Adipokines, Myokines, and Cardiokines: The Role of Nutritional Interventions




Abstract
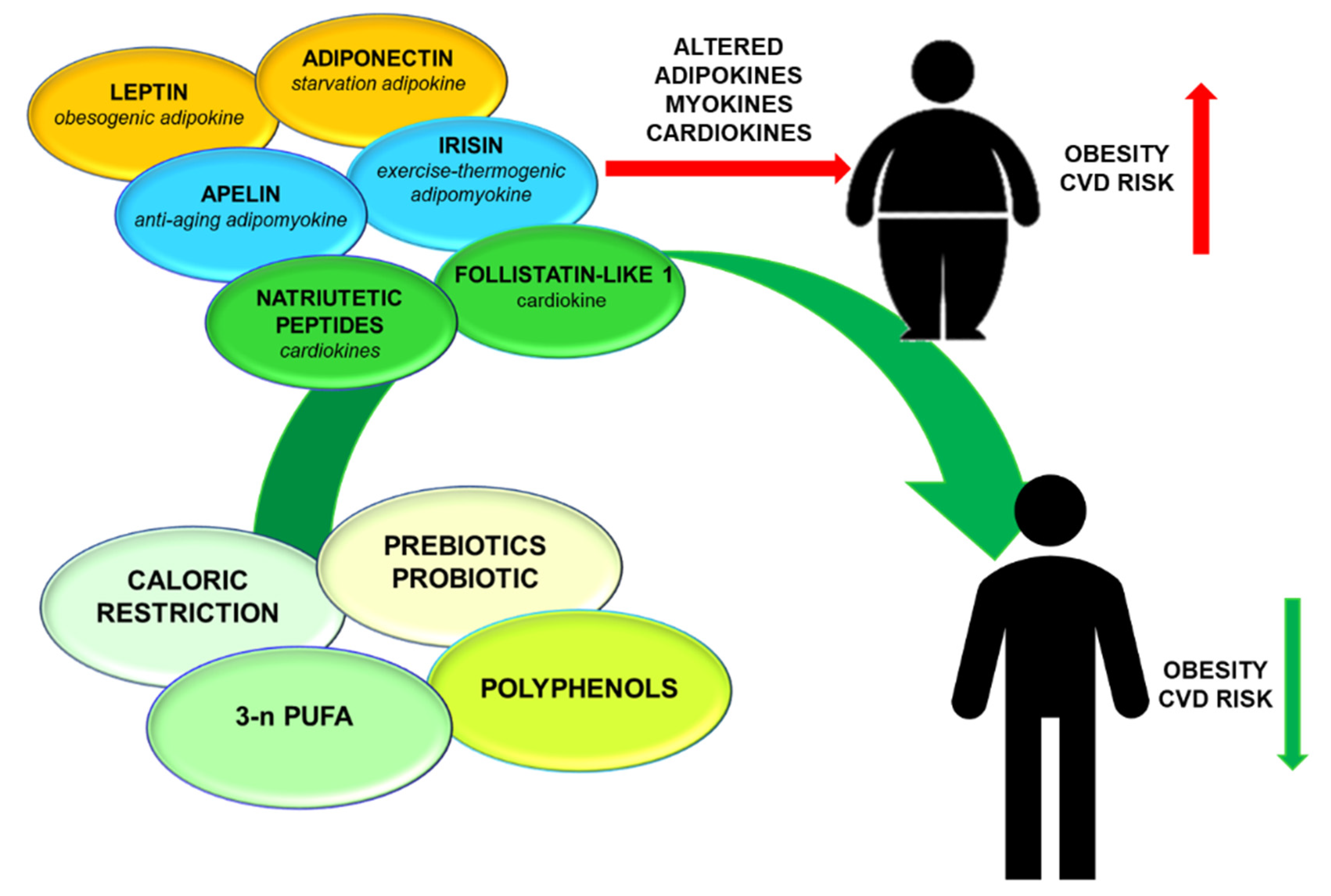
1. Introduction
2. Leptin
2.1. Leptin: Biological Role
2.2. Leptin: Nutritional Interventions
3. Adiponectin
3.1. Adiponectin: Biological Role
3.2. Adiponectin: Nutritional Interventions
4. Apelin/APJ Axis
4.1. Apelin/APJ Axis: Biological Role
4.2. Apelin/APJ Axis: Nutritional Interventions
5. Irisin
5.1. Irisin: Biological Role
5.2. Irisin: Nutritional Interventions
6. Natriuretic Peptides
6.1. Natriuretic Peptides: Biological Role
6.2. Natriuretic Peptides: Nutritional Interventions
7. Follistatin-Like 1
7.1. Follistatin-Like 1: Biological Role
7.2. Follistatin-Like 1: Nutritional Interventions
8. Discussion
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALA | Linolenic acid |
| ANP | Atrial natriuretic peptide |
| APLN | Apelin |
| APN | Adiponectin |
| BNP | B-type natriuretic peptide |
| CR | Caloric restriction diet |
| CVD | Cardiovascular disease |
| DHA | Docosahexaenoic acid |
| EPA | n-3 fatty acids eicosapentaenoic acid |
| FSTL-1 | Follistatin-like 1 |
| n-3 PUFA | Omega-3 polyunsaturated fatty acids |
| NAFLD | Non-alcoholic fatty liver disease |
| ROS | Reactive oxygen species |
References
- Shim, K.; Begum, R.; Yang, C.; Wang, H. Complement activation in obesity, insulin resistance, and type 2 diabetes mellitus. World J. Diabetes 2020, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Avgerinos, K.I.; Spyrou, N.; Mantzoros, C.S.; Dalamaga, M. Obesity and cancer risk: Emerging biological mechanisms and perspectives. Metabolism 2019, 92, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Hu, J.; Zhu, C. Obesity aggravates COVID-19: A systematic review and meta-analysis. J. Med. Virol. 2020, 30. [Google Scholar] [CrossRef]
- Kachur, S.; Lavie, C.J.; de Schutter, A.; Milani, R.V.; Ventura, H.O. Obesity and cardiovascular diseases. Minerva Med. 2017, 108, 212–228. [Google Scholar] [CrossRef]
- Vona, R.; Gambardella, L.; Cittadini, C.; Straface, E.; Pietraforte, D. Biomarkers of Oxidative Stress in Metabolic Syndrome and Associated Diseases. Oxid. Med. Cell Longev. 2019. [Google Scholar] [CrossRef]
- Saltiel, A.R.; Olefsky, J.M. Inflammatory mechanisms linking obesity and metabolic disease. J. Clin. Investig. 2017, 127. [Google Scholar] [CrossRef]
- Su, X.; Peng, D. Emerging functions of adipokines in linking the development of obesity and cardiovascular diseases. Mol. Biol. Rep. 2020. [Google Scholar] [CrossRef]
- Wu, Y.S.; Zhu, B.; Luo, A.L.; Yang, L.; Yang, C. The Role of Cardiokines in Heart Diseases: Beneficial or Detrimental? Biomed. Res. Int. 2018. [Google Scholar] [CrossRef]
- Ferrero, K.M.; Koch, W.J. Metabolic Crosstalk between the Heart and Fat. Korean Circ. J. 2020, 50, 379–394. [Google Scholar] [CrossRef]
- Chen, W.; Wang, L.; You, W.; Shan, T. Myokines mediate the cross talk between skeletal muscle and other organs. J. Cell Physiol. 2020. [Google Scholar] [CrossRef]
- Bergamin, A.; Mantzioris, E.; Cross, G.; Deo, P.; Garg, S.; Hill, A.M. Nutraceuticals: Reviewing their Role in Chronic Disease Prevention and Management. Pharm. Med. 2019, 33, 291–309. [Google Scholar] [CrossRef]
- Santos, J.; Leitão-Correia, F.; Sousa, M.J.; Leão, C. Dietary Restriction and Nutrient Balance in Aging. Oxid. Med. Cell Longev. 2016. [Google Scholar] [CrossRef]
- Patel, S.A.; Chaudhari, A.; Gupta, R.; Velingkaar, N.; Kondratov, R.V. Circadian clocks govern calorie restriction-mediated life span extension through BMAL1- and IGF-1-dependent mechanisms. FASEB J. 2016, 30, 1634–1642. [Google Scholar] [CrossRef]
- Pifferi, F.; Aujard, F. Caloric restriction, longevity and aging: Recent contributions from human and non-human primate studies. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 95. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Casado, E.; Khraiwesh, H.; López-Domínguez, J.A.; Montero-Guisado, J.; López-Lluch, G.; Navas, P.; de Cabo, R.; Ramsey, J.J.; González-Reyes, J.A.; Villalba, J.M. The Impact of Aging, Calorie Restriction and Dietary Fat on Autophagy Markers and Mitochondrial Ultrastructure and Dynamics in Mouse Skeletal Muscle. J. Gerontol A Biol. Sci. Med. Sci. 2019, 74, 760–769. [Google Scholar] [CrossRef]
- Colman, R.J.; Beasley, T.M.; Kemnitz, J.W.; Johnson, S.C.; Weindruch, R.; Anderson, R.M. Caloric restriction reduces age-related and all-cause mortality in rhesus monkeys. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef]
- Das, S.K.; Roberts, S.B.; Bhapkar, M.V.; Villareal, D.T.; Fontana, L.; Martin, C.K.; Racette, S.B.; Fuss, P.J.; Kraus, W.E.; Wong, W.W.; et al. CALERIE-2 Study Group. Body-composition changes in the Comprehensive Assessment of Long-term Effects of Reducing Intake of Energy (CALERIE)-2 study: A 2-y randomized controlled trial of calorie restriction in nonobese humans. Am. J. Clin. Nutr. 2017, 105, 913–927. [Google Scholar] [CrossRef]
- Kraus, W.E.; Bhapkar, M.; Huffman, K.M.; Pieper, C.F.; Krupa Das, S.; Redman, L.M.; Villareal, D.T.; Rochon, J.; Roberts, S.B.; Ravussin, E.; et al. CALERIE Investigators. 2 years of calorie restriction and cardiometabolic risk (CALERIE): Exploratory outcomes of a multicentre, phase 2, randomised controlled trial. Lancet Diabetes Endocrinol. 2019, 7, 673–683. [Google Scholar] [CrossRef]
- Novakovic, M.; Rout, A.; Kingsley, T.; Kirchoff, R.; Singh, A.; Verma, V.; Kant, R.; Chaudhary, R. Role of gut microbiota in cardiovascular diseases. World J. Cardiol. 2020, 12, 110–122. [Google Scholar] [CrossRef]
- Ejtahed, H.S.; Angoorani, P.; Soroush, A.R.; Hasani-Ranjbar, S.; Siadat, S.D.; Larijani, B. Gut microbiota-derived metabolites in obesity: A systematic review. Biosci. Microbiota. Food Health 2020, 39, 65–76. [Google Scholar] [CrossRef]
- Louis-Jean, S.; Martirosyan, D. Nutritionally Attenuating the Human Gut Microbiome to Prevent and Manage Metabolic Syndrome. J. Agric. Food Chem. 2019, 67, 12675–12684. [Google Scholar] [CrossRef]
- Tortosa-Caparrós, E.; Navas-Carrillo, D.; Marín, F.; Orenes-Piñero, E. Anti-inflammatory effects of omega 3 and omega 6 polyunsaturated fatty acids in cardiovascular disease and metabolic syndrome. Crit. Rev. Food Sci. Nutr. 2017, 57, 3421–3429. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Rodrigues, C.F.; Sharopov, F.; Docea, A.O.; Can Karaca, A.; Sharifi-Rad, M.; Kahveci Karıncaoglu, D.; Gülseren, G.; Şenol, E.; Demircan, E.; et al. Diet, Lifestyle and Cardiovascular Diseases: Linking Pathophysiology to Cardioprotective Effects of Natural Bioactive Compounds. Int. J. Environ. Res. Public Health. 2020, 17, 2326. [Google Scholar] [CrossRef] [PubMed]
- Boccellino, M.; D’Angelo, S. Anti-Obesity Effects of Polyphenol Intake: Current Status and Future Possibilities. Int. J. Mol. Sci. 2020, 21, 5642. [Google Scholar] [CrossRef]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372. [Google Scholar] [CrossRef]
- Seoane-Collazo, P.; Martínez-Sánchez, N.; Milbank, E.; Contreras, C. Incendiary Leptin. Nutrients 2020, 12, 472. [Google Scholar] [CrossRef]
- De Git, K.C.G.; Peterse, C.; Beerens, S.; Luijendijk, M.C.M.; van der Plasse, G.; la Fleur, S.E.; Adan, R.A.H. Is leptin resistance the cause or the consequence of diet-induced obesity? Int. J. Obes. 2018, 42, 1445–1457. [Google Scholar] [CrossRef]
- Gonzalez-Carter, D.; Goode, A.E.; Fiammengo, R.; Dunlop, I.E.; Dexter, D.T.; Porter, A.E. Inhibition of Leptin-ObR Interaction Does not Prevent Leptin Translocation Across a Human Blood-Brain Barrier Model. J. Neuroendocrol. 2016, 28. [Google Scholar] [CrossRef]
