Rational Design of a Glycoconjugate Vaccine against Group A Streptococcus

 ,
,  , and
, and 
Abstract
1. Introduction
2. Results
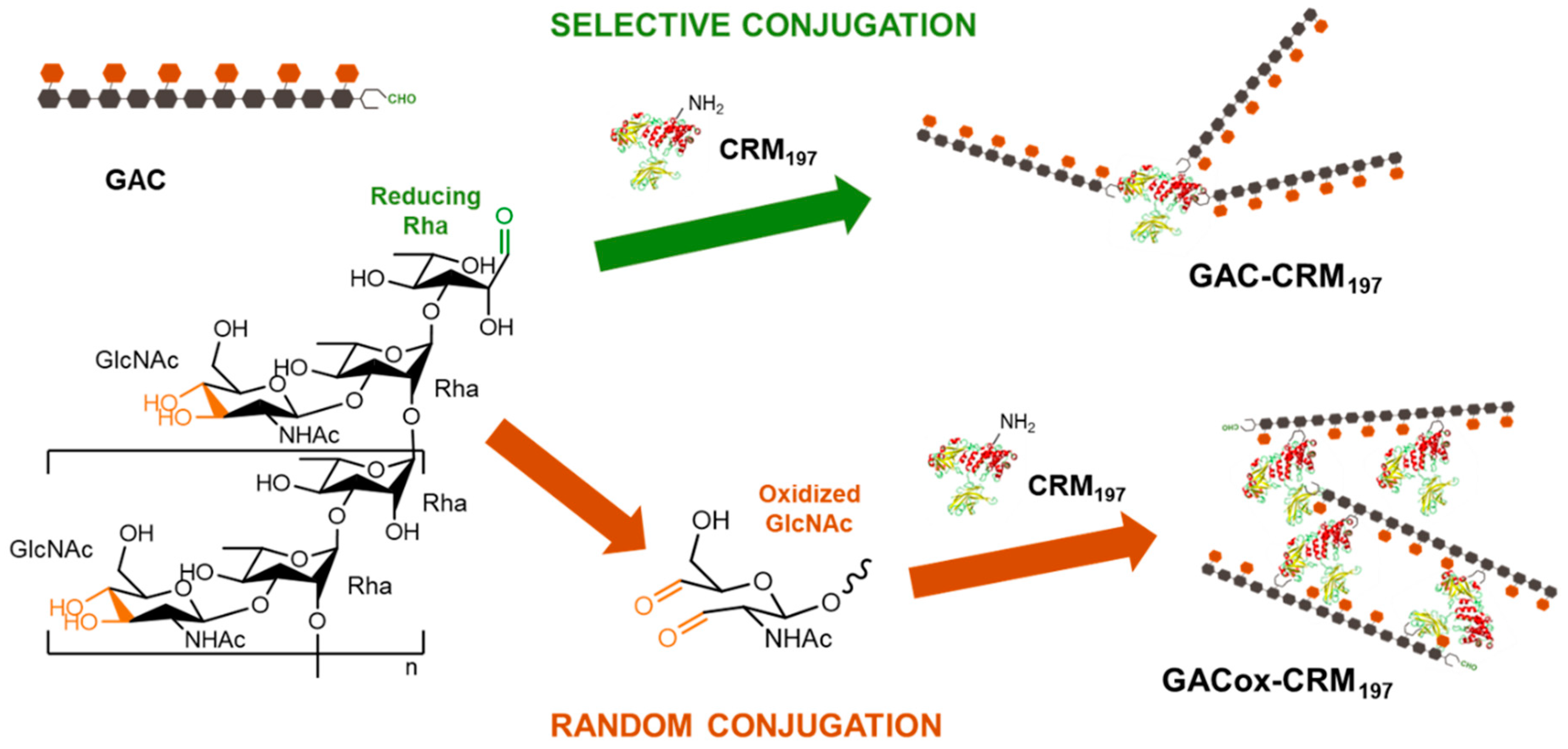
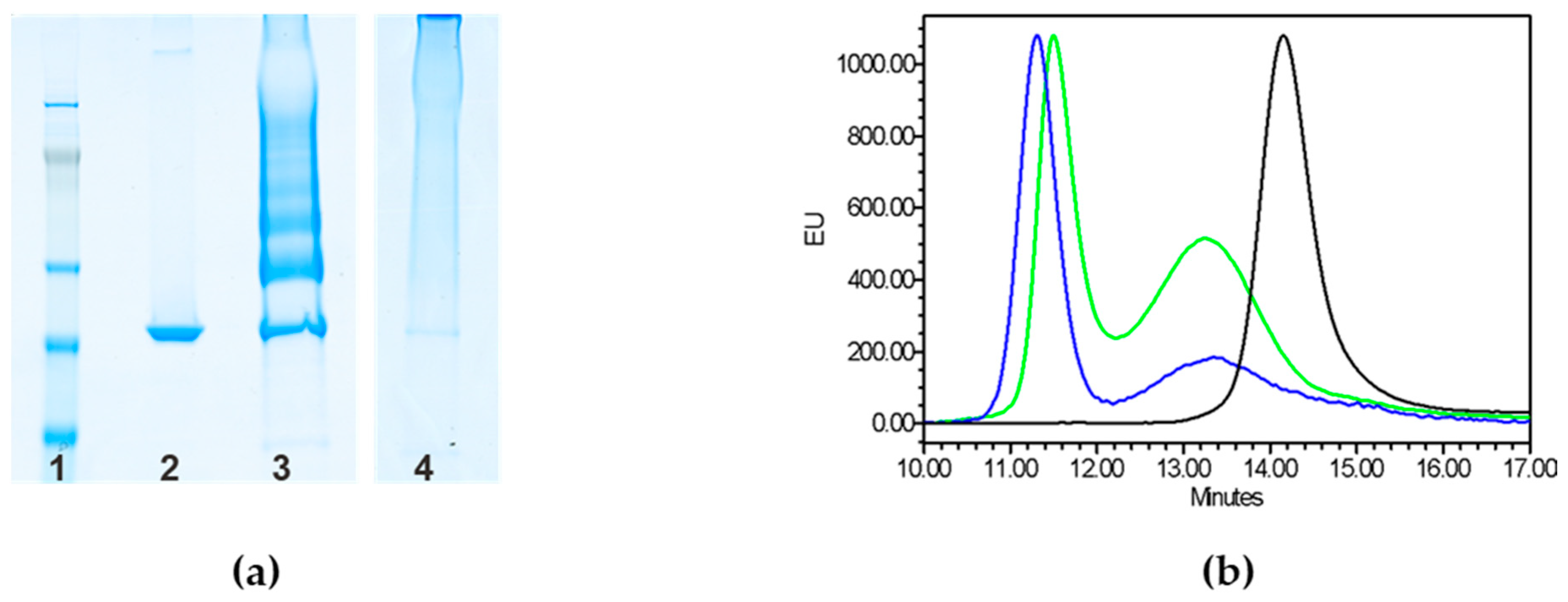
2.1. Testing Random and Selective Conjugation Chemistries for Linkage of GAC to CRM197
2.2. Applying Random Chemistry for Linkage of GAC to GAS Proteins
2.3. Optimization of the Random Chemistry through a DoE Approach
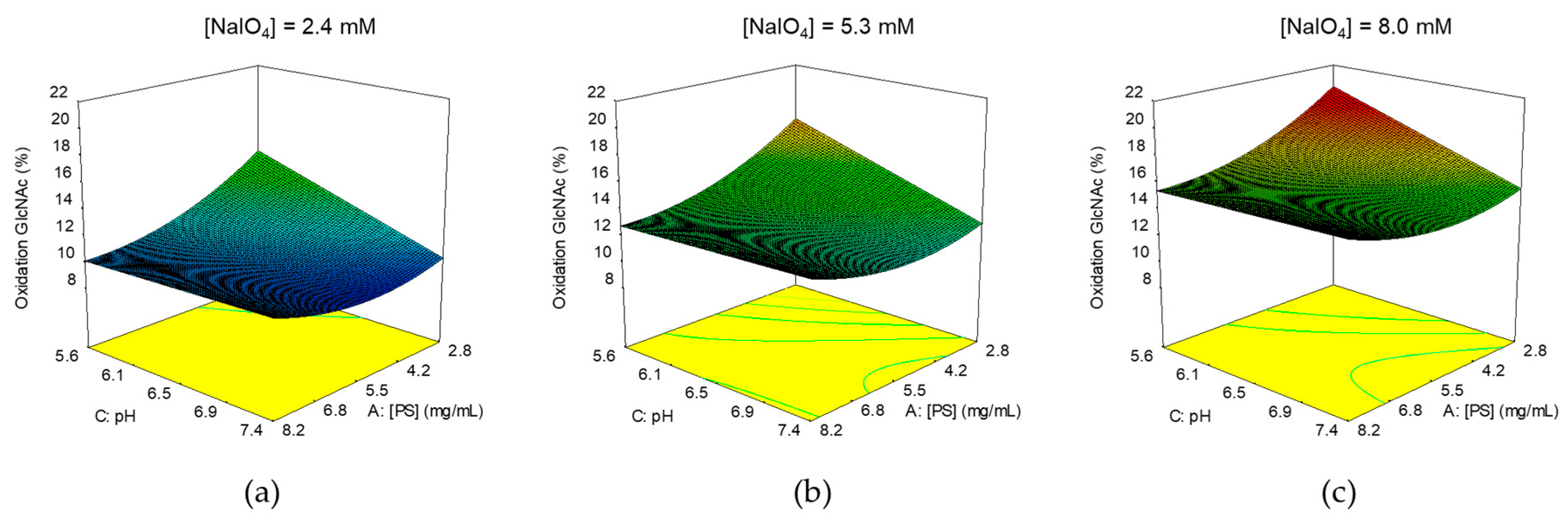
2.3.1. Identification of Optimal Conditions for GAC Oxidation
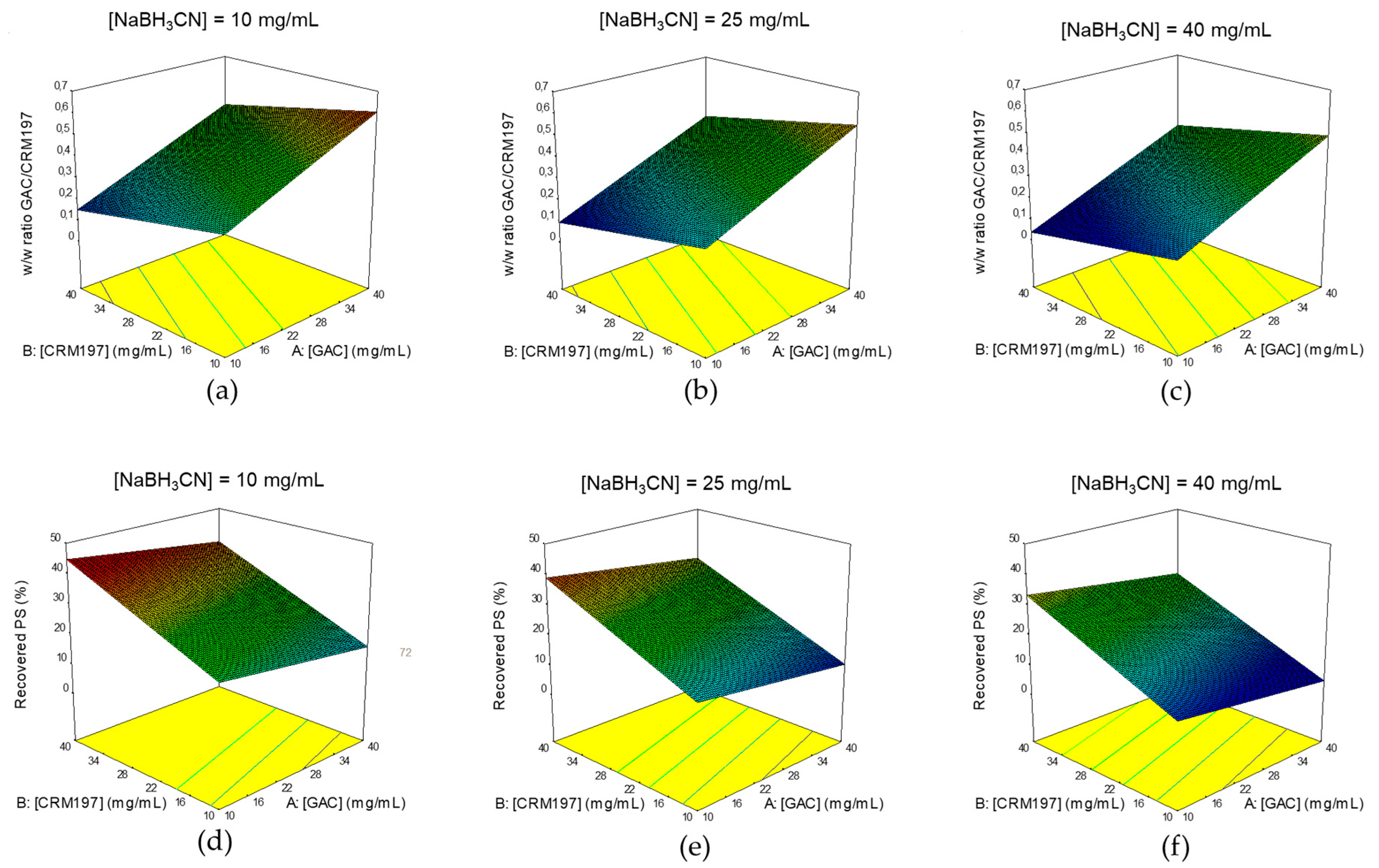
2.3.2. Identification of Optimal Conditions for GAC Conjugation to CRM197
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Conjugation of GAC to CRM197 through Selective Direct Reductive Amination
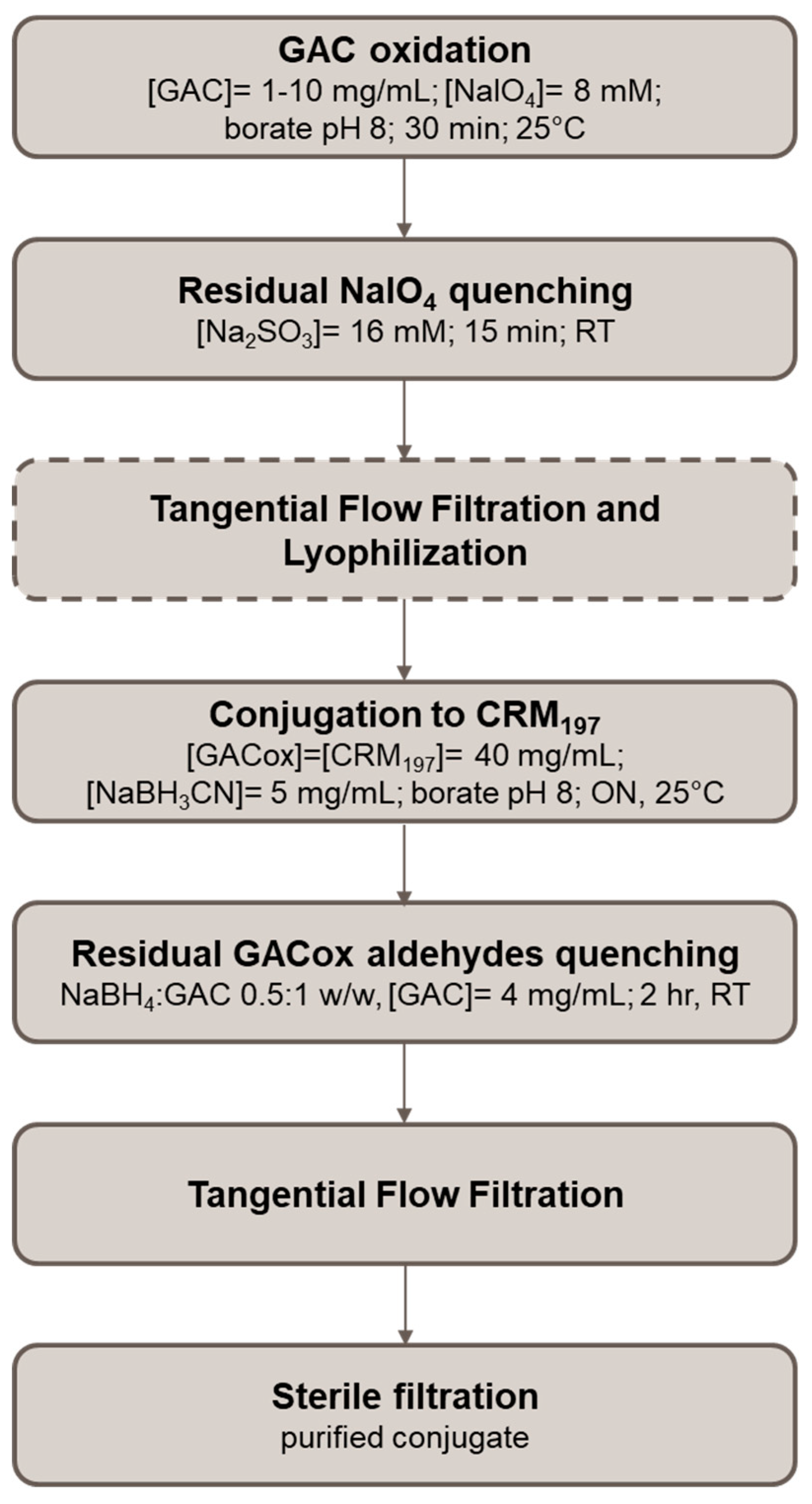
4.3. Conjugation of GAC to Different Carrier Proteins through Random Oxidation Followed by Reductive Amination
4.3.1. GAC Oxidation
4.3.2. Conjugation
4.4. Design of Experiment (DoE)
4.5. Analytical Methods
4.5.1. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
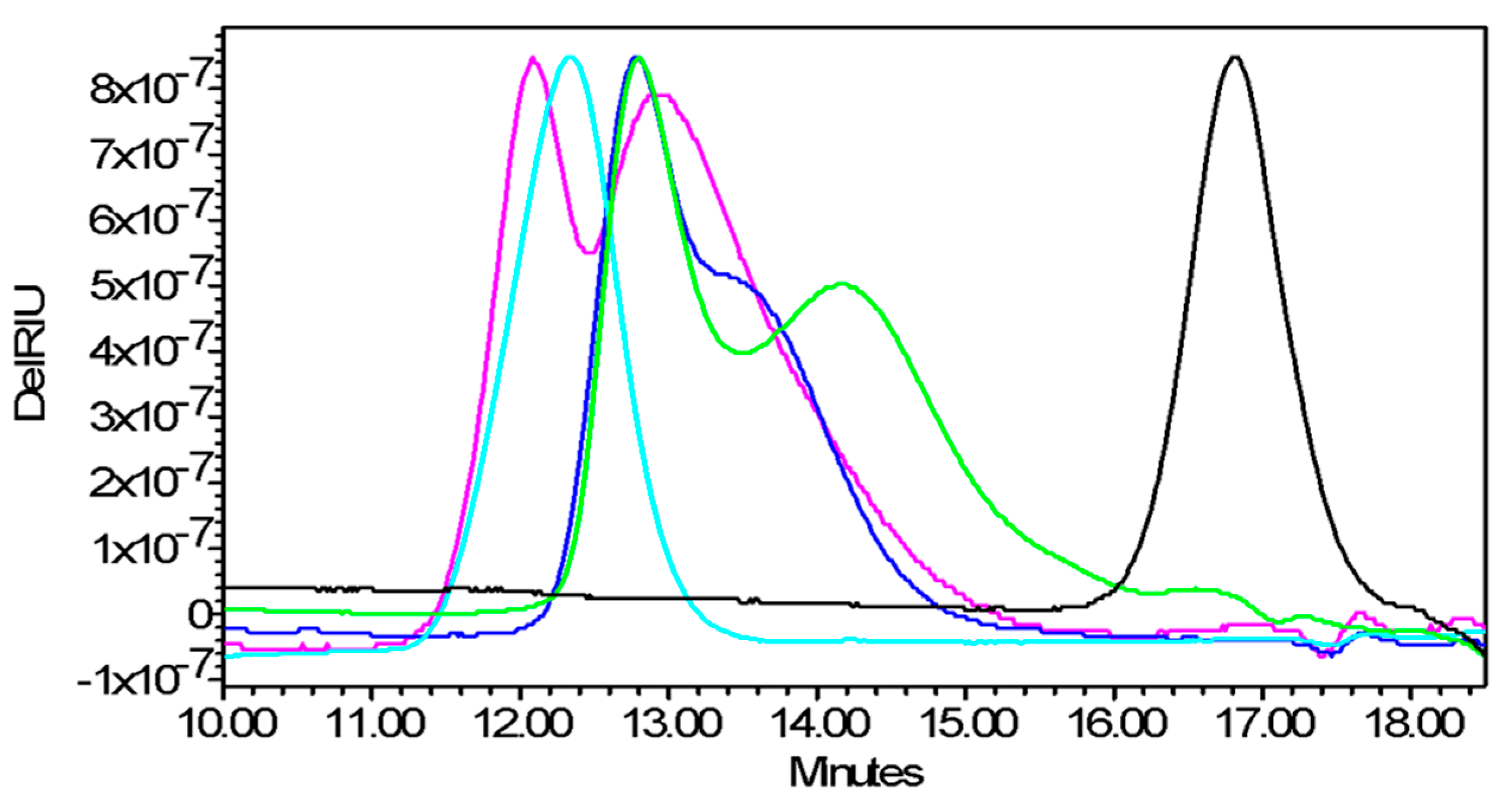
4.5.2. High Performance Liquid Chromatography–Size Exclusion Chromatography (HPLC–SEC)
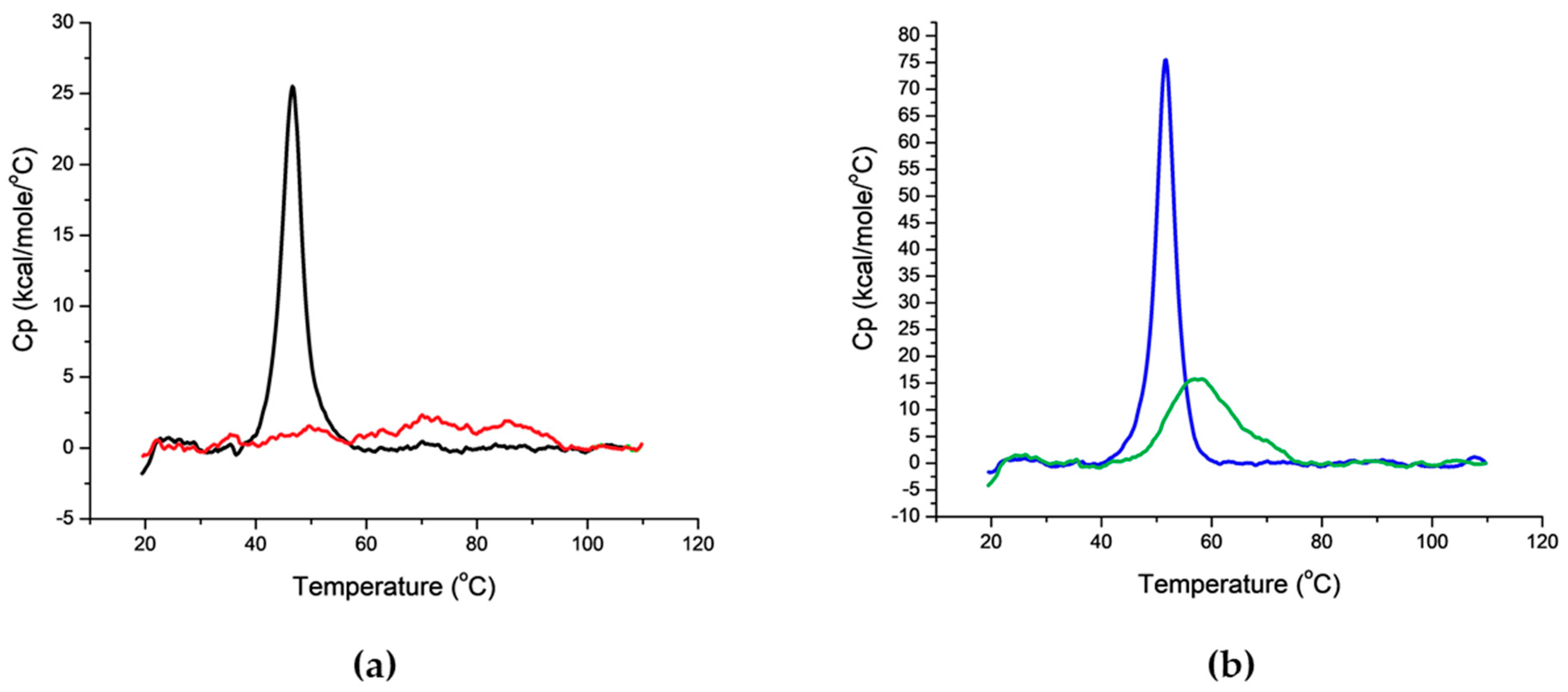
4.5.3. Differential Scanning Calorimetry (DSC)
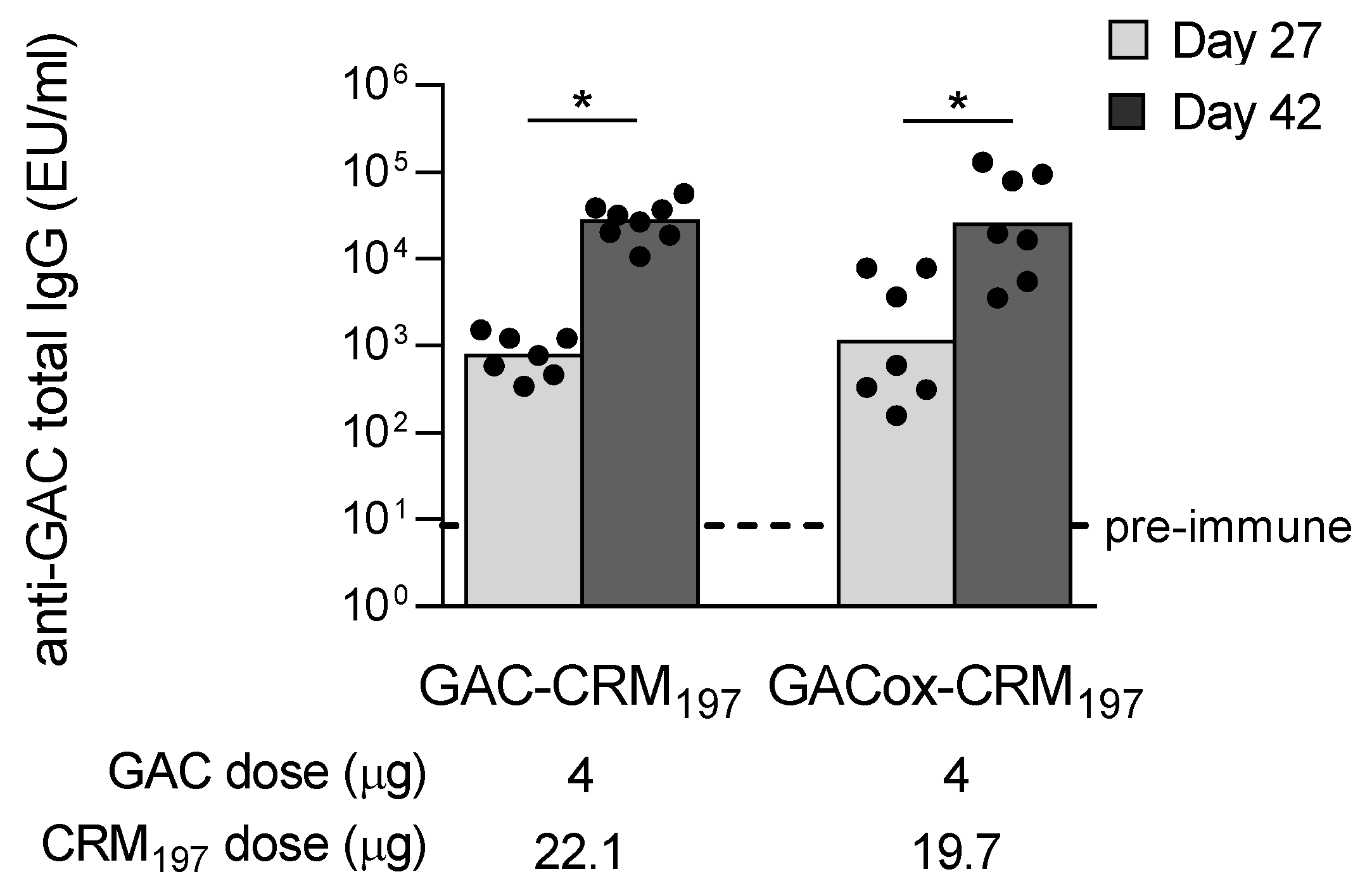
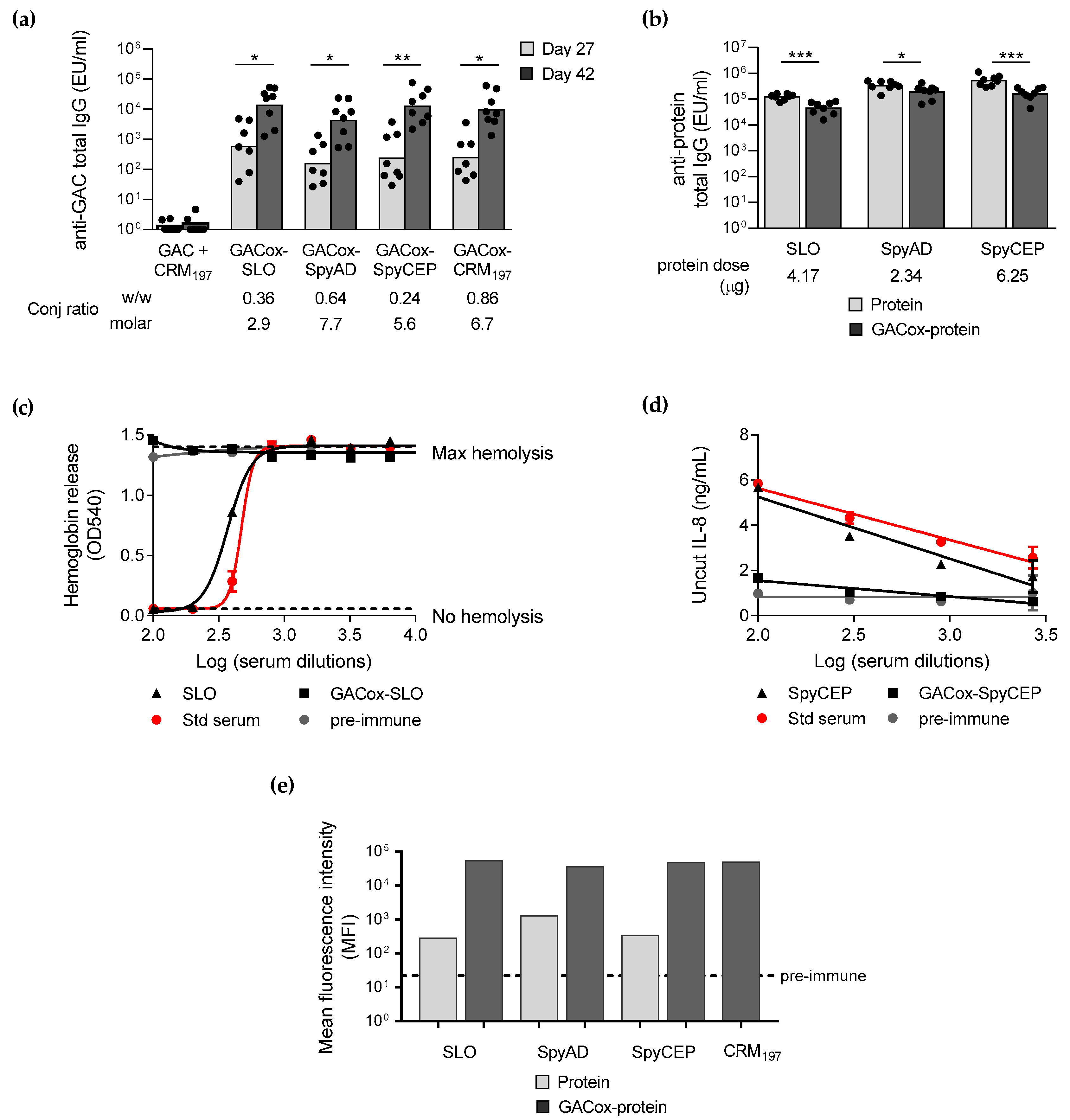
4.6. Immunogenicity Studies in Mice
4.7. Assessment of Anti-GAC and Anti-GAS Carrier Protein Immune Responses in Mice
4.8. Flow Cytometry (FACS)
4.9. Functional Assays
4.9.1. IL-8 Cleavage Inhibition Assay
4.9.2. In Vitro Hemolysis Assay
4.10. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GAS | Group A Streptococcus |
| GAC | Group A Carbohydrate |
| SLO | Streptolysin O |
| LMICs | Low- and Middle-income Countries |
| RHD | Rheumatic Hearth Disease |
| GlcNAc | N-acetylglucosamine |
| PS | Polysaccharide |
| SDS-PAGE | Sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| MW | Molecular Weight |
| HPLC-SEC | High Performance Liquid Chromatography–Size Exclusion Chromatography |
| i.p. | Intraperitoneally |
| DSC | Differential Scanning Calorimetry |
| FACS | Flow cytometry |
| DoE | Design of Experiment |
| RT | Room Temperature |
| Rha | Rhamnose |
| HPAEC-PAD | Anion Exchange Chromatography coupled with Pulsed Amperometric Detection |
| TFF | Tangential Flow Filtration |
| ELISA | Enzyme-linked immunosorbent assay |
References
- Ralph, A.P.; Carapetis, J.R. Group A streptococcal diseases and their global burden. Curr. Top. Microbiol. Immunol. 2013, 368, 1–27. [Google Scholar]
- Carapetis, J.R.; Steer, A.C.; Mulholland, E.K.; Weber, M. The global burden of group A streptococcal diseases. Lancet Infect. Dis. 2005, 5, 685–694. [Google Scholar] [CrossRef]
