Role of Conserved Residues and F322 in the Extracellular Vestibule of the Rat P2X7 Receptor in Its Expression, Function and Dye Uptake Ability

 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Results
2.1. Electrophysiological Characterization of Alanine Mutants of Conserved and Unique Amino Acid Residues in the Extracellular Vestibule of Rat P2X7
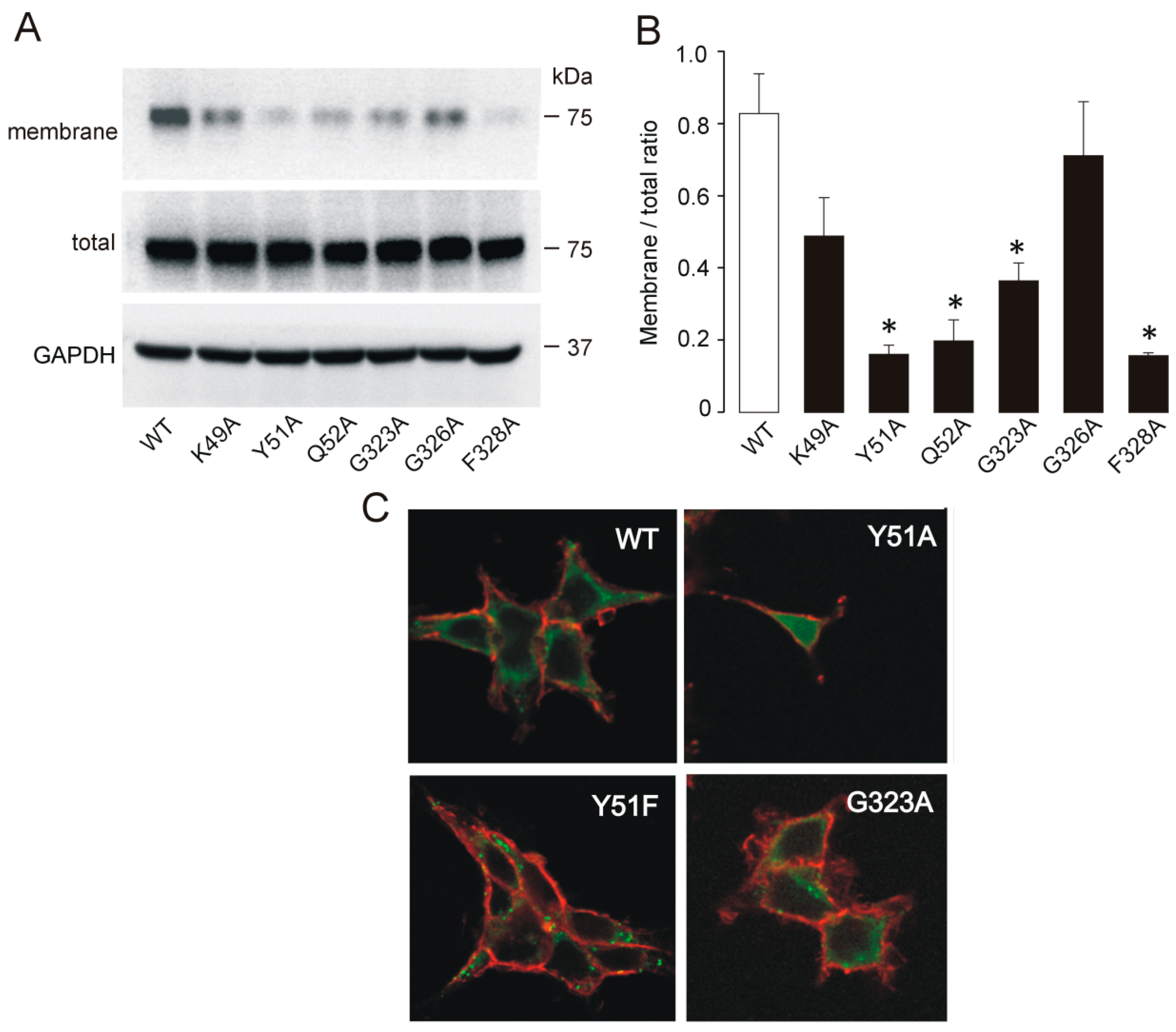
2.2. Membrane Expression of Nonfunctional and Low-Functioning Mutants
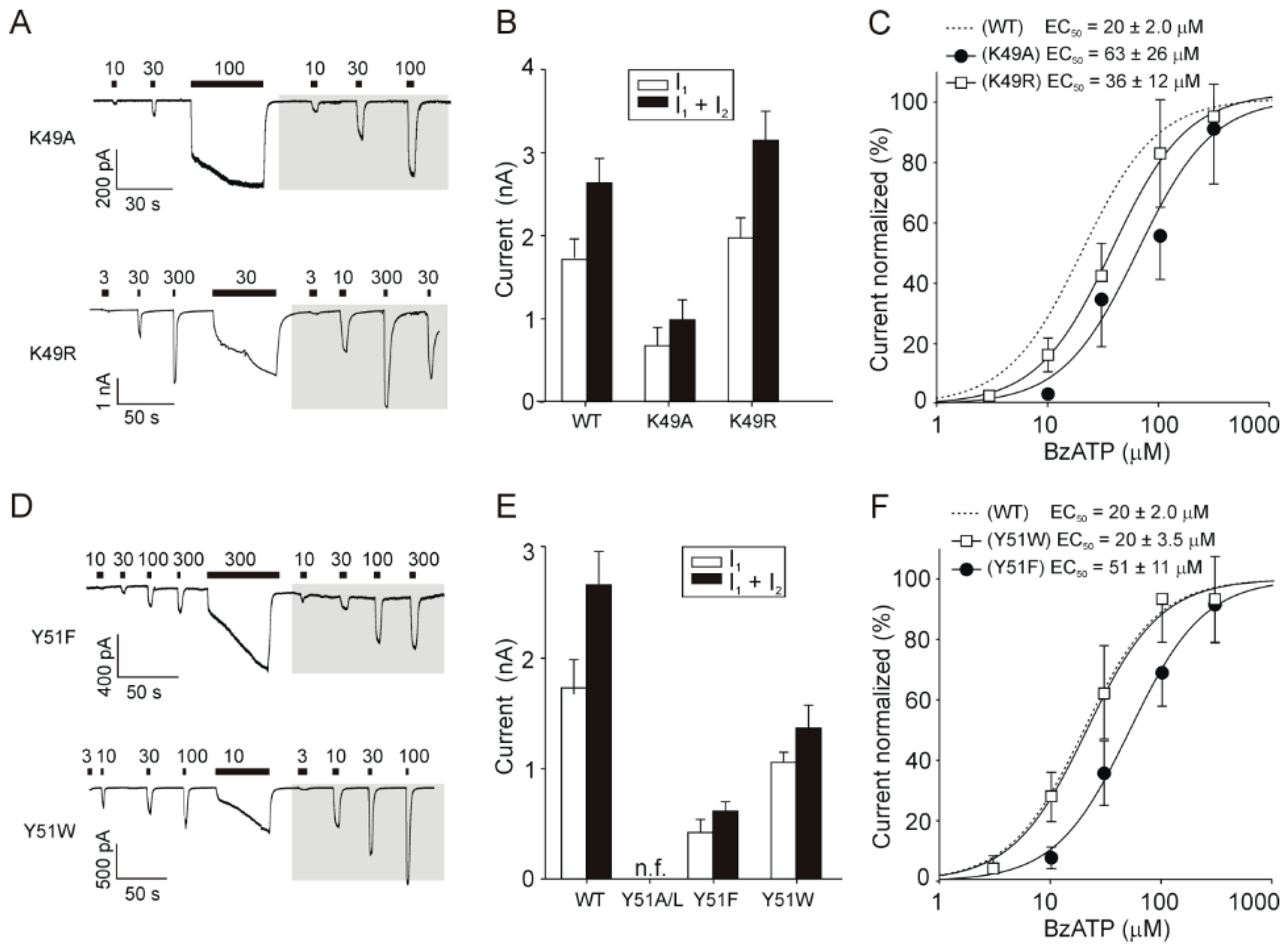
2.3. Rescue of Receptor Function by Replacing K49 with a Positively Charged Residue
2.4. Rescue of the Receptor Function by Replacing Y51 with Aromatic Residues
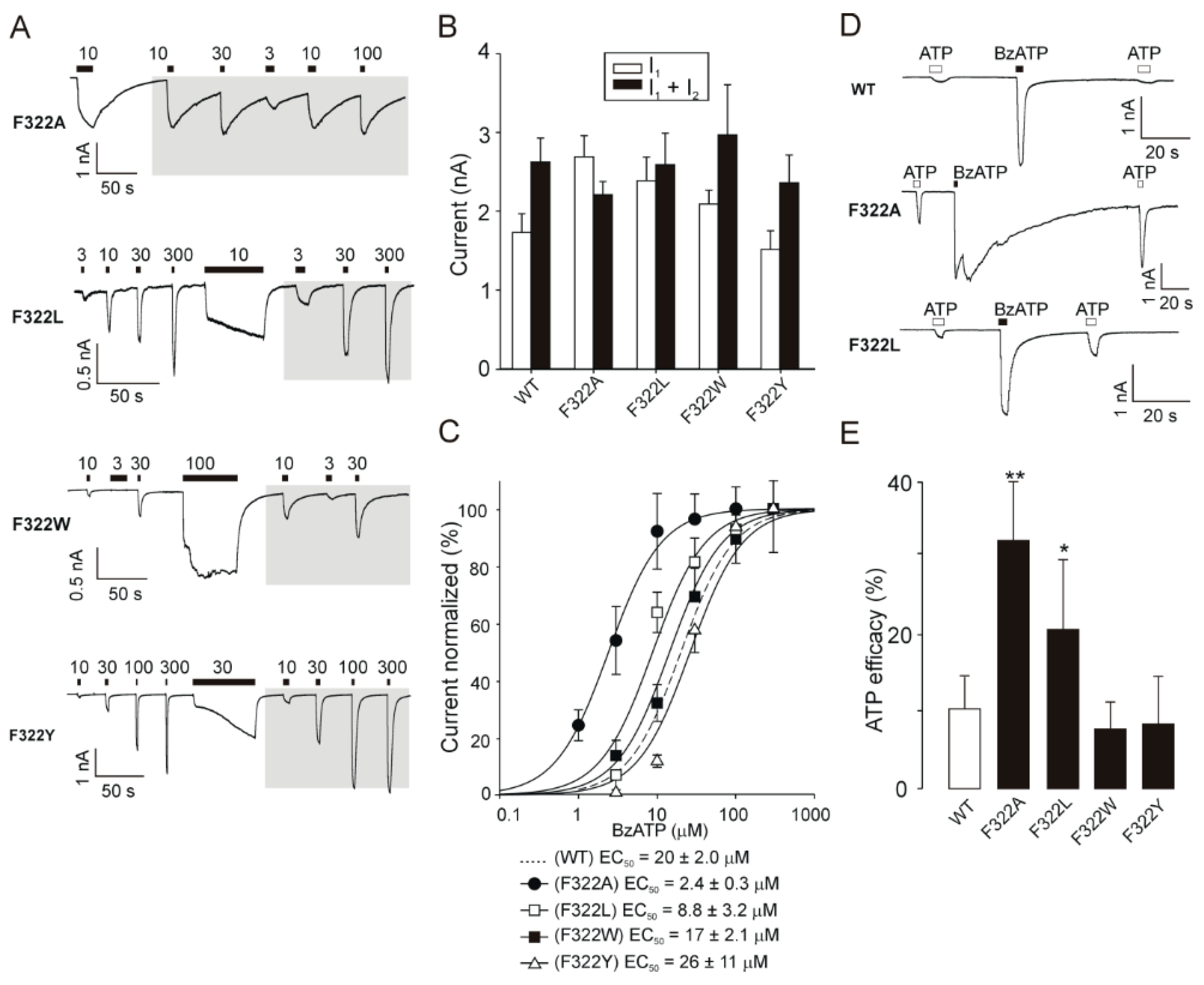
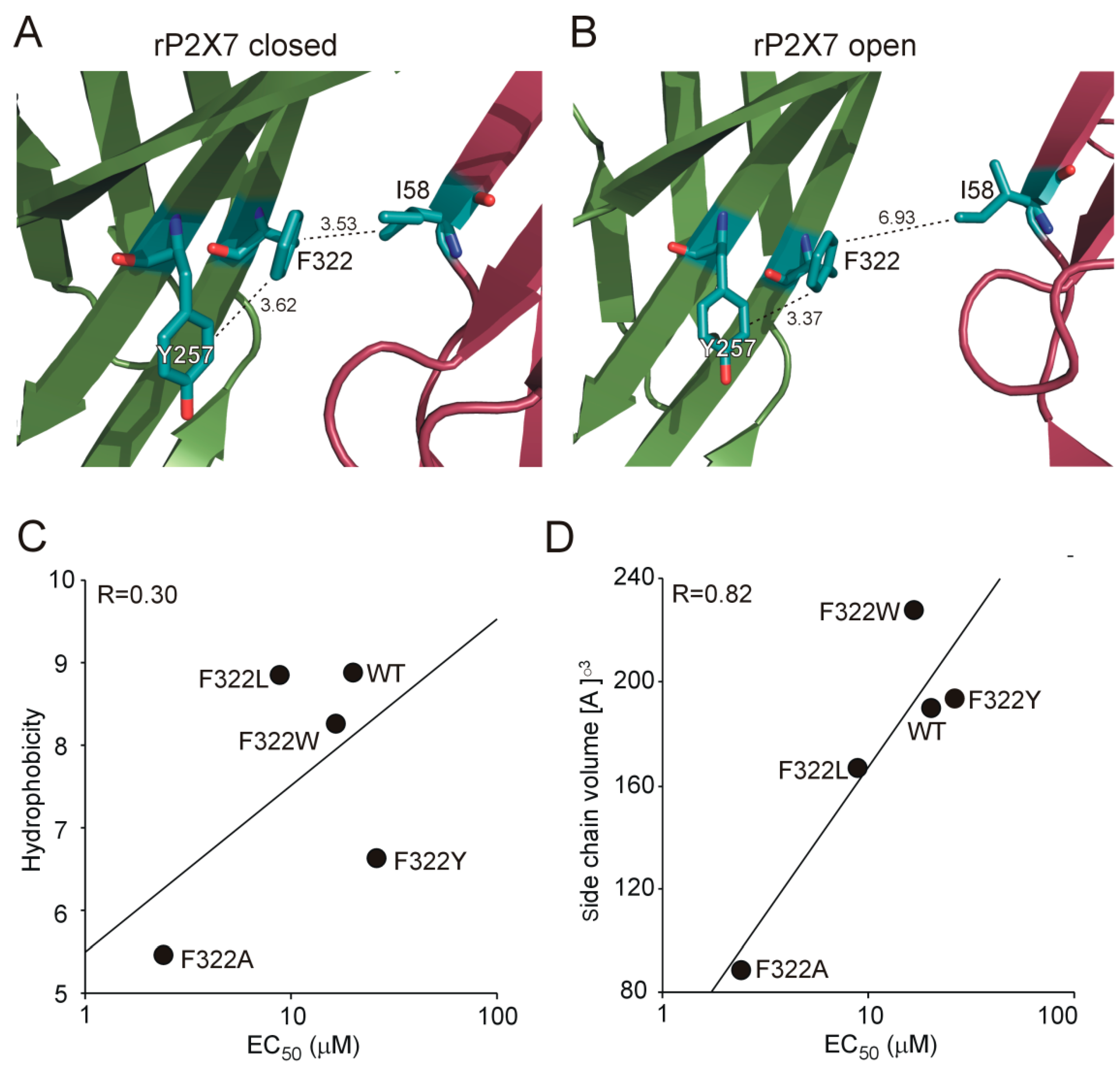
2.5. Recovery of Normal Receptor Function by Replacing F322 with Hydrophobic/Aromatic Residues
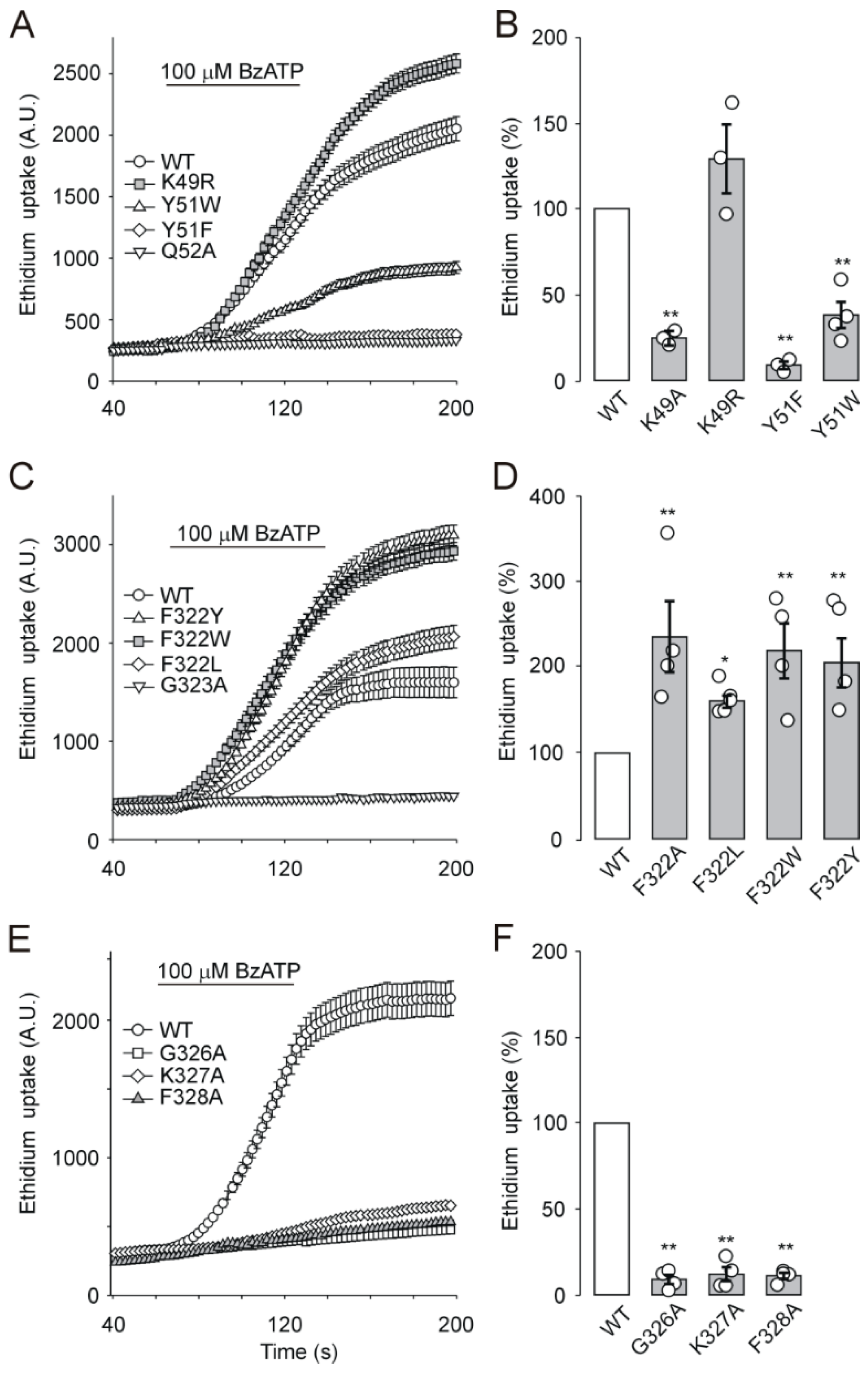
2.6. EtBr Accumulation by Cells Expressing Extracellular Vestibule Mutants
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Transfection
4.2. DNA Constructs
4.3. Whole-Cell Patch-Clamp Recording
4.4. Ethidium Bromide Uptake
4.5. Cell-Surface Biotinylation Assay
4.6. Membrane Expression of YFP-Tagged Receptors
4.7. Calculations
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ATP | Adenosine 5′-triphosphate |
| BzATP | 2′,3′-O-(Benzoyl-4-benzoyl)-ATP |
| EC50 | Half maximal effective concentration |
| HEK293T | Human embryonic kidney 293T |
| TM1 | Transmembrane domain 1 |
| TM2 | Transmembrane domain 2 |
| WT | Wild-type |
References
- Adinolfi, E.; Pizzirani, C.; Idzko, M.; Panther, E.; Norgauer, J.; Di Virgilio, F.; Ferrari, D. P2X(7) receptor: Death or life? Purinergic Signal. 2005, 1, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Pathophysiology and therapeutic potential of purinergic signaling. Pharmacol. Rev. 2006, 58, 58–86. [Google Scholar] [CrossRef] [PubMed]
- Surprenant, A.; North, R.A. Signaling at purinergic P2X receptors. Annu. Rev. Physiol. 2009, 71, 333–359. [Google Scholar] [CrossRef] [PubMed]
