Neural Tissue Degeneration in Rosenthal’s Canal and Its Impact on Electrical Stimulation of the Auditory Nerve by Cochlear Implants: An Image-Based Modeling Study

 ,
,  ,
, 

Abstract
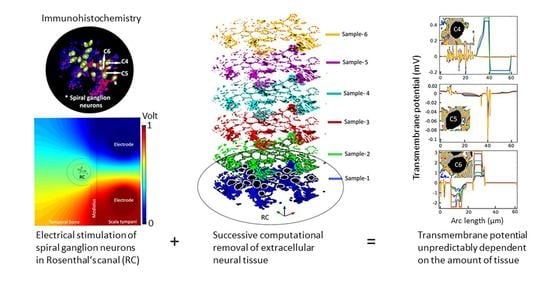
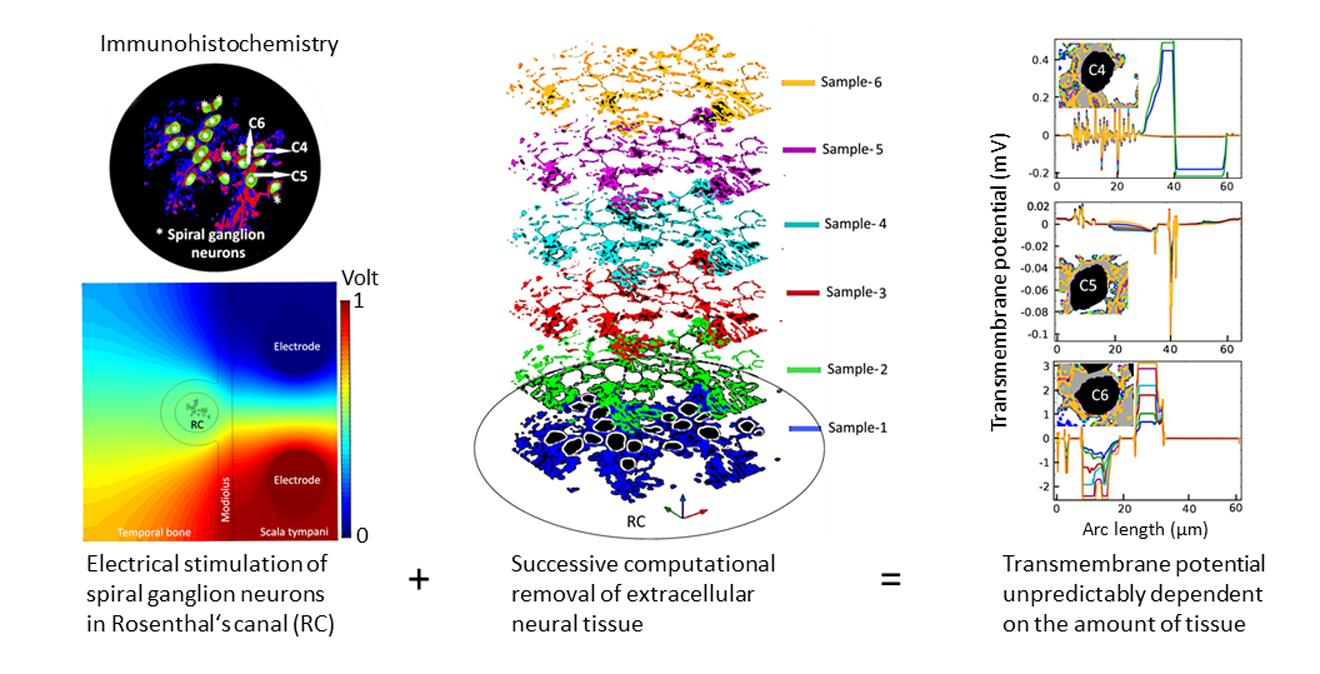
1. Introduction
2. Results
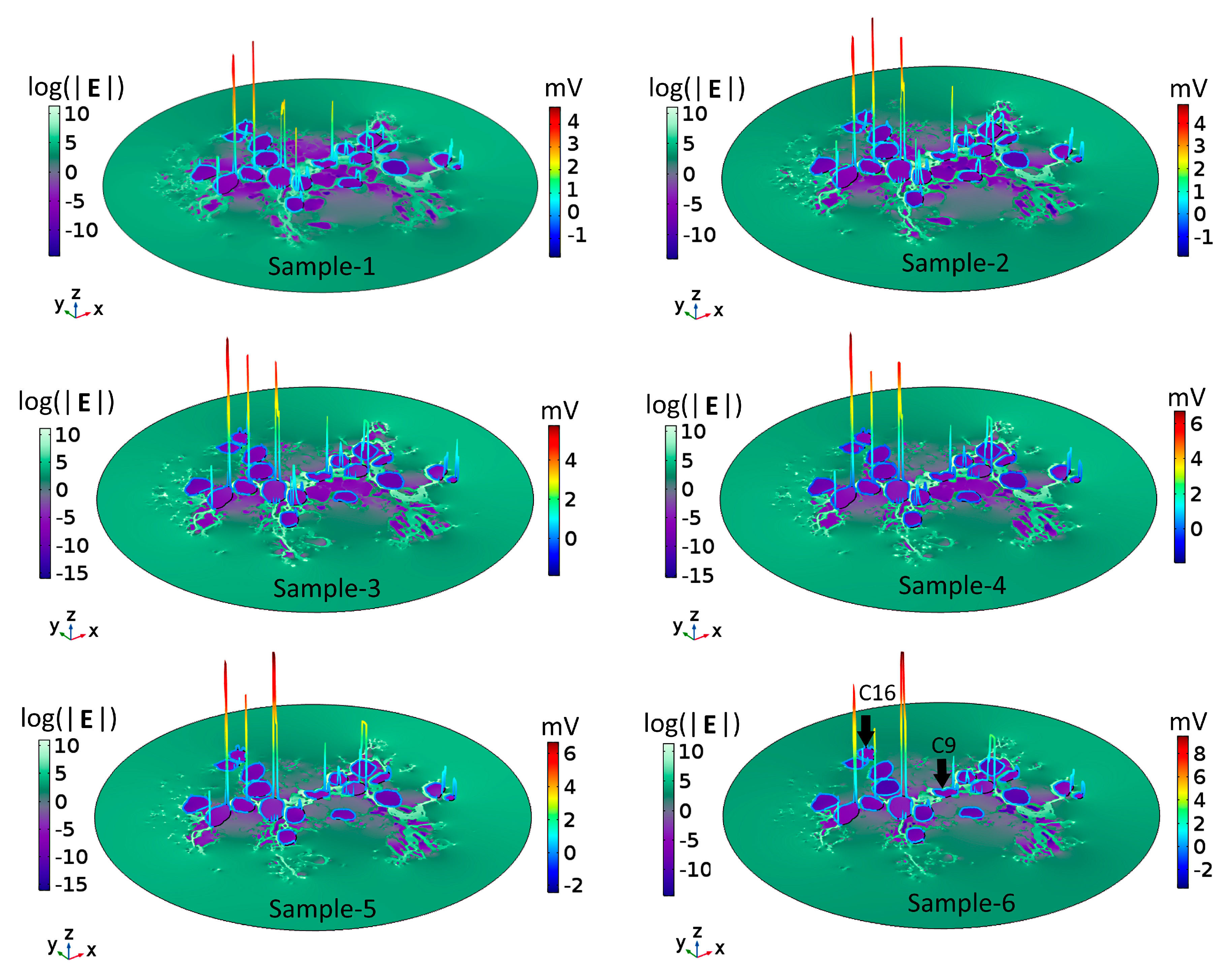
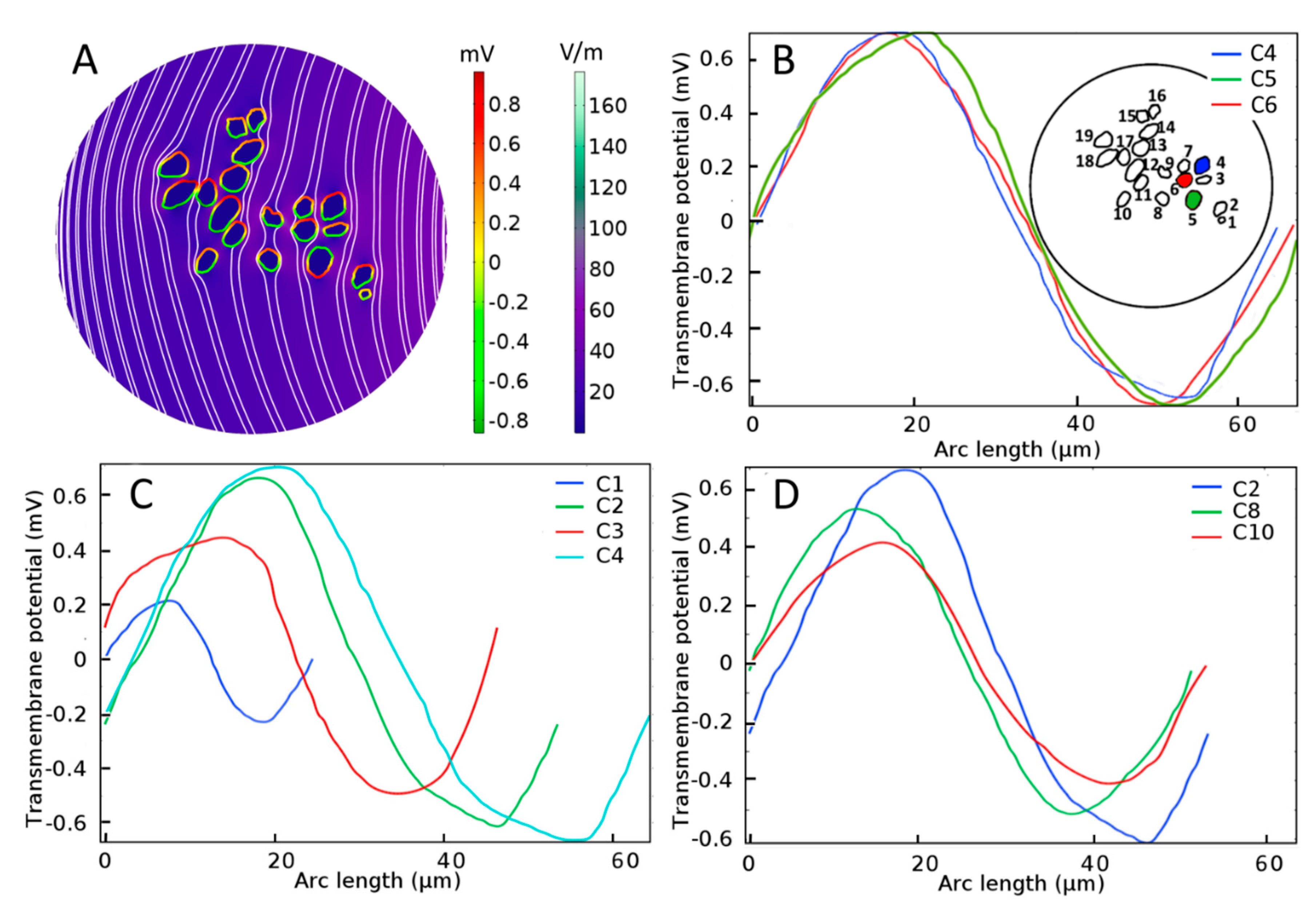
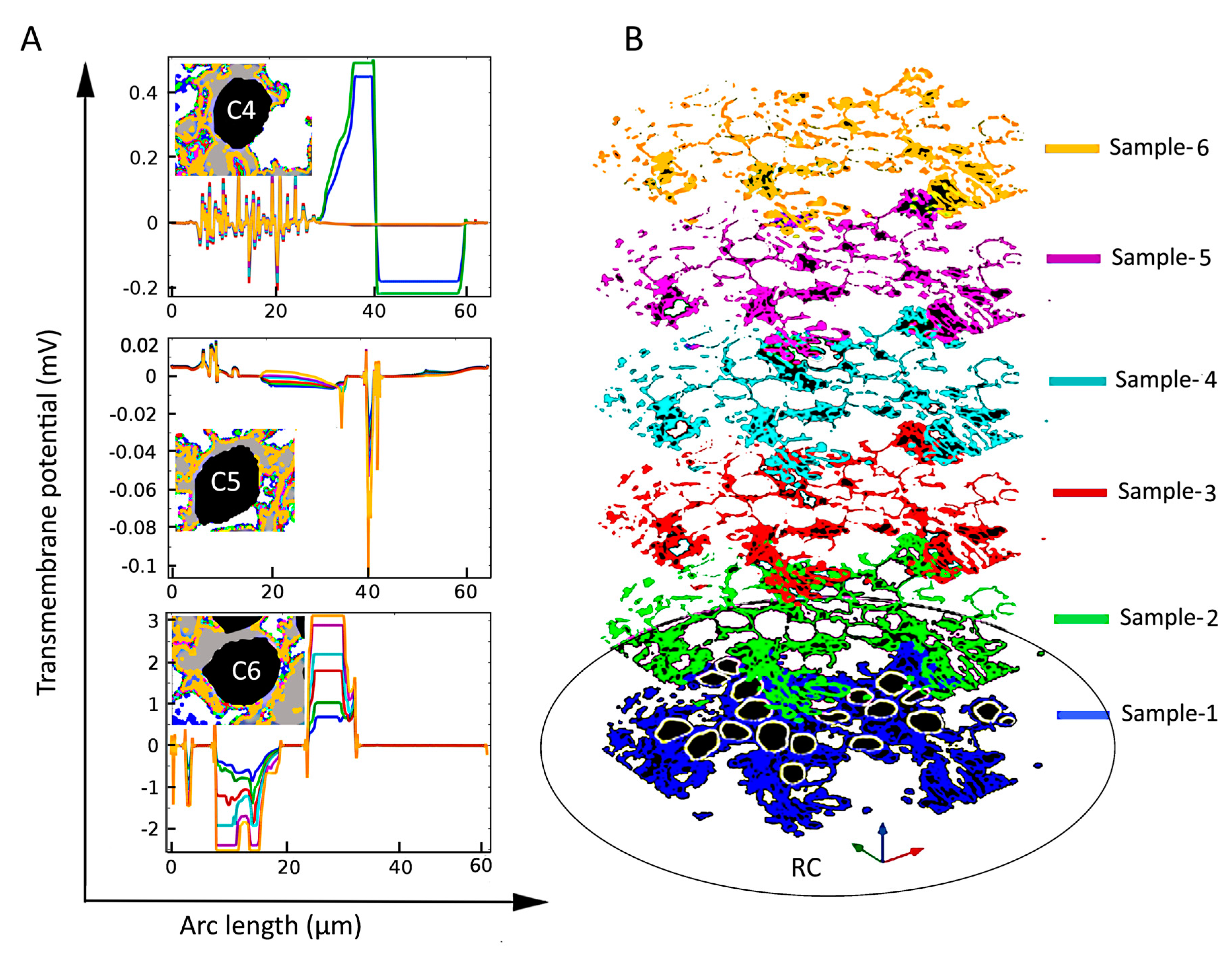
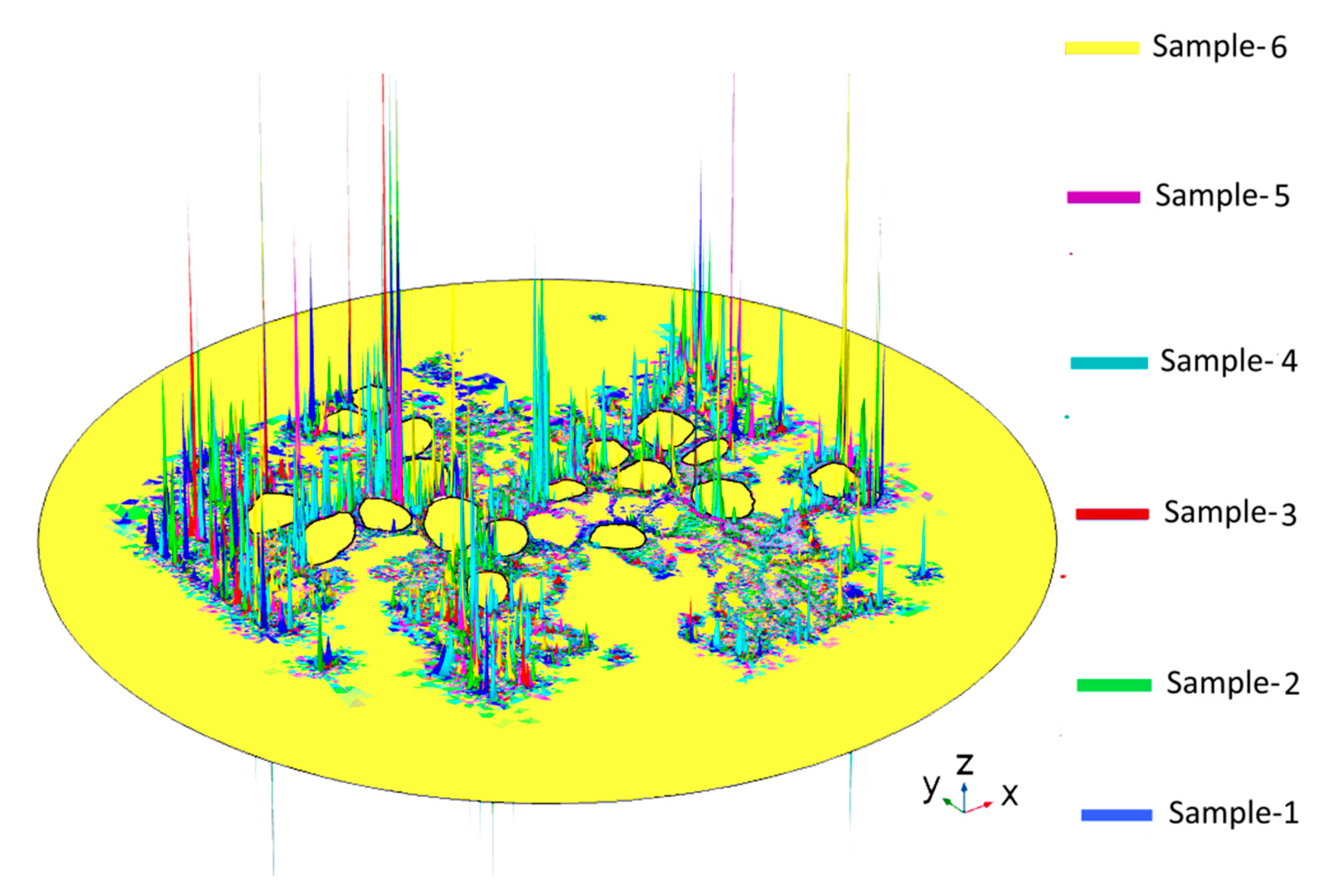