- Izquierdo, A.G.; Crujeiras, A.B.; Casanueva, F.F.; Carreira, M.C. Leptin, Obesity, and Leptin Resistance: Where Are We 25 Years Later? Nutrients 2019, 11, 2704. [Google Scholar] [CrossRef]
- Friedman, J.M. Leptin and the endocrine control of energy balance. Nat. Metab. 2019, 1, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Charchour, R.; Dufour-Rainfray, D.; Morineau, G.; Vatier, C.; Fellahi, S.; Vigouroux, C.; Genoux, A.; Capeau, J.; Lacorte, J.-M.; Collet, C.; et al. Mutltifaceted biological roles of leptin. Ann. Biol. Clin. 2020, 78, 231–242. [Google Scholar] [CrossRef]
- Pérez-Pérez, A.; Sánchez-Jiménez, F.; Vilariño-García, T.; Sánchez-Margalet, V. Role of Leptin in Inflammation and Vice Versa. Int. J. Mol. Sci. 2020, 21, 5887. [Google Scholar] [CrossRef]
- Poetsch, M.S.; Strano, A.; Guan, K. Role of Leptin in Cardiovascular Diseases. Front. Endocrinol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Becerril, S.; Rodríguez, A.; Catalán, V.; Ramírez, B.; Unamuno, X.; Portincasa, P.; Gómez-Ambrosi, J.; Frühbeck, G. Functional Relationship between Leptin and Nitric Oxide in Metabolism. Nutrients 2019, 11, 2129. [Google Scholar] [CrossRef]
- Simonds, S.E.; Pryor, J.T.; Ravussin, E.; Greenway, F.L.; Dileone, R.; Allen, A.M.; Bassi, J.; Elmquist, J.K.; Keogh, J.M.; Henning, E.; et al. Leptin mediates the increase in blood pressure associated with obesity. Cell 2014, 159. [Google Scholar] [CrossRef]
- Ghaedian, M.M.; Nazari Jaz, A.; Momeni, M.; Ghaedian, T.; Samiei, N. Plasma leptin level is positively associated with blood pressure measures independent of gender and BMI. Clin. Exp. Hypertens. 2020, 42, 31–35. [Google Scholar] [CrossRef]
- Alpert, M.A.; Karthikeyan, K.; Abdullah, O.; Ghadban, R. Obesity and Cardiac Remodeling in Adults: Mechanisms and Clinical Implications. Prog. Cardiovasc. Dis. 2018, 61, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.E.; Harmancey, R.; Stec, D.E. Lean heart: Role of leptin in cardiac hypertrophy and metabolism. World J. Cardiol. 2015, 7. [Google Scholar] [CrossRef]
- Martínez-Martínez, E.; Jurado-López, R.; Valero-Muñoz, M.; Bartolomé, M.V.; Ballesteros, S.; Luaces, M.; Briones, A.M.; López-Andrés, N.; Miana, M.; Cachofeiro, V. Leptin induces cardiac fibrosis through galectin-3, mTOR and oxidative stress: Potential role in obesity. J. Hypertens. 2014, 32, 1104–1114. [Google Scholar] [CrossRef]
- An, H.S.; Lee, J.Y.; Choi, E.B.; Jeong, E.A.; Shin, H.J.; Kim, K.E.; Park, K.A.; Jin, Z.; Lee, J.E.; Koh, J.S.; et al. Caloric restriction reverses left ventricular hypertrophy through the regulation of cardiac iron homeostasis in impaired leptin signaling mice. Sci. Rep. 2020, 10, 7176. [Google Scholar] [CrossRef]
- Bagarolli, R.A.; Tobar, N.; Oliveira, A.G.; Araújo, T.G.; Carvalho, B.M.; Rocha, G.Z.; Vecina, J.F.; Calisto, K.; Guadagnini, D.; Prada, P.O.; et al. Probiotics modulate gut microbiota and improve insulin sensitivity in DIO mice. J. Nutr. Biochem. 2017, 50, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Al-Muzafar, H.M.; Amin, K.A. Probiotic mixture improves fatty liver disease by virtue of its action on lipid profiles, leptin, and inflammatory biomarkers. BMC Complement. Altern. Med. 2017, 17, 1–9. [Google Scholar] [CrossRef]
- Behrouz, V.; Jazayeri, S.; Aryaeian, N.; Zahedi, M.J.; Hosseini, F. Effects of Probiotic and Prebiotic Supplementation on Leptin, Adiponectin, and Glycemic Parameters in Non-alcoholic Fatty Liver Disease: A Randomized Clinical Trial. Middle East. J. Dig. Dis. 2017, 9, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Sabico, S.; Al-Mashharawi, A.; Al-Daghri, N.M.; Wani, K.; Amer, O.E.; Hussain, D.S.; Ahmed Ansari, M.G.; Masoud, M.S.; Alokail, M.S.; McTernan, P.G. Effects of a 6-month multi-strain probiotics supplementation in endotoxemic, inflammatory and cardiometabolic status of T2DM patients: A randomized, double-blind, placebo-controlled trial. Clin. Nutr. 2019, 38, 1561–1569. [Google Scholar] [CrossRef]
- Rouhani, M.H.; Hadi, A.; Ghaedi, E.; Salehi, M.; Mahdavi, A.; Mohammadi, H. Do probiotics, prebiotics and synbiotics affect adiponectin and leptin in adults? A systematic review and meta-analysis of clinical trials. Clin. Nutr. 2019, 38, 2031–2037. [Google Scholar] [CrossRef]
- Gray, B.; Steyn, F.; Davies, P.S.; Vitetta, L. Omega-3 fatty acids: A review of the effects on adiponectin and leptin and potential implications for obesity management. Eur. J. Clin. Nutr. 2013, 67, 1234–1242. [Google Scholar] [CrossRef]
- Sedláček, P.; Plavinová, I.; Langmajerová, J.; Dvořáková, J.; Novák, J.; Trefil, L.; Müller, L.; Buňatová, P.; Zeman, V.; Müllerová, D. Effect of n-3 fatty acids supplementation during life style modification in women with overweight. Cent. Eur. J. Public Health 2018, 26, 265–271. [Google Scholar] [CrossRef]
- Huang, F.; Del-Río-Navarro, B.E.; Leija-Martinez, J.; Torres-Alcantara, S.; Ruiz-Bedolla, E.; Hernández-Cadena, L.; Barraza-Villarreal, A.; Romero-Nava, R.; Sanchéz-Muñoz, F.; Villafaña, S.; et al. Effect of omega-3 fatty acids supplementation combined with lifestyle intervention on adipokines and biomarkers of endothelial dysfunction in obese adolescents with hypertriglyceridemia. J. Nutr. Biochem. 2019, 64, 162–169. [Google Scholar] [CrossRef]
- Russell, J.S.; Griffith, T.A.; Naghipour, S.; Vider, J.; Du Toit, E.F.; Patel, H.H.; Peart, J.N.; Headrick, J.P. Dietary α-Linolenic Acid Counters Cardioprotective Dysfunction in Diabetic Mice: Unconventional PUFA Protection. Nutrients 2020, 12, 2679. [Google Scholar] [CrossRef]
- Mostowik, M.; Gajos, G.; Zalewski, J.; Nessler, J.; Undas, A. Omega-3 polyunsaturated fatty acids increase plasma adiponectin to leptin ratio in stable coronary artery disease. Cardiovasc. Drugs Ther. 2013, 27, 289–295. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Q.; Ma, W.; Tian, F.; Shen, H.; Zhou, M. A combination of quercetin and resveratrol reduces obesity in high-fat diet-fed rats by modulation of gut microbiota. Food Funct. 2017, 8, 4644–4656. [Google Scholar] [CrossRef]
- Luvizotto Rde, A.; Nascimento, A.F.; Imaizumi, E.; Pierine, D.T.; Conde, S.J.; Correa, C.R.; Yeum, K.J.; Ferreira, A.L. Lycopene supplementation modulates plasma concentrations and epididymal adipose tissue mRNA of leptin, resistin and IL-6 in diet-induced obese rats. Br. J. Nutr. 2013, 110, 1803–1809. [Google Scholar] [CrossRef]
- Cheng, H.M.; Koutsidis, G.; Lodge, J.K.; Ashor, A.; Siervo, M.; Lara, J. Tomato and lycopene supplementation and cardiovascular risk factors: A systematic review and meta-analysis. Atherosclerosis 2017, 257, 100–108. [Google Scholar] [CrossRef]
- Mazidi, M.; Rezaie, P.; Ferns, G.A.; Gao, H.K. Impact of different types of tree nut, peanut, and soy nut consumption on serum C-reactive protein (CRP): A systematic review and meta-analysis of randomized controlled clinical trials. Medicine 2016, 95, e5165. [Google Scholar] [CrossRef] [PubMed]
- Gencoglu, H.; Orhan, C.; Tuzcu, M.; Sahin, N.; Juturu, V.; Sahin, K. Effects of walnut oil on metabolic profile and transcription factors in rats fed high-carbohydrate-/-fat diets. J. Food Biochem. 2020, 44. [Google Scholar] [CrossRef]
- Godwin, N.; Roberts, T.; Hooshmand, S.; Kern, M.; Hong, M.Y. Mixed Nuts May Promote Satiety While Maintaining Stable Blood Glucose and Insulin in Healthy, Obese, and Overweight Adults in a Two-Arm Randomized Controlled Trial. J. Med. Food 2019, 22, 427–432. [Google Scholar] [CrossRef]