- Stojilkovic, S.S. Purinergic regulation of hypothalamopituitary functions. Trends Endocrinol. Metab. 2009, 20, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Khakh, B.S.; North, R.A. Neuromodulation by extracellular ATP and P2X receptors in the CNS. Neuron 2012, 76, 51–69. [Google Scholar] [CrossRef] [PubMed]
- Coddou, C.; Yan, Z.; Obsil, T.; Huidobro-Toro, J.P.; Stojilkovic, S.S. Activation and regulation of purinergic P2X receptor channels. Pharmacol. Rev. 2011, 63, 641–683. [Google Scholar] [CrossRef]
- Surprenant, A.; Rassendren, F.; Kawashima, E.; North, R.A.; Buell, G. The cytolytic P2Z receptor for extracellular ATP identified as a P2X receptor (P2X7). Science 1996, 272, 735–738. [Google Scholar] [CrossRef]
- Rassendren, F.; Buell, G.N.; Virginio, C.; Collo, G.; North, R.A.; Surprenant, A. The permeabilizing ATP receptor, P2X7. Cloning and expression of a human cDNA. J. Biol. Chem. 1997, 272, 5482–5486. [Google Scholar] [CrossRef]
- Kopp, R.; Krautloher, A.; Ramirez-Fernandez, A.; Nicke, A. P2X7 Interactions and Signaling—Making Head or Tail of It. Front. Mol. Neurosci. 2019, 12, 183. [Google Scholar] [CrossRef]
- Di Virgilio, F.; Dal Ben, D.; Sarti, A.C.; Giuliani, A.L.; Falzoni, S. The P2X7 Receptor in Infection and Inflammation. Immunity 2017, 47, 15–31. [Google Scholar] [CrossRef]
- Vavra, V.; Bhattacharya, A.; Zemkova, H. Facilitation of glutamate and GABA release by P2X receptor activation in supraoptic neurons from freshly isolated rat brain slices. Neuroscience 2011, 188, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Illes, P.; Khan, T.M.; Rubini, P. Neuronal P2X7 Receptors Revisited: Do They Really Exist? J. Neurosci. 2017, 37, 7049–7062. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek-Hajek, K.; Zhang, J.; Kopp, R.; Grosche, A.; Rissiek, B.; Saul, A.; Bruzzone, S.; Engel, T.; Jooss, T.; Krautloher, A.; et al. Re-evaluation of neuronal P2X7 expression using novel mouse models and a P2X7-specific nanobody. eLife 2018, 7. [Google Scholar] [CrossRef]
- Burnstock, G. P2X ion channel receptors and inflammation. Purinergic Signal. 2016, 12, 59–67. [Google Scholar] [CrossRef]