- Dooling, K.L.; Shapiro, D.J.; Van Beneden, C.; Hersh, A.L.; Hicks, L.A. Overprescribing and inappropriate antibiotic selection for children with pharyngitis in the United States, 1997–2010. JAMA Pediatr. 2014, 168, 1073–1074. [Google Scholar] [CrossRef] [PubMed]
- Jansen, K.U.; Knirsch, C.; Anderson, A.S. The role of vaccines in preventing bacterial antimicrobial resistance. Nat. Med. 2018, 24, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Watkins, D.A.; Johnson, C.O.; Colquhoun, S.M.; Karthikeyan, G.; Beaton, A.; Bukhman, G.; Forouzanfar, M.H.; Longenecker, C.T.; Mayosi, B.M.; Mensah, G.A.; et al. Global, regional, and national burden of rheumatic heart disease, 1990–2015. N. Engl. J. Med. 2017, 377, 713–722. [Google Scholar] [CrossRef]
- Vekemans, J.; Gouvea-Reis, F.; Kim, J.H.; Excler, J.L.; Smeesters, P.R.; O’Brien, K.L.; Van Beneden, C.A.; Steer, A.C.; Carapetis, J.R.; Kaslow, D.C. The path to group A streptococcus vaccines: World Health Organization research and development technology roadmap and preferred product characteristics. Clin. Infect. Dis. 2019, 69, 877–883. [Google Scholar] [CrossRef]
- Walker, M.J.; Barnett, T.C.; McArthur, J.D.; Cole, J.N.; Gillen, C.M.; Henningham, A.; Sriprakash, K.S.; Sanderson-Smith, M.L.; Nizet, V. Disease manifestations and pathogenic mechanisms of group A streptococcus. Clin. Microbiol. Rev. 2014, 27, 264–301. [Google Scholar] [CrossRef]
- Schneerson, R.; Barrera, O.; Sutton, A.; Robbins, J.B. Preparation, characterization, and immunogenicity of Haemophilus influenzae type b polysaccharide-protein conjugates. J. Exp. Med. 1980, 152, 361–376. [Google Scholar] [CrossRef]
- Berti, F.; Micoli, F. Improving efficacy of glycoconjugate vaccines: From chemical conjugates to next generation constructs. Curr. Opin. Immunol. 2020, 65, 42–49. [Google Scholar] [CrossRef]
- Rappuoli, R. Glycoconjugate vaccines: Principles and mechanisms. Sci. Transl. Med. 2018, 10, 456. [Google Scholar] [CrossRef]
- Salvadori, L.G.; Blake, M.S.; McCarty, M.; Tai, J.Y.; Zabriskie, J.B. Group A streptococcus-liposome ELISA antibody titers to group A polysaccharide and opsonophagocytic capabilities of the antibodies. J. Infect. Dis. 1995, 171, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Kabanova, A.; Margarit, I.; Berti, F.; Romano, M.R.; Grandi, G.; Bensi, G.; Chiarot, E.; Proietti, D.; Swennen, E.; Cappelletti, E.; et al. Evaluation of a group A streptococcus synthetic oligosaccharide as vaccine candidate. Vaccine 2010, 29, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Sabharwal, H.; Michon, F.; Nelson, D.; Dong, W.; Fuchs, K.; Manjarrez, R.C.; Sarkar, A.; Uitz, C.; Viteri-Jackson, A.; Suarez, R.S.; et al. Group A streptococcus (GAS) carbohydrate as an immunogen for protection against GAS infection. J. Infect. Dis. 2006, 193, 129–135. [Google Scholar] [CrossRef]