- Heydari, M.; Cornide-Petronio, M.E.; Jiménez-Castro, M.B.; Peralta, C. Data on Adiponectin from 2010 to 2020: Therapeutic Target and Prognostic Factor for Liver Diseases? Int. J. Mol. Sci. 2020, 21, 5242. [Google Scholar] [CrossRef]
- Choi, H.M.; Doss, H.M.; Kim, K.S. Multifaceted Physiological Roles of Adiponectin in Inflammation and Diseases. Int. J. Mol. Sci. 2020, 21, 1219. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, T.; Yamauchi, T.; Kubota, N. The physiological and pathophysiological role of adiponectin and adiponectin receptors in the peripheral tissues and CNS. FEBS Lett. 2008, 582, 74–80. [Google Scholar] [CrossRef]
- Ryu, J.; Loza, C.A.; Xu, H.; Zhou, M.; Hadley, J.T.; Wu, J.; You, H.; Wang, H.; Yang, J.; Bai, J.; et al. Potential Roles of Adiponectin Isoforms in Human Obesity with Delayed Wound Healing. Cells 2019, 8, 1134. [Google Scholar] [CrossRef]
- López-Jaramillo, P.; Gómez-Arbeláez, D.; López-López, J.; López-López, C.; Martínez-Ortega, J.; Gómez-Rodríguez, A.; Triana-Cubillos, S. The role of leptin/adiponectin ratio in metabolic syndrome and diabetes. Horm. Mol. Biol. Clin. Investig. 2014, 18, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, Q.; Pu, H.; Wei, Q.; Duan, M.; Zhang, C.; Jiang, T.; Shou, X.; Zhang, J.; Yang, Y. Adiponectin improves NF-κB-mediated inflammation and abates atherosclerosis progression in apolipoprotein E-deficient mice. Lipids Health Dis. 2016, 15, 1–14. [Google Scholar] [CrossRef]
- Wang, X.; Pu, H.; Ma, C.; Jiang, T.; Wei, Q.; Zhang, C.; Duan, M.; Shou, X.; Su, L.; Zhang, J.; et al. Adiponectin abates atherosclerosis by reducing oxidative stress. Int. Med. J. Exp. Clin. Res. 2014, 20, 1792. [Google Scholar] [CrossRef]
- Ogunwobi, O.O.; Beales, I.L. Adiponectin stimulates proliferation and cytokine secretion in colonic epithelial cells. Regul. Pept. 2006, 134, 105–113. [Google Scholar] [CrossRef]
- Xu, X.J.; Babo, E.; Qin, F.; Croteau, D.; Colucci, W.S. Short-term caloric restriction in db/db mice improves myocardial function and increases high molecular weight (HMW) adiponectin. IJC Metab. Endocr. 2016, 13, 28–34. [Google Scholar] [CrossRef][Green Version]
- Ding, Q.; Ash, C.; Mracek, T.; Merry, B.; Bing, C. Caloric restriction increases adiponectin expression by adipose tissue and prevents the inhibitory effect of insulin on circulating adiponectin in rats. J. Nutr. Biochem. 2012, 23, 867–874. [Google Scholar] [CrossRef]
- Hołowko, J.; Michalczyk, M.M.; Zając, A.; Czerwińska-Rogowska, M.; Ryterska, K.; Banaszczak, M.; Jakubczyk, K.; Stachowska, E. Six Weeks of Calorie Restriction Improves Body Composition and Lipid Profile in Obese and Overweight Former Athletes. Nutrients 2019, 11, 1461. [Google Scholar] [CrossRef]
- Ott, B.; Skurk, T.; Hastreiter, L.; Lagkouvardos, I.; Fischer, S.; Büttner, J.; Kellerer, T.; Clavel, T.; Rychlik, M.; Haller, D.; et al. Effect of caloric restriction on gut permeability, inflammation markers, and fecal microbiota in obese women. Sci. Rep. 2017, 7, 11955. [Google Scholar] [CrossRef]
- Cho, J.; Koh, Y.; Han, J.; Kim, D.; Kim, T.; Kang, H. Adiponectin mediates the additive effects of combining daily exercise with caloric restriction for treatment of non-alcoholic fatty liver. Int. J. Obes. 2016, 40, 1760–1767. [Google Scholar] [CrossRef] [PubMed]
- Yazdanparast, F.; Jafarirad, S.; Borazjani, F.; Haghighizadeh, M.H.; Jahanshahi, A. Comparing between the effect of energy-restricted diet and yoga on the resting metabolic rate, anthropometric indices, and serum adipokine levels in overweight and obese staff women. J. Res. Med. Sci. 2020, 25. [Google Scholar] [CrossRef]
- Hume, M.P.; Nicolucci, A.C.; Reimer, R.A. Prebiotic supplementation improves appetite control in children with overweight and obesity: A randomized controlled trial. Am. J. Clin. Nutr. 2017, 105, 790–799. [Google Scholar] [CrossRef]
- Bernini, L.J.; Simão, A.N.C.; de Souza, C.H.B.; Alfieri, D.F.; Segura, L.G.; Costa, G.N.; Dichi, I. Effect of Bifidobacterium lactis HN019 on inflammatory markers and oxidative stress in subjects with and without the metabolic syndrome. Br. J. Nutr. 2018, 120, 645–652. [Google Scholar] [CrossRef]
- Feizollahzadeh, S.; Ghiasvand, R.; Rezaei, A.; Khanahmad, H.; Sadeghi, A.; Hariri, M. Effect of Probiotic Soy Milk on Serum Levels of Adiponectin, Inflammatory Mediators, Lipid Profile, and Fasting Blood Glucose Among Patients with Type II Diabetes Mellitus. Probiotics Antimicrob. Proteins 2017, 9, 41–47. [Google Scholar] [CrossRef]
- Shahrokhi, M.; Nagalli, S. Probiotics. In StatPearls [Internet]. Treasure Island (FL); StatPearls Publishing LLC.: Bethesda, MD, USA, 2020. [Google Scholar]
- Albracht-Schulte, K.; Kalupahana, N.S.; Ramalingam, L.; Wang, S.; Rahman, S.M.; Robert-McComb, J.; Moustaid-Moussa, N. Omega-3 fatty acids in obesity and metabolic syndrome: A mechanistic update. J. Nutr. Biochem. 2018, 58, 1–16. [Google Scholar] [CrossRef]
- Redondo Useros, N.; Gheorghe, A.; Perez de Heredia, F.; Díaz, L.E.; Baccan, G.C.; De la Fuente, M.; Marcos, A. 2-OHOA supplementation reduced adiposity and improved cardiometabolic risk to a greater extent than n-3 PUFA in obese mice. Obes. Res. Clin. Pract. 2019, 13, 579–585. [Google Scholar] [CrossRef]
- Mohammadi-Sartang, M.; Mazloom, Z.; Sohrabi, Z.; Sherafatmanesh, S.; Barati-Boldaji, R. Resveratrol supplementation and plasma adipokines concentrations? A systematic review and meta-analysis of randomized controlled trials. Pharmacol. Res. 2017, 117, 394–405. [Google Scholar] [CrossRef]
- Clark, C.C.T.; Ghaedi, E.; Arab, A.; Pourmasoumi, M.; Hadi, A. The effect of curcumin supplementation on circulating adiponectin: A systematic review and meta-analysis of randomized controlled trials. Diabetes Metab. Syndr. 2019, 13, 2819–2825. [Google Scholar] [CrossRef]
- Williams, C.J.; Fargnoli, J.L.; Hwang, J.J.; van Dam, R.M.; Blackburn, G.L.; Hu, F.B.; Mantzoros, C.S. Coffee consumption is associated with higher plasma adiponectin concentrations in women with or without type 2 diabetes: A prospective cohort study. Diabetes Care 2008, 31, 504–507. [Google Scholar] [CrossRef]
- Gressner, O.A.; Lahme, B.; Rehbein, K.; Siluschek, M.; Weiskirchen, R.; Gressner, A.M. Pharmacological application of caffeine inhibits TGF-beta-stimulated connective tissue growth factor expression in hepatocytes via PPARgamma and SMAD2/3-dependent pathways. J. Hepatol. 2008, 49, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Shokouh, P.; Jeppesen, P.B.; Christiansen, C.B.; Mellbye, F.B.; Hermansen, K.; Gregersen, S. Efficacy of Arabica Versus Robusta Coffee in Improving Weight, Insulin Resistance, and Liver Steatosis in a Rat Model of Type-2 Diabetes. Nutrients 2019, 11, 2074. [Google Scholar] [CrossRef]
- Van Dam, R.M.; Hu, F.B.; Willett, W.C. Coffee, Caffeine, and Health. N. Engl. J. Med. 2020, 383, 369–378. [Google Scholar] [CrossRef]
- Bagheri, R.; Rashidlamir, A.; Ashtary-Larky, D.; Wong, A.; Grubbs, B.; Motevalli, M.S.; Baker, J.S.; Laher, I.; Zouhal, H. Effects of green tea extract supplementation and endurance training on irisin, pro-inflammatory cytokines, and adiponectin concentrations in overweight middle-aged men. Eur. J. Appl. Physiol. 2020, 120, 915–923. [Google Scholar] [CrossRef]
- Antushevich, H.; Wójcik, M. Review: Apelin in disease. Clin. Chim. Acta. 2018, 483, 241–248. [Google Scholar] [CrossRef]
- O’Dowd, B.F.; Heiber, M.; Chan, A.; Heng, H.H.; Tsui, L.C.; Kennedy, J.L.; Shi, X.; Petronis, A.; George, S.R.; Nguyen, T. A human gene that shows identity with the gene encoding the angiotensin receptor is located on chromosome 11. Gene 1993, 136, 355–360. [Google Scholar] [CrossRef]
