]
- Telegdy, G.; Kokavszky, K. The action of pituitary adenylate cyclase activating polypeptide (PACAP) on passive avoidance learning. The role of transmitters. Brain Res. 2000, 874, 194–199. [Google Scholar] [CrossRef]
- McDONALD, M.P.; Gleason, T.C.; Robinson, J.K.; Crawley, J.N. Galanin inhibits performance on rodent memory tasks. Ann. N. Y. Acad. Sci. 1998, 863, 305–322. [Google Scholar] [CrossRef]
- Counts, S.E.; Perez, S.E.; Kahl, U.; Bartfai, T.; Bowser, R.P.; Deecher, D.C.; Mash, D.C.; Crawley, J.N.; Mufson, E.J. Galanin: Neurobiologic mechanisms and therapeutic potential for Alzheimer’s disease. CNS Drug Rev. 2001, 7, 445–470. [Google Scholar] [CrossRef]
- Wrenn, C.; Marriott, L.; Kinney, J.; Holmes, A.; Wenk, G.; Crawley, J. Galanin peptide levels in hippocampus and cortex of galanin-overexpressing transgenic mice evaluated for cognitive performance. Neuropeptides 2002, 36, 413–426. [Google Scholar] [CrossRef]
- Santo-Yamada, Y.; Yamada, K.; Wada, E.; Goto, Y.-I.; Wada, K. Blockade of bombesin-like peptide receptors impairs inhibitory avoidance learning in mice. Neurosci. Lett. 2003, 340, 65–68. [Google Scholar] [CrossRef]
- Yamada, K.; Santo-Yamada, Y.; Wada, K. Stress-induced impairment of inhibitory avoidance learning in female neuromedin B receptor-deficient mice. Physiol. Behav. 2003, 78, 303–309. [Google Scholar] [CrossRef]
- Ojika, K.; Mitake, S.; Tohdoh, N.; Appel, S.H.; Otsuka, Y.; Katada, E.; Matsukawa, N. Hippocampal cholinergic neurostimulating peptides (HCNP). Prog. Neurobiol. 2000, 60, 37–83. [Google Scholar] [CrossRef]
- Gasparini, L.; Netzer, W.J.; Greengard, P.; Xu, H. Does insulin dysfunction play a role in Alzheimer’s disease? Trends Pharmacol. Sci. 2002, 23, 288–293. [Google Scholar] [CrossRef]
- Moriguchi, T.; Sakurai, T.; Nambu, T.; Yanagisawa, M.; Goto, K. Neurons containing orexin in the lateral hypothalamic area of the adult rat brain are activated by insulin-induced acute hypoglycemia. Neurosci. Lett. 1999, 264, 101–104. [Google Scholar] [CrossRef]
- Aou, S.; Li, X.-L.; Li, A.-J.; Oomura, Y.; Shiraishi, T.; Sasaki, K.; Imamura, T.; Wayner, M. Orexin-A (hypocretin-1) impairs Morris water maze performance and CA1-Schaffer collateral long-term potentiation in rats. Neuroscience 2003, 119, 1221–1228. [Google Scholar] [CrossRef]
- Janecka, A.; Fichna, J.; Janecki, T. Opioid receptors and their ligands. Curr. Top. Med. Chem. 2004, 4, 1–17. [Google Scholar] [CrossRef]
- Koneru, A.; Satyanarayana, S.; Rizwan, S. Endogenous opioids: Their physiological role and receptors. Glob. J. Pharm. 2009, 3, 149–153. [Google Scholar]
- Law, P.-Y.; Wong, Y.H.; Loh, H.H. Mutational analysis of the structure and function of opioid receptors. Pept. Sci. 1999, 51, 440–455. [Google Scholar] [CrossRef]
- Yang, Y.-R.; Chiu, T.-H.; Chen, C.-L. Structure–activity relationships of naturally occurring and synthetic opioid tetrapeptides acting on locus coeruleus neurons. Eur. J. Pharmacol. 1999, 372, 229–236. [Google Scholar] [CrossRef]
- Kaur, J.; Kumar, V.; Sharma, K.; Kaur, S.; Gat, Y.; Goyal, A.; Tanwar, B. Opioid Peptides: An Overview of Functional Significance. Int. J. Pept. Res. Ther. 2020, 26, 33–41. [Google Scholar] [CrossRef]
- Froehlich, J.C. Opioid peptides. Alcohol Health Res. World 1997, 21, 132. [Google Scholar] [PubMed]
- Garg, S.; Nurgali, K.; Kumar Mishra, V. Food proteins as source of opioid peptides-a review. Curr. Med. Chem. 2016, 23, 893–910. [Google Scholar] [CrossRef] [PubMed]
- Hackler, L.; Zadina, J.E.; Ge, L.-J.; Kastin, A.J. Isolation of relatively large amounts of endomorphin-1 and endomorphin-2 from human brain cortex. Peptides 1997, 18, 1635–1639. [Google Scholar] [CrossRef]
- Aldrich, J.V.; Kulkarni, S.S.; Senadheera, S.N.; Ross, N.C.; Reilley, K.J.; Eans, S.O.; Ganno, M.L.; Murray, T.F.; McLaughlin, J.P. Unexpected Opioid Activity Profiles of Analogues of the Novel Peptide Kappa Opioid Receptor Ligand CJ-15,208. ChemMedChem 2011, 6, 1739–1745. [Google Scholar] [CrossRef]
- Goldstein, A. Opioid peptides (endorphins) in pituitary and brain. Science 1976, 193, 1081–1086. [Google Scholar] [CrossRef]
- Li, C.H.; Chung, D. Isolation and structure of an untriakontapeptide with opiate activity from camel pituitary glands. Proc. Natl. Acad. Sci. USA 1976, 73, 1145–1148. [Google Scholar] [CrossRef]
- Goldstein, A.; Fischli, W.; Lowney, L.I.; Hunkapiller, M.; Hood, L. Porcine pituitary dynorphin: Complete amino acid sequence of the biologically active heptadecapeptide. Proc. Natl. Acad. Sci. USA 1981, 78, 7219–7223. [Google Scholar] [CrossRef]
- Chavkin, C.; James, I.F.; Goldstein, A. Dynorphin is a specific endogenous ligand of the kappa opioid receptor. Science 1982, 215, 413–415. [Google Scholar] [CrossRef]
- Okada, Y.; Fukumizu, A.; Takahashi, M.; Shimizu, Y.; Tsuda, Y.; Yokoi, T.; Bryant, S.D.; Lazarus, L.H. Synthesis of stereoisomeric analogues of endomorphin-2, H-Tyr-Pro-Phe-Phe-NH2, and examination of their opioid receptor binding activities and solution conformation. Biochem. Biophys. Res. Commun. 2000, 276, 7–11. [Google Scholar] [CrossRef]
- Li, T.; Fujita, Y.; Tsuda, Y.; Miyazaki, A.; Ambo, A.; Sasaki, Y.; Jinsmaa, Y.; Bryant, S.D.; Lazarus, L.H.; Okada, Y. Development of Potent μ-Opioid Receptor Ligands Using Unique Tyrosine Analogues of Endomorphin-2. J. Med. Chem. 2005, 48, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Torino, D.; Mollica, A.; Pinnen, F.; Lucente, G.; Feliciani, F.; Davis, P.; Lai, J.; Ma, S.-W.; Porreca, F.; Hruby, V.J. Synthesis and evaluation of new endomorphin analogues modified at the Pro2 residue. Bioorgan. Med. Chem. Lett. 2009, 19, 4115–4118. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Murray, T.F.; Aldrich, J.V. Synthesis and evaluation of potential affinity labels derived from endomorphin-2. J. Pept. Res. 2003, 61, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Tömböly, C.; Kövér, K.E.; Péter, A.; Tourwé, D.; Biyashev, D.; Benyhe, S.; Borsodi, A.; Al-Khrasani, M.; Rónai, A.Z.; Tóth, G. Structure−Activity Study on the Phe Side Chain Arrangement of Endomorphins Using Conformationally Constrained Analogues. J. Med. Chem. 2004, 47, 735–743. [Google Scholar] [CrossRef]
