Intramolecular Lactones of Sialic Acids



Abstract
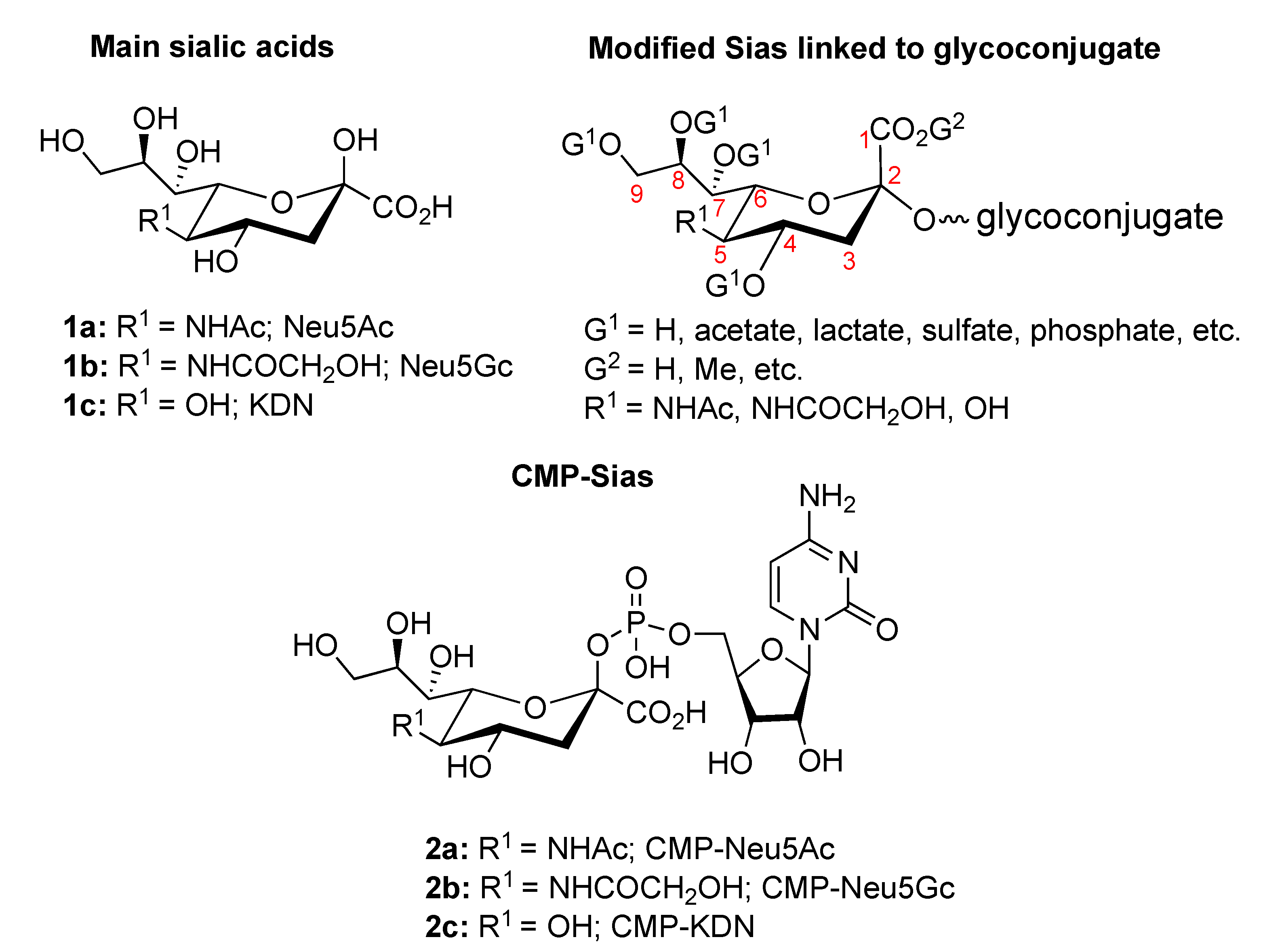
:1. Introduction
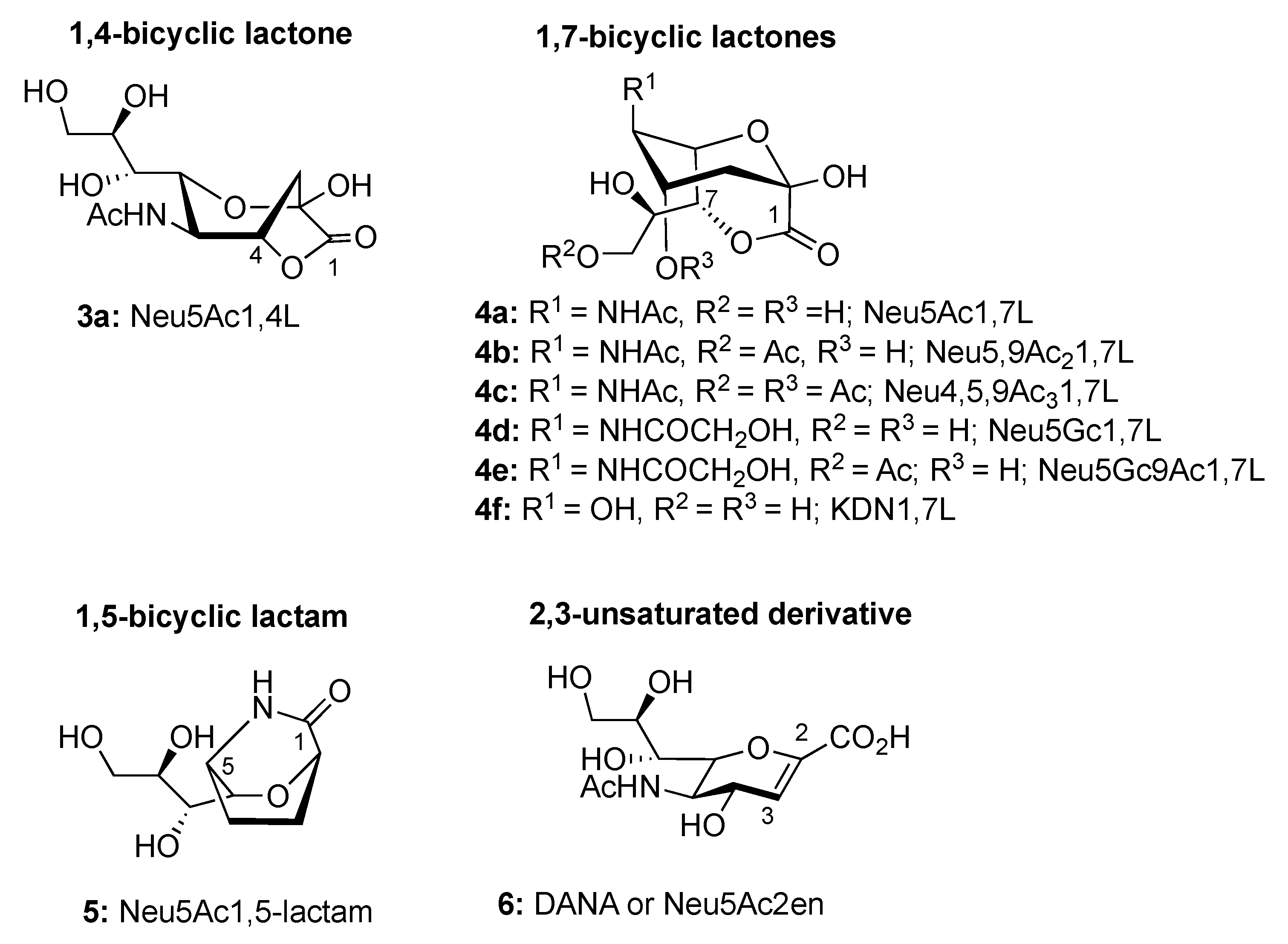
2. Overview of the Sias Intramolecular Lactones: Synthesis and Rearrangement Mechanism