- Klaft, Z.J.; Schulz, S.B.; Maslarova, A.; Gabriel, S.; Heinemann, U.; Gerevich, Z. Extracellular ATP differentially affects epileptiform activity via purinergic P2X7 and adenosine A1 receptors in naive and chronic epileptic rats. Epilepsia 2012, 53, 1978–1986. [Google Scholar] [CrossRef]
- Nurkhametova, D.; Kudryavtsev, I.; Guselnikova, V.; Serebryakova, M.; Giniatullina, R.R.; Wojciechowski, S.; Tore, F.; Rizvanov, A.; Koistinaho, J.; Malm, T.; et al. Activation of P2X7 Receptors in Peritoneal and Meningeal Mast Cells Detected by Uptake of Organic Dyes: Possible Purinergic Triggers of Neuroinflammation in Meninges. Front. Cell. Neurosci. 2019, 13, 45. [Google Scholar] [CrossRef]
- Sperlagh, B.; Illes, P. P2X7 receptor: An emerging target in central nervous system diseases. Trends Pharmacol. Sci. 2014, 35, 537–547. [Google Scholar] [CrossRef]
- Stokes, L.; Spencer, S.J.; Jenkins, T.A. Understanding the role of P2X7 in affective disorders-are glial cells the major players? Front. Cell. Neurosci. 2015, 9, 258. [Google Scholar] [CrossRef]
- Garre, J.M.; Silva, H.M.; Lafaille, J.J.; Yang, G. P2X7 receptor inhibition ameliorates dendritic spine pathology and social behavioral deficits in Rett syndrome mice. Nat. Commun. 2020, 11, 1784. [Google Scholar] [CrossRef]
- Koldej, R.M.; Perera, T.; Collins, J.; Ritchie, D.S. Association between P2X7 Polymorphisms and Post-Transplant Outcomes in Allogeneic Haematopoietic Stem Cell Transplantation. Int. J. Mol. Sci. 2020, 21, 3772. [Google Scholar] [CrossRef]
- Wang, X.; Arcuino, G.; Takano, T.; Lin, J.; Peng, W.G.; Wan, P.; Li, P.; Xu, Q.; Liu, Q.S.; Goldman, S.A.; et al. P2X7 receptor inhibition improves recovery after spinal cord injury. Nat. Med. 2004, 10, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Cekic, C.; Linden, J. Purinergic regulation of the immune system. Nat. Rev. Immunol. 2016, 16, 177–192. [Google Scholar] [CrossRef]
- Danquah, W.; Meyer-Schwesinger, C.; Rissiek, B.; Pinto, C.; Serracant-Prat, A.; Amadi, M.; Iacenda, D.; Knop, J.H.; Hammel, A.; Bergmann, P.; et al. Nanobodies that block gating of the P2X7 ion channel ameliorate inflammation. Sci. Transl. Med. 2016, 8, 366ra162. [Google Scholar] [CrossRef] [PubMed]