- Honda, T.; Shirasu, N.; Isozaki, K.; Kawano, M.; Shigehiro, D.; Chuman, Y.; Fujita, T.; Nose, T.; Shimohigashi, Y. Differential receptor binding characteristics of consecutive phenylalanines in μ-opioid specific peptide ligand endomorphin-2. Bioorgan. Med. Chem. 2007, 15, 3883–3888. [Google Scholar] [CrossRef]
- Sasaki, Y.; Sasaki, A.; Niizuma, H.; Goto, H.; Ambo, A. Endomorphin 2 analogues containing Dmp residue as an aromatic amino acid surrogate with high μ-opioid receptor affinity and selectivity. Bioorgan. Med. Chem. 2003, 11, 675–678. [Google Scholar] [CrossRef]
- Vázquez, M.E.; Blanco, J.B.; Salvadori, S.; Trapella, C.; Argazzi, R.; Bryant, S.D.; Jinsmaa, Y.; Lazarus, L.H.; Negri, L.; Giannini, E. 6-N, N-Dimethylamino-2, 3-naphthalimide: A new environment-sensitive fluorescent probe in δ-and μ-selective opioid peptides. J. Med. Chem. 2006, 49, 3653–3658. [Google Scholar] [CrossRef]
- Yu, Y.; Shao, X.; Cui, Y.; Liu, H.M.; Wang, C.L.; Fan, Y.Z.; Liu, J.; Dong, S.L.; Cui, Y.X.; Wang, R. Structure–activity study on the spatial arrangement of the third aromatic ring of endomorphins 1 and 2 using an atypical constrained C terminus. ChemMedChem 2007, 2, 309–317. [Google Scholar] [CrossRef]
- Liu, W.X.; Wang, R. Endomorphins: Potential roles and therapeutic indications in the development of opioid peptide analgesic drugs. Med. Res. Rev. 2012, 32, 536–580. [Google Scholar] [CrossRef]
- Cardillo, G.; Gentilucci, L.; Tolomelli, A. Unusual amino acids: Synthesis and introduction into naturally occurring peptides and biologically active analogues. Mini Rev. Med. Chem. 2006, 6, 293–304. [Google Scholar] [CrossRef]
- Gentilucci, L. SNew Trends in the Development of Opioid Peptide Analogues as Advanced Remedies for Pain Relief. Curr. Top. Med. Chem. 2004, 4, 19–38. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Udenigwe, C.C. Role of food-derived opioid peptides in the central nervous and gastrointestinal systems. J. Food Biochem. 2019, 43, e12629. [Google Scholar] [CrossRef] [PubMed]
- Kastin, A. Handbook of Biologically Active Peptides; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Yoshikawa, M.; Takahashi, M.; Yang, S. Delta opioid peptides derived from plant proteins. Curr. Pharm. Des. 2003, 9, 1325–1330. [Google Scholar] [CrossRef] [PubMed]
- Zioudrou, C.; Streaty, R.A.; Klee, W.A. Opioid peptides derived from food proteins. The exorphins. J. Biol. Chem. 1979, 254, 2446–2449. [Google Scholar]
- Bell, S.J.; Grochoski, G.T.; Clarke, A.J. Health implications of milk containing β-casein with the A2 genetic variant. Crit. Rev. Food Sci. Nutr. 2006, 46, 93–100. [Google Scholar] [CrossRef]
- Nguyen, D.D.; Johnson, S.K.; Busetti, F.; Solah, V.A. Formation and degradation of beta-casomorphins in dairy processing. Crit. Rev. Food Sci. Nutr. 2015, 55, 1955–1967. [Google Scholar] [CrossRef]
- El-Salam, M.A.; El-Shibiny, S. Bioactive peptides of buffalo, camel, goat, sheep, mare, and yak milks and milk products. Food Rev. Int. 2013, 29, 1–23. [Google Scholar] [CrossRef]
- Brantl, V.; Teschemacher, H. A material with opioid activity in bovine milk and milk products. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1979, 306, 301–304. [Google Scholar] [CrossRef]
- Hazum, E.; Sabatka, J.J.; Chang, K.-J.; Brent, D.A.; Findlay, J.; Cuatrecasas, P. Morphine in cow and human milk: Could dietary morphine constitute a ligand for specific morphine (mu) receptors? Science 1981, 213, 1010–1012. [Google Scholar] [CrossRef]
- Henschen, A.; Brantl, V.; Teschemacher, H.; Lottspeich, F. β-Casomorphins–Novel Opioid Peptides Derived from Bovine Casein–Isolation and Structure. In Endogenous and Exogenous Opiate Agonists and Antagonists; Elsevier: Amsterdam, The Netherlands, 1980; pp. 233–236. [Google Scholar]
- Loukas, S.; Varoucha, D.; Zioudrou, C.; Streaty, R.A.; Klee, W.A. Opioid activities and structures of alpha-casein-derived exorphins. Biochemistry 1983, 22, 4567–4573. [Google Scholar] [CrossRef]
- Brantl, V.; Teschemacher, H.; Bläsig, J.; Henschen, A.; Lottspeich, F. Opioid activities of β-casomorphins. Life Sci. 1981, 28, 1903–1909. [Google Scholar] [CrossRef]
- Koch, G.; Wiedemann, K.; Teschemacher, H. Opioid activities of human β-casomorphins. Naunyn-Schmiedeberg Arch. Pharmacol. 1985, 331, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Bouglé, D.; Bouhallab, S. Dietary bioactive peptides: Human studies. Crit. Rev. Food Sci. Nutr. 2017, 57, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Calvo, C.F.; Cesselin, F.; Gelman, M.; Glowinski, J. Identification of an opioid peptide secreted by rat embryonic mixed brain cells as a promoter of macrophage migration. Eur. J. Neurosci. 2000, 12, 2676–2684. [Google Scholar] [CrossRef] [PubMed]
- Lister, J.; Fletcher, P.J.; Nobrega, J.N.; Remington, G. Behavioral effects of food-derived opioid-like peptides in rodents: Implications for schizophrenia? Pharmacol. Biochem. Behav. 2015, 134, 70–78. [Google Scholar] [CrossRef]
- Pruimboom, L.; De Punder, K. The opioid effects of gluten exorphins: Asymptomatic celiac disease. J. HealthPopul. Nutr. 2015, 33, 24. [Google Scholar] [CrossRef] [PubMed]
- Ohinata, K.; Agui, S.; Yoshikawa, M. Soymorphins, novel μ opioid peptides derived from soy β-conglycinin β-subunit, have anxiolytic activities. Biosci. Biotechnol. Biochem. 2007, 71, 2618–2621. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Tani, F.; Yoshimura, T.; Chiba, H. Opioid peptides from milk proteins. Agric. Biol. Chem. 1986, 50, 2419–2421. [Google Scholar] [CrossRef]