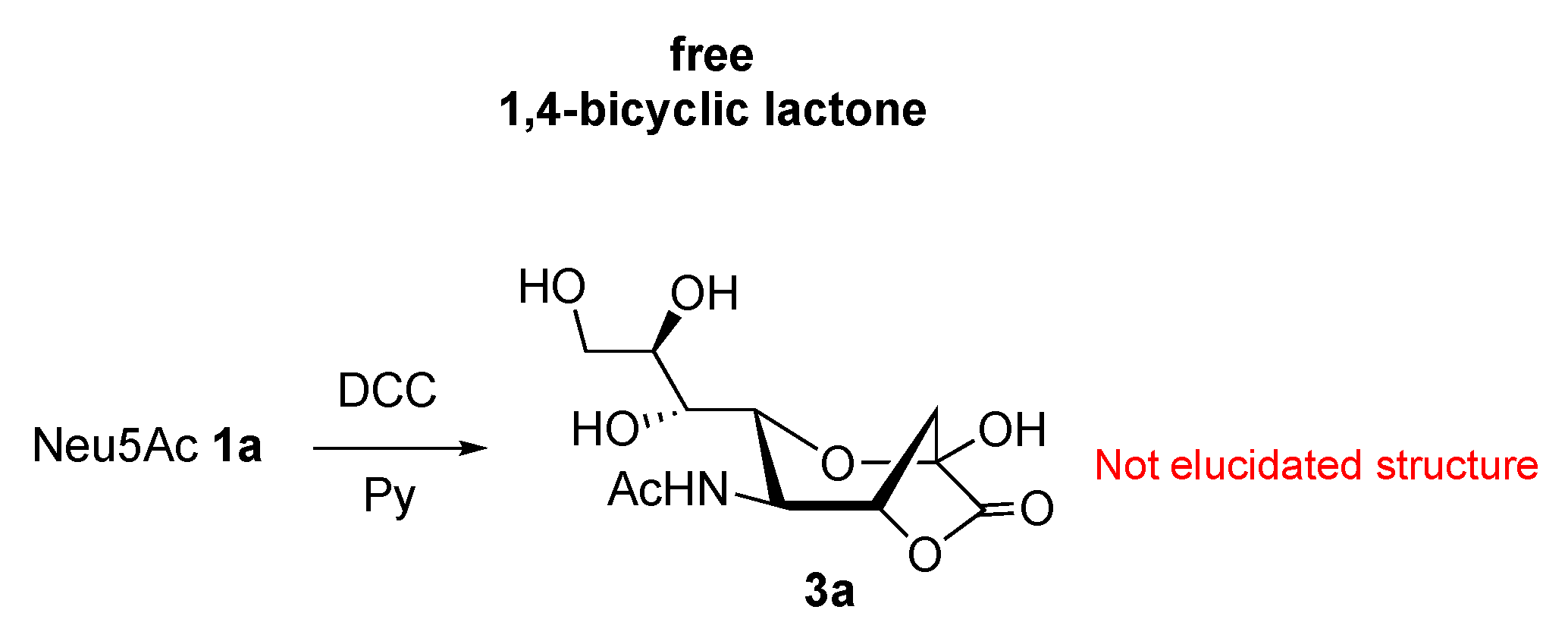
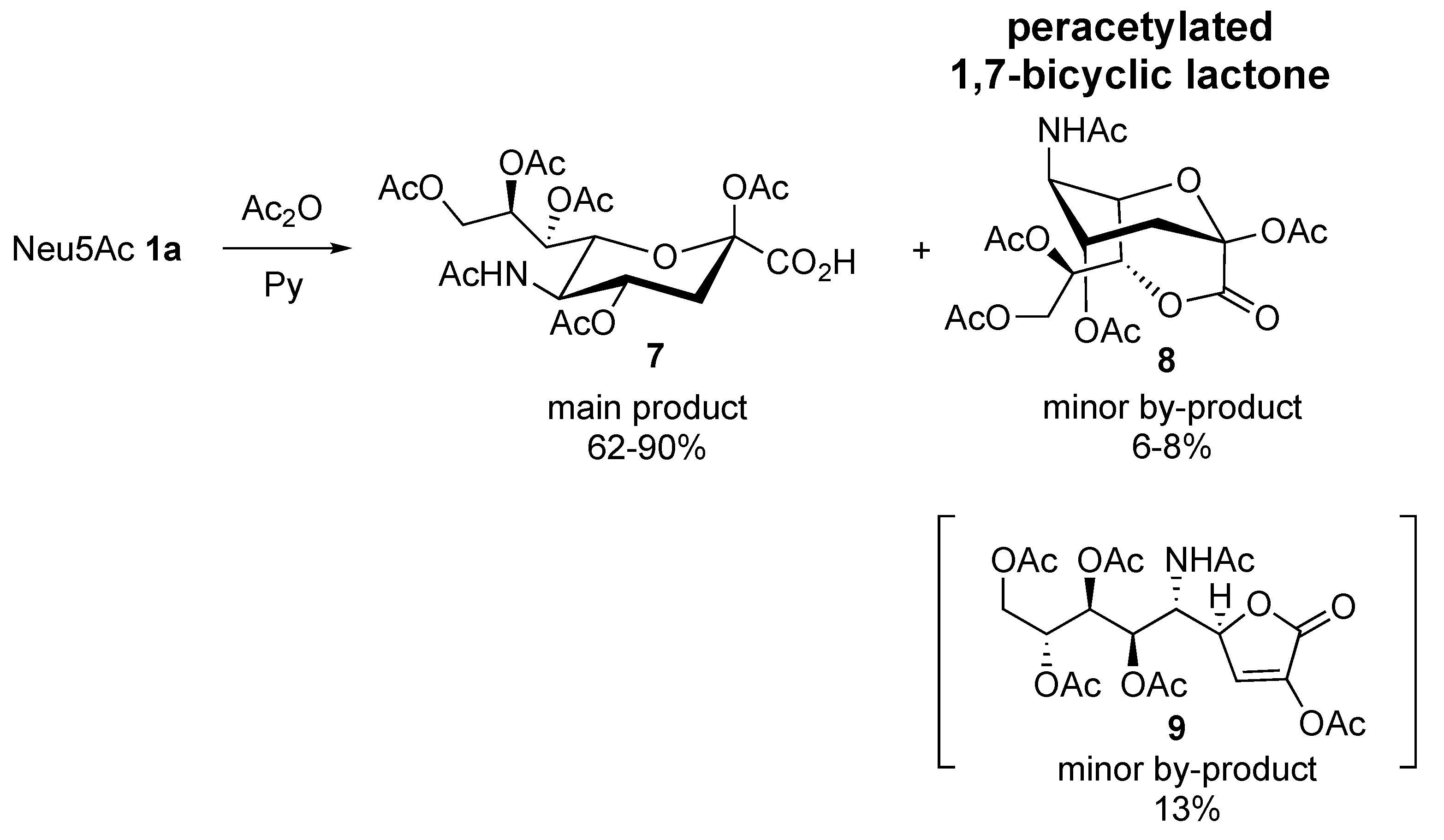
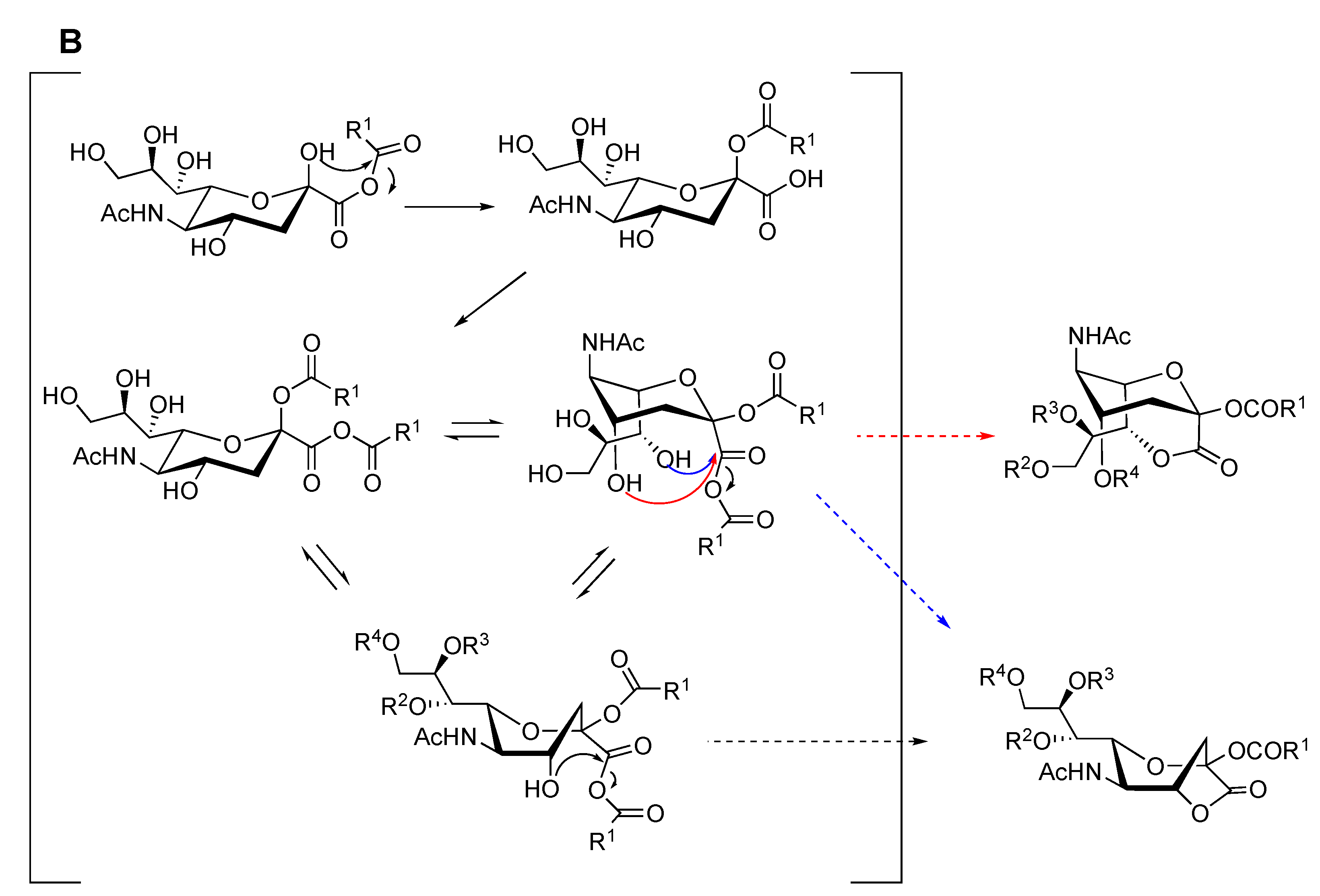
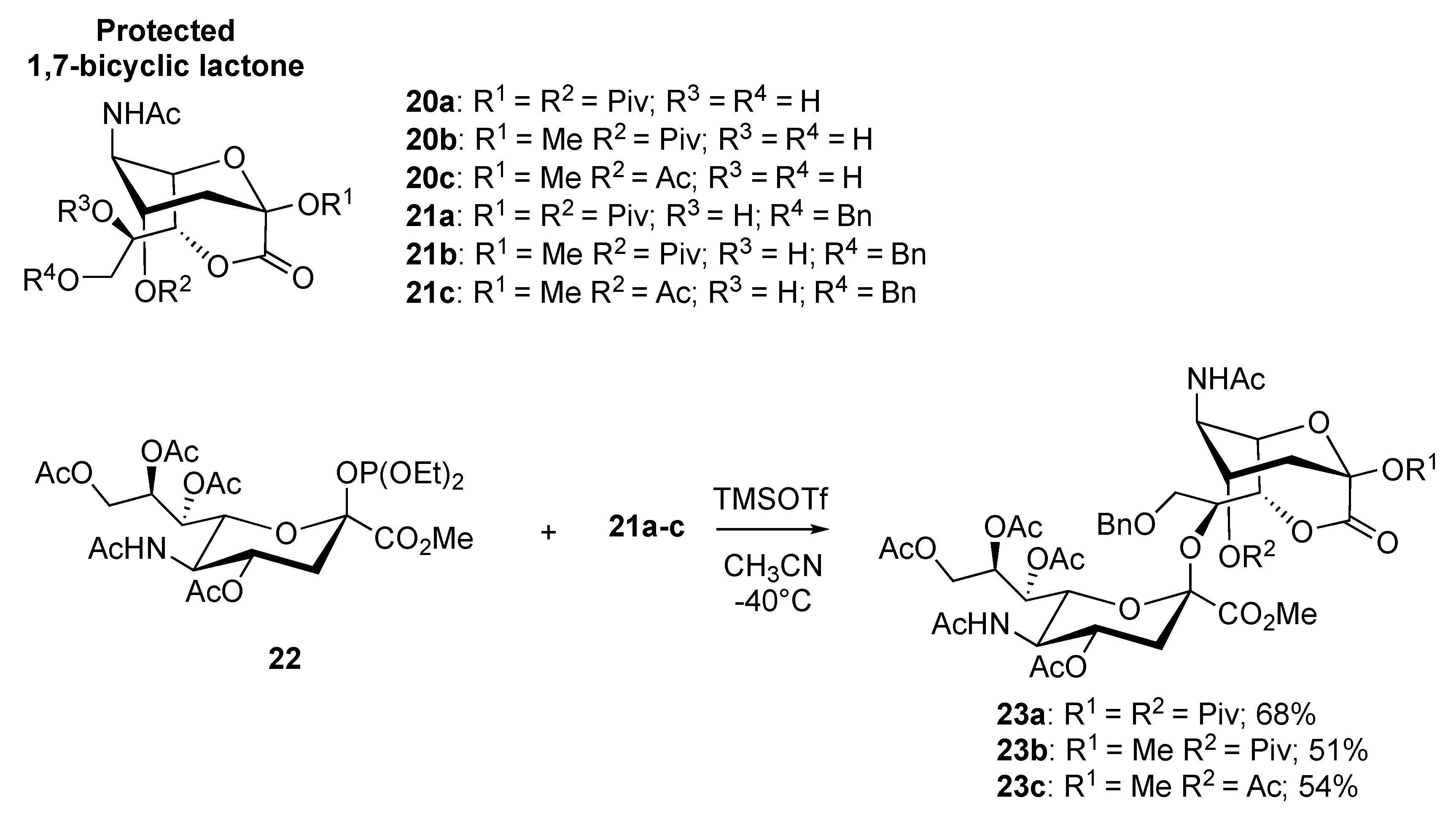
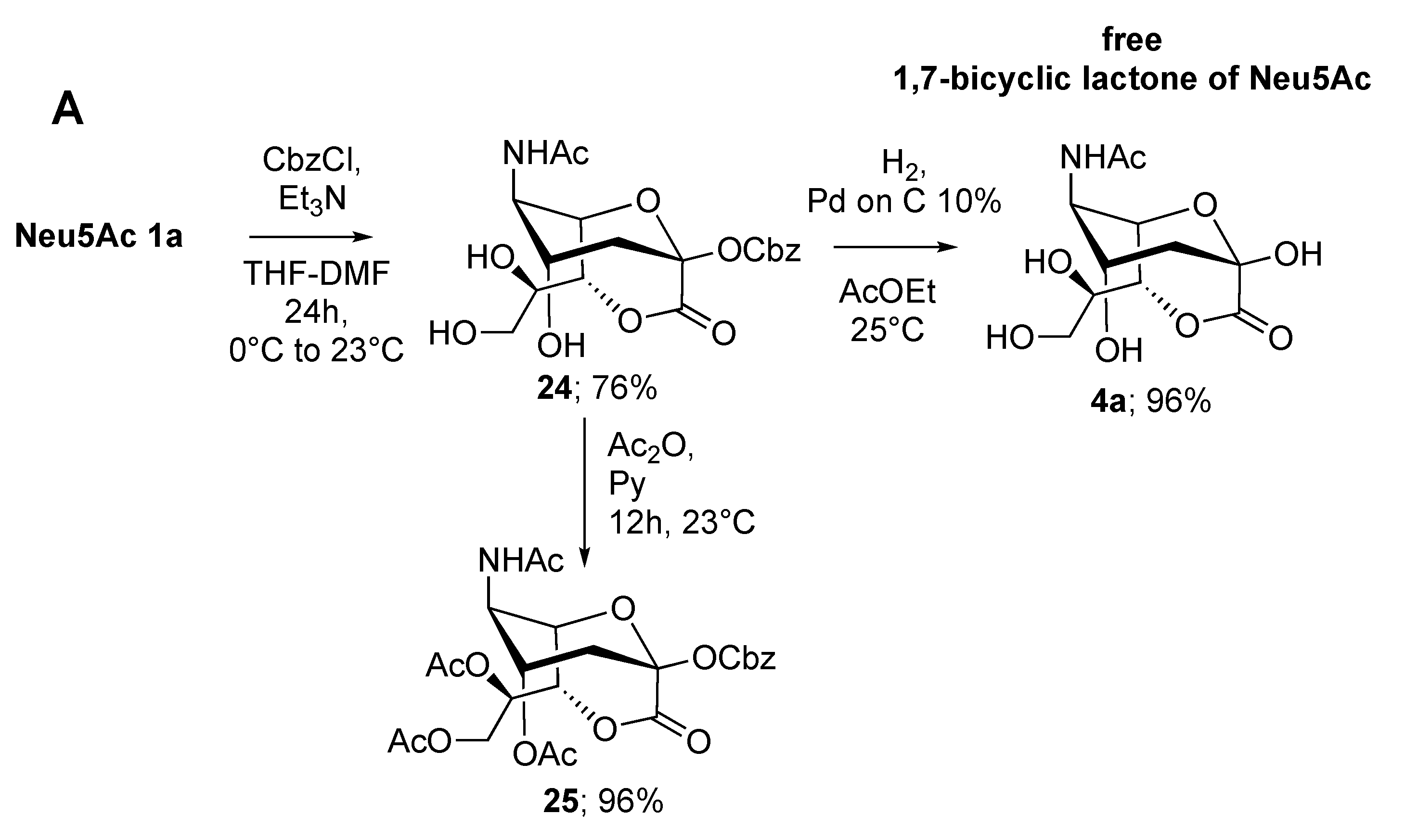
2.1. Synthesis of 1,4- and 1,7-bicyclic Lactones of Neu5Ac