- Choi, G.B.; Yim, Y.S.; Wong, H.; Kim, S.; Kim, H.; Kim, S.V.; Hoeffer, C.A.; Littman, D.R.; Huh, J.R. The maternal interleukin-17a pathway in mice promotes autism-like phenotypes in offspring. Science 2016, 351, 933–939. [Google Scholar] [CrossRef]
- Horvath, G.; Otrokocsi, L.; Beko, K.; Baranyi, M.; Kittel, A.; Fritz-Ruenes, P.A.; Sperlagh, B. P2X7 Receptors Drive Poly(I:C) Induced Autism-like Behavior in Mice. J. Neurosci. 2019, 39, 2542–2561. [Google Scholar] [CrossRef] [PubMed]
- Nicke, A.; Baumert, H.G.; Rettinger, J.; Eichele, A.; Lambrecht, G.; Mutschler, E.; Schmalzing, G. P2X1 and P2X3 receptors form stable trimers: A novel structural motif of ligand-gated ion channels. EMBO J. 1998, 17, 3016–3028. [Google Scholar] [CrossRef] [PubMed]
- North, R.A. Molecular physiology of P2X receptors. Physiol. Rev. 2002, 82, 1013–1067. [Google Scholar] [CrossRef]
- Kawate, T.; Michel, J.C.; Birdsong, W.T.; Gouaux, E. Crystal structure of the ATP-gated P2X(4) ion channel in the closed state. Nature 2009, 460, 592–598. [Google Scholar] [CrossRef]
- Browne, L.E.; Compan, V.; Bragg, L.; North, R.A. P2X7 receptor channels allow direct permeation of nanometer-sized dyes. J. Neurosci. 2013, 33, 3557–3566. [Google Scholar] [CrossRef]
- Harkat, M.; Peverini, L.; Cerdan, A.H.; Dunning, K.; Beudez, J.; Martz, A.; Calimet, N.; Specht, A.; Cecchini, M.; Chataigneau, T.; et al. On the permeation of large organic cations through the pore of ATP-gated P2X receptors. Proc. Natl. Acad. Sci. USA 2017, 114, E3786–E3795. [Google Scholar] [CrossRef]
- McCarthy, A.E.; Yoshioka, C.; Mansoor, S.E. Full-Length P2X7 Structures Reveal How Palmitoylation Prevents Channel Desensitization. Cell 2019, 179, 659–670.e613. [Google Scholar] [CrossRef] [PubMed]
- Pippel, A.; Stolz, M.; Woltersdorf, R.; Kless, A.; Schmalzing, G.; Markwardt, F. Localization of the gate and selectivity filter of the full-length P2X7 receptor. Proc. Natl. Acad. Sci. USA 2017, 114, E2156–E2165. [Google Scholar] [CrossRef] [PubMed]