- Huang, Z.; He, L.; Chen, Z.; Chen, L. Targeting drugs to APJ receptor: From signaling to pathophysiological effects. J. Cell Physiol. 2018, 234, 61–74. [Google Scholar] [CrossRef]
- Tatemoto, K.; Hosoya, M.; Habata, Y.; Fujii, R.; Kakegawa, T.; Zou, M.X.; Kawamata, Y.; Fukusumi, S.; Hinuma, S.; Kitada, C.; et al. Isolation and characterization of a novel endogenous peptide ligand for the human APJ receptor. Biochem. Biophys. Res. Commun. 1998, 251, 471–476. [Google Scholar] [CrossRef]
- Wysocka, M.B.; Pietraszek-Gremplewicz, K.; Nowak, D. The Role of Apelin in Cardiovascular Diseases, Obesity and Cancer. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef]
- Goidescu, C.M.; Vida-Simiti, L.A. The Apelin-APJ System in the Evolution of Heart Failure. Clujul Med. 2015, 88, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Castan-Laurell, I.; Masri, B.; Valet, P. The apelin/APJ system as a therapeutic target in metabolic diseases. Expert Opin. Ther. Targets 2019, 23, 215–225. [Google Scholar] [CrossRef]
- Guo, R.; Rogers, O.; Nair, S. Targeting Apelinergic System in Cardiometabolic Disease. Curr. Drug Targets 2017, 18, 1785–1791. [Google Scholar] [CrossRef]
- Mughal, A.; O’Rourke, S.T. Vascular effects of apelin: Mechanisms and therapeutic potential. Pharmacol. Therapeutic 2018, 190, 139–147. [Google Scholar] [CrossRef]
- Zhang, Z.Z.; Wang, W.; Jin, H.Y.; Chen, X.; Cheng, Y.W.; Xu, Y.L.; Song, B.; Penninger, J.M.; Oudit, G.Y.; Zhong, J.C. Apelin Is a Negative Regulator of Angiotensin II-Mediated Adverse Myocardial Remodeling and Dysfunction. Hypertension 2017, 70, 1165–1175. [Google Scholar] [CrossRef]
- Xie, H.; Luo, G.; Zheng, Y.; Hu, D.; Peng, F.; Xie, L. Lowered circulating apelin is significantly associated with an increased risk for hypertension: A meta-analysis. Clin. Exp. Hypertens. 2017, 39, 435–440. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, W.; Feng, F.; Xu, J.; Wu, F. Apelin-13 protects against myocardial infarction-induced myocardial fibrosis. Mol. Med. Rep. 2016, 13, 5262–5268. [Google Scholar] [CrossRef]
- Ouyang, Q.; You, T.; Guo, J.; Xu, R.; Guo, Q.; Lin, J.; Zhao, H. Effects of Apelin on Left Ventricular-Arterial Coupling and Mechanical Efficiency in Rats with Ischemic Heart Failure. Dis Markers. 2019, 2019, 4823156. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Wu, D.; Li, L.; Chen, L. Apelin/APJ system: A bifunctional target for cardiac hypertrophy. Int. J. Cardiol. 2017, 230, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.H.; Guo, C.X.; Wang, H.X.; Lu, L.Q.; Wang, Y.J.; Zhang, L.K.; Du, F.H.; Zeng, X.J. Cardioprotective effects of adipokine apelin on myocardial infarction. Heart Vessels 2014, 29, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.C.; Zhang, Z.Z.; Wang, W.; McKinnie, S.M.K.; Vederas, J.C.; Oudit, G.Y. Targeting the apelin pathway as a novel therapeutic approach for cardiovascular diseases. Biochim. Biophys. Acta. Mol. Basis Dis. 2017, 1863, 1942–1950. [Google Scholar] [CrossRef]
- Yue, P.; Jin, H.; Aillaud, M.; Deng, A.C.; Azuma, J.; Asagami, T.; Kundu, R.K.; Reaven, G.M.; Quertermous, T.; Tsao, P.S. Apelin is necessary for the maintenance of insulin sensitivity. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E59–E67. [Google Scholar] [CrossRef]
- Zhou, Q.; Cao, J.; Chen, L. Apelin/APJ system: A novel therapeutic target for oxidative stress-related inflammatory diseases (Review). Int. J. Mol. Med. 2016, 37, 1159–1169. [Google Scholar] [CrossRef]
- Chen, Z.; Wu, D.; Li, L.; Chen, L. Apelin/APJ System: A Novel Therapeutic Target for Myocardial Ischemia/Reperfusion Injury. DNA Cell Biol. 2016, 35, 766–775. [Google Scholar] [CrossRef] [PubMed]
- Sabry, M.M.; Ramadan, N.M.; Al Dreny, B.A.; Rashed, L.A.; Abo El Enein, A. Protective effect of apelin preconditioning in a rat model of hepatic ischemia reperfusion injury; possible interaction between the apelin/APJ system, Ang II/AT1R system and eNOS. United Eur. Gastroenterol. J. 2019, 7, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Tao, Y.; Jiang, Y. Apelin activates the expression of inflammatory cytokines in microglial BV2 cells via PI-3K/Akt and MEK/Erk pathways. Sci. China Life Sci. 2015, 58, 531–540. [Google Scholar] [CrossRef]
- Heinonen, M.V.; Laaksonen, D.E.; Karhu, T.; Karhunen, L.; Laitinen, T.; Kainulainen, S.; Rissanen, A.; Niskanen, L.; Herzig, K.H. Effect of diet-induced weight loss on plasma apelin and cytokine levels in individuals with the metabolic syndrome. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 626–633. [Google Scholar] [CrossRef]
- Chu, J.; Zhang, H.; Huang, X.; Lin, Y.; Shen, T.; Chen, B.; Man, Y.; Wang, S.; Li, J. Apelin ameliorates TNF-α-induced reduction of glycogen synthesis in the hepatocytes through G protein-coupled receptor APJ. PLoS ONE 2013, 8, e57231. [Google Scholar] [CrossRef]
- Li, M.; Fang, H.; Hu, J. Apelin-13 ameliorates metabolic and cardiovascular disorders in a rat model of type 2 diabetes with a high-fat diet. Mol. Med. Rep. 2018, 18, 5784–5790. [Google Scholar] [CrossRef]
- Gunes, I.; Kartal, H.; Dursun, A.D.; Sungu, N.; Polat, Y.S.; Erkent, F.D.; Arslan, M.; Kucuk, A. Effects of apelin-13 on myocardial ischemia reperfusion injury in streptozotocine induced diabetic rats. Bratisl. Lek. Listy 2018, 119, 348–354. [Google Scholar] [CrossRef]
- Pan, C.S.; Teng, X.; Zhang, J.; Cai, Y.; Zhao, J.; Wu, W.; Wang, X.; Tang, C.S.; Qi, Y.F. Apelin antagonizes myocardial impairment in sepsis. J. Card Fail 2010, 16, 609–617. [Google Scholar] [CrossRef]
- Zhou, Q.; Chen, L.; Tang, M.; Guo, Y.; Li, L. Apelin/APJ system: A novel promising target for anti-aging intervention. Clin. Chim. Acta 2018, 487, 233–240. [Google Scholar] [CrossRef]
- Than, A.; Zhang, X.; Leow, M.K.; Poh, C.L.; Chong, S.K.; Chen, P. Apelin attenuates oxidative stress in human adipocytes. J. Biol. Chem. 2014, 289, 3763–3774. [Google Scholar] [CrossRef]
- Yang, R.; Fang, W.; Liang, J.; Lin, C.; Wu, S.; Yan, S.; Hu, C.; Ke, X. Apelin/APJ axis improves angiotensin II-induced endothelial cell senescence through AMPK/SIRT1 signaling pathway. Arch. Med. Sci. 2018, 14, 725–734. [Google Scholar] [CrossRef]
- Rai, R.; Ghosh, A.K.; Eren, M.; Mackie, A.R.; Levine, D.C.; Kim, S.Y.; Cedernaes, J.; Ramirez, V.; Procissi, D.; Smith, L.H.; et al. Downregulation of the Apelinergic Axis Accelerates Aging, whereas Its Systemic Restoration Improves the Mammalian Healthspan. Cell Rep. 2017, 21, 1471–1480. [Google Scholar] [CrossRef]
- Izadi, M.R.; Ghardashi Afousi, A.; Asvadi Fard, M.; Babaee Bigi, M.A. High-intensity interval training lowers blood pressure and improves apelin and NOx plasma levels in older treated hypertensive individuals. J. Physiol. Biochem. 2018, 74, 47–55. [Google Scholar] [CrossRef]
- Fujie, S.; Sato, K.; Miyamoto-Mikami, E.; Hasegawa, N.; Fujita, S.; Sanada, K.; Hamaoka, T.; Iemitsu, M. Reduction of arterial stiffness by exercise training is associated with increasing plasma apelin level in middle-aged and older adults. PLoS ONE 2014, 9, e93545. [Google Scholar] [CrossRef]
- Kwak, S.E.; Cho, S.C.; Bae, J.H.; Lee, J.; Shin, H.E.; Di Zhang, D.; Lee, Y.I.; Song, W. Effects of exercise-induced apelin on muscle function and cognitive function in aged mice. Exp. Gerontol. 2019, 127, 110710. [Google Scholar] [CrossRef]