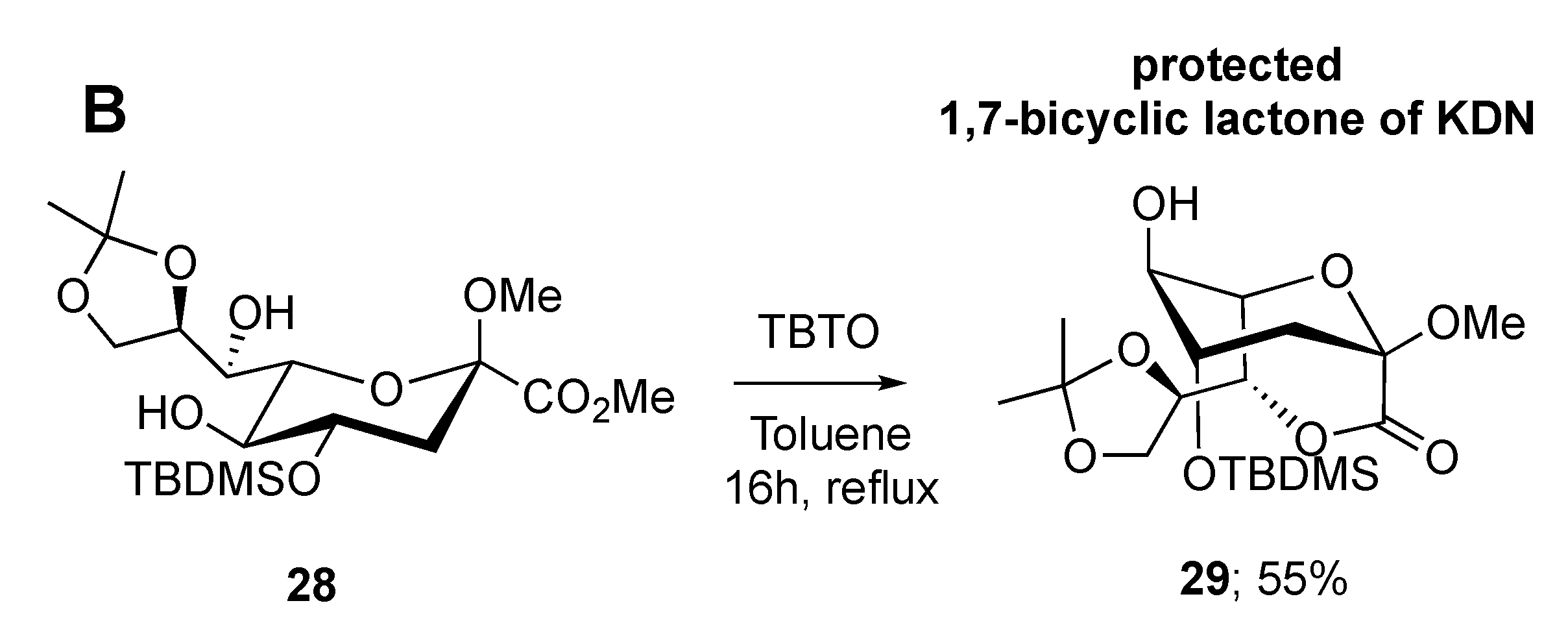
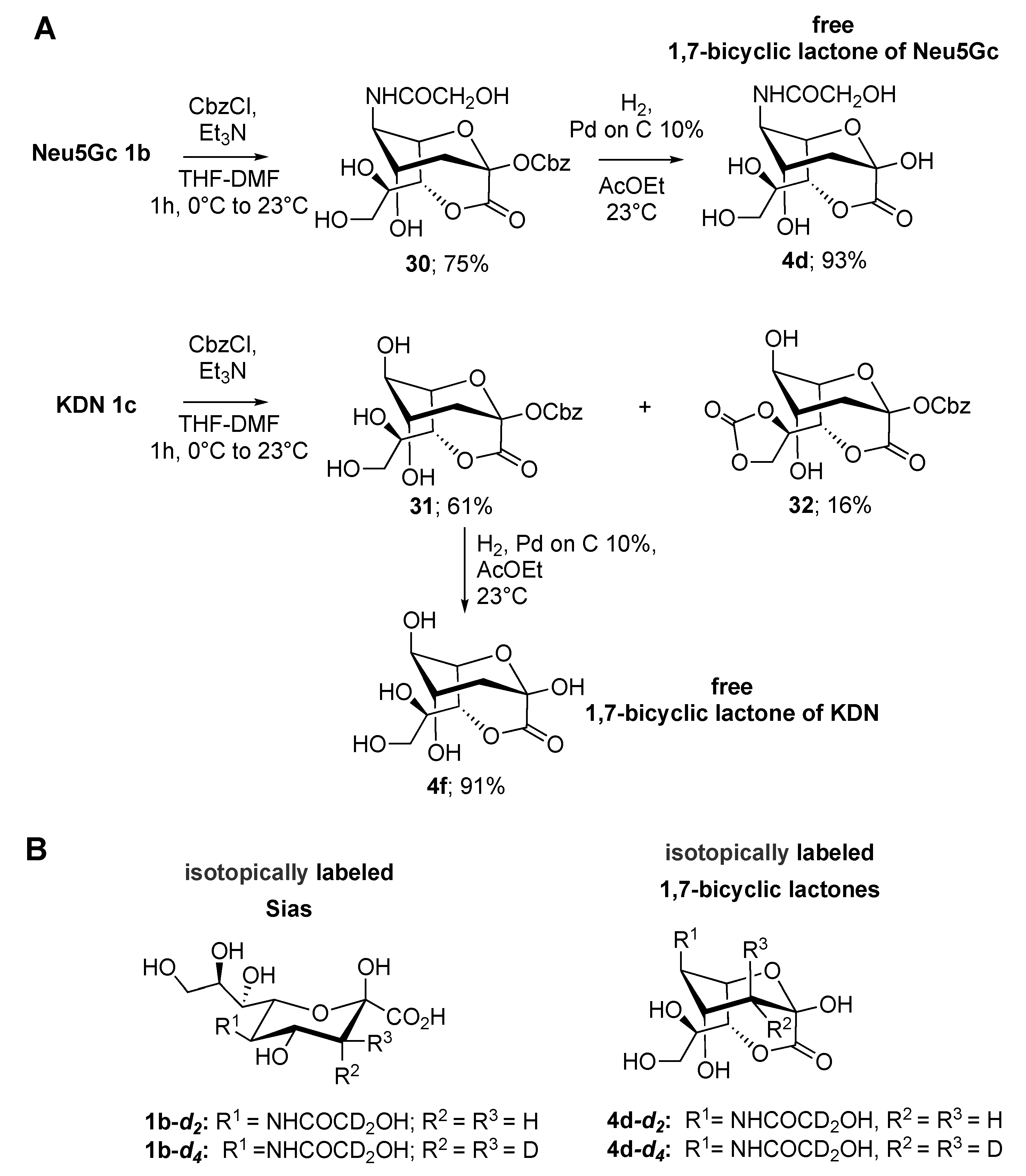
2.2. Synthesis of 1,4- and 1,7-bicyclic Lactones of Neu5Gc and KDN
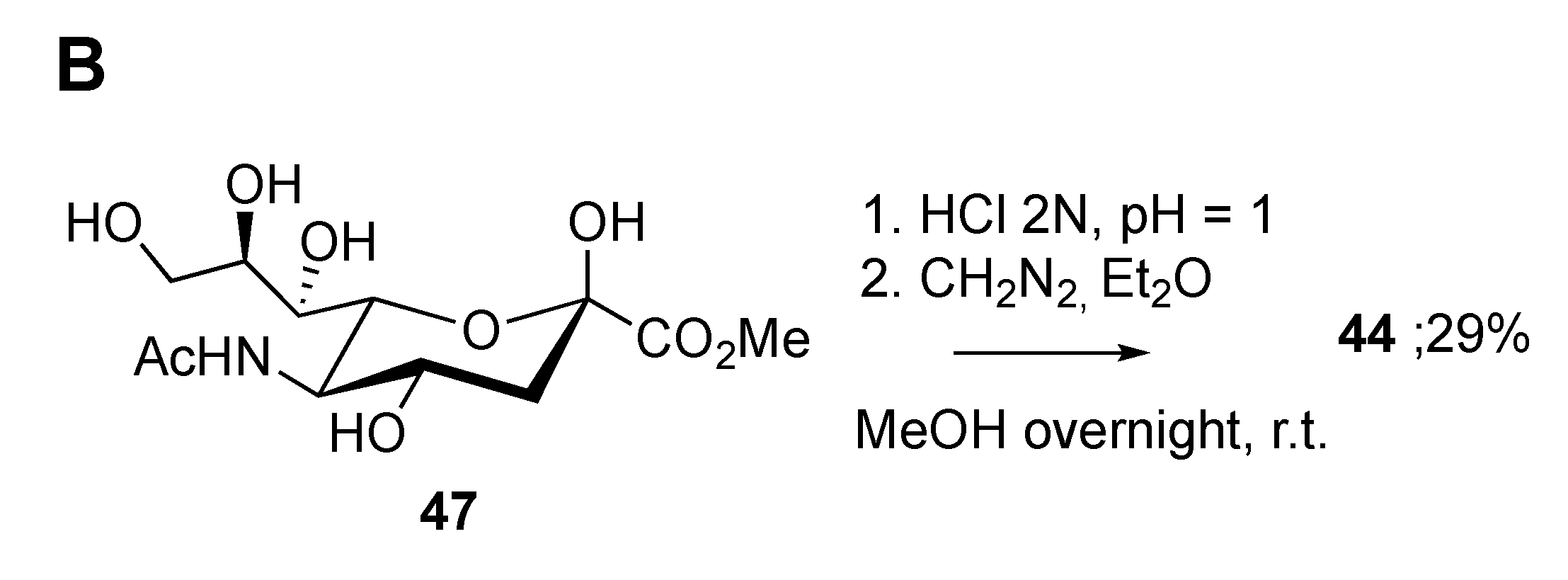
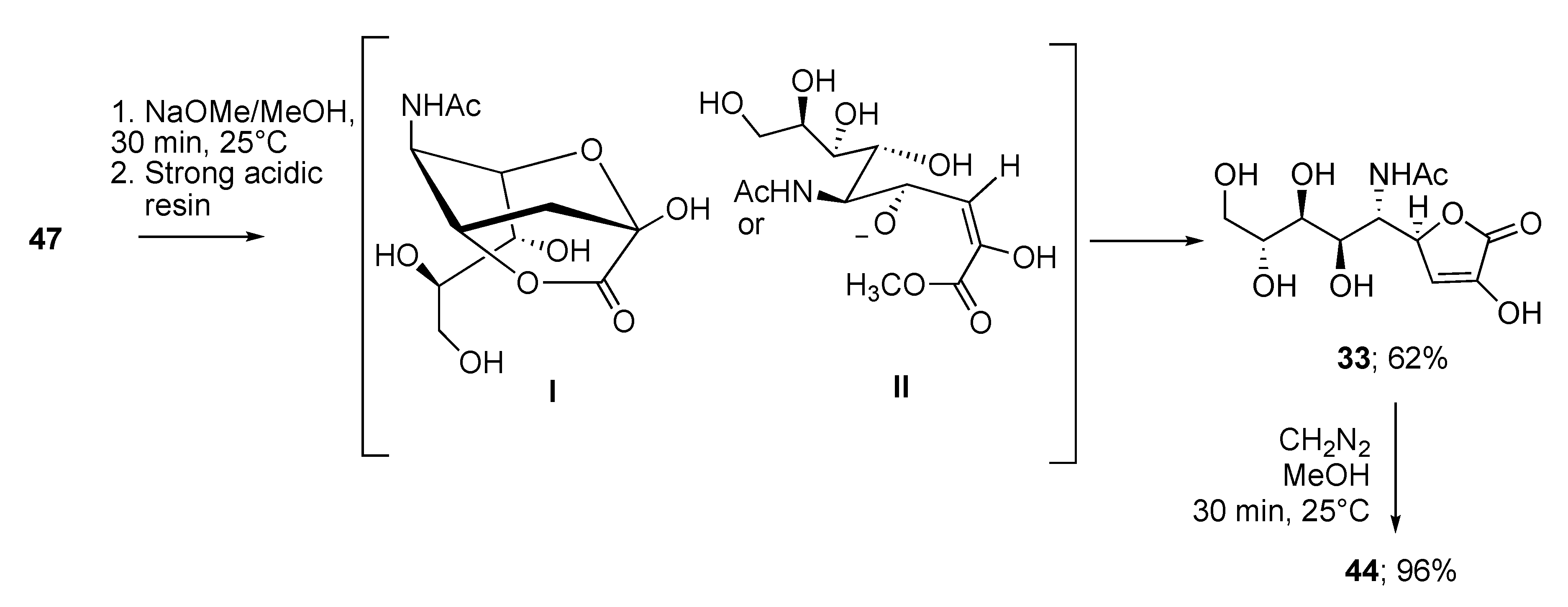