- Bensi, G.; Mora, M.; Tuscano, G.; Biagini, M.; Chiarot, E.; Bombaci, M.; Capo, S.; Falugi, F.; Manetti, A.G.; Donato, P.; et al. Multi high-throughput approach for highly selective identification of vaccine candidates: The group A streptococcus case. Mol. Cell. Proteom. 2012, 11, M111 015693. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, M.W. Pathogenesis of group A streptococcal infections. Clin. Microbiol. Rev. 2000, 13, 470–511. [Google Scholar] [CrossRef]
- Uchiyama, S.; Dohrmann, S.; Timmer, A.M.; Dixit, N.; Ghochani, M.; Bhandari, T.; Timmer, J.C.; Sprague, K.; Bubeck-Wardenburg, J.; Simon, S.I.; et al. Rapidly impairs neutrophil oxidative burst and antibacterial responses to group A streptococcus. Front. Immunol. 2015, 6, 581. [Google Scholar] [CrossRef]
- Gallotta, M.; Gancitano, G.; Pietrocola, G.; Mora, M.; Pezzicoli, A.; Tuscano, G.; Chiarot, E.; Nardi-Dei, V.; Taddei, A.R.; Rindi, S.; et al. SpyAD, a moonlighting protein of group A streptococcus contributing to bacterial division and host cell adhesion. Infect. Immun. 2014, 82, 2890–2901. [Google Scholar] [CrossRef]
- Edwards, R.J.; Taylor, G.W.; Ferguson, M.; Murray, S.; Rendell, N.; Wrigley, A.; Bai, Z.; Boyle, J.; Finney, S.J.; Jones, A.; et al. Specific C-terminal cleavage and inactivation of interleukin-8 by invasive disease isolates of Streptococcus pyogenes. J. Infect. Dis. 2005, 192, 783–790. [Google Scholar] [CrossRef]
- Jobichen, C.; Tan, Y.C.; Prabhakar, M.T.; Nayak, D.; Biswas, D.; Pannu, N.S.; Hanski, E.; Sivaraman, J. Structure of ScpC, a virulence protease from Streptococcus pyogenes, reveals the functional domains and maturation mechanism. Biochem. J. 2018, 475, 2847–2860. [Google Scholar] [CrossRef]
- Davies, M.R.; McIntyre, L.; Mutreja, A.; Lacey, J.A.; Lees, J.A.; Towers, R.J.; Duchene, S.; Smeesters, P.R.; Frost, H.R.; Price, D.J.; et al. Atlas of group A streptococcal vaccine candidates compiled using large-scale comparative genomics. Nat. Genet. 2019, 51, 1035–1043. [Google Scholar] [CrossRef]
- Micoli, F.; Adamo, R.; Costantino, P. Protein carriers for glycoconjugate vaccines: History, selection criteria, characterization and new trends. Molecules 2018, 23, 6. [Google Scholar] [CrossRef] [PubMed]
- Tontini, M.; Romano, M.R.; Proietti, D.; Balducci, E.; Micoli, F.; Balocchi, C.; Santini, L.; Masignani, V.; Berti, F.; Costantino, P. Preclinical studies on new proteins as carrier for glycoconjugate vaccines. Vaccine 2016, 34, 4235–4242. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Katzenellenbogen, E.; Jennings, H.J. Improved procedures for the conjugation of oligosaccharides to protein by reductive amination. Can. J. Biochem. Cell. Biol. 1984, 62, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Steer, A.C.; Law, I.; Matatolu, L.; Beall, B.W.; Carapetis, J.R. Global emm type distribution of group A streptococci: Systematic review and implications for vaccine development. Lancet Infect. Dis. 2009, 9, 611–616. [Google Scholar] [CrossRef]