- Karasawa, A.; Kawate, T. Structural basis for subtype-specific inhibition of the P2X7 receptor. eLife 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Kawate, T.; Robertson, J.L.; Li, M.; Silberberg, S.D.; Swartz, K.J. Ion access pathway to the transmembrane pore in P2X receptor channels. J. Gen. Physiol. 2011, 137, 579–590. [Google Scholar] [CrossRef]
- Egan, T.M.; Khakh, B.S. Contribution of calcium ions to P2X channel responses. J. Neurosci. 2004, 24, 3413–3420. [Google Scholar] [CrossRef]
- Samways, D.S.; Khakh, B.S.; Egan, T.M. Allosteric Modulation of Ca2+ flux in Ligand-gated Cation Channel (P2X4) by Actions on Lateral Portals. J. Biol. Chem. 2012, 287, 7594–7602. [Google Scholar] [CrossRef]
- Samways, D.S.; Khakh, B.S.; Dutertre, S.; Egan, T.M. Preferential use of unobstructed lateral portals as the access route to the pore of human ATP-gated ion channels (P2X receptors). Proc. Natl. Acad. Sci. USA 2011, 108, 13800–13805. [Google Scholar] [CrossRef]
- Samways, D.S.; Egan, T.M. Acidic amino acids impart enhanced Ca2+ permeability and flux in two members of the ATP-gated P2X receptor family. J. Gen. Physiol. 2007, 129, 245–256. [Google Scholar] [CrossRef]
- Yan, Z.; Liang, Z.; Obsil, T.; Stojilkovic, S.S. Participation of the Lys313-Ile333 sequence of the purinergic P2X4 receptor in agonist binding and transduction of signals to the channel gate. J. Biol. Chem. 2006, 281, 32649–32659. [Google Scholar] [CrossRef]
- Rokic, M.B.; Stojilkovic, S.S.; Vavra, V.; Kuzyk, P.; Tvrdonova, V.; Zemkova, H. Multiple roles of the extracellular vestibule amino acid residues in the function of the rat P2X4 receptor. PLoS ONE. 2013, 8, e59411. [Google Scholar] [CrossRef]
- Virginio, C.; MacKenzie, A.; North, R.A.; Surprenant, A. Kinetics of cell lysis, dye uptake and permeability changes in cells expressing the rat P2X7 receptor. J. Physiol. 1999, 519 Pt 2, 335–346. [Google Scholar] [CrossRef]
- Roger, S.; Pelegrin, P.; Surprenant, A. Facilitation of P2X7 receptor currents and membrane blebbing via constitutive and dynamic calmodulin binding. J. Neurosci. 2008, 28, 6393–6401. [Google Scholar] [CrossRef] [PubMed]