- Brantl, V.; Pfeiffer, A.; Herz, A.; Henschen, A.; Lottspeich, F. Antinociceptive potencies of β-casomorphin analogs as compared to their affinities towards μ and δ opiate receptor sites in brain and periphery. Peptides 1982, 3, 793–797. [Google Scholar] [CrossRef]
- Tani, F.; Iio, K.; Chiba, H.; Yoshikawa, M. Isolation and characterization of opioid antagonist peptides derived from human lactoferrin. Agric. Biol. Chem. 1990, 54, 1803–1810. [Google Scholar]
- Sharma, S.K.; Klee, W.A.; Nirenberg, M. Dual regulation of adenylate cyclase accounts for narcotic dependence and tolerance. Proc. Natl. Acad. Sci. USA 1975, 72, 3092–3096. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.; Kosterlitz, H.; Leslie, F.M. Effect of morphine on adrenergic transmission in the mouse vas deferens. Assessment of agonist and antogonist potencies of narcotic analgesics. Br. J. Pharmacol. 1975, 53, 371. [Google Scholar] [CrossRef] [PubMed]
- Pert, C.B.; Pasternak, G.; Snyder, S.H. Opiate agonists and antagonists discriminated by receptor binding in brain. Science 1973, 182, 1359–1361. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Kawamura, Y.; Yoshikawa, M. Effect of rubiscolin, a δ opioid peptide derived from Rubisco, on memory consolidation. Peptides 2003, 24, 325–328. [Google Scholar] [CrossRef]
- Kaneko, K.; Lazarus, M.; Miyamoto, C.; Oishi, Y.; Nagata, N.; Yang, S.; Yoshikawa, M.; Aritake, K.; Furuyashiki, T.; Narumiya, S. Orally administered rubiscolin-6, a δ opioid peptide derived from Rubisco, stimulates food intake via leptomeningeal lipocallin-type prostaglandin D synthase in mice. Mol. Nutr. Food Res. 2012, 56, 1315–1323. [Google Scholar] [CrossRef]
- Miyazaki, Y.; Kaneko, K.; Iguchi, S.; Mizushige, T.; Kanamoto, R.; Yoshikawa, M.; Shimizu, T.; Ohinata, K. Orally administered δ opioid agonist peptide rubiscolin-6 stimulates food intake in aged mice with ghrelin resistance. Mol. Nutr. Food Res. 2014, 58, 2046–2052. [Google Scholar] [CrossRef]
- Hirata, H.; Sonoda, S.; Agui, S.; Yoshida, M.; Ohinata, K.; Yoshikawa, M. Rubiscolin-6, a δ opioid peptide derived from spinach Rubisco, has anxiolytic effect via activating σ1 and dopamine D1 receptors. Peptides 2007, 28, 1998–2003. [Google Scholar] [CrossRef]
- Kaneko, K.; Mizushige, T.; Miyazaki, Y.; Lazarus, M.; Urade, Y.; Yoshikawa, M.; Kanamoto, R.; Ohinata, K. δ-Opioid receptor activation stimulates normal diet intake but conversely suppresses high-fat diet intake in mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 306, R265–R272. [Google Scholar] [CrossRef]
- Schusdziarra, V.; Schick, A.; de la Fuente, A.; Specht, J.; Klier, M.; Brantl, V.; Pfeiffer, E.-F. Effect of β-casomorphins and analogs on insulin release in dogs. Endocrinology 1983, 112, 885–889. [Google Scholar] [CrossRef]
- Mihatsch, W.; Franz, A.; Kuhnt, B.; Högel, J.; Pohlandt, F. Hydrolysis of casein accelerates gastrointestinal transit via reduction of opioid receptor agonists released from casein in rats. Neonatology 2005, 87, 160–163. [Google Scholar] [CrossRef]
- Zoghbi, S.; Trompette, A.; Claustre, J.; Homsi, M.E.; Garzón, J.; Jourdan, G.; Scoazec, J.-Y.; Plaisancié, P. β-Casomorphin-7 regulates the secretion and expression of gastrointestinal mucins through a μ-opioid pathway. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G1105–G1113. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Miao, J.; Zhang, Y. Protective effect of β-casomorphin-7 on type 1 diabetes rats induced with streptozotocin. Peptides 2010, 31, 1725–1729. [Google Scholar] [CrossRef]
- Fanciulli, G.; Dettori, A.; Demontis, M.P.; Tomasi, P.A.; Anania, V.; Delitala, G. Gluten exorphin B5 stimulates prolactin secretion through opioid receptors located outside the blood-brain barrier. Life Sci. 2005, 76, 1713–1719. [Google Scholar] [CrossRef] [PubMed]
- Belyaeva, Y.A.; Dubynin, V.; Stovolosov, I.; Dobryakova, Y.V.; Bespalova, Z.D.; Kamenskii, A. Effects of acute and chronic administration of exorphin C on behavior and learning in white rat pups. Mosc. Univ. Biol. Sci. Bull. 2009, 64, 66–70. [Google Scholar] [CrossRef]
- Jinsmaa, Y.; Yoshikawa, M. Enzymatic release of neocasomorphin and β-casomorphin from bovine β-casein. Peptides 1999, 20, 957–962. [Google Scholar] [CrossRef]
- Fukudome, S.-I.; Yoshikawa, M. Opioid peptides derived from wheat gluten: Their isolation and characterization. Febs Lett. 1992, 296, 107–111. [Google Scholar] [CrossRef]
- Fukudome, S.-I.; Yoshikawa, M. Gluten exorphin C: A novel opioid peptide derived from wheat gluten. FEBS Lett. 1993, 316, 17–19. [Google Scholar] [CrossRef]
- Yang, S.; Yunden, J.; Sonoda, S.; Doyama, N.; Lipkowski, A.W.; Kawamura, Y.; Yoshikawa, M. Rubiscolin, a δ selective opioid peptide derived from plant Rubisco. FEBS Lett. 2001, 509, 213–217. [Google Scholar] [CrossRef]
- Matthies, H.; Stark, H.; Hartrodt, B.; Ruethrich, H.-L.; Spieler, H.-T.; Barth, A.; Neubert, K. Derivatives of β-casomorphins with high analgesic potency. Peptides 1984, 5, 463–470. [Google Scholar] [CrossRef]
- Sakaguchi, M.; Koseki, M.; Wakamatsu, M.; Matsumura, E. Effects of systemic administration of β-casomorphin-5 on learning and memory in mice. Eur. J. Pharmacol. 2006, 530, 81–87. [Google Scholar] [CrossRef]
- Grecksch, G.; Schweigert, C.; Matthies, H. Evidence for analgesic activity of β-casomorphin in rats. Neurosci. Lett. 1981, 27, 325–328. [Google Scholar] [CrossRef]
- Lin, L.; Umahara, M.; York, D.; Bray, G. β-Casomorphins stimulate and enterostatin inhibits the intake of dietary fat in rats. Peptides 1998, 19, 325–331. [Google Scholar] [CrossRef]
- Kaneko, K.; Iwasaki, M.; Yoshikawa, M.; Ohinata, K. Orally administered soymorphins, soy-derived opioid peptides, suppress feeding and intestinal transit via gut μ1-receptor coupled to 5-HT1A, D2, and GABAB systems. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G799–G805. [Google Scholar] [CrossRef]