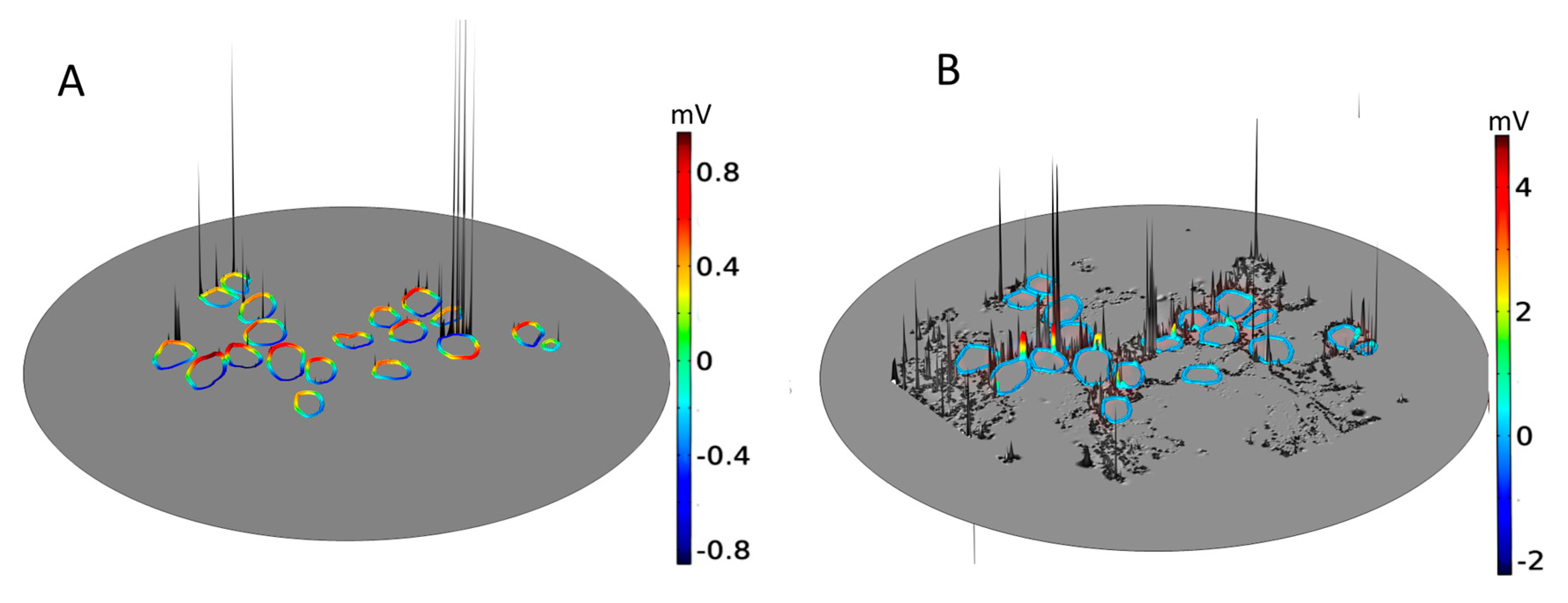
2.1. Dependence of the Maximal Transmembrane Potential on Tissue Density
2.2. Dependence of the Electric Field on Tissue Density
3. Discussion
3.1. Neural Stimulation in Homogeneous and Inhomogeneous Tissue Environments
3.2. The Activating Function as an Indicator of AP Initiation Site
3.3. Limitations of the Analysis
3.4. Potential Clinical Applications
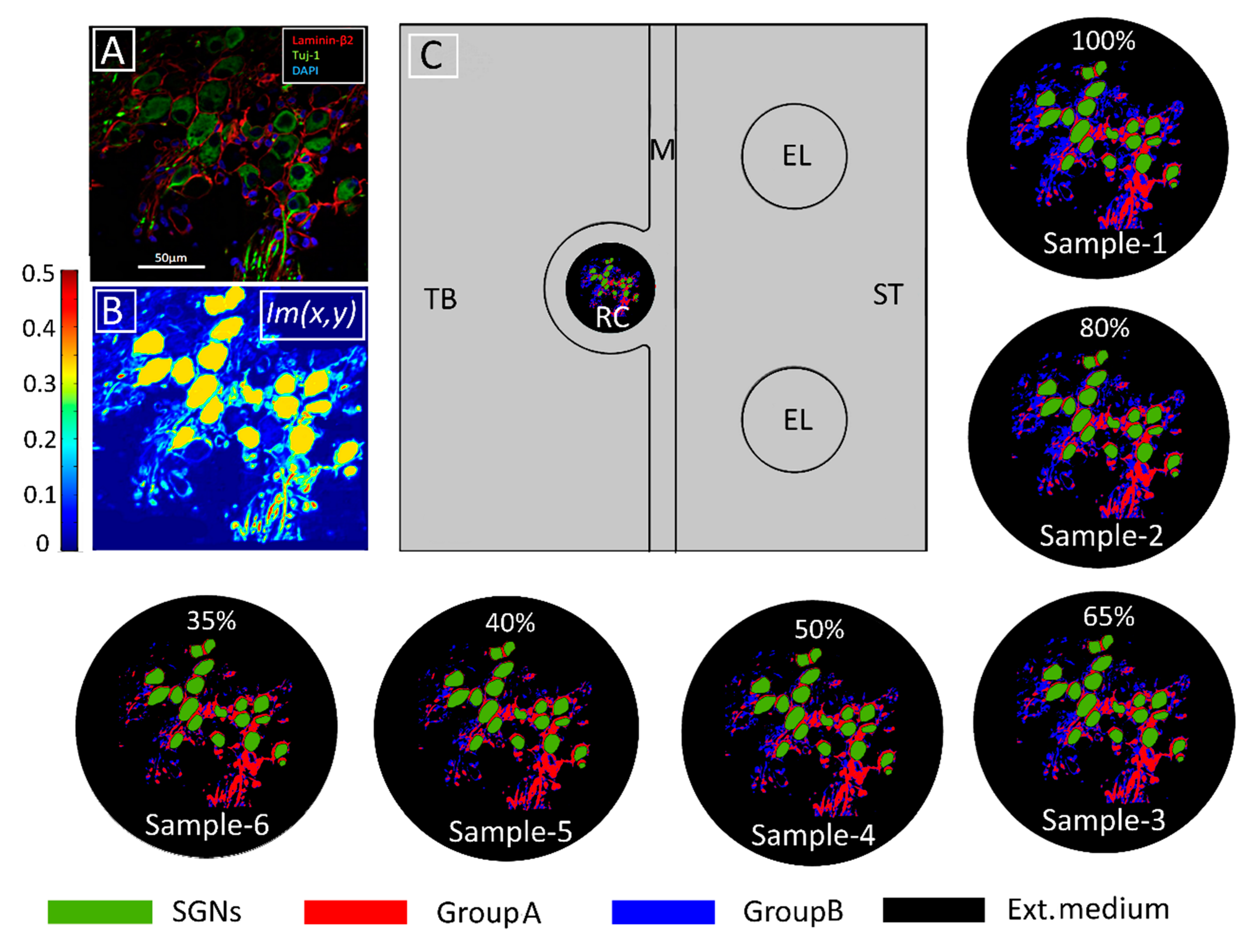