- Roger, S.; Gillet, L.; Baroja-Mazo, A.; Surprenant, A.; Pelegrin, P. C-terminal calmodulin-binding motif differentially controls human and rat P2X7 receptor current facilitation. J. Biol. Chem. 2010, 285, 17514–17524. [Google Scholar] [CrossRef] [PubMed]
- Khadra, A.; Tomic, M.; Yan, Z.; Zemkova, H.; Sherman, A.; Stojilkovic, S.S. Dual Gating Mechanism and Function of P2X7 Receptor Channels. Biophys. J. 2013, 104, 2612–2621. [Google Scholar] [CrossRef] [PubMed]
- Allsopp, R.C.; Evans, R.J. Contribution of the Juxtatransmembrane Intracellular Regions to the Time Course and Permeation of ATP-gated P2X7 Receptor Ion Channels. J. Biol. Chem. 2015, 290, 14556–14566. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Li, S.; Liang, Z.; Tomic, M.; Stojilkovic, S.S. The P2X7 receptor channel pore dilates under physiological ion conditions. J. Gen. Physiol. 2008, 132, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.H.; Rassendren, F.; Spelta, V.; Surprenant, A.; North, R.A. Amino acid residues involved in gating identified in the first membrane-spanning domain of the rat P2X(2) receptor. J. Biol. Chem. 2001, 276, 14902–14908. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.A.; Evans, R.J. Contribution of conserved polar glutamine, asparagine and threonine residues and glycosylation to agonist action at human P2X1 receptors for ATP. J. Neurochem. 2006, 96, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Allsopp, R.C.; El Ajouz, S.; Schmid, R.; Evans, R.J. Cysteine scanning mutagenesis (residues Glu52-Gly96) of the human P2X1 receptor for ATP: Mapping agonist binding and channel gating. J. Biol. Chem. 2011, 286, 29207–29217. [Google Scholar] [CrossRef]
- Du, J.; Dong, H.; Zhou, H.X. Gating mechanism of a P2X4 receptor developed from normal mode analysis and molecular dynamics simulations. Proc. Natl. Acad. Sci. USA 2012, 109, 4140–4145. [Google Scholar] [CrossRef]
- Digby, H.R.; Roberts, J.A.; Sutcliffe, M.J.; Evans, R.J. Contribution of conserved glycine residues to ATP action at human P2X1 receptors: Mutagenesis indicates that the glycine at position 250 is important for channel function. J. Neurochem. 2005, 95, 1746–1754. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.A.; Evans, R.J. Cysteine substitution mutants give structural insight and identify ATP binding and activation sites at P2X receptors. J. Neurosci. 2007, 27, 4072–4082. [Google Scholar] [CrossRef] [PubMed]