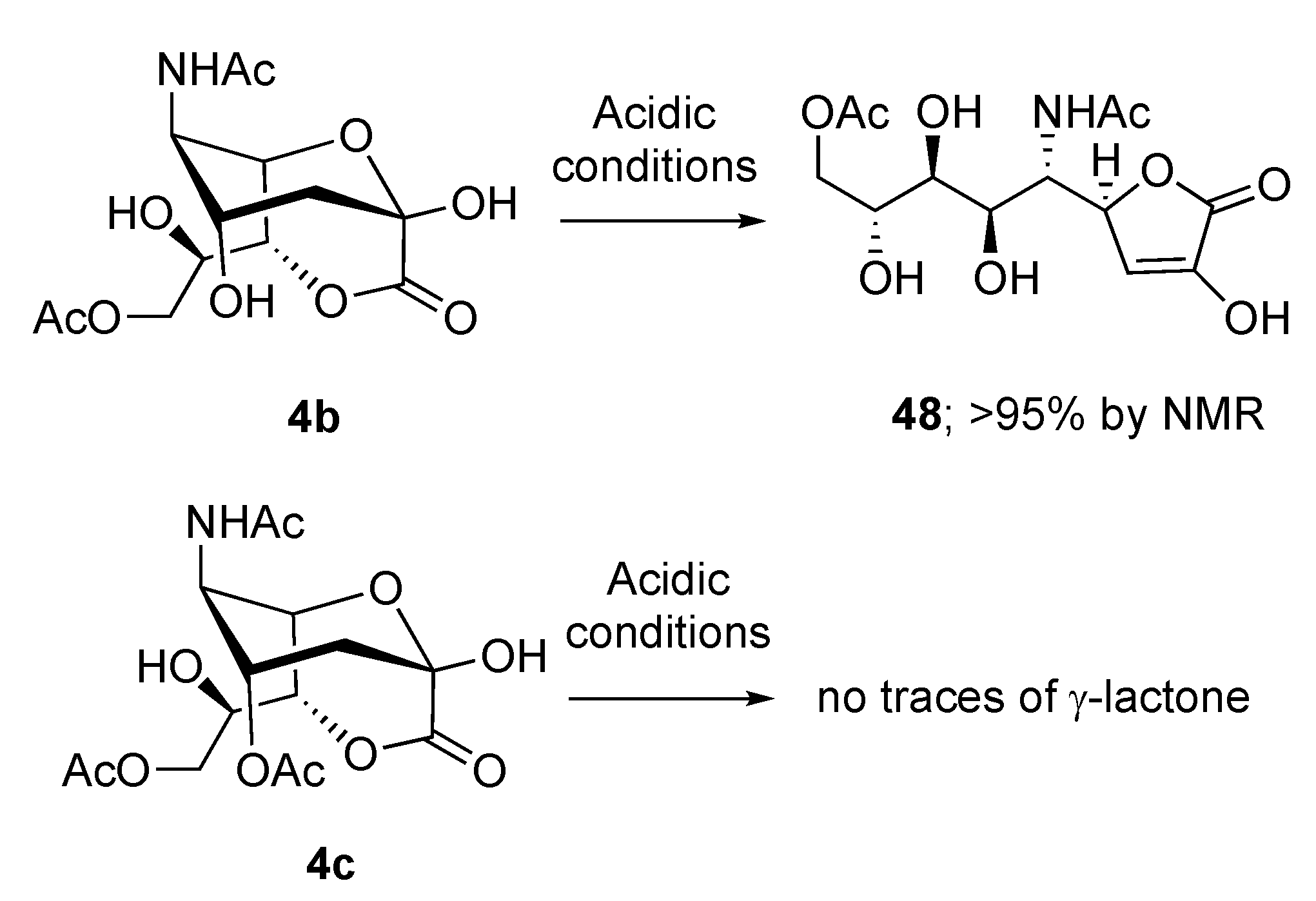
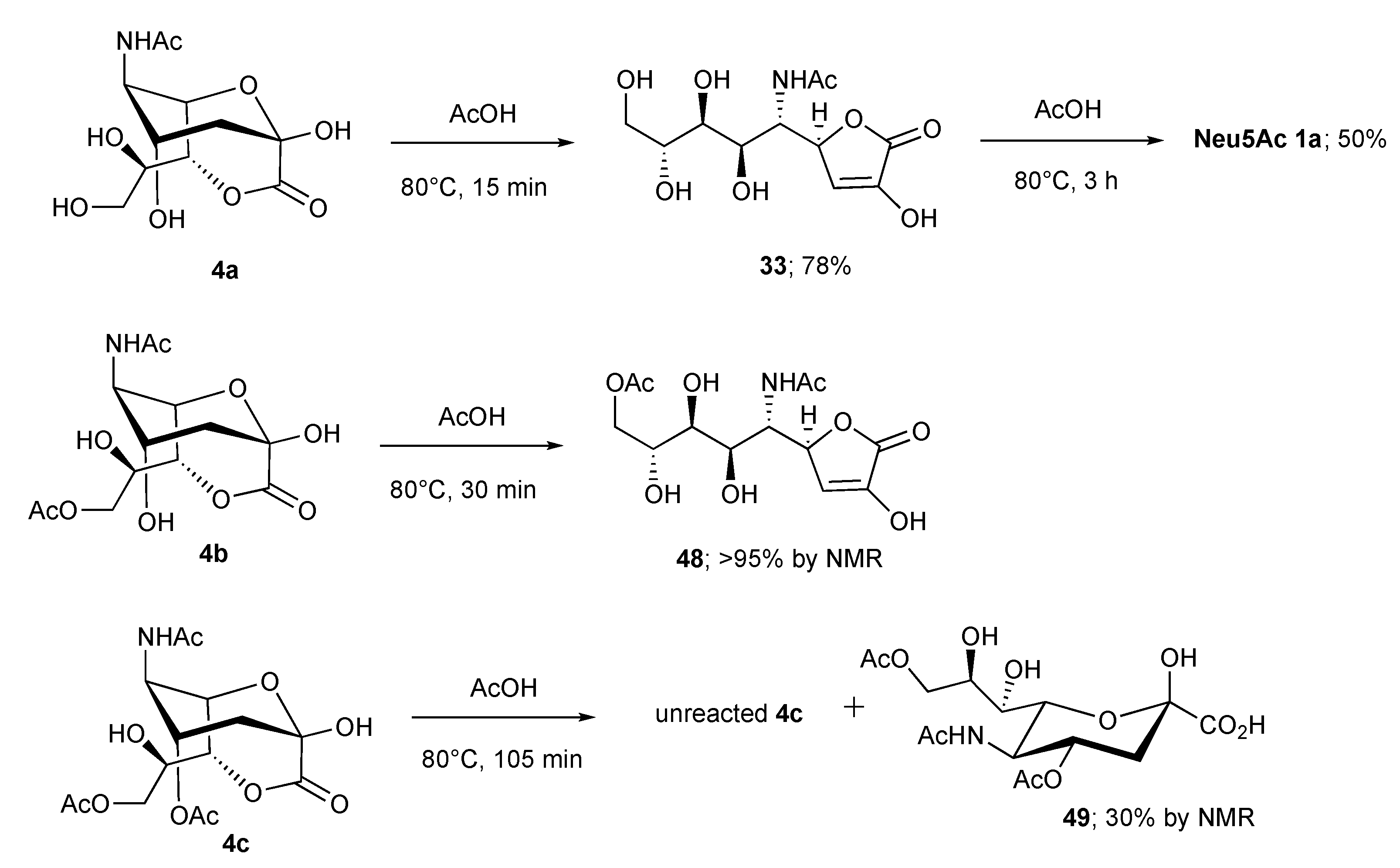
2.3. Synthesis of γ-Lactones of Sias
3. Biological Relevance of Neu5ac 1,7-Lactones
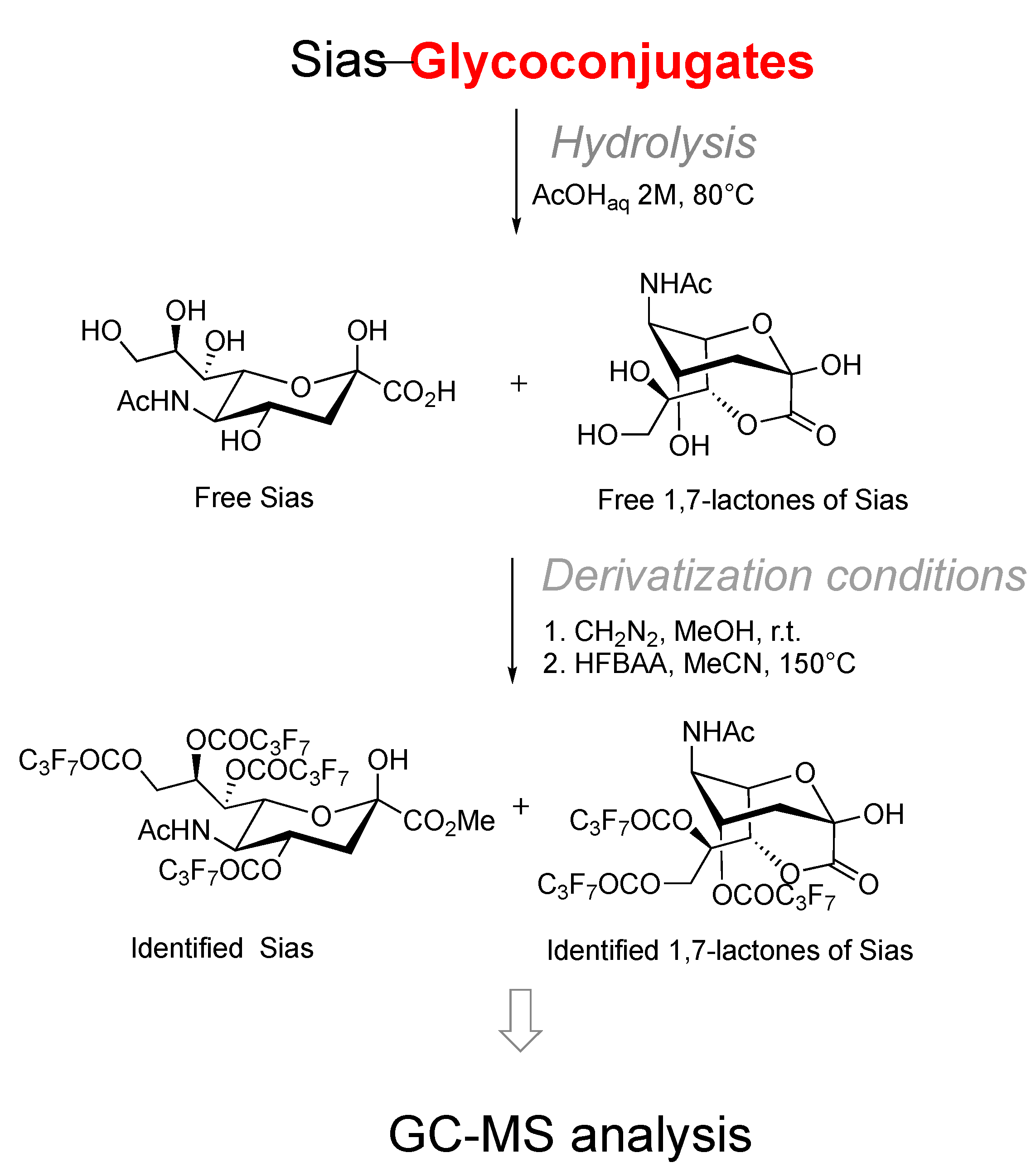
3.1. Analytical and Biochemical Techniques