4. Materials and Methods
4.1. Basic Assumptions
4.2. Derivation of the Distribution of Electrical Conductivities
4.3. The Constitutive Equations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AN | Auditory Nerve |
| AP | Action Potential |
| CI | Cochlear Implant |
| SGN | Spiral Ganglion Neuron |
| RC | Rosenthal’s Canal |
| Transmembrane Potential |
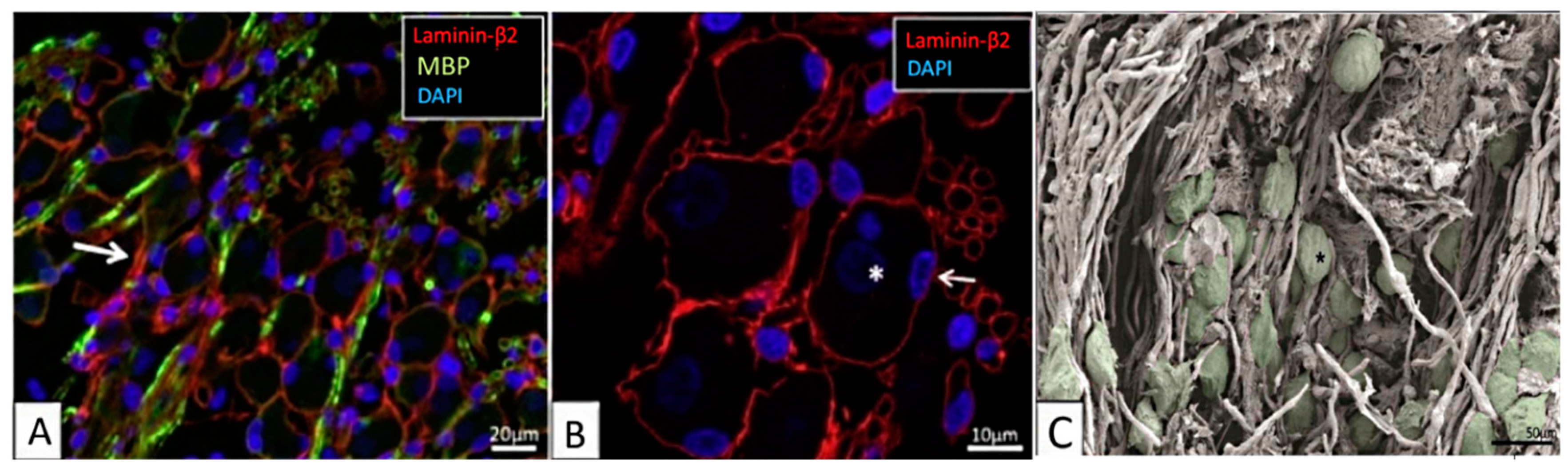
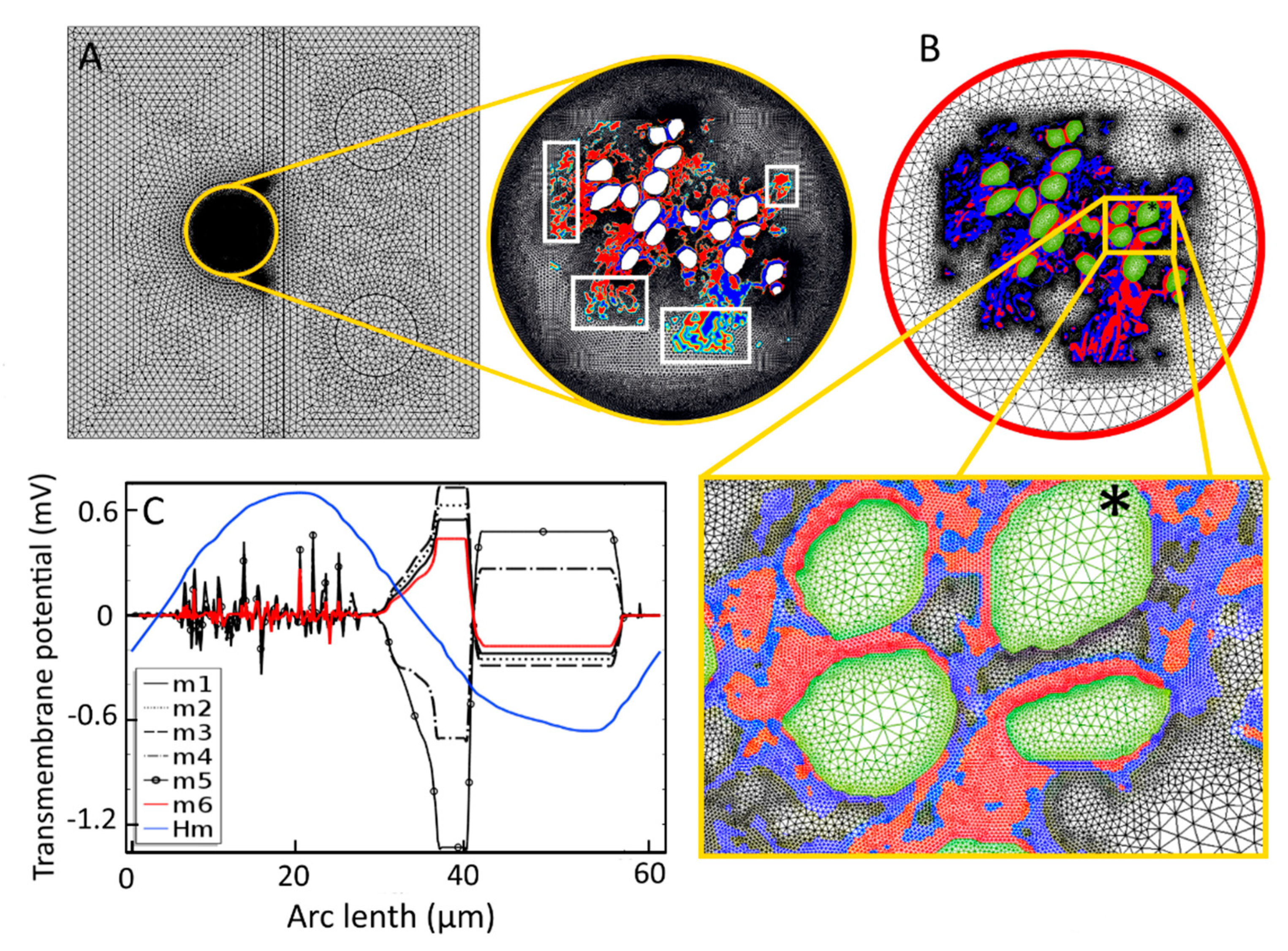
Appendix A. Mesh Convergence Study

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mesh Type | Max | Min | GR | CF | Res |
|---|---|---|---|---|---|
| Finer | 0.1 mm | 0.6 µm | 1.3 | 0.3 | 1 |
| Fine | 1 mm | 6 µm | 1.3 | 0.3 | 1 |
| Normal | 20 mm | 4 µm | 1.4 | 0.4 | 1 |

Appendix B. Activating Function


References
- WHO. 2018 Hearing Loss Estimates. Available online: https://www.who.int/deafness/estimates/en/ (accessed on 21 September 2020).