- Rassendren, F.; Buell, G.; Newbolt, A.; North, R.A.; Surprenant, A. Identification of amino acid residues contributing to the pore of a P2X receptor. EMBO J. 1997, 16, 3446–3454. [Google Scholar] [CrossRef] [PubMed]
- Boue-Grabot, E.; Emerit, M.B.; Toulme, E.; Seguela, P.; Garret, M. Cross-talk and co-trafficking between rho1/GABA receptors and ATP-gated channels. J. Biol. Chem. 2004, 279, 6967–6975. [Google Scholar] [CrossRef]
- Boue-Grabot, E.; Barajas-Lopez, C.; Chakfe, Y.; Blais, D.; Belanger, D.; Emerit, M.B.; Seguela, P. Intracellular cross talk and physical interaction between two classes of neurotransmitter-gated channels. J. Neurosci. 2003, 23, 1246–1253. [Google Scholar] [CrossRef]
- Lukacs, G.L.; Segal, G.; Kartner, N.; Grinstein, S.; Zhang, F. Constitutive internalization of cystic fibrosis transmembrane conductance regulator occurs via clathrin-dependent endocytosis and is regulated by protein phosphorylation. Biochem. J. 1997, 328 Pt 2, 353–361. [Google Scholar] [CrossRef]
- Caohuy, H.; Jozwik, C.; Pollard, H.B. Rescue of DeltaF508-CFTR by the SGK1/Nedd4-2 signaling pathway. J. Biol. Chem. 2009, 284, 25241–25253. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| rP2X7 | I1 (nA) | I1 + I2 (nA) | (I1) EC50 (µM) | (I1 + I2) EC50 (µM) | τoff (s) | EtBr Uptake (%) |
|---|---|---|---|---|---|---|
| WT | 1.7 ± 0.2 | 2.6 ± 0.3 | 45.2 ± 4.6 | 20.9 ± 2.5 | 0.45 ± 0.06 | 100 ± 5 |
| K49A | 0.7 ± 0.2 ** | 1.0 ± 0.2 ** | 92 ± 28 * | 63 ± 26 * | 0.25 ± 0.41 | 25 ± 4 ** |
| K49R | 1.9 ± 0.2 | 3.1 ± 0.4 | 70.9 ± 10.3 | 36.2 ± 12 | 0.62 ± 0.10 | 129 ± 22 |
| Y51A/L | n.f. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Y51F | 0.4 ± 0.1 ** | 0.6 ± 0.1 ** | 80.6 ± 19 * | 51.2 ± 11 * | 0.72 ± 0.27 | 9.2 ± 2 ** |
| Y51W | 1.0 ± 0.1 * | 1.4 ± 0.2 * | 30.8 ± 2.8 | 20.3 ± 3.5 | 0.84 ± 0.22 | 38 ± 7.5 ** |
| Q52A | n.f. | n.d. | n.d. | n.d. | n.d. | n.d. |
| F322A | 2.7 ± 0.3 * | 2.2 ± 0.2 | 8.6 ± 3.4 ** | 2.4 ± 0.3 ** | 27.1 ± 2.7 ** | 235 ± 41 ** |