- Dale, J.B.; Penfound, T.A.; Chiang, E.Y.; Walton, W.J. New 30-valent M protein-based vaccine evokes cross-opsonic antibodies against non-vaccine serotypes of group A streptococci. Vaccine 2011, 29, 8175–8178. [Google Scholar] [CrossRef]
- Postol, E.; Alencar, R.; Higa, F.T.; Freschi de Barros, S.; Demarchi, L.M.; Kalil, J.; Guilherme, L. StreptInCor: A candidate vaccine epitope against S. pyogenes infections induces protection in outbred mice. PLoS ONE 2013, 8, e60969. [Google Scholar] [CrossRef]
- Sekuloski, S.; Batzloff, M.R.; Griffin, P.; Parsonage, W.; Elliott, S.; Hartas, J.; O’Rourke, P.; Marquart, L.; Pandey, M.; Rubin, F.A.; et al. Evaluation of safety and immunogenicity of a group A streptococcus vaccine candidate (MJ8VAX) in a randomized clinical trial. PLoS ONE 2018, 13, e0198658. [Google Scholar] [CrossRef]
- Steer, A.C.; Carapetis, J.R.; Dale, J.B.; Fraser, J.D.; Good, M.F.; Guilherme, L.; Moreland, N.J.; Mulholland, E.K.; Schodel, F.; Smeesters, P.R. Status of research and development of vaccines for Streptococcus pyogenes. Vaccine 2016, 34, 2953–2958. [Google Scholar] [CrossRef]
- Avci, F.; Berti, F.; Dull, P.; Hennessey, J.; Pavliak, V.; Prasad, A.K.; Vann, W.; Wacker, M.; Marcq, O. Glycoconjugates: What it would take to master these well-known yet little-understood immunogens for vaccine development. mSphere 2019, 4, 5. [Google Scholar] [CrossRef]
- Dagan, R.; Poolman, J.; Siegrist, C.A. Glycoconjugate vaccines and immune interference: A review. Vaccine 2010, 28, 5513–5523. [Google Scholar] [CrossRef]
- Michon, F.; Fusco, P.C.; Minetti, C.A.; Laude-Sharp, M.; Uitz, C.; Huang, C.H.; D’Ambra, A.J.; Moore, S.; Remeta, D.P.; Heron, I.; et al. Multivalent pneumococcal capsular polysaccharide conjugate vaccines employing genetically detoxified pneumolysin as a carrier protein. Vaccine 1998, 16, 1732–1741. [Google Scholar] [CrossRef]
- Nilo, A.; Morelli, L.; Passalacqua, I.; Brogioni, B.; Allan, M.; Carboni, F.; Pezzicoli, A.; Zerbini, F.; Maione, D.; Fabbrini, M.; et al. Anti-group B Streptococcus glycan-conjugate vaccines using pilus protein GBS80 as carrier and antigen: Comparing Lysine and Tyrosine-directed conjugation. ACS Chem. Biol. 2015, 10, 1737–1746. [Google Scholar] [CrossRef] [PubMed]
- Nilo, A.; Passalacqua, I.; Fabbrini, M.; Allan, M.; Usera, A.; Carboni, F.; Brogioni, B.; Pezzicoli, A.; Cobb, J.; Romano, M.R.; et al. Exploring the effect of conjugation site and chemistry on the immunogenicity of an anti-group B streptococcus glycoconjugate vaccine based on GBS67 pilus protein and type V Polysaccharide. Bioconjug. Chem. 2015, 26, 1839–1849. [Google Scholar] [CrossRef] [PubMed]