- Chesnokova, E.; Saricheva, N.; Dubynin, V.; Kamenskij, A.; Kalikhevich, V.; Adermasova, Z. Behavioral effect of soymorphin-5-amide in rats. Mosc. Univ. Biol. Sci. Bull. 2014, 69, 103–107. [Google Scholar] [CrossRef]
- Boullin, D.; Crampton, R.; Heading, C.E.; Pelling, D. Intestinal absorption of dipeptides containing glycine, phenylalanine, proline, β-alanine or histidine in the rat. Clin. Sci. Mol. Med. 1973, 45, 849–858. [Google Scholar] [CrossRef]
- Matthews, D. Intestinal absorption of peptides. Physiol. Rev. 1975, 55, 537–608. [Google Scholar] [CrossRef]
- Gardner, M. Absorption of amino acids and peptides from a complex mixture in the isolated small intestine of the rat. J. Physiol. 1975, 253, 233–256. [Google Scholar] [CrossRef]
- Zaloga, G.P.; Siddiqui, R.A. Biologically active dietary peptides. Mini Rev. Med. Chem. 2004, 4, 815–821. [Google Scholar] [CrossRef]
- Kaspar, A.A.; Reichert, J.M. Future directions for peptide therapeutics development. Drug Discov. Today 2013, 18, 807–817. [Google Scholar] [CrossRef]
- Houston, M.E.; Campbell, A.P.; Lix, B.; Kay, C.M.; Sykes, B.D.; Hodges, R.S. Lactam bridge stabilization of α-helices: The role of hydrophobicity in controlling dimeric versus monomeric α-helices. Biochemistry 1996, 35, 10041–10050. [Google Scholar] [CrossRef]
- Timmerman, P.; Puijk, W.C.; Meloen, R.H. Functional reconstruction and synthetic mimicry of a conformational epitope using CLIPS™ technology. J. Mol. Recognit. Interdiscip. J. 2007, 20, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.; Kim, Y.; Kim, T.; Lim, S.; Lee, M. Directional assembly of α-helical peptides induced by cyclization. J. Am. Chem. Soc. 2012, 134, 20270–20272. [Google Scholar] [CrossRef]
- Bougatef, A.; Nedjar-Arroume, N.; Ravallec-Plé, R.; Leroy, Y.; Guillochon, D.; Barkia, A.; Nasri, M. Angiotensin I-converting enzyme (ACE) inhibitory activities of sardinelle (Sardinella aurita) by-products protein hydrolysates obtained by treatment with microbial and visceral fish serine proteases. Food Chem. 2008, 111, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, Y.; Zhong, Q.; Wu, Y.; Xia, W. Purification and characterization of a novel angiotensin-I converting enzyme (ACE) inhibitory peptide derived from enzymatic hydrolysate of grass carp protein. Peptides 2012, 33, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Silvestre, M.P.C.; Silva, M.R.; Silva, V.D.M.; Souza, M.W.S.d.; Junior, L.; de Oliveira, C.; Afonso, W.d.O. Analysis of whey protein hydrolysates: Peptide profile and ACE inhibitory activity. Braz. J. Pharm. Sci. 2012, 48, 747–757. [Google Scholar] [CrossRef]
- Darewicz, M.; Borawska, J.; Vegarud, G.E.; Minkiewicz, P.; Iwaniak, A. Angiotensin I-converting enzyme (ACE) inhibitory activity and ACE inhibitory peptides of salmon (Salmo salar) protein hydrolysates obtained by human and porcine gastrointestinal enzymes. Int. J. Mol. Sci. 2014, 15, 14077–14101. [Google Scholar] [CrossRef]
- Nakamura, K.; Naramoto, K.; Koyama, M. Blood-pressure-lowering effect of fermented buckwheat sprouts in spontaneously hypertensive rats. J. Funct. Foods 2013, 5, 406–415. [Google Scholar] [CrossRef]
- García-Tejedor, A.; Saánchez-Rivera, L.; Castelló-Ruiz, M.; Recio, I.; Salom, J.B.; Manzanares, P. Novel antihypertensive lactoferrin-derived peptides produced by Kluyveromyces marxianus: Gastrointestinal stability profile and in vivo angiotensin I-converting enzyme (ACE) inhibition. J. Agric. Food Chem. 2014, 62, 1609–1616. [Google Scholar] [CrossRef]
- Ha, G.E.; Chang, O.K.; Jo, S.-M.; Han, G.-S.; Park, B.-Y.; Ham, J.-S.; Jeong, S.-G. Identification of antihypertensive peptides derived from low molecular weight casein hydrolysates generated during fermentation by Bifidobacterium longum KACC 91563. Korean J. Food Sci. Anim. Resour. 2015, 35, 738. [Google Scholar] [CrossRef]
- Jarmołowska, B.; Kostyra, E.; Krawczuk, S.; Kostyra, H. β-Casomorphin-7 isolated from Brie cheese. J. Sci. Food Agric. 1999, 79, 1788–1792. [Google Scholar] [CrossRef]
- Stuknytė, M.; Maggioni, M.; Cattaneo, S.; De Luca, P.; Fiorilli, A.; Ferraretto, A.; De Noni, I. Release of wheat gluten exorphins A5 and C5 during in vitro gastrointestinal digestion of bread and pasta and their absorption through an in vitro model of intestinal epithelium. Food Res. Int. 2015, 72, 208–214. [Google Scholar] [CrossRef]
- De Noni, I.; Cattaneo, S. Occurrence of β-casomorphins 5 and 7 in commercial dairy products and in their digests following in vitro simulated gastro-intestinal digestion. Food Chem. 2010, 119, 560–566. [Google Scholar] [CrossRef]
- Coda, R.; Rizzello, C.G.; Pinto, D.; Gobbetti, M. Selected lactic acid bacteria synthesize antioxidant peptides during sourdough fermentation of cereal flours. Appl. Environ. Microbiol. 2012, 78, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Zou, Y.; Yu, N.; Gu, Z. Accumulation and identification of angiotensin-converting enzyme inhibitory peptides from wheat germ. J. Agric. Food Chem. 2011, 59, 3598–3605. [Google Scholar] [CrossRef] [PubMed]
- Farvin, K.S.; Baron, C.P.; Nielsen, N.S.; Otte, J.; Jacobsen, C. Antioxidant activity of yoghurt peptides: Part 2–characterisation of peptide fractions. Food Chem. 2010, 123, 1090–1097. [Google Scholar] [CrossRef]
- Pripp, A.H.; Isaksson, T.; Stepaniak, L.; Sørhaug, T. Quantitative structure-activity relationship modelling of ACE-inhibitory peptides derived from milk proteins. Eur. Food Res. Technol. 2004, 219, 579–583. [Google Scholar] [CrossRef]
- Wu, C.-H.; Kuo, Y.-H.; Hong, R.-L.; Wu, H.-C. α-Enolase–binding peptide enhances drug delivery efficiency and therapeutic efficacy against colorectal cancer. Sci. Transl. Med. 2015, 7, 290ra291. [Google Scholar] [CrossRef]