| F322W | 2.1 ± 0.2 | 3.0 ± 0.6 | 35.5 ± 5.3 | 16.5 ± 2.1 | 1.39 ± 0.13 ** | 219 ± 31 ** |
| F322Y | 1.5 ± 0.2 | 2.3 ± 0.4 | 57.0 ± 27.2 | 26 ± 10.9 | 1.47 ± 0.25 ** | 205 ± 28 ** |
| F322L | 2.4 ± 0.3 * | 2.6 ± 0.4 | 12.7 ± 8.1 ** | 8.8 ± 3.2 ** | 2.19 ± 0.34 ** | 160 ± 6 * |
| G323A/S/P | n.f. | n.d. | n.d. | n.d. | n.d. | n.d. |
| G326A | 0.5 ± 0.1 ** | 0.8 ± 0.2 ** | 25.8 ± 4.5 * | 11.6 ±2.0 * | 1.28 ± 0.15 ** | 9 ± 3 ** |
| K327A | 0.8 ± 0.3 * | 1.1 ± 0.2 * | 53.4 ± 11.8 | 26.7 ± 7.1 | 0.58 ± 0.09 | 12 ± 4 ** |
| F328A | 0.3 ± 0.1 ** | 0.5 ± 0.3 ** | 46.0 ± 25 | 23.5 ±5.2 | 0.23 ± 0.06 | 11 ± 2 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rupert, M.; Bhattacharya, A.; Stillerova, V.T.; Jindrichova, M.; Mokdad, A.; Boué-Grabot, E.; Zemkova, H. Role of Conserved Residues and F322 in the Extracellular Vestibule of the Rat P2X7 Receptor in Its Expression, Function and Dye Uptake Ability. Int. J. Mol. Sci. 2020, 21, 8446. https://doi.org/10.3390/ijms21228446
Rupert M, Bhattacharya A, Stillerova VT, Jindrichova M, Mokdad A, Boué-Grabot E, Zemkova H. Role of Conserved Residues and F322 in the Extracellular Vestibule of the Rat P2X7 Receptor in Its Expression, Function and Dye Uptake Ability. International Journal of Molecular Sciences. 2020; 21(22):8446. https://doi.org/10.3390/ijms21228446
Chicago/Turabian StyleRupert, Marian, Anirban Bhattacharya, Vendula Tvrdonova Stillerova, Marie Jindrichova, Audrey Mokdad, Eric Boué-Grabot, and Hana Zemkova. 2020. "Role of Conserved Residues and F322 in the Extracellular Vestibule of the Rat P2X7 Receptor in Its Expression, Function and Dye Uptake Ability" International Journal of Molecular Sciences 21, no. 22: 8446. https://doi.org/10.3390/ijms21228446
APA StyleRupert, M., Bhattacharya, A., Stillerova, V. T., Jindrichova, M., Mokdad, A., Boué-Grabot, E., & Zemkova, H. (2020). Role of Conserved Residues and F322 in the Extracellular Vestibule of the Rat P2X7 Receptor in Its Expression, Function and Dye Uptake Ability. International Journal of Molecular Sciences, 21(22), 8446. https://doi.org/10.3390/ijms21228446