- Pinto, V.B.; Burden, R.; Wagner, A.; Moran, E.E.; Lee, C.H. The development of an experimental multiple serogroups vaccine for Neisseria meningitidis. PLoS ONE 2013, 8, e79304. [Google Scholar] [CrossRef] [PubMed]
- Pozzi, C.; Wilk, K.; Lee, J.C.; Gening, M.; Nifantiev, N.; Pier, G.B. Opsonic and protective properties of antibodies raised to conjugate vaccines targeting six Staphylococcus aureus antigens. PLoS ONE 2012, 7, e46648. [Google Scholar] [CrossRef] [PubMed]
- Simon, R.; Tennant, S.M.; Wang, J.Y.; Schmidlein, P.J.; Lees, A.; Ernst, R.K.; Pasetti, M.F.; Galen, J.E.; Levine, M.M. Salmonella enterica serovar enteritidis core O polysaccharide conjugated to H: g,m flagellin as a candidate vaccine for protection against invasive infection with S. enteritidis. Infect. Immun. 2011, 79, 4240–4249. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Hernandez, T.; Pandey, M.; Henningham, A.; Cole, J.; Choudhury, B.; Cork, A.J.; Gillen, C.M.; Ghaffar, K.A.; West, N.P.; Silvestri, G.; et al. Differing efficacies of lead group A streptococcal vaccine candidates and full-length M protein in cutaneous and invasive disease models. mBio 2016, 7. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, Y.; Wang, G.; Feng, S.; Guo, Z.; Gu, G. Group A Streptococcus cell wall oligosaccharide-streptococcal C5a peptidase conjugates as effective antibacterial vaccines. ACS Infect. Dis. 2020, 6, 281–290. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, S.; Wang, G.; Li, H.; Guo, Z.; Gu, G. Synthesis and immunological studies of group A Streptococcus cell-wall oligosaccharide–streptococcal C5a peptidase conjugates as bivalent vaccines. Org. Chem. Front. 2019, 6, 3589–3596. [Google Scholar] [CrossRef]
- Chiarot, E.; Faralla, C.; Chiappini, N.; Tuscano, G.; Falugi, F.; Gambellini, G.; Taddei, A.; Capo, S.; Cartocci, E.; Veggi, D.; et al. Targeted amino acid substitutions impair streptolysin O toxicity and group A Streptococcus virulence. mBio 2013, 4, e00387-12. [Google Scholar] [CrossRef]
- McKenna, S.; Malito, E.; Rouse, S.L.; Abate, F.; Bensi, G.; Chiarot, E.; Micoli, F.; Mancini, F.; Gomes Moriel, D.; Grandi, G.; et al. Structure, dynamics and immunogenicity of a catalytically inactive CXC chemokine-degrading protease SpyCEP from Streptococcus pyogenes. Comput. Struct. Biotechnol. J. 2020, 18, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Stefanetti, G.; Hu, Q.Y.; Usera, A.; Robinson, Z.; Allan, M.; Singh, A.; Imase, H.; Cobb, J.; Zhai, H.; Quinn, D.; et al. Sugar-protein connectivity impacts on the immunogenicity of site-selective salmonella O-antigen glycoconjugate vaccines. Angew. Chem. Int. Ed. Engl. 2015, 54, 13198–13203. [Google Scholar] [CrossRef] [PubMed]
- Wacker, M.; Linton, D.; Hitchen, P.G.; Nita-Lazar, M.; Haslam, S.M.; North, S.J.; Panico, M.; Morris, H.R.; Dell, A.; Wren, B.W.; et al. N-linked glycosylation in Campylobacter jejuni and its functional transfer into E. coli. Science 2002, 298, 1790–1793. [Google Scholar] [CrossRef] [PubMed]