- Wu, C.-H.; Liu, I.-J.; Lu, R.-M.; Wu, H.-C. Advancement and applications of peptide phage display technology in biomedical science. J. Biomed. Sci. 2016, 23, 8. [Google Scholar] [CrossRef]
- Cavazos, A.; Gonzalez de Mejia, E. Identification of bioactive peptides from cereal storage proteins and their potential role in prevention of chronic diseases. Compr. Rev. Food Sci. Food Saf. 2013, 12, 364–380. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Gong, M.; Wu, S. In silico analysis of the large and small subunits of cereal RuBisCO as precursors of cryptic bioactive peptides. Process Biochem. 2013, 48, 1794–1799. [Google Scholar] [CrossRef]
- Lacroix, I.M.; Li-Chan, E.C. Evaluation of the potential of dietary proteins as precursors of dipeptidyl peptidase (DPP)-IV inhibitors by an in silico approach. J. Funct. Foods 2012, 4, 403–422. [Google Scholar] [CrossRef]
- Holton, T.A.; Pollastri, G.; Shields, D.C.; Mooney, C. CPPpred: Prediction of cell penetrating peptides. Bioinformatics 2013, 29, 3094–3096. [Google Scholar] [CrossRef] [PubMed]
- Chandrudu, S.; Simerska, P.; Toth, I. Chemical methods for peptide and protein production. Molecules 2013, 18, 4373–4388. [Google Scholar] [CrossRef] [PubMed]
- Nishiuchi, Y.; Inui, T.; Nishio, H.; BOdi, J.; Kimura, T.; Tsuji, F.I.; Sakakibara, S. Chemical synthesis of the precursor molecule of the Aequorea green fluorescent protein, subsequent folding, and development of fluorescence. Proc. Natl. Acad. Sci. USA 1998, 95, 13549–13554. [Google Scholar] [CrossRef] [PubMed]
- Carpino, L.A.; Ghassemi, S.; Ionescu, D.; Ismail, M.; Sadat-Aalaee, D.; Truran, G.A.; Mansour, E.; Siwruk, G.A.; Eynon, J.S.; Morgan, B. Rapid, continuous solution-phase peptide synthesis: Application to peptides of pharmaceutical interest. Org. Process Res. Dev. 2003, 7, 28–37. [Google Scholar] [CrossRef]
- Murata, H.; Carmali, S.; Baker, S.L.; Matyjaszewski, K.; Russell, A.J. Solid-phase synthesis of protein-polymers on reversible immobilization supports. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Dirksen, A.; Meijer, E.; Adriaens, W.; Hackeng, T.M. Strategy for the synthesis of multivalent peptide-based nonsymmetric dendrimers by native chemical ligation. Chem. Commun. 2006, 1667–1669. [Google Scholar] [CrossRef]
- Lovrinovic, M.; Niemeyer, C.M. Microtiter plate-based screening for the optimization of DNA–protein conjugate synthesis by means of expressed protein ligation. ChemBioChem 2007, 8, 61–67. [Google Scholar] [CrossRef]
- Hartmann, R.; Meisel, H. Food-derived peptides with biological activity: From research to food applications. Curr. Opin. Biotechnol. 2007, 18, 163–169. [Google Scholar] [CrossRef]
- Meisel, H.; FitzGerald, R.J. Biofunctional peptides from milk proteins: Mineral binding and cytomodulatory effects. Curr. Pharm. Des. 2003, 9, 1289–1296. [Google Scholar]
- Mendis, E.; Rajapakse, N.; Byun, H.-G.; Kim, S.-K. Investigation of jumbo squid (Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects. Life Sci. 2005, 77, 2166–2178. [Google Scholar] [CrossRef] [PubMed]
- González-García, E.; Marina, M.L.; García, M.C. Plum (Prunus domestica L.) by-product as a new and cheap source of bioactive peptides: Extraction method and peptides characterization. J. Funct. Foods 2014, 11, 428–437. [Google Scholar] [CrossRef]
- Agyei, D.; Danquah, M.K. Industrial-scale manufacturing of pharmaceutical-grade bioactive peptides. Biotechnol. Adv. 2011, 29, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, S.-K. Bioactive peptides from marine sources as potential anti-inflammatory therapeutics. Curr. Protein Pept. Sci. 2013, 14, 177–182. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Chemometric analysis of the amino acid requirements of antioxidant food protein hydrolysates. Int. J. Mol. Sci. 2011, 12, 3148–3161. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Park, S.; Atwal, A.; Gibbs, B.; Lee, B. Angiotensin I-converting enzyme (ACE) inhibitory peptides from whey fermented by Lactobacillus species. J. Food Biochem. 2009, 33, 587–602. [Google Scholar] [CrossRef]
- Hou, P.; Zhao, G.; He, C.; Wang, H.; He, H. Biopanning of polypeptides binding to bovine ephemeral fever virus G 1 protein from phage display peptide library. BMC Vet. Res. 2018, 14, 3. [Google Scholar] [CrossRef]
- Suire, C.N.; Nainar, S.; Fazio, M.; Kreutzer, A.G.; Paymozd-Yazdi, T.; Topper, C.L.; Thompson, C.R.; Leissring, M.A. Peptidic inhibitors of insulin-degrading enzyme with potential for dermatological applications discovered via phage display. PLoS ONE 2018, 13, e0193101. [Google Scholar] [CrossRef]
- Khondee, S.; Piyawattanametha, W. Targeting Peptides Derived from Phage Display for Clinical Imaging. In Bacteriophages-Biology and Applications; IntechOpen: London, UK, 2019. [Google Scholar]
- Shen, Y.; Ruan, L.; Lian, C.; Li, R.; Tu, Z.; Liu, H. Discovery of HB-EGF binding peptides and their functional characterization in ovarian cancer cell lines. Cell Death Discov. 2019, 5, 1–11. [Google Scholar] [CrossRef]
- Christiansen, A.; Kringelum, J.V.; Hansen, C.S.; Bøgh, K.L.; Sullivan, E.; Patel, J.; Rigby, N.M.; Eiwegger, T.; Szépfalusi, Z.; De Masi, F. High-throughput sequencing enhanced phage display enables the identification of patient-specific epitope motifs in serum. Sci. Rep. 2015, 5, 12913. [Google Scholar] [CrossRef]
- Ramaraju, H.; Miller, S.J.; Kohn, D.H. Dual-functioning peptides discovered by phage display increase the magnitude and specificity of BMSC attachment to mineralized biomaterials. Biomaterials 2017, 134, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, F.; Luo, J.; Pang, J.; Yan, S.; Luo, F.; Liu, J.; Wang, W.; Cui, Y.; Su, X. A new non-muscle-invasive bladder tumor-homing peptide identified by phage display in vivo. Oncol. Rep. 2016, 36, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Fricker, L.D. Neuropeptides and other bioactive peptides: From discovery to function. In Colloquium Series on Neuropeptides; Morgan & Claypool Publishers: Williston, VT, USA, 2012. [Google Scholar]