- Nance, W.E. The genetics of deafness. Ment. Retard. Dev. Disabil. Res. Rev. 2003, 9, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Hinojosa, R.; Lindsay, J.R. Profound deafness. Associated sensory and neural degeneration. Arch. Otolaryngol. 1980, 106, 193–209. [Google Scholar] [CrossRef] [PubMed]
- Hinojosa, R.; Marion, M. Histopathology of profound sensorineural deafness. Ann. N. Y. Acad. Sci. 1983, 405, 459–484. [Google Scholar] [CrossRef] [PubMed]
- Makary, C.A.; Shin, J.; Kujawa, S.G.; Liberman, M.C.; Merchant, S.N. Age-related primary cochlear neuronal degeneration in human temporal bones. Assoc. Res. Otolaryngol. 2011, 12, 711–717. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.S.; Dorman, M.F. Cochlear implants: A remarkable past and a brilliant future. Hear. Res. 2008, 242, 3–21. [Google Scholar] [CrossRef]
- Yawn, R.; Hunter, J.B.; Sweeney, A.D.; Bennett, M.L. Cochlear implantation: A biomechanical prosthesis for hearing loss. F1000Prime Rep. 2015, 7, 45. [Google Scholar] [CrossRef]
- Hochmair, I.; Hochmair, E.; Nopp, P.; Waller, M.; Jolly, C. Deep electrode insertion and sound coding in cochlear implants. Hear. Res. 2015, 322, 14–23. [Google Scholar] [CrossRef]
- Spelman, F.A. Cochlear electrode arrays: Past, present and future. Audiol. Neurootol. 2006, 11, 77–85. [Google Scholar] [CrossRef]
- Van Schoonhoven, J.; Sparreboom, M.; van Zanten, B.G.A.; Scholten, R.J.P.M.; Mylanus, E.A.M.; Dreschler, W.A.; Grolman, W.; Maat, B. The effectiveness of bilateral cochlear implants for severe-to-profound deafness in adults: A systematic review. Otol. Neurotol. 2013, 34, 190–198. [Google Scholar] [CrossRef]
- Sparreboom, M.; van Schoonhoven, J.; van Zanten, B.G.A.; Scholten, R.J.P.M.; Mylanus, E.A.M.; Grolman, W.; Maat, B. The effectiveness of bilateral cochlear implants for severe-to-profound deafness in children: A systematic review. Otol. Neurotol. 2010, 31, 1062–1071. [Google Scholar] [CrossRef]
- O’Donoghue, G. Cochlear implants—Science, serendipity, and success. N. Engl. J. Med. 2013, 369, 1190–1193. [Google Scholar] [CrossRef] [PubMed]
- Kalkman, R.K.; Briaire, J.J.; Frijns, J.H.M. Stimulation strategies and electrode design in computational models of the electrically stimulated cochlea: An overview of existing literature. Netw. Comput. Neural Syst. 2016, 27, 107–134. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Encke, J.; Obando-Leitón, M.; Weiß, R.; Schäfer, F.; Eberharter, J.; Böhnke, F.; Hemmert, W. Electrical stimulation in the human cochlea: A computational study based on high-resolution micro-CT scans. Front. Neurosci. 2019, 13, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kral, A.; Hartmann, R.; Mortazavi, D.; Klinke, R. Spatial resolution of cochlear implants: The electrical field and excitation of auditory afferents. Hear. Res. 1998, 121, 11–28. [Google Scholar] [CrossRef]
- Liu, W.; Edin, F.; Atturo, F.; Rieger, G.; Löwenheim, H.; Senn, P.; Blumer, M.; Schrott-Fisher, A.; Rask-Andersen, H.; Glueckert, R. The pre- and post- somatic segements of the human type I spiral ganglion neurons—Structural and functional considerations related to cochlear implantation. J. Neurosci. 2015, 284, 470–482. [Google Scholar] [CrossRef]
- Glueckert, R.; Pfaller, K.; Kinnefors, A.; Rask-Andersen, H.; Schrott-Fischer, A. The human spiral ganglion: New insights into ultrastructure, survival rate and implications for cochlear implants. Audiol. Neurotol. 2005, 10, 258–273. [Google Scholar] [CrossRef]
- Ota, C.Y.; Kimura, R.S. Ultrastructural study of the human spiral ganglion. Acta Otolaryngol. 1980, 89, 53–62. [Google Scholar] [CrossRef]
- Liu, W.; Glueckert, R.; Linthicum, F.H.; Rieger, G.; Blumer, M.; Bitsche, M.; Pechriggl, E.; Rask-Andersen, H.; Schrott-Fischer, A. Possible role of gap junction intercellular channels and connexin 43 in satellite glial cells (SGCs) for preservation of human spiral ganglion neurons: A comparative study with clinical implications. Cell Tissue Res. 2014, 355, 267–278. [Google Scholar] [CrossRef]
- Goldwyn, J.H.; Bierer, S.M.; Bierer, J.A. Modeling the electrode-neuron interface of cochlear implants: Effects of neural survival, electrode placement, and the partial tripolar configuration. Hear. Res. 2010, 268, 93–104. [Google Scholar] [CrossRef]
- Kang, S.; Chwodhury, T.; Moon, I.J.; Hong, S.H.; Yang, H.; Won, J.H.; Woo, J. Effects of electrode position on spatiotemporal auditory nerve fiber responses: A 3D computational model study. Comput. Math. Methods Med. 2015, 2015, 93438. [Google Scholar] [CrossRef]
- Rattay, F.; Potrusil, T.; Wenger, C.; Wise, A.K.; Glueckert, R.; Schrott-Fischer, A. Impact of morphometry, myelinization and synaptic current strength on spike conduction in human and cat spiral ganglion neurons. PLoS ONE 2013, 8, e79256. [Google Scholar] [CrossRef]
- Rattay, F.; Leao, R.N.; Felix, H. A model of the electrically excited human cochlear neuron. II. Influence of the three-dimensional cochlear structure on neural excitability. Hear. Res. 2001, 153, 64–79. [Google Scholar] [CrossRef]
- Miller, C.A.; Abbas, P.J.; Nourski, K.V.; Hu, N.; Robinson, B.K. Electrode configuration influences action potential initiation site and ensemble stochastic response properties. Hear. Res. 2003, 175, 200–214. [Google Scholar] [CrossRef]
- Bonnet, R.M.; Boermans, P.P.B.M.; Avenarius, O.F.; Briaire, J.J.; Frijns, J.H.M. Effects of pulse width, pulse rate and paired electrode stimulation on psychophysical measures of dynamic range and speech recognition in cochlear implants. Ear Hear. 2012, 33, 489–496. [Google Scholar] [CrossRef]
- Cohen, L.T. Practical model description of peripheral neural excitation in cochlear implant recipients: 4. Model development at low pulse rates: General model and application to individuals. Hear. Res. 2009, 248, 15–30. [Google Scholar] [CrossRef]
- Woo, J.; Miller, C.A.; Abbas, P.J. Simulation of the electrically stimulated cochlear neuron: Modeling adaptation to trains of electric pulses. IEEE Trans. Biomed. Eng. 2009, 56, 1348–1359. [Google Scholar] [CrossRef]
- Nogueira, W.; Schurzig, D.; Büchner, A.; Penninger, R.T.; Würfel, W. Validation of a cochlear implant patient-specific model of the voltage distribution in a clinical setting. Front. Bioeng. Biotechnol. 2016, 4, 1–16. [Google Scholar] [CrossRef]