- Attané, C.; Foussal, C.; Le Gonidec, S.; Benani, A.; Daviaud, D.; Wanecq, E.; Guzmán-Ruiz, R.; Dray, C.; Bezaire, V.; Rancoule, C.; et al. Apelin treatment increases complete Fatty Acid oxidation, mitochondrial oxidative capacity, and biogenesis in muscle of insulin-resistant mice. Diabetes 2012, 61, 310–320. [Google Scholar] [CrossRef]
- Clarke, K.J.; Whitaker, K.W.; Reyes, T.M. Diminished metabolic responses to centrally-administered apelin-13 in diet-induced obese rats fed a high-fat diet. J. Neuroendocrol. 2009, 21, 83–89. [Google Scholar] [CrossRef]
- Bertrand, C.; Pradère, J.P.; Geoffre, N.; Deleruyelle, S.; Masri, B.; Personnaz, J.; Le Gonidec, S.; Batut, A.; Louche, K.; Moro, C.; et al. Chronic apelin treatment improves hepatic lipid metabolism in obese and insulin-resistant mice by an indirect mechanism. Endocrine 2018, 60, 112–121. [Google Scholar] [CrossRef]
- El Wakeel, M.A.; El-Kassas, G.M.; Kamhawy, A.H.; Galal, E.M.; Nassar, M.S.; Hammad, E.M.; El-Zayat, S.R. Serum Apelin and Obesity-Related Complications in Egyptian Children. Open Access Maced. J. Med. Sci. 2018, 6, 1354–1358. [Google Scholar] [CrossRef]
- Zaki, M.; Kamal, S.; Ezzat, W.; Hassan, N.; Yousef, W.; Ryad, H.; Mohamed, R.; Youness, E.; Basha, W.; Elhosary, Y. Serum apelin levels and metabolic risk markers in obese women. J. Genet. Eng. Biotechnol. 2017, 15, 423–429. [Google Scholar] [CrossRef]
- Higuchi, K.; Masaki, T.; Gotoh, K.; Chiba, S.; Katsuragi, I.; Tanaka, K.; Kakuma, T.; Yoshimatsu, H. Apelin, an APJ receptor ligand, regulates body adiposity and favors the messenger ribonucleic acid expression of uncoupling proteins in mice. Endocrinology 2007, 148, 2690–2697. [Google Scholar] [CrossRef]
- Yuzbashian, E.; Zarkesh, M.; Asghari, G.; Hedayati, M.; Safarian, M.; Mirmiran, P.; Khalaj, A. Is apelin gene expression and concentration affected by dietary intakes? A systematic review. Crit. Rev. Food Sci. Nutr. 2018, 58, 680–688. [Google Scholar] [CrossRef]
- Yuzbashian, E.; Asghari, G.; Aghayan, M.; Hedayati, M.; Zarkesh, M.; Mirmiran, P.; Khalaj, A. Dietary glycemic index and dietary glycemic load is associated with apelin gene expression in visceral and subcutaneous adipose tissues of adults. Nutr. Metab. 2019, 16, 68. [Google Scholar] [CrossRef]
- Krist, J.; Wieder, K.; Klöting, N.; Oberbach, A.; Kralisch, S.; Wiesner, T.; Schön, M.R.; Gärtner, D.; Dietrich, A.; Shang, E.; et al. Effects of weight loss and exercise on apelin serum concentrations and adipose tissue expression in human obesity. Obes. Facts 2013, 6, 57–69. [Google Scholar] [CrossRef]
- Celik, A.; Saricicek, E.; Saricicek, V.; Sahin, E.; Ozdemir, G.; Bozkurt, S.; Okumus, M.; Sucakli, M.H.; Cikim, G.; Coskun, Y.; et al. Effect of Ramadan fasting on serum concentration of apelin-13 and new obesity indices in healthy adult men. Med. Sci. Monit. 2014, 20, 337–342. [Google Scholar] [CrossRef]
- Karimi, E.; Moini, A.; Yaseri, M.; Shirzad, N.; Sepidarkish, M.; Hossein-Boroujerdi, M.; Hosseinzadeh-Attar, M.J. Effects of synbiotic supplementation on metabolic parameters and apelin in women with polycystic ovary syndrome: A randomised double-blind placebo-controlled trial. Br. J. Nutr. 2018, 119, 398–406. [Google Scholar] [CrossRef]
- Bertrand, C.; Pignalosa, A.; Wanecq, E.; Rancoule, C.; Batut, A.; Deleruyelle, S.; Lionetti, L.; Valet, P.; Castan-Laurell, I. Effects of dietary eicosapentaenoic acid (EPA) supplementation in high-fat fed mice on lipid metabolism and apelin/APJ system in skeletal muscle. PLoS ONE 2013, 8, e78874. [Google Scholar] [CrossRef]
- Pérez-Echarri, N.; Pérez-Matute, P.; Marcos-Gómez, B.; Martínez, J.A.; Moreno-Aliaga, M.J. Effects of eicosapentaenoic acid ethyl ester on visfatin and apelin in lean and overweight (cafeteria diet-fed) rats. Br. J. Nutr. 2009, 101, 1059–1067. [Google Scholar] [CrossRef]
- Shamsi-Goushki, A.; Mortazavi, Z.; Mirshekar, M.A.; Mohammadi, M.; Moradi-Kor, N.; Jafari-Maskouni, S.; Shahraki, M. Comparative Effects of Curcumin versus Nano-Curcumin on Insulin Resistance, Serum Levels of Apelin and Lipid Profile in Type 2 Diabetic Rats. Diabetes Metab. Syndr. Obes. 2020, 13, 2337–2346. [Google Scholar] [CrossRef]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef]
- Benedini, S.; Dozio, E.; Invernizzi, P.L.; Vianello, E.; Banfi, G.; Terruzzi, I.; Luzi, L.; Corsi Romanelli, M.M. Irisin: A Potential Link between Physical Exercise and Metabolism-An Observational Study in Differently Trained Subjects, from Elite Athletes to Sedentary People. J. Diabetes Res. 2017, 2017, 1039161. [Google Scholar] [CrossRef]
- Korta, P.; Pocheć, E.; Mazur-Biały, A. Irisin as a Multifunctional Protein: Implications for Health and Certain Diseases. Medicina 2019, 55, 485. [Google Scholar] [CrossRef]
- Rodríguez, A.; Becerril, S.; Ezquerro, S.; Méndez-Giménez, L.; Frühbeck, G. Crosstalk between adipokines and myokines in fat browning. Acta. Physiol. 2017, 219, 362–381. [Google Scholar] [CrossRef]
- Perakakis, N.; Triantafyllou, G.A.; Fernández-Real, J.M.; Huh, J.Y.; Park, K.H.; Seufert, J.; Mantzoros, C.S. Physiology and role of irisin in glucose homeostasis. Nat. Rev. Endocrinol. 2017, 13, 324–337. [Google Scholar] [CrossRef] [PubMed]
- So, W.Y.; Leung, P.S. Irisin ameliorates hepatic glucose/lipid metabolism and enhances cell survival in insulin-resistant human HepG2 cells through adenosine monophosphate-activated protein kinase signaling. Int. J. Biochem. Cell Biol. 2016, 78, 237–247. [Google Scholar] [CrossRef]
- Xin, C.; Liu, J.; Zhang, J.; Zhu, D.; Wang, H.; Xiong, L.; Lee, Y.; Ye, J.; Lian, K.; Xu, C.; et al. Irisin improves fatty acid oxidation and glucose utilization in type 2 diabetes by regulating the AMPK signaling pathway. Int. J. Obes. 2016, 40, 443–451. [Google Scholar] [CrossRef]
- Liu, T.Y.; Shi, C.X.; Gao, R.; Sun, H.J.; Xiong, X.Q.; Ding, L.; Chen, Q.; Li, Y.H.; Wang, J.J.; Kang, Y.M.; et al. Irisin inhibits hepatic gluconeogenesis and increases glycogen synthesis via the PI3K/Akt pathway in type 2 diabetic mice and hepatocytes. Clin. Sci. 2015, 129, 839–850. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, J.O.; Kim, N.; Kim, J.K.; Kim, H.I.; Lee, Y.W.; Kim, S.J.; Choi, J.I.; Oh, Y.; Kim, J.H.; et al. Irisin, a Novel Myokine, Regulates Glucose Uptake in Skeletal Muscle Cells via AMPK. Mol. Endocrinol. 2015, 29, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Arhire, L.I.; Mihalache, L.; Covasa, M. Irisin: A Hope in Understanding and Managing Obesity and Metabolic Syndrome. Front. Endocrinol. 2019, 10, 524. [Google Scholar] [CrossRef]
- Monserrat-Mesquida, M.; Quetglas-Llabrés, M.; Abbate, M.; Montemayor, S.; Mascaró, C.M.; Casares, M.; Tejada, S.; Abete, I.; Zulet, M.A.; Tur, J.A.; et al. Oxidative Stress and Pro-Inflammatory Status in Patients with Non-Alcoholic Fatty Liver Disease. Antioxidants 2020, 9, 759. [Google Scholar] [CrossRef] [PubMed]
- Askari, H.; Rajani, S.F.; Poorebrahim, M.; Haghi-Aminjan, H.; Raeis-Abdollahi, E.; Abdollahi, M. A glance at the therapeutic potential of irisin against diseases involving inflammation, oxidative stress, and apoptosis: An introductory review. Pharm. Res. 2018, 129, 44–55. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, K.; Han, Y.; Zhu, H.; Zhou, X.; Tan, T.; Zeng, J.; Zhang, J.; Liu, Y.; Li, Y.; et al. Irisin Protects Heart Against Ischemia-Reperfusion Injury Through a SOD2-Dependent Mitochondria Mechanism. J. Cardiovasc. Pharm. 2018, 72, 259–269. [Google Scholar] [CrossRef]
- Deng, J.; Zhang, N.; Chen, F.; Yang, C.; Ning, H.; Xiao, C.; Sun, K.; Liu, Y.; Yang, M.; Hu, T.; et al. Irisin ameliorates high glucose-induced cardiomyocytes injury via AMPK/mTOR signal pathway. Cell Biol. Int. 2020. [Google Scholar] [CrossRef]