- Law, P.-Y.; Wong, Y.H.; Loh, H.H. Molecular mechanisms and regulation of opioid receptor signaling. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 389–430. [Google Scholar] [CrossRef] [PubMed]
- De Noni, I.; FitzGerald, R.J.; Korhonen, H.J.; Le Roux, Y.; Livesey, C.T.; Thorsdottir, I.; Tomé, D.; Witkamp, R. Review of the potential health impact of β-casomorphins and related peptides. EFSA Sci. Rep. 2009, 231, 1–107. [Google Scholar]
- Ganapathy, V.; Miyauchi, S. Transport systems for opioid peptides in mammalian tissues. AAPS J. 2005, 7, E852–E856. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.-J. The enteric nervous system and gastrointestinal innervation: Integrated local and central control. In Microbial Endocrinology: The Microbiota-Gut-Brain Axis in Health and Disease; Springer: Berlin/Heidelberg, Germany, 2014; pp. 39–71. [Google Scholar]
- Brasnjevic, I.; Steinbusch, H.W.; Schmitz, C.; Martinez-Martinez, P.; Initiative, E.N.R. Delivery of peptide and protein drugs over the blood–brain barrier. Prog. Neurobiol. 2009, 87, 212–251. [Google Scholar] [CrossRef]
- Wang, J.; Hogenkamp, D.J.; Tran, M.; Li, W.-Y.; Yoshimura, R.F.; Johnstone, T.B.; Shen, W.-C.; Gee, K.W. Reversible lipidization for the oral delivery of leu-enkephalin. J. Drug Target. 2006, 14, 127–136. [Google Scholar] [CrossRef]
- Brugos, B.; Hochhaus, G. Metabolism of dynorphin A (1–13). Die Pharm.-Int. J. Pharm. Sci. 2004, 59, 339–343. [Google Scholar]
- Negri, L.; Lattanzi, R.; Tabacco, F.; Scolaro, B.; Rocchi, R. Glycodermorphins: Opioid peptides with potent and prolonged analgesic activity and enhanced blood-brain barrier penetration. Br. J. Pharmacol. 1998, 124, 1516–1522. [Google Scholar] [CrossRef]
- Kim, B.-J.; Zhou, J.; Martin, B.; Carlson, O.D.; Maudsley, S.; Greig, N.H.; Mattson, M.P.; Ladenheim, E.E.; Wustner, J.; Turner, A. Transferrin fusion technology: A novel approach to prolonging biological half-life of insulinotropic peptides. J. Pharmacol. Exp. Ther. 2010, 334, 682–692. [Google Scholar] [CrossRef]
- Dennis, M.S.; Zhang, M.; Meng, Y.G.; Kadkhodayan, M.; Kirchhofer, D.; Combs, D.; Damico, L.A. Albumin binding as a general strategy for improving the pharmacokinetics of proteins. J. Biol. Chem. 2002, 277, 35035–35043. [Google Scholar] [CrossRef] [PubMed]
- Egleton, R.D.; Mitchell, S.A.; Huber, J.D.; Janders, J.; Stropova, D.; Polt, R.; Yamamura, H.I.; Hruby, V.J.; Davis, T.P. Improved bioavailability to the brain of glycosylated Met-enkephalin analogs. Brain Res. 2000, 881, 37–46. [Google Scholar] [CrossRef]
- Perlikowska, R.; Gach, K.; Fichna, J.; Toth, G.; Walkowiak, B.; do-Rego, J.-C.; Janecka, A. Biological activity of endomorphin and [Dmt1]endomorphin analogs with six-membered proline surrogates in position 2. Bioorganic Med. Chem. 2009, 17, 3789–3794. [Google Scholar] [CrossRef] [PubMed]
- Tóth, G.; Keresztes, A.; Tömböly, C.; Péter, A.; Fülöp, F.; Tourwé, D.; Navratilova, E.; Varga, É.; Roeske, W.R.; Yamamura, H.I.; et al. New endomorphin analogs with mu-agonist and delta-antagonist properties. Pure Appl. Chem. 2004, 76, 951–957. [Google Scholar] [CrossRef][Green Version]
- Bali, A.; Singh, N.; Singh Jaggi, A. Neuropeptides as therapeutic targets to combat stress-associated behavioral and neuroendocrinological effects. CNS Neurol. Disord. Drug Targets (Former. Curr. Drug Targets-Cns Neurol. Disord.) 2014, 13, 347–368. [Google Scholar] [CrossRef]
- Clynen, E.; Swijsen, A.; Raijmakers, M.; Hoogland, G.; Rigo, J.-M. Neuropeptides as targets for the development of anticonvulsant drugs. Mol. Neurobiol. 2014, 50, 626–646. [Google Scholar] [CrossRef]
- Yimit, D.; Hoxur, P.; Amat, N.; Uchikawa, K.; Yamaguchi, N. Effects of soybean peptide on immune function, brain function, and neurochemistry in healthy volunteers. Nutrition 2012, 28, 154–159. [Google Scholar] [CrossRef]
- Bernet, F.; Montel, V.; Noël, B.; Dupouy, J.P. Diazepam-like effects of a fish protein hydrolysate (Gabolysat PC60) on stress responsiveness of the rat pituitary-adrenal system and sympathoadrenal activity. Psychopharmacology 2000, 149, 34–40. [Google Scholar] [CrossRef]
- Perlikowska, R.; Janecka, A. Rubiscolins-highly potent peptides derived from plant proteins. Mini Rev. Med. Chem. 2018, 18, 104–112. [Google Scholar] [CrossRef]
- Jahan-Mihan, A.; Luhovyy, B.L.; El Khoury, D.; Anderson, G.H. Dietary proteins as determinants of metabolic and physiologic functions of the gastrointestinal tract. Nutrients 2011, 3, 574–603. [Google Scholar] [CrossRef]
- Yoshikawa, M. Bioactive peptides derived from natural proteins with respect to diversity of their receptors and physiological effects. Peptides 2015, 72, 208–225. [Google Scholar] [CrossRef] [PubMed]
- Booij, L.; Merens, W.; Markus, C.R.; Van der Does, A.W. Diet rich in α-lactalbumin improves memory in unmedicated recovered depressed patients and matched controls. J. Psychopharmacol. 2006, 20, 526–535. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptides | Functions | References |
|---|---|---|
| Colostrinin | Promotes acquisition of spatial learning in aged rats | [28] |
| Corticotropin-releasing factor (CRF) | Improves memory retention Enhance learning performance | [17,29] |
| Neuropeptide Y (NPY) | Neuroprotection as by control of feeding works against neurodegenerative diseases | [17,30] |
| Substance P (SP) | Improves functional recovery and increases the learning ability | [17,31] |
| Nociceptin/orphanin FQ (N/OFQ) | Impairs spatial learning in animal models. Facilitate memory | [32,33,34] |