- Wacker, M.; Wang, L.; Kowarik, M.; Dowd, M.; Lipowsky, G.; Faridmoayer, A.; Shields, K.; Park, S.; Alaimo, C.; Kelley, K.A.; et al. Prevention of Staphylococcus aureus infections by glycoprotein vaccines synthesized in Escherichia coli. J. Infect. Dis. 2014, 209, 1551–1561. [Google Scholar] [CrossRef]
- Carmenate, T.; Canaan, L.; Alvarez, A.; Delgado, M.; Gonzalez, S.; Menendez, T.; Rodes, L.; Guillen, G. Effect of conjugation methodology on the immunogenicity and protective efficacy of meningococcal group C polysaccharide-P64k protein conjugates. FEMS Immunol. Med. Microbiol. 2004, 40, 193–199. [Google Scholar] [CrossRef]
- Costantino, P.; Rappuoli, R.; Berti, F. The design of semi-synthetic and synthetic glycoconjugate vaccines. Expert Opin. Drug. Discov. 2011, 6, 1045–1066. [Google Scholar] [CrossRef]
- Fattom, A.; Li, X.; Cho, Y.H.; Burns, A.; Hawwari, A.; Shepherd, S.E.; Coughlin, R.; Winston, S.; Naso, R. Effect of conjugation methodology, carrier protein, and adjuvants on the immune response to Staphylococcus aureus capsular polysaccharides. Vaccine 1995, 13, 1288–1293. [Google Scholar] [CrossRef]
- Rush, J.S.; Edgar, R.J.; Deng, P.; Chen, J.; Zhu, H.; van Sorge, N.M.; Morris, A.J.; Korotkov, K.V.; Korotkova, N. The molecular mechanism of N-acetylglucosamine side-chain attachment to the Lancefield group A carbohydrate in Streptococcus pyogenes. J. Biol. Chem. 2017, 292, 19441–19457. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Kasper, D.L. Oxidative depolymerization of polysaccharides by reactive oxygen/nitrogen species. Glycobiology 2011, 21, 401–409. [Google Scholar] [CrossRef]
- Kholy, A.E.; Facklam, R.; Sabri, G.; Rotta, J. Serological identification of group A streptococci from throat scrapings before culture. J. Clin. Microbiol. 1978, 8, 725–728. [Google Scholar]
- Pancholi, V.; Fischetti, V.A. Isolation and characterization of the cell-associated region of group A streptococcal M6 protein. J. Bacteriol. 1988, 170, 2618–2624. [Google Scholar] [CrossRef] [PubMed]
- Beg, S.; Swain, S.; Rahman, M.; Hasnain, M.S.; Imam, S.S. Application of design of experiments (DoE) in pharmaceutical product and process optimization. In Pharmaceutical Quality by Design; Beg, S., Hasnain, M.S., Eds.; Academic Press: Amsterdam, The Netherlands, 2019; pp. 43–64. [Google Scholar]
- Montgomery, D.C. (Ed.) Response surface methods and other approaches to process optimization. In Design and Analysis of Experiments; John Wiley and Sons: New York, NY, USA, 1997. [Google Scholar]
- Giannelli, C.; Raso, M.M.; Palmieri, E.; De Felice, A.; Pippi, F.; Micoli, F. Development of a specific and sensitive HPAEC-PAD method for quantification of Vi Polysaccharide applicable to other polysaccharides containing amino uronic acids. Anal. Chem. 2020, 92, 6304–6311. [Google Scholar] [CrossRef] [PubMed]