- Moscoso, I.; Cebro-Márquez, M.; Rodríguez-Mañero, M.; González-Juanatey, J.R.; Lage, R. FNDC5/Irisin counteracts lipotoxic-induced apoptosis in hypoxic H9c2 cells. J. Mol. Endocrinol. 2019, 63, 151–159. [Google Scholar] [CrossRef]
- Khorasani, Z.M.; Bagheri, R.K.; Yaghoubi, M.A.; Chobkar, S.; Aghaee, M.A.; Abbaszadegan, M.R.; Sahebkar, A. The association between serum irisin levels and cardiovascular disease in diabetic patients. Diabetes Metab. Syndr. 2019, 13, 786–790. [Google Scholar] [CrossRef]
- Abd El-Mottaleb, N.A.; Galal, H.M.; El Maghraby, K.M.; Gadallah, A.I. Serum irisin level in myocardial infarction patients with or without heart failure. Can. J. Physiol. Pharm. 2019, 97, 932–938. [Google Scholar] [CrossRef] [PubMed]
- Liao, Q.; Qu, S.; Tang, L.X.; Li, L.P.; He, D.F.; Zeng, C.Y.; Wang, W.E. Irisin exerts a therapeutic effect against myocardial infarction via promoting angiogenesis. Acta. Pharm. Sin. 2019, 40, 1314–1321. [Google Scholar] [CrossRef]
- Shirvani, H.; Delpasand, A.; Bazgir, B. The data on the aerobic training with or without calorie restriction and muscular levels of Irisin and muscular FNDC5 concentration in obese male Wistar rats. Data Brief. 2018, 21, 888–892. [Google Scholar] [CrossRef]
- Batitucci, G.; Brandao, C.F.C.; De Carvalho, F.G.; Marchini, J.S.; Pfrimer, K.; Ferrioli, E.; Cunha, F.Q.; Papoti, M.; Terrazas, S.I.B.M.; Junqueira-Franco, M.V.M.; et al. Taurine supplementation increases irisin levels after high intensity physical training in obese women. Cytokine 2019, 123, 154741. [Google Scholar] [CrossRef] [PubMed]
- Eskandari, M.; Hooshmand Moghadam, B.; Bagheri, R.; Ashtary-Larky, D.; Eskandari, E.; Nordvall, M.; Dutheil, F.; Wong, A. Effects of Interval Jump Rope Exercise Combined with Dark Chocolate Supplementation on Inflammatory Adipokine, Cytokine Concentrations, and Body Composition in Obese Adolescent Boys. Nutrients 2020, 12, 3011. [Google Scholar] [CrossRef]
- Kwon, J.; Kim, B.; Lee, C.; Joung, H.; Kim, B.K.; Choi, I.S.; Hyun, C.K. Comprehensive amelioration of high-fat diet-induced metabolic dysfunctions through activation of the PGC-1α pathway by probiotics treatment in mice. PLoS ONE 2020, 15, e0228932. [Google Scholar] [CrossRef]
- Ansari, S.; Djalali, M.; Mohammadzadeh Honarvar, N.; Mazaherioun, M.; Zarei, M.; Agh, F.; Gholampour, Z.; Javanbakht, M.H. The Effect of n-3 Polyunsaturated Fatty Acids Supplementation on Serum Irisin in Patients with Type 2 Diabetes: A Randomized, Double-Blind, Placebo-Controlled Trial. Int. J. Endocrinol. Metab. 2017, 15, e40614. [Google Scholar] [CrossRef]
- Palacios-González, B.; Vargas-Castillo, A.; Velázquez-Villegas, L.A.; Vasquez-Reyes, S.; López, P.; Noriega, L.G.; Aleman, G.; Tovar-Palacio, C.; Torre-Villalvazo, I.; Yang, L.J.; et al. Genistein increases the thermogenic program of subcutaneous WAT and increases energy expenditure in mice. J. Nutr. Biochem. 2019, 68, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Kheiripour, N.; Karimi, J.; Khodadadi, I.; Tavilani, H.; Taghi Goodarzi, M.; Hashemnia, M. Hepatoprotective Effects of Silymarin on Liver Injury via Irisin Upregulation and Oxidative Stress Reduction in Rats with Type 2 Diabetes. Iran J. Med. Sci. 2019, 44, 108–117. [Google Scholar]
- Daneshi-Maskooni, M.; Keshavarz, S.A.; Qorbani, M.; Mansouri, S.; Alavian, S.M.; Badri-Fariman, M.; Jazayeri-Tehrani, S.A.; Sotoudeh, G. Green cardamom supplementation improves serum irisin, glucose indices, and lipid profiles in overweight or obese non-alcoholic fatty liver disease patients: A double-blind randomized placebo-controlled clinical trial. BMC Complement. Altern. Med. 2019, 19, 59. [Google Scholar] [CrossRef]
- Rodriguez Lanzi, C.; Perdicaro, D.J.; Gambarte Tudela, J.; Muscia, V.; Fontana, A.R.; Oteiza, P.I.; Vazquez Prieto, M.A. Grape pomace extract supplementation activates FNDC5/irisin in muscle and promotes white adipose browning in rats fed a high-fat diet. Food Funct. 2020, 11, 1537–1546. [Google Scholar] [CrossRef]
- Jeon, Y.J.; Jung, S.J.; Kim, H.C. Does serum vitamin D level affect the association between cardiovascular health and cognition? Results of the Cardiovascular and Metabolic Diseases Etiology Research Center (CMERC) study. Eur. J. Neurol. 2020. [Google Scholar] [CrossRef]
- Latic, N.; Erben, R.G. Vitamin D and Cardiovascular Disease, with Emphasis on Hypertension, Atherosclerosis, and Heart Failure. Int. J. Mol. Sci. 2020, 21, 6483. [Google Scholar] [CrossRef] [PubMed]
- Nadimi, H.; Djazayery, A.; Javanbakht, M.H.; Dehpour, A.; Ghaedi, E.; Derakhshanian, H.; Mohammadi, H.; Zarei, M.; Djalali, M. The Effect of Vitamin D Supplementation on Serum and Muscle Irisin Levels, and FNDC5 Expression in Diabetic Rats. Rep. Biochem. Mol. Biol. 2019, 8, 236–243. [Google Scholar] [PubMed]
- Chang, E.; Kim, Y. Vitamin D Insufficiency Exacerbates Adipose Tissue Macrophage Infiltration and Decreases AMPK/SIRT1 Activity in Obese Rats. Nutrients 2017, 9, 338. [Google Scholar] [CrossRef]
- Sajjadi, S.F.; Mirzaei, K.; Khorrami-Nezhad, L.; Maghbooli, Z.; Keshavarz, S.A. Vitamin D Status and Resting Metabolic Rate May Modify through Expression of Vitamin D Receptor and Peroxisome Proliferator-Activated Receptor Gamma Coactivator-1 Alpha Gene in Overweight and Obese Adults. Ann. Nutr. Metab. 2018, 72, 43–49. [Google Scholar] [CrossRef]
- Safarpour, P.; Daneshi-Maskooni, M.; Vafa, M.; Nourbakhsh, M.; Janani, L.; Maddah, M.; Amiri, F.S.; Mohammadi, F.; Sadeghi, H. Vitamin D supplementation improves SIRT1, Irisin, and glucose indices in overweight or obese type 2 diabetic patients: A double-blind randomized placebo-controlled clinical trial. BMC Fam. Prac. 2020, 21, 26. [Google Scholar] [CrossRef]
- Vinnakota, S.; Chen, H.H. The Importance of Natriuretic Peptides in Cardiometabolic Diseases. J. Endoc. Soc. 2020, 4. [Google Scholar] [CrossRef]
- Goetze, J.P.; Bruneau, B.G.; Ramos, H.R.; Ogawa, T.; de Bold, M.K.; de Bold, A.J. Cardiac natriuretic peptides. Nat. Rev. Cardiol. 2020. [Google Scholar] [CrossRef]
- Matsuo, A.; Nagai-Okatani, C.; Nishigori, M.; Kangawa, K.; Minamino, N. Natriuretic peptides in human heart: Novel insight into their molecular forms, functions, and diagnostic use. Peptides 2019, 111, 3–17. [Google Scholar] [CrossRef]
- Berezin, A.E. Prognostication of clinical outcomes in diabetes mellitus: Emerging role of cardiac biomarkers. Diabetes Metab. Syndr. 2019, 13, 995–1003. [Google Scholar] [CrossRef]
- Cannone, V.; Cabassi, A.; Volpi, R.; Burnett, J.C., Jr. Atrial Natriuretic Peptide: A Molecular Target of Novel Therapeutic Approaches to Cardio-Metabolic Disease. Int. J. Mol. Sci. 2019, 20, 3265. [Google Scholar] [CrossRef]
- Bordicchia, M.; Liu, D.; Amri, E.Z.; Ailhaud, G.; Dessì-Fulgheri, P.; Zhang, C.; Takahashi, N.; Sarzani, R.; Collins, S. Cardiac natriuretic peptides act via p38 MAPK to induce the brown fat thermogenic program in mouse and human adipocytes. J. Clin. Invest. 2012, 122, 1022–1036. [Google Scholar] [CrossRef]
- Liu, D.; Ceddia, R.P.; Collins, S. Cardiac natriuretic peptides promote adipose ‘browning’ through mTOR complex-1. Mol. Metab. 2018, 9, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, K.; Nishida, M.; Yamamoto, R.; Koseki, M.; Moriyama, T.; Yamauchi-Takihara, K. Association between N-terminal pro-brain natriuretic peptide and adiponectin in healthy Japanese men. Clin. Chim. Acta. 2016, 460, 138–141. [Google Scholar] [CrossRef] [PubMed]