| Angiotensin-vasopressin (AVP) and Oxytocin (OT) | Promote social memory and learning behaviors. Deficiency of AVP results in memory impairment | [35,36] |
| Cholecystokinin (CCK) | CCK peptides improve learning and memory performance in the patients Help in anxiety states Lack of CCK-A receptors cause impaired learning and memory functions. Play a role in conditioned fear stress and anxiety | [29,37,38] |
| Atrial natriuretic peptide (ANP), Brain-derived natriuretic peptide (BNP), C-type natriuretic peptide (CNP) | Promote action on memory consolidation | [39,40] |
| Pituitary adenylate cyclase-activating polypeptide (PACAP) | Promote learning (consolidation and retrieval) | [41] |
| Galanin | Impairs the learning and memory performances overexpression impairs cognition | [42,43,44] |
| Bombesin/gastrin-releasing peptide (BN/GRP) and Neuromedin (NM) | Improve memory performance | [45,46] |
| Hippocampal cholinergic neurostimulation Peptide (HCNP) | Abnormal accumulation and expression associated with memory and learning disorders | [47] |
| Calcitonin-gene related peptide (CGRP), Substance P(SP) and Neuropeptide Y (NPY) | Enhance memory retention. SP improves functional recovery and increases learning ability. NPY enhances memory | [30] |
| Insulin | Improves short-term memory | [48] |
| Orexin-A | Inhibits long-term potentiation (LTP) and retards spatial learning | [49,50] |
| (A) | ||||
| Opioid Peptide | Amino-Acid Sequence | Protein Precursor | References | |
| endomorphin-1 | Tyr-Pro-Trp-Phe-NH2 | pro-endomorphin | [57,58] | |
| endomorphin-2 | Tyr-Pro-Phe-Phe-NH2 | pro-endomorphin | [57,58] | |
| met-enkephalin | Tyr-Gly-Gly-Phe-Met | pro-enkephalin | [16], | |
| leu-enkephalin | Tyr-Gly-Gly-Phe-Leu | pro-enkephalin | [16,59] | |
| β-endorphin | Tyr-Gly-Gly-Phe-Met-Thr-Ser-Glu-Lys-Ser-Gln-ThrPro-Leu-Val-Thr-Leu-Phe-Lys-Asn-Ala-Ile-Ile-LysAsn-Ala-Tyr-Lys-Lys-Gly-Glu | pro-opiomelanocortin | [60,61] | |
| dynorphin A | Tyr-Gly-Gly-Phe-Leu-Arg-Arg-Ile-Arg-Pro-Lys-LeuLys-Trp-Asp-Asn-Gln | pro-dynorphin | [62,63] | |
| dynorphin B | Phe-Gly-Gly-Phe-Thr-Gly-Ala-Arg-Lys-Ser-Ala-ArgLys-Leu-Ala-Asn-Gln | pronociceptin | [62,63] | |
(B) | ||||
| Sequence | IC50 (nM) | Ratio of IC50 Ratio δ/µ | References | |
| Affity (µ-Receptor) | Affity (δ-Receptor) | |||
| Endomorphins Modified at First Amino Acid Position | ||||
| d-Tyr1-Pro-Phe-Phe-NH2 | 32.1 ± 1.5 | 4121 ± 1492 | 128 | [64] |
| Dmt1-Pro-Trp-Phe-NH2 | 0.014 ± 0.003 | 12.0 ± 4.05 | 857 | [65] |
| Mmt1-Pro-Phe-Phe-NH2 | 0.132 ± 0.008 | 528.6 ± 47 | 4005 | [65] |
| Emt1-Pro-Phe-Phe NH2 | 0.063 ± 0.006 | 55.7 ± 6.2 | 884 | [65] |
| Dit1-Pro-Phe-Phe-NH2 | 2.29 ± 0.37 | 105 ± 16 | 46 | [65] |
| Det1-Pro-Phe-Phe-NH2 | 0.084 ± 0.006 | 69.7 ± 5.3 | 830 | [65] |
| Tmt1-Pro-Phe-Phe-NH2 | 1.111 ± 0.002 | 593.5 ± 80 | 5347 | [65] |
| Endomorphins Modified at Second Amino Acid Position | ||||
| Tyr-d-Pro2-Phe-Phe-NH2 | 512.4 ± 29 | 30,641 ± 419 | 60 | [64] |
| Tyr-Aze2-Trp-Phe-NH2 | 2.3 ± 0.23 | 3500 ± 360 | 1500 | [66] |
| Tyr-δAla2-Phe-Phe-NH2 | 34 ± 6.3 | 710 ± 130 | 21 | [66] |
| Tyr-3Aze2-Phe-Phe-NH2 | 210 ± 51 | 6900 ± 1200 | 32 | [66] |
| Tyr-Aze2-Phe-Phe-NH2 | 5.6 ± 1.2 | 5100 ± 600 | 920 | [66] |
| Endomorphins Modified at Third Amino Acid Position | ||||
| Tyr-Pro-d-Phe3-Phe-NH2 | 203.2 ± 83 | 4230 ± 344 | 21 | [64] |
| Tyr-Pro-Phe3-(p-NH2)-Phe-NH2 | 185 ± 36 | >10,000 | >1.9 | [67] |
| TyrProPhe3(p-NHCOCH2Br)-PheNH2 | 7210 ± 820 | >10,000 | >1.4 | [67] |
| Tyr-Pro-(2S,3R)-βMePhe3-Phe-NH2 | 106 ± 9 | >10,000 | >10 | [68] |
| Tyr-Pro-(2S,3S)-βMePhe3-Phe-NH2 | 45.3 ± 4.1 | 179 ± 15 | 4 | [68] |
| Tyr-Pro-(2R,3S)-βMePhe3-Phe-NH2 | 4910 ± 328 | >10,000 | >2 | [68] |
| Tyr-Pro-(2R,3R)-βMePhe3-Phe-NH2 | 7090 ± 131 | 6760 ± 865 | 1 | [68] |
| Tyr-Pro-(F5)-Phe3-Phe-NH2 | 11.7 ± 0.503 | 11,700 ± 1010 | 1000 | [69] |
| Endomorphins Modified at C-TERMINAL Position | ||||
| Tyr-Pro-Phe-d-Phe4-NH2 | 45.9 ± 8.6 | 8159 ± 1569 | 177 | [64] |
| Tyr-Pro-Phe-(p-NH2)-Phe4-NH2 | 36.7 ± 2.2 | >10,000 | >270 | [67] |
| TyrProPhePhe4(p-NHCOCH2Br)-NH2 | 158 ± 23 | 1940 ± 310 | 12 | [67] |
| Tyr-Pro-Phe-(p-NCS)-Phe4-NH2 | 345 ± 128 | >10,000 | >29 | [67] |
| Tyr-Pro-Trp-Dmp4-NH2 | 13.2 ± 1.9 | 7624 ± 2571 | 578 | [70] |
| Tyr-Pro-Trp-d-Dmp4-NH2 | 106 ± 20 | 1765 ± 834 | 17 | [70] |
| Tyr-Pro-Phe-Phe4-NH-(CH2)5-CODap(6DMN)-NH2 | 244.5 ± 14 | 5939 ± 1396 | 24 | [71] |
| Tyr-Pro-Phe-d-Val4-NH-Bn | 4.97 ± 1.24 | 3358 ± 414 | 676 | [72] |
| Tyr-Pro-Trp-d-Val4-NH-Bn | 2.32 ± 0.15 | 3287 ± 456 | 1417 | [72] |
| Source | Sequences | Peptide Name | References |
|---|---|---|---|
| Bovine milk β-casein | Tyr-Pro-Phe-Pro | βb-casomorphin-4 | [87] |
| Tyr-Pro-Phe-Pro-Gly | βb-casomorphin-5 | ||
| Tyr-Pro-Phe-Pro-Gly-Pro | βb-casomorphin-6 | ||
| Tyr-Pro-Phe-Pro-Gly-Pro-Ile | βb-casomorphin-7 | ||
| Tyr-Pro-Val-Glu-Pro-Phe | Neocasomorphin-6 | [111] | |
| Bovine milk α-lactalbumin | Tyr-Gly-Leu-Phe-NH2 | αb-lactorphin | [22] |
| Human milk β-casein | Tyr-Pro-Phe-Val | βh-casomorphin-4 | [22] |
| Tyr-Pro-Phe-Val-Glu | βh-casomorphin-5 | [57] | |
| Tyr-Pro-Phe-Val-Glu-Pro-Ile | βh-casomorphin-7 | [88] | |
| Tyr-Pro-Phe-Val-Glu-Pro-Ile-pro | βh-casomorphin-8 | [22,88] | |
| Human milk lactalbumin | Tyr-Gly-Leu-Phe-NH2 | αh-lactorphin | [22,94] |
| Bovine/bovine milk lactoferrin | Tyr-Leu-Gly-Ser-Gly-Tyr-OCH3 | lactoferrsoxin A | [96] |
| Arg-Tyr-Tyr-Gly-Tyr-OCH3 | lactoferrsoxin B | ||
| Lys-Tyr-Leu-Gly-Pro-Gln-Tyr-OCH3 | lactoferrsoxin C | ||
| Soy β-conglycinin | Tyr-Pro-Phe-Val-Val | Soymorphin-5 | [93] |
| Tyr-Pro-Phe-Val-Val-Asn | Soymorphin-6 | ||
| Tyr-Pro-Phe-Val-Val-Asn-Ala | Soymorphin-7 | ||
| Wheat HMW glutenin | Gly-Tyr-Tyr-Pro | gluten exorphin A4 | [78,112] |
| Gly-Tyr-Tyr-Pro-Thr | gluten exorphin A5 | ||
| Tyr-Gly-Gly-Trp | gluten exorphin B4 | ||
| Tyr-Gly-Gly-Trp-Leu | gluten exorphin B5 | ||
| Tyr-Pro-Ile-Ser-Leu | gluten exorphin C | [78,113] | |