- Micco, A.G.; Richter, C.-P. Tissue resistivities determine the current flow in the cochlea. Curr. Opin. Otolaryngol. Head Neck Surg. 2006, 14, 352–355. [Google Scholar] [CrossRef]
- Malherbe, T.K.; Hanekom, T.; Hanekom, J.J. The effect of the resistive properties of bone on neural excitation and electric fields in cochlear implant models. Hear. Res. 2015, 327, 126–135. [Google Scholar] [CrossRef]
- Finley, C.C.; Wilson, B.S.; White, M.W. Models of Neural Responsiveness to Electrical Stimulation. In Cochlear Implants: Models of the Electrically Stimulated Ear; Miller, J.M., Spelman, F.A., Eds.; Springer: New York, NY, USA, 1990; pp. 55–96. ISBN 978-1-4612-3256-8. [Google Scholar]
- Rattay, F.; Lutter, P.; Felix, H. A model of the electrically excited human cochlear neuron. I. Contribution of neural substructures to the generation and propagation of spikes. Hear. Res. 2001, 153, 43–63. [Google Scholar] [CrossRef]
- Kalkman, R.K.; Briaire, J.J.; Frijns, J.H.M. Current focussing in cochlear implants: An analysis of neural recruitment in a computational model. Hear. Res. 2015, 322, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Sriperumbudur, K.K.; Pau, H.W.; van Rienen, U. Effect of tissue heterogeneity on the transmembrane potential of type-1 spiral ganglion neurons: A simulation study. IEEE Trans. Biomed. Eng. 2018, 65, 658–668. [Google Scholar] [CrossRef] [PubMed]
- Hodgkin, A.L.; Huxley, A.F. A quantitative description of membrane current and its application to conduction and excitation in nerve. Bull. Math. Biol. 1990, 52, 25–71. [Google Scholar] [CrossRef]
- Hodgkin, A.L. The membrane resistance of a non-medullated nerve fibre. J. Physiol. 1947, 106, 305–318. [Google Scholar] [CrossRef]
- Plonsey, R.; Barr, R.C. Electric field stimulation of excitable tissue. IEEE Eng. Med. Biol. Mag. 1998, 17, 130–137. [Google Scholar] [CrossRef]
- Sagers, J.E.; Landegger, L.D.; Worthington, S.; Nadol, J.B.; Stankovic, K.M. Human cochlear histopathology reflects clinical signatures of primary neural degeneration. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Felix, H.; Pollak, A.; Gleeson, M.; Johnsson, L.-G. Degeneration pattern of human first-order cochlear neurons. Adv. Otorhinolaryngol. 2002, 59, 116–123. [Google Scholar]
- Xing, Y.; Samuvel, D.J.; Stevens, S.M.; Dubno, J.R.; Schulte, B.A.; Lang, H. Age-related changes of myelin basic protein in mouse and human auditory nerve. PLoS ONE 2012, 7, e34500. [Google Scholar] [CrossRef]
- Wu, P.Z.; Liberman, L.D.; Bennett, K.; de Gruttola, V.; O’Malley, J.T.; Liberman, M.C. Primary neural degeneration in the human cochlea: Evidence for hidden hearing loss in the aging ear. Neuroscience 2019, 407, 8–20. [Google Scholar] [CrossRef]
- Pfingst, B.E.; Zhou, N.; Colesa, D.J.; Watts, M.M.; Strahl, S.B.; Garadat, S.N.; Schvartz-Leyzac, K.C.; Budenz, C.L.; Raphael, Y.; Zwolan, T.A. Importance of cochlear health for implant function. Hear. Res. 2015, 322, 77–88. [Google Scholar] [CrossRef]
- Lundin, K.; Stillesjö, F.; Rask-Andersen, H. Experiences and results from cochlear implantation in patients with long duration of deafness. Audiol. Neurotol. Extra 2014, 4, 46–55. [Google Scholar] [CrossRef]
- Sly, D.J.; Heffer, L.F.; White, M.W.; Shepherd, R.K.; Birch, M.G.J.; Minter, R.L.; Nelson, N.E.; Wise, A.K.; O’Leary, S.J. Deafness alters auditory nerve fibre responses to cochlear implant stimulation. Eur. J. Neurosci. 2007, 26, 510–522. [Google Scholar] [CrossRef]
- Nadol, J.B., Jr. Degeneration of cochlear neurons as seen in the spiral ganglion of man. Hear. Res. 1990, 49, 141–154. [Google Scholar] [CrossRef]
- Liberman, M.C. Hidden hearing loss: Primary neural degeneration in the noise-damaged and aging cochlea. Acoust. Sci. Technol. 2020, 41, 59–62. [Google Scholar] [CrossRef]
- Zimmermann, C.E.; Burgess, B.J.; Nadol, J.B. Patterns of degeneration in the human cochlear nerve. Hear. Res. 1995, 90, 192–201. [Google Scholar] [CrossRef]
- Schwieger, J.; Warnecke, A.; Lenarz, T.; Esser, K.H.; Scheper, V.; Forsythe, J. Neuronal survival, morphology and outgrowth of spiral ganglion neurons using a defined growth factor combination. PLoS ONE 2015, 10, e0133680. [Google Scholar] [CrossRef]
- Cartee, L.A. Spiral ganglion cell site of excitation II: Numerical model analysis. Hear. Res. 2006, 215, 22–30. [Google Scholar] [CrossRef]
- Cartee, L.A.; Miller, C.A.; van den Honert, C. Spiral ganglion cell site of excitation I: Comparison of scala tympani and intrameatal electrode responses. Hear. Res. 2006, 215, 10–21. [Google Scholar] [CrossRef]
- Hossain, W.A.; Antic, S.D.; Yang, Y.; Rasband, M.N.; Morest, D.K. Where is the spike generator of the cochlear nerve? Voltage-gated sodium channels in the mouse cochlea. J. Neurosci. 2005, 25, 6857–6868. [Google Scholar] [CrossRef]
- Rattay, F.; Danner, S.M. Peak I of the human auditory brainstem response results from the somatic regions of type I spiral ganglion cells: Evidence from computer modeling. Hear. Res. 2014, 315, 67–79. [Google Scholar] [CrossRef]
- Pelliccia, P.; Venail, F.; Bonafé, A.; Makeieff, M.; Iannetti, G.; Bartolomeo, M.; Mondain, M. Cochlea size variability and implications in clinical practice. Acta Otorhinolaryngol. Ital. 2014, 34, 42–49. [Google Scholar] [PubMed]
- Rask-Andersen, H.; Erixon, E.; Kinnefors, A.; Löwenheim, H.; Schrott-Fischer, A.; Liu, W. Anatomy of the human cochlea—Implications for cochlear implantation. Cochlear Implants Int. 2011, 12, S13–S18. [Google Scholar] [CrossRef] [PubMed]
- Erixon, E.; Högstorp, H.; Wadin, K.; Rask-Andersen, H. Variational anatomy of the human cochlea: Implications for cochlear implantation. Otol. Neurotol. 2009, 30, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Meng, F.; Cai, J.; Lu, S. Beyond pixels: A comprehensive survey from bottom-up to semantic image segmentation and cosegmentation. J. Vis. Commun. Image Represent. 2016, 34, 12–27. [Google Scholar] [CrossRef]
- Mossop, B.J.; Barr, R.C.; Zaharoff, D.A.; Yuan, F. Electric fields within cells as a function of membrane resistivity—A model study. IEEE Trans. Nanobiosci. 2004, 3, 225–231. [Google Scholar] [CrossRef]
- Klee, M.; Plonsey, R. Extracellular stimulation of a cell having a non-uniform membrane. IEEE Trans. Biomed. Eng. 1974, 21, 452–460. [Google Scholar] [CrossRef]