- Verboven, K.; Hansen, D.; Jocken, J.W.E.; Blaak, E.E. Natriuretic peptides in the control of lipid metabolism and insulin sensitivity. Obes. Rev. 2017, 18, 1243–1259. [Google Scholar] [CrossRef]
- Haufe, S.; Kaminski, J.; Utz, W.; Haas, V.; Mähler, A.; Daniels, M.A.; Birkenfeld, A.L.; Lichtinghagen, R.; Luft, F.C.; Schulz-Menger, J.; et al. Differential response of the natriuretic peptide system to weight loss and exercise in overweight or obese patients. J. Hypertens. 2015, 33, 1458–1464. [Google Scholar] [CrossRef]
- Kirkham, A.A.; Beka, V.; Prado, C.M. The effect of caloric restriction on blood pressure and cardiovascular function: A systematic review and meta-analysis of randomized controlled trials. Clin. Nutr. 2020, 1. [Google Scholar] [CrossRef] [PubMed]
- Nicoll, R.; Henein, M.Y. Caloric Restriction and Its Effect on Blood Pressure, Heart Rate Variability and Arterial Stiffness and Dilatation: A Review of the Evidence. Int. J. Mol. Sci. 2018, 19, 751. [Google Scholar] [CrossRef]
- Gan, X.T.; Ettinger, G.; Huang, C.X.; Burton, J.P.; Haist, J.V.; Rajapurohitam, V.; Sidaway, J.E.; Martin, G.; Gloor, G.B.; Swann, J.R.; et al. Probiotic administration attenuates myocardial hypertrophy and heart failure after myocardial infarction in the rat. Circ. Heart Fail. 2014, 7, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.P.; Hsieh, Y.M.; Kuo, W.W.; Lin, C.C.; Tsai, F.J.; Tsai, C.H.; Huang, C.Y.; Tsai, C.C. Inhibition of cardiac hypertrophy by probiotic-fermented purple sweet potato yogurt in spontaneously hypertensive rat hearts. Int. J. Mol. Med. 2012, 30, 1365–1375. [Google Scholar] [CrossRef][Green Version]
- Wang, C.; Xiong, B.; Huang, J. The Role of Omega-3 Polyunsaturated Fatty Acids in Heart Failure: A Meta-Analysis of Randomised Controlled Trials. Nutrients 2016, 9, 18. [Google Scholar] [CrossRef]
- Shoieb, S.M.; El-Kadi, A.O.S. Resveratrol attenuates angiotensin II-induced cellular hypertrophy through the inhibition of CYP1B1 and the cardiotoxic mid-chain HETE metabolites. Mol. Cell Biochem. 2020, 471, 165–176. [Google Scholar] [CrossRef]
- Liu, R.; Zhang, H.B.; Yang, J.; Wang, J.R.; Liu, J.X.; Li, C.L. Curcumin alleviates isoproterenol-induced cardiac hypertrophy and fibrosis through inhibition of autophagy and activation of mTOR. Eur. Rev. Med. Pharm. Sci. 2018, 22, 7500–7508. [Google Scholar] [CrossRef]
- Ren, B.C.; Zhang, Y.F.; Liu, S.S.; Cheng, X.J.; Yang, X.; Cui, X.G.; Zhao, X.R.; Zhao, H.; Hao, M.F.; Li, M.D.; et al. Curcumin alleviates oxidative stress and inhibits apoptosis in diabetic cardiomyopathy via Sirt1-Foxo1 and PI3K-Akt signalling pathways. J. Cell Mol. Med. 2020. [Google Scholar] [CrossRef]
- Yao, Q.; Ke, Z.Q.; Guo, S.; Yang, X.S.; Zhang, F.X.; Liu, X.F.; Chen, X.; Chen, H.G.; Ke, H.Y.; Liu, C. Curcumin protects against diabetic cardiomyopathy by promoting autophagy and alleviating apoptosis. J. Mol. Cell Cardiol. 2018, 124, 26–34. [Google Scholar] [CrossRef]
- Li, H.; Sureda, A.; Devkota, H.P.; Pittalà, V.; Barreca, D.; Silva, A.S.; Tewari, D.; Xu, S.; Nabavi, S.M. Curcumin, the golden spice in treating cardiovascular diseases. Biotechnol. Adv. 2020, 38, 107343. [Google Scholar] [CrossRef]
- Moretti, H.D.; Colucci, V.J.; Berry, B.D. Vitamin D3 repletion versus placebo as adjunctive treatment of heart failure patient quality of life and hormonal indices: A randomized, double-blind, placebo-controlled trial. BMC Cardiovasc. Disord. 2017, 17, 274. [Google Scholar] [CrossRef]
- Panizo, S.; Barrio-Vázquez, S.; Naves-Díaz, M.; Carrillo-López, N.; Rodríguez, I.; Fernández-Vázquez, A.; Valdivielso, J.M.; Thadhani, R.; Cannata-Andía, J.B. Vitamin D receptor activation, left ventricular hypertrophy and myocardial fibrosis. Nephrol. Dial. Transplatation 2013, 28, 2735–2744. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Su, H.; Zhang, Y.; Kong, J. The protective effect of 1,25(OH)2D3 against cardiac hypertrophy is mediated by the cyclin-dependent kinase inhibitor p21. Eur. J. Pharmacol. 2020, 888, 173510. [Google Scholar] [CrossRef]
- Shibanuma, M.; Mashimo, J.; Mita, A.; Kuroki, T.; Nose, K. Cloning from a mouse osteoblastic cell line of a set of transforming-growth-factor-beta 1-regulated genes, one of which seems to encode a follistatin-related polypeptide. Eur. J. Biochem. 1993, 217, 13–19. [Google Scholar] [CrossRef]
- Mattiotti, A.; Prakash, S.; Barnett, P.; van den Hoff, M.J.B. Follistatin-like 1 in development and human diseases. Cell Mol. Life Sci. 2018, 75, 2339–2354. [Google Scholar] [CrossRef]
- Peters, M.M.C.; Meijs, T.A.; Gathier, W.; Doevendans, P.A.M.; Sluijter, J.P.G.; Chamuleau, S.A.J.; Neef, K. Follistatin-like 1 in Cardiovascular Disease and Inflammation. Mini Rev. Med. Chem. 2019, 19, 1379–1389. [Google Scholar] [CrossRef]
- Wollert, K.C. Follistatin-Like 1: Growing Evidence for Its Beneficial Role in Heart Failure. JACC Basic Transl. Sci. 2016, 1, 222–223. [Google Scholar] [CrossRef][Green Version]
- Yang, W.; Duan, Q.; Zhu, X.; Tao, K.; Dong, A. Follistatin-Like 1 Attenuates Ischemia/Reperfusion Injury in Cardiomyocytes via Regulation of Autophagy. Biomed. Res. Int. 2019, 2019, 9537382. [Google Scholar] [CrossRef]
- Tanaka, K.; Valero-Muñoz, M.; Wilson, R.M.; Essick, E.E.; Fowler, C.T.; Nakamura, K.; van den Hoff, M.; Ouchi, N.; Sam, F. Follistatin like 1 Regulates Hypertrophy in Heart Failure with Preserved Ejection Fraction. JACC Basic Transl. Sci. 2016, 1, 207–221. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, J.; Zhang, W.; Peng, M.; Chen, J.; Zheng, L.; Zhang, X.; Yang, H.; Liu, Y. Follistatin-Like 1 Protects against Doxorubicin-Induced Cardiomyopathy through Upregulation of Nrf2. Oxid. Med. Cell Longev. 2020, 2020, 3598715. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, T.; Mokou, M.; Li, L.; Li, P.; Song, J.; Liu, H.; Zhu, Z.; Liu, D.; Yang, M.; et al. Follistatin-like 1 as a novel adipo-myokine related to insulin resistance and physical activity. J. Clin. Endocrinol. Metab. 2020. [Google Scholar] [CrossRef]
- Kon, M.; Tanimura, Y.; Yoshizato, H. Effects of acute endurance exercise on follistatin-like 1 and apelin in the circulation and metabolic organs in rats. Arch. Physiol. Biochem. 2020, 15, 1–5. [Google Scholar] [CrossRef]
- Fang, D.; Shi, X.; Lu, T.; Ruan, H.; Gao, Y. The glycoprotein follistatin-like 1 promotes brown adipose thermogenesis. Metabolism 2019, 98, 16–26. [Google Scholar] [CrossRef]
- Jafari Salim, S.; Alizadeh, S.; Djalali, M.; Nematipour, E.; Hassan Javanbakht, M. Effect of Omega-3 Polyunsaturated Fatty Acids Supplementation on Body Composition and Circulating Levels of Follistatin-Like 1 in Males With Coronary Artery Disease: A Randomized Double-Blind Clinical Trial. Am. J. Mens. Health 2017, 11, 1758–1764. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.Y.; Liu, Y.; Han, Y.G.; Li, J.K.; Jia, J.L.; Chen, B.; Yao, Z.X.; Nie, L.; Cheng, L. Follistatin-like protein 1 suppressed pro-inflammatory cytokines expression during neuroinflammation induced by lipopolysaccharide. J. Mol. Histol. 2017, 48, 63–72. [Google Scholar] [CrossRef]
- Murakami, K.; Tanaka, M.; Usui, T.; Kawabata, D.; Shiomi, A.; Iguchi-Hashimoto, M.; Shimizu, M.; Yukawa, N.; Yoshifuji, H.; Nojima, T.; et al. Follistatin-related protein/follistatin-like 1 evokes an innate immune response via CD14 and toll-like receptor 4. FEBS Lett. 2012, 586, 319–324. [Google Scholar] [CrossRef]