| Spinach RuBisCo | Tyr-Pro-Leu-Asp-Leu | rubiscolin-5 | [93,114] |
| Tyr-Pro-Leu-Asp-Leu-Phe | rubiscolin-6 |
| Opioid Peptide | Opioid Activity (IC50 in µM) | µ/δ Ratio | Reference | |
|---|---|---|---|---|
| Mouse (vas Deferens) (δ) | Guinea-Pig (ileum) (µ) | |||
| rubiscolin-5 | 51 | 1110 | 21.8 | [114] |
| rubiscolin-6 | 24.4 | 748 | 30.7 | [114] |
| βb-casomorphin-4 | 84 | 22 | 0.26 | [87] |
| βb-casomorphin-5 | 40 | 6.5 | 0.16 | [87] |
| βb-casomorphin-6 | >150 | 27.4 | <0.18 | [87] |
| βb-casomorphin-7 | >200 | 57 | <0.29 | [87] |
| βh-casomorphin-4 | 750 | 19 | 0.025 | [94] |
| βh-casomorphin-5 | ND | 14 | ND | [94] |
| βh-casomorphin-6 | 350 | 25 | 0.071 | [94] |
| βh-casomorphin-8 | 540 | 25 | 0.047 | [94] |
| gluten-exorphin A4 | 70 | >1000 | ND | [112] |
| gluten exorphin A5 | 60 | 1000 | 60.7 | [112] |
| gluten exorphin B4 | 3.4 | 1.5 | 0.44 | [112] |
| gluten exorphin B5 | 0.017 | 0.05 | 2.9 | [112] |
| gluten exorphin C | 30 | 110 | 3.7 | [113] |
| soymorphin-5 | 50 | 6 | 0.12 | [93] |
| soymorphin-6 | 32 | 9.2 | 0.287 | [93] |
| soymorphin-7 | 50 | 13 | 0.26 | [93] |
| Human milk lactalbumin (α-lactorphin) | >1000 | 50 | ND | [94] |
| Bovine milk lactoferrin (lactoferrsoxin) | 4.38 | 5.68 | 0.77 | [96] |
| Opioid Peptide | Animal Model | Dosage | Administration Route | Effect | Time Duration | Reference |
|---|---|---|---|---|---|---|
| rubiscolin-5 | Mice | 3 nM/mouse | i.c.v | antinociception | Effects observed up to 30 min post-injection | [114] |
| rubiscolin-6 | Mice | 1 nM/mouse | i.c.v | |||
| rubiscolin–6 | Mice | 100 mg/kg 3 nM/mouse | Oral i.c.v | enhancement in memory consolidation | Effects observed up to 24 h post-injection | [100] |
| β-casomorphin- 4,-5,-6,-7 | Rat | 60–2000 nM | i.c.v | analgesic, naloxone reversible | Effects observed up to 30–40 min post-injection | [115] |
| β-casomorphin-5 | Mice Rat | 1mg/kg 166 nM | i.p i.v | improvement in learning and memory, analgestic | Effects observed up to 30-min post-injection Significant analgesia l0-min after injection up to 60 min post-injection | [116,117] |
| β-casomorphin-7 | Rat | 0.1–20 nM | i.c.v | food-intake stimulation | Effects observed up to 6 h post-injection | [118] |
| gluten exorphin C | Mice | 5mg/kg | i.p | improvement in learning and behavior, decreased anxiety | Effects observed up to 15–20 min post-injection | [110] |
| Gluten-exorphin B5 | Rat | 3 mg/kg | i.v | stimulated prolactin secretion | Effects observed after 20 min post-injection | [109] |
| soymorphin-5, 6, and 7 | Mice | 10–30 mg/kg or 3 mg/kg | Oral i.p | anxiolytic effect | Oral—Effects observed up to 20–25 min post-injection i.p—Effects observed <30min of post-injection | [93] |
| soymorphin-5 and 7 | Mice | 30 mg/kg or 48 µ mol/kg | oral | reduced food intake and showed anorexigenic activity | Effects observed after 2 h of oral administration | [119] |
| soymorphin-5 amide | Rat | 5 mg/kg | i.p | decreased anxiety | Effects observed after 30 min of administration | [120] |
| Databases Name | Address | Role |
|---|---|---|
| NeuroPIpred | https://webs.iiitd.edu.in/raghava/neuropipred | Neuropeptide database |
| NeuroPP | http://i.uestc.edu.cn/neuropeptide/neuropp/home.html | Neuropeptide database |
| BIOPEP (Bioactivity) (digestion) (Protein) (toxicity) | http://www.uwm.edu.pl/biochemia/index.php/en/biopep | Prediction for precursors of bioactive peptides, |
| ToxinPred (Toxicity) | http://crdd.osdd.net/raghava//toxinpred/ | Prediction of toxicity of peptides |
| I-TASSER (Protein Structure) | https://zhanglab.ccmb.med.umich.edu/I-TASSER/ | Structure and function prediction |
| NCBI (Protein Database) | https://www.ncbi.nlm.nih.gov/ | Protein sequences information |
| AlgPred (Toxicity) | http://crdd.osdd.net/raghava//algpred/ | Prediction of toxicity of peptides |
| ProtParam (phytochemical) | http://web.expasy.org/protparam/ | Compute GRAVY (grand average of hydropathicity) |
| UniProtKB (Protein database) | http://www.uniprot.org/ | Structure and sequences information |
| APD (Peptide database) | http://aps.unmc.edu/AP/main.html | Bioactive peptide prediction |
| AntiBP2 (Bioactivity prediction) | http://crdd.osdd.net/raghava//antibp2/ | Antibacterial peptide prediction |
| PEPstrMOD (Peptide database) | http://osddlinux.osdd.net/raghava/pepstrmod | Prediction of tertiary structures |
| Sequence | Half-Life (Mouse Brain) [min] | Reference |
|---|---|---|
| [d-Ala2, p-Cl-Phe4]EM-1 | >300 | [181] |
| [Dmt1,Nip2]EM-1 | 30.9 ± 3.29 | [182] |
| [(2S,3S)β-MePhe4]EM-2 | 35.8 ± 1.8 | [68] |
| [(1S,2R)ACHC2]EM-1 | >12 h | [183] |
| Guanidino-[d-Pro2Gly3, p-Cl-Phe5]EM-1 | 187.3 ± 24 | [181] |
| [(1S,2R)ACPC2]EM-2 | >12 h | [183] |
| [Dmt1,Nip2]EM-2 | 10.7 ± 0.3 | [182] |
| Guanidino-[d-Pro2-Gly3]EM-1 | 111.8 ± 19.2 | [181] |
| Guanidino-[Sar2]EM-1 | 43.9 ± 2.4 | [181] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tyagi, A.; Daliri, E.B.-M.; Kwami Ofosu, F.; Yeon, S.-J.; Oh, D.-H. Food-Derived Opioid Peptides in Human Health: A Review. Int. J. Mol. Sci. 2020, 21, 8825. https://doi.org/10.3390/ijms21228825
Tyagi A, Daliri EB-M, Kwami Ofosu F, Yeon S-J, Oh D-H. Food-Derived Opioid Peptides in Human Health: A Review. International Journal of Molecular Sciences. 2020; 21(22):8825. https://doi.org/10.3390/ijms21228825
Chicago/Turabian StyleTyagi, Akanksha, Eric Banan-Mwine Daliri, Fred Kwami Ofosu, Su-Jung Yeon, and Deog-Hwan Oh. 2020. "Food-Derived Opioid Peptides in Human Health: A Review" International Journal of Molecular Sciences 21, no. 22: 8825. https://doi.org/10.3390/ijms21228825
APA StyleTyagi, A., Daliri, E. B.-M., Kwami Ofosu, F., Yeon, S.-J., & Oh, D.-H. (2020). Food-Derived Opioid Peptides in Human Health: A Review. International Journal of Molecular Sciences, 21(22), 8825. https://doi.org/10.3390/ijms21228825