- Miranda, P.C.; Correia, L.; Salvador, R.; Basser, P.J. Tissue heterogeneity as a mechanism for localized neural stimulation by applied electric fields. Phys. Med. Biol. 2007, 52, 5603–5617. [Google Scholar] [CrossRef]
- Aslanidi, O.V.; Colman, M.A.; Varela, M.; Zhao, J.; Smaill, B.H.; Hancox, J.C.; Boyett, M.R.; Zhang, H. Heterogeneous and anisotropic integrative model of pulmonary veins: Computational study of arrhythmogenic substrate for atrial fibrillation. Interface Focus 2013, 3, 20120069. [Google Scholar] [CrossRef]
- Golberg, A.; Bruinsma, B.G.; Uygun, B.E.; Yarmush, M.L. Tissue heterogeneity in structure and conductivity contribute to cell survival during irreversible electroporation ablation by “electric field sinks”. Sci. Rep. 2015, 5, 8485. [Google Scholar] [CrossRef]
- Pucihar, G.; Kotnik, T.; Valič, B.; Miklavčič, D. Numerical determination of transmembrane voltage induced on irregularly shaped cells. Ann. Biomed. Eng. 2006, 34, 642–652. [Google Scholar] [CrossRef]
- Spoendlin, H.; Schrott-Fischer, A. Analysis of the human auditory nerve. Hear. Res. 1989, 43, 25–38. [Google Scholar] [CrossRef]
- Ho, S.Y.; Wiet, R.J.; Richter, C.-P. Modifying cochlear implant design: Advantages of placing a return electrode in the modiolus. Otol. Neurotol. 2004, 25, 497–503. [Google Scholar] [CrossRef]
- Frijns, J.H.M.; Briaire, J.J.; Grote, J.J. The importance of human cochlear anatomy for the results of modiolus-hugging multichannel cochlear implants. Otol. Neurotol. 2001, 22, 340–349. [Google Scholar] [CrossRef]
- Holden, L.K.; Firszt, J.B.; Reeder, R.M.; Uchanski, R.M.; Dwyer, N.Y.; Holden, T.A. Factors affecting outcomes in cochlear implant recipients implanted with a perimodiolar electrode array located in scala tympani. Otol. Neurotol. 2016, 37, 1662–1668. [Google Scholar] [CrossRef]
- Kole, M.H.P.; Ilschner, S.U.; Kampa, B.M.; Williams, S.R.; Ruben, P.C.; Stuart, G.J. Action potential generation requires a high sodium channel density in the axon initial segment. Nat. Neurosci. 2008, 11, 178–186. [Google Scholar] [CrossRef]
- Van der Marel, K.S.; Briaire, J.J.; Verbist, B.M.; Muurling, T.J.; Frijns, J.H.M. The influence of cochlear implant electrode position on performance. Audiol. Neurotol. 2015, 20, 202–211. [Google Scholar] [CrossRef]
- Kós, M.I.; Boëx, C.; Sigrist, A.; Guyot, J.-P.; Pelizzone, M. Measurements of electrode position inside the cochlea for different cochlear implant systems. Acta Otolaryngol. 2005, 125, 474–480. [Google Scholar] [CrossRef]
- Skinner, M.W.; Ketten, D.R.; Holden, L.K.; Harding, G.W.; Smith, P.G.; Gates, G.A.; Neely, J.G.; Kletzker, G.R.; Brunsden, B.; Blocker, B. CT-derived estimation of cochlear morphology and electrode array position in relation to word recognition in Nucleus-22 recipients. J. Assoc. Res. Otolaryngol. 2002, 3, 332–350. [Google Scholar] [CrossRef]
- Rattay, F. The basic mechanism for the electrical stimulation of the nervous system. Neuroscience 1999, 89, 335–346. [Google Scholar] [CrossRef]
- Lai, W.D.; Choi, C.T.M. Incorporating the electrode-tissue interface to cochlear implant models. IEEE Trans. Magn. 2007, 43, 1721–1724. [Google Scholar] [CrossRef]
- Choi, C.T.M.; Lai, W.-D.; Chen, Y.-B. Optimization of cochlear implant electrode array using genetic algorithms and computational neuroscience models. IEEE Trans. Magn. 2004, 40, 639–642. [Google Scholar] [CrossRef]
- Rattay, F. Basics of hearing theory and noise in cochlear implants. Chaos Solitons Fractals 2000, 11, 1875–1884. [Google Scholar] [CrossRef]
- Altman, K.W.; Plonsey, R. Analysis of excitable cell activation: Relative effects of external electrical stimuli. Med. Biol. Eng. Comput. 1990, 28, 574–580. [Google Scholar] [CrossRef]
- Joucla, S.; Yvert, B. The “mirror” estimate: An intuitive predictor of membrane polarization during extracellular stimulation. Biophys. J. 2009, 96, 3495–3508. [Google Scholar] [CrossRef]
- Agudelo-Toro, A.; Neef, A. Computationally efficient simulation of electrical activity at cell membranes interacting with self-generated and externally imposed electric fields. J. Neural Eng. 2013, 10, 026019. [Google Scholar] [CrossRef]
- Appali, R.; Sriperumbudur, K.K.; van Rienen, U. Extracellular stimulation of neural tissues: Activating function and sub-threshold potential perspective. In Proceedings of the Annual International Conference of the IEEE Engineering in Medicine and Biology Society, EMBS, Berlin, Germany, 23–27 July 2019. [Google Scholar]
- Rask-Andersen, H.; Liu, W. Auditory nerve preservation and regeneration in man: Relevance for cochlear implantation. Neural Regen. Res. 2015, 10, 710. [Google Scholar] [CrossRef]
- Hahnewald, S.; Roccio, M.; Tscherter, A.; Streit, J.; Ambett, R.; Senn, P. Spiral ganglion neuron explant culture and electrophysiology on multi electrode arrays. J. Vis. Exp. 2016, e54538. [Google Scholar] [CrossRef]
- Brill, S.; Müller, J.; Hagen, R.; Möltner, A.; Brockmeier, S.J.; Stark, T.; Helbig, S.; Maurer, J.; Zahnert, T.; Zierhofer, C.; et al. Site of cochlear stimulation and its effect on electrically evoked compound action potentials using the MED-EL standard electrode array. Biomed. Eng. Online 2009, 8. [Google Scholar] [CrossRef]
- Kashio, A.; Tejani, V.D.; Scheperle, R.A.; Brown, C.J.; Abbas, P.J. Exploring the source of neural responses of different latencies obtained from different recording electrodes in cochlear implant users. Audiol. Neurotol. 2016, 21, 141–149. [Google Scholar] [CrossRef]
- Smit, J.E.; Hanekom, T.; Hanekom, J.J. Predicting action potential characteristics of human auditory nerve fibres through modification of the Hodgkin-Huxley equations. S. Afr. J. Sci. 2008, 104, 284–292. [Google Scholar]
- Pisoni, D.B.; Kronenberger, W.G.; Harris, M.S.; Moberly, A.C. Three challenges for future research on cochlear implants. World J. Otorhinolaryngol. Head Neck Surg. 2017, 3, 240–254. [Google Scholar] [CrossRef]
- Hanekom, T. Modelling encapsulation tissue around cochlear implant electrodes. Med. Biol. Eng. Comput. 2005, 43, 47–55. [Google Scholar] [CrossRef]