- Ward, W.E.; Chilibeck, P.D.; Comelli, E.M.; Duncan, A.M.; Phillips, S.M.; Robinson, L.E.; Stellingwerff, T. Research in nutritional supplements and nutraceuticals for health, physical activity, and performance: Moving forward 1. Appl. Physiol. Nutr. Metab. 2019, 44, 455–460. [Google Scholar] [CrossRef]
- Koletzko, B.; Godfrey, K.M.; Poston, L.; Szajewska, H.; van Goudoever, J.B.; de Waard, M.; Brands, B.; Grivell, R.M.; Deussen, A.R.; Dodd, J.M.; et al. EarlyNutrition Project Systematic Review Group. Nutrition During Pregnancy, Lactation and Early Childhood and its Implications for Maternal and Long-Term Child Health: The Early Nutrition Project Recommendations. Ann. Nutr. Metab. 2019, 74, 93–106. [Google Scholar] [CrossRef]
- Lind, M.V.; Larnkjær, A.; Mølgaard, C.; Michaelsen, K.F. Breastfeeding, Breast Milk Composition, and Growth Outcomes. Nestle Nutr. Inst. Workshop Ser. 2018, 89, 63–77. [Google Scholar] [CrossRef]
- Palou, M.; Picó, C.; Palou, A. Leptin as a breast milk component for the prevention of obesity. Nutr. Rev. 2018, 76, 875–892. [Google Scholar] [CrossRef]
- Kratzsch, J.; Bae, Y.J.; Kiess, W. Adipokines in human breast milk. Best Pract. Res. Clin. Endocrinol. Metab. 2018, 32, 27–38. [Google Scholar] [CrossRef]
- Pomar, C.A.; Sánchez, J.; Palou, A. The Intake of a Cafeteria Diet in Nursing Rats Alters the Breast Milk Concentration of Proteins Important for the Development of Offspring. Nutrients 2020, 12, 2470. [Google Scholar] [CrossRef] [PubMed]
- Psaltopoulou, T.; Hatzis, G.; Papageorgiou, N.; Androulakis, E.; Briasoulis, A.; Tousoulis, D. Socioeconomic status and risk factors for cardiovascular disease: Impact of dietary mediators. Hell. J. Cardiol. 2017, 58, 32–42. [Google Scholar] [CrossRef]


{kind=link}
| Biological Action | Adipo-myo-cardiokines | Type of Study |
|---|---|---|
| ↓left ventricular hypertrophy | ↓leptin resistance | Obese mice a |
| ↓myocardial function | ↑adiponectin levels | Obese mice b |
| ↓weight loss | ↑adiponectin levels | Rats b |
| ↑body composition, ↑metabolic parameters | ↑adiponectin levels | Obese and overweight former athletes b |
| ↑liver steatosis | ↑adiponectin levels | Obese mice b |
| ↑resting metabolic rate | ↑adiponectin levels | Obese or overweight women b,* |
| ↑insulin sensitivity | ↑apelin expression and levels | T2DM patients c,* |
| ↑metabolic state | = apelin levels | Healthy adult men c |
| ↑aerobic exercise | ↑irisin levels | Obese rats d,* |
| Type of Intervention | Biological Action | Adipo-myo-cardiokines | Type of Study |
|---|---|---|---|
| Lactobacillus rhamnosus, L. acidophilus, Bifidobacterium bifidumi | ↑insulin sensitivity | ↓leptin resistance | Obese mice a |
| Probiotic mixture | ↓liver steatosis inflammatory state | ↓leptin levels | Mouse model of NAFLD a |
| Oligofructose dietary fiber + lifestyle modifications | ↑metabolic condition | ↓leptin levels = adiponectin levels | Patients with NAFLD a |
| Multistrain probiotic | ↑metabolic and cardiac state ↓ inflammatory state | =leptin levels ↑adiponectin levels | T2DM patients a |
| Oligofructose-enriched inulin/d | ↑satiety | ↑adiponectin levels | Children with overweight and obesity b |
| Bifidobacterium lactis HN019 | ↑nitric oxide levels | ↑adiponectin levels | Subjects with and without the metabolic syndrome b |
| Probiotic soy milk | ↑lipid profile | =adiponectin levels | T2DM patients b |
| Synbiotic | no significant beneficial metabolic effects | ↓apelin levels | PCOS women c |
| L. plantarum | ↑browning and thermogenesis of adipose tissue | ↑irisin levels | Obese mice d |
| Lactobacillus rhamnosus GR-1 | ↓myocardial hypertrophy | ↓ANP levels | Rats after myocardial infarction e |
| Type of Intervention | Biological Action | Adipo-myo-cardiokines | Type of Study |
|---|---|---|---|
| EPA-DHA + lifestyle modifications | ↓inflammatory state ↑metabolic state | =leptin levels | Obese women a |
| Omega-3 + lifestyle modifications | ↓insulin resistance ↓endothelial dysfunction | ↓leptin levels | Obese adolescents a |
| Linolenic acid | ↑cardiometabolic parameters ↓inflammatory state | ↓leptin levels | Obese-diabetic mice a |
| 3-n PUFA | ↑cardiometabolic state ↓inflammatory state | ↓leptin levels | Obese mice a |
| ↑insulin sensitivity, improve blood pressure | ↑irisin levels | T2DM patients b | |
| ↑heart failure | ↓BNP levels | Meta-analysis of randomized controlled trials c | |
| ↓inflammatory state | ↓follistatin-like 1 levels | Patients with coronary artery disease d | |
| 2-hydroxyoleic acid (2-OHOA) and n-3 PUFA | ↑body composition ↑cardioprotection mechanims | ↑adiponectin levels | Obese mice e |
| EPA | ↑insulin signaling | ↑apelin levels | Lean/obese mice f |
| ↑insulin signaling in adipose tissue | ↑apelin levels | Lean/overweight rats f |
| Type of Intervention | Biological Action | Adipo-myo-cardiokines | Type of Study |
|---|---|---|---|
| Combination of quercetin and resveratrol | ↑body composition ↓ inflammatory state and gut dysbiosis | ↑leptin levels | Obese mice a |
| Lycopene | ↓inflammatory state | ↓leptin expression and levels | Obese mice a |
| Resveratrol | ↑body composition | ↑adiponectin levels, = leptin | Meta-analysis of randomized controlled trials b |
| ↓cardiac fibrosis ↓inflammatory state | ↓ANP and BNP levels | Cardiomyocytes (in vitro) c | |
| Curcumin | ↑body composition | ↑adiponectin levels | Meta-analysis of randomized controlled trials-obese mice d |
| ↑insulin sensitivity | ↓apelin levels | T2DM rats e | |
| ↓cardiac hypertrophy and fibrosis | ↓ANP and BNP levels | Rats f | |
| ↓cardiac hypertrophy oxidative stress | ↓ANP and BNP levels | Diabetic rats f | |
| Genistein | ↑browning of adipose tissue | ↑irisin levels | Adipocytes-mice g |
| Silymarin | ↑hepatic condition ↑anti-oxidative mechanisms | ↑irisin levels | T2DM rats g |
| Type of Intervention | Biological Action | Adipo-myo-cardiokines | Type of Study |
|---|---|---|---|
| Walnut oil | ↑insulin sensitivity ↑antioxidative capacity | ↓leptin levels | Obese rats a |
| Mixed nuts | ↑satiety | ↓leptin levels | Overweight and obese adults a |
| Coffee | ↓inflammatory state | ↑adiponectin levels | Women with or without type 2 diabetes b |
| ↑body composition ↑metabolic state | ↑adiponectin levels | Diabetic rats b | |
| Green tea extract + exercise | ↑metabolic state ↓inflammatory state | ↑adiponectin levels | Overweight middle-aged men b |
| Green cardamom | ↑liver steatosis and insulin signaling ↓inflammatory state | ↑irisin levels | Overweight or obese with NAFLD c |
| Grape pomace extract | ↑browning of adipose tissue | ↑irisin levels | Obese rats c |
| Probiotic-fermented purple sweet potato yogurt | ↓cardiac hypertrophy | ↓ANP and BNP levels | Hypertensive rat d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Senesi, P.; Luzi, L.; Terruzzi, I. Adipokines, Myokines, and Cardiokines: The Role of Nutritional Interventions. Int. J. Mol. Sci. 2020, 21, 8372. https://doi.org/10.3390/ijms21218372
Senesi P, Luzi L, Terruzzi I. Adipokines, Myokines, and Cardiokines: The Role of Nutritional Interventions. International Journal of Molecular Sciences. 2020; 21(21):8372. https://doi.org/10.3390/ijms21218372
Chicago/Turabian StyleSenesi, Pamela, Livio Luzi, and Ileana Terruzzi. 2020. "Adipokines, Myokines, and Cardiokines: The Role of Nutritional Interventions" International Journal of Molecular Sciences 21, no. 21: 8372. https://doi.org/10.3390/ijms21218372
APA StyleSenesi, P., Luzi, L., & Terruzzi, I. (2020). Adipokines, Myokines, and Cardiokines: The Role of Nutritional Interventions. International Journal of Molecular Sciences, 21(21), 8372. https://doi.org/10.3390/ijms21218372