- Joelsson, D.; Moravec, P.; Troutman, M.; Pigeon, J.; DePhillips, P. Optimizing ELISAs for precision and robustness using laboratory automation and statistical design of experiments. J. Immunol. Methods 2008, 337, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Necchi, F.; Carducci, M.; Pisoni, I.; Rossi, O.; Saul, A.; Rondini, S. Development of FAcE (formulated Alhydrogel competitive ELISA) method for direct quantification of OAg present in Shigella sonnei GMMA-based vaccine and its optimization using design of experiments approach. J. Immunol. Methods 2019, 471, 11–17. [Google Scholar] [CrossRef]
- Ahl, P.L.; Mensch, C.; Hu, B.; Pixley, H.; Zhang, L.; Dieter, L.; Russell, R.; Smith, W.J.; Przysiecki, C.; Kosinski, M.; et al. Accelerating vaccine formulation development using design of experiment stability studies. J. Pharm. Sci. 2016, 105, 3046–3056. [Google Scholar] [CrossRef]
- Ji, Y.; Tian, Y.; Ahnfelt, M.; Sui, L. Design and optimization of a chromatographic purification process for Streptococcus pneumoniae serotype 23F capsular polysaccharide by a design of experiments approach. J. Chromatogr. A 2014, 1348, 137–149. [Google Scholar] [CrossRef]
- Kanojia, G.; Willems, G.J.; Frijlink, H.W.; Kersten, G.F.; Soema, P.C.; Amorij, J.P. A design of experiment approach to predict product and process parameters for a spray dried influenza vaccine. Int. J. Pharm. 2016, 511, 1098–1111. [Google Scholar] [CrossRef]
- Patel, A.; Erb, S.M.; Strange, L.; Shukla, R.S.; Kumru, O.S.; Smith, L.; Nelson, P.; Joshi, S.B.; Livengood, J.A.; Volkin, D.B. Combined semi-empirical screening and design of experiments (DOE) approach to identify candidate formulations of a lyophilized live attenuated tetravalent viral vaccine candidate. Vaccine 2018, 36, 3169–3179. [Google Scholar] [CrossRef]
- Abate, F.; Malito, E.; Falugi, F.; Margarit, Y.R.I.; Bottomley, M.J. Cloning, expression, purification, crystallization and preliminary X-ray diffraction analysis of SpyCEP, a candidate antigen for a vaccine against Streptococcus pyogenes. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2013, 69, 1103–1106. [Google Scholar] [CrossRef]
- Stefanetti, G.; Rondini, S.; Lanzilao, L.; Saul, A.; MacLennan, C.A.; Micoli, F. Impact of conjugation chemistry on the immunogenicity of S. Typhimurium conjugate vaccines. Vaccine 2014, 32, 6122–6129. [Google Scholar] [CrossRef]
- Pitirollo, O.; Micoli, F.; Necchi, F.; Mancini, F.; Carducci, M.; Adamo, R.; Evangelisti, C.; Morelli, L.; Polito, L.; Lay, L. Gold nanoparticles morphology does not affect the multivalent presentation and antibody recognition of Group A Streptococcus synthetic oligorhamnans. Bioorg. Chem. 2020, 99, 103815. [Google Scholar] [CrossRef] [PubMed]