- Choong, J.K.; Hampson, A.J.; Brody, K.M.; Lo, J.; Bester, C.W.; Gummer, A.W.; Reynolds, N.P.; O’Leary, S.J. Nanomechanical mapping reveals localized stiffening of the basilar membrane after cochlear implantation. Hear. Res. 2020, 385. [Google Scholar] [CrossRef]
- Lazard, D.S.; Vincent, C.; Venail, F.; van de Heyning, P.; Truy, E.; Sterkers, O.; Skarzynski, P.H.; Skarzynski, H.; Schauwers, K.; O’Leary, S.; et al. Pre-, per- and postoperative factors affecting performance of postlinguistically deaf adults using cochlear implants: A new conceptual model over time. PLoS ONE 2012, 7, e48739. [Google Scholar] [CrossRef]
- Boisvert, I.; Reis, M.; Au, A.; Cowan, R.; Dowell, R.C. Cochlear implantation outcomes in adults: A scoping review. PLoS ONE 2020, 15, e0232421. [Google Scholar] [CrossRef]
- Moberly, A.C.; Bates, C.; Harris, M.S.; Pisoni, D.B. The enigma of poor performance by adults with cochlear implants. Otol. Neurotol. 2016, 37, 1522–1528. [Google Scholar] [CrossRef]
- Mangado, N.; Ceresa, M.; Benav, H.; Mistrik, P.; Piella, G.; González Ballester, M.A. Towards a complete in-silico assessment of the outcome of cochlear implantation surgery. Mol. Neurobiol. 2018, 55, 173–186. [Google Scholar] [CrossRef]
- Mangado, N.; Pons-Prats, J.; Coma, M.; Mistrík, P.; Piella, G.; Ceresa, M.; González Ballester, M.Á. Computational evaluation of cochlear implant surgery outcomes accounting for uncertainty and parameter variability. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef]
- Evans, A.J.; Thompson, B.C.; Wallace, G.G.; Millard, R.; O’Leary, S.J.; Clark, G.M.; Shepherd, R.K.; Richardson, R.T. Promoting neurite outgrowth from spiral ganglion neuron explants using polypyrrole/BDNF-coated electrodes. J. Biomed. Mater. Res. Part A 2009, 91, 241–250. [Google Scholar] [CrossRef]
- Cai, Y.; Edin, F.; Jin, Z.; Alexsson, A.; Gudjonsson, O.; Liu, W.; Rask-Andersen, H.; Karlsson, M.; Li, H. Strategy towards independent electrical stimulation from cochlear implants: Guided auditory neuron growth on topographically modified nanocrystalline diamond. Acta Biomater. 2016, 31, 211–220. [Google Scholar] [CrossRef]
- Whitlon, D.S.; Grover, M.; Dunne, S.F.; Richter, S.; Luan, C.-H.; Richter, C.-P. Novel high content screen detects compounds that promote neurite regeneration from cochlear spiral ganglion neurons. Sci. Rep. 2015, 5, 15960. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.-Z.; He, Y.; Jiang, X.-W.; Chen, F.-Q.; Chen, Y.; Shi, L.; Chen, J.; Chen, X.; Li, X.; Xue, T.; et al. Stem cell transplantation via the cochlear lateral wall for replacement of degenerated spiral ganglion neurons. Hear. Res. 2013, 298, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Snyder, R.L.; Middlebrooks, J.C.; Bonham, B.H. Cochlear implant electrode configuration effects on activation threshold and tonotopic selectivity. Hear. Res. 2008, 235, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Tang, Q.; Zeng, F.-G.G.; Guan, T.; Ye, D. Cochlear implant spatial selectivity with monopolar, bipolar and tripolar stimulation. Hear. Res. 2012, 283, 45–58. [Google Scholar] [CrossRef]
- Akil, O.; Sun, Y.; Vijayakumar, S.; Zhang, W.; Ku, T.; Lee, C.-K.; Jones, S.; Grabowski, G.A.; Lustig, L.R. Spiral ganglion degeneration and hearing loss as a consequence of satellite cell death in saposin B-deficient mice. J. Neurosci. 2015, 35, 3263–3275. [Google Scholar] [CrossRef]
- Matsumura, R.; Yamamoto, H.; Niwano, M.; Hirano-Iwata, A. An electrically resistive sheet of glial cells for amplifying signals of neuronal extracellular recordings. Appl. Phys. Lett. 2016, 108, 023701. [Google Scholar] [CrossRef]
- Cheng, A.H.-D.; Cheng, D.T. Heritage and early history of the boundary element method. Eng. Anal. Bound. Elem. 2005, 29, 268–302. [Google Scholar] [CrossRef]
- Plonsey, R.; Heppner, D.B. Considerations of quasi-stationarity in electrophysiological systems. Bull. Math. Biophys. 1967, 29, 657–664. [Google Scholar] [CrossRef]
- Van Rienen, U.; Flehr, J.; Schreiber, U.; Schulze, S.; Gimsa, U.; Baumann, W.; Weiss, D.G.; Gimsa, J.; Benecke, R.; Pau, H.W. Electro-quasistatic simulations in bio-systems engineering and medical engineering. Adv. Radio Sci. 2005, 3, 39–49. [Google Scholar] [CrossRef]
- Amestoy, P.R.; Duff, I.; L’Excellent, J.-Y. Multifrontal parallel distributed symmetric and unsymmetric solvers. Comput. Methods Appl. Mech. Eng. 2000, 184, 501–520. [Google Scholar] [CrossRef]






| Subdomain | Dimensions |
|---|---|
| Temporal bone | 0.5 cm × 0.5 cm |
| Modiolus bone | 100 µm × 2 mm |
| Electrode | 0.3 mm (diameter) |
| Scala tympani | 1 mm × 1 mm |
| Rosenthal’s canal | 250 µm (diameter) |
| Spiral ganglion neurons | 20–30 µm (diameter) |
| Subdomain | Electric Conductivity (S/m) |
|---|---|
| Temporal bone | 0.016 |
| Modiolus bone | 0.0334 |
| Electrode | 9 × 106 |
| Tympanic medium | 1.43 |
| GroupA tissues | 4 × 10−9 |
| Intracellular medium | 0.31 |
| Extracellular medium | 1.2 |
| GroupB tissues | 3.45 × 10−6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sriperumbudur, K.K.; Appali, R.; Gummer, A.W.; van Rienen, U. Neural Tissue Degeneration in Rosenthal’s Canal and Its Impact on Electrical Stimulation of the Auditory Nerve by Cochlear Implants: An Image-Based Modeling Study. Int. J. Mol. Sci. 2020, 21, 8511. https://doi.org/10.3390/ijms21228511
Sriperumbudur KK, Appali R, Gummer AW, van Rienen U. Neural Tissue Degeneration in Rosenthal’s Canal and Its Impact on Electrical Stimulation of the Auditory Nerve by Cochlear Implants: An Image-Based Modeling Study. International Journal of Molecular Sciences. 2020; 21(22):8511. https://doi.org/10.3390/ijms21228511
Chicago/Turabian StyleSriperumbudur, Kiran Kumar, Revathi Appali, Anthony W. Gummer, and Ursula van Rienen. 2020. "Neural Tissue Degeneration in Rosenthal’s Canal and Its Impact on Electrical Stimulation of the Auditory Nerve by Cochlear Implants: An Image-Based Modeling Study" International Journal of Molecular Sciences 21, no. 22: 8511. https://doi.org/10.3390/ijms21228511
APA StyleSriperumbudur, K. K., Appali, R., Gummer, A. W., & van Rienen, U. (2020). Neural Tissue Degeneration in Rosenthal’s Canal and Its Impact on Electrical Stimulation of the Auditory Nerve by Cochlear Implants: An Image-Based Modeling Study. International Journal of Molecular Sciences, 21(22), 8511. https://doi.org/10.3390/ijms21228511