- Lei, Q.P.; Lamb, D.H.; Heller, R.; Pietrobon, P. Quantitation of low level unconjugated polysaccharide in tetanus toxoid-conjugate vaccine by HPAEC/PAD following rapid separation by deoxycholate/HCl. J. Pharm. Biomed. Anal. 2000, 21, 1087–1091. [Google Scholar] [CrossRef]
- Lanzilao, L.; Stefanetti, G.; Saul, A.; MacLennan, C.A.; Micoli, F.; Rondini, S. Strain selection for generation of O-antigen-based glycoconjugate vaccines against invasive nontyphoidal Salmonella disease. PLoS ONE 2015, 10, e0139847. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conjugate | GAC/Protein Molar Ratio | GAC/Protein w/w Ratio |
|---|---|---|
| GACox-CRM197 | 7.2 | 0.86 |
| GACox-SLO | 3.3 | 0.36 |
| GACox-SpyAD | 8.2 | 0.64 |
| GACox-SpyCEP | 6.0 | 0.24 |
| Optimized Conditions | GAC/CRM197 w/w | GAC Recovery % | ||
|---|---|---|---|---|
| Predicted (95% CI for Mean) | Actual | Predicted (95% CI for Mean) | Actual | |
| [GACox] = [CRM197] = 40 mg/mL; [NaBH3CN] = 10 mg/mL; borate buffer pH 8; T = 25 °C; 2 days reaction time | 0.46 (0.39–0.52) | 0.39 | 38 (32–43) | 39 |
| [NaBH3CN] in Reaction (mg/mL) | Reaction Time | GAC/CRM197 w/w Ratio in Purified Conjugate |
|---|---|---|
| 10 | 4 h | 0.34 |
| ON | 0.41 | |
| 2 days | 0.39 | |
| 5 | 4 h | 0.36 |
| ON | 0.46 | |
| 2 days | 0.48 | |
| 1 | 4 h | 0.42 |
| ON | 0.45 | |
| 2 days | 0.47 |
| GAC/CRM197 w/w | GAC Recovery % | ||
|---|---|---|---|
| Small Scale | Large Scale | Small Scale | Large Scale |
| 0.44 | 0.51 | 44 | 39 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Benedetto, R.; Mancini, F.; Carducci, M.; Gasperini, G.; Moriel, D.G.; Saul, A.; Necchi, F.; Rappuoli, R.; Micoli, F. Rational Design of a Glycoconjugate Vaccine against Group A Streptococcus. Int. J. Mol. Sci. 2020, 21, 8558. https://doi.org/10.3390/ijms21228558
Di Benedetto R, Mancini F, Carducci M, Gasperini G, Moriel DG, Saul A, Necchi F, Rappuoli R, Micoli F. Rational Design of a Glycoconjugate Vaccine against Group A Streptococcus. International Journal of Molecular Sciences. 2020; 21(22):8558. https://doi.org/10.3390/ijms21228558
Chicago/Turabian StyleDi Benedetto, Roberta, Francesca Mancini, Martina Carducci, Gianmarco Gasperini, Danilo Gomes Moriel, Allan Saul, Francesca Necchi, Rino Rappuoli, and Francesca Micoli. 2020. "Rational Design of a Glycoconjugate Vaccine against Group A Streptococcus" International Journal of Molecular Sciences 21, no. 22: 8558. https://doi.org/10.3390/ijms21228558
APA StyleDi Benedetto, R., Mancini, F., Carducci, M., Gasperini, G., Moriel, D. G., Saul, A., Necchi, F., Rappuoli, R., & Micoli, F. (2020). Rational Design of a Glycoconjugate Vaccine against Group A Streptococcus. International Journal of Molecular Sciences, 21(22), 8558. https://doi.org/10.3390/ijms21228558