Applied to Identify Neu5Ac 1,7-Lactones
3.2. 1,7-Lactones of Sias in Biological Samples
3.2.1. Animal Tissues
3.2.2. Human Healthy and Diseased Tissues
3.3. Recent Insights into the Biological Role of Neu5Ac 1,7-Lactone
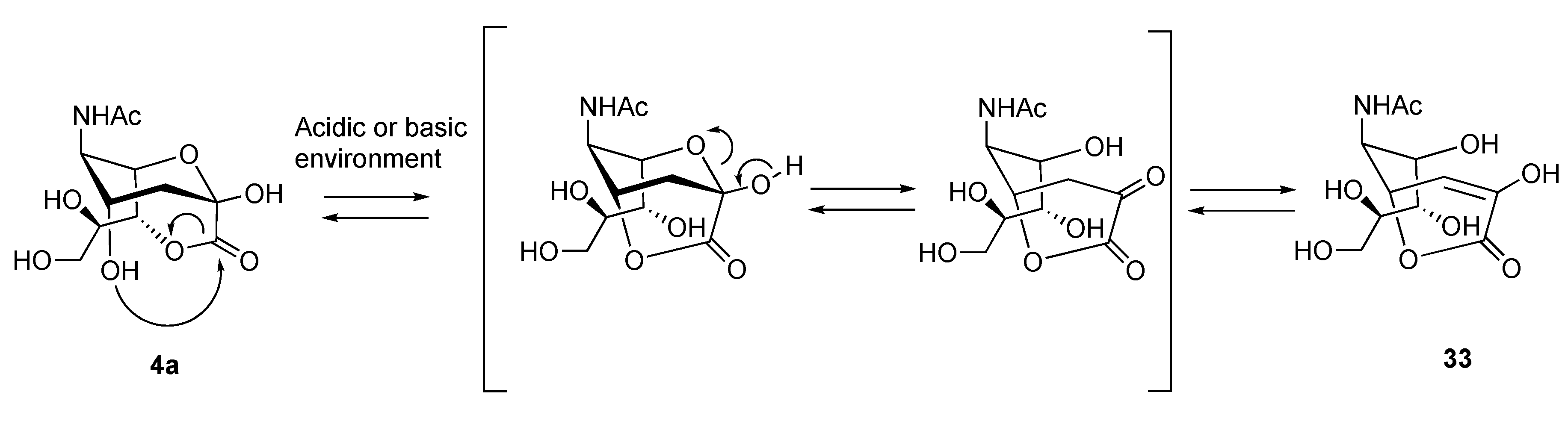
3.4. Stability of the Lactones of Biological Interest and Questioning on the Real Presence of 1,7-Lactones of Sias in Biological Tissues
4. Conclusions and Future Perspectives
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AcOH | Acetic acid |
| CMP | Cytidine mononucleotide |
| Cs2CO3 | Cesium carbonate |
| DANA | 2-Deoxy-2,3-dehydro N-acetylneuraminic acid |
| DCC | N,N′-dicyclohexylcarbodiimide |
| DMF | Dimethylformamide |
| GC | Gas chromatography |
| GC-MS | Gas chromatography–mass spectrometry |
| GM3 | Monosialodihexosylganglioside |
| HCl | Hydrochloric acid |
| HFB | Heptafluorobutyrate |
| HFBAA | heptafluorobutyric anhydride |
| HPLC | High-performance liquid chromatography |
| IL-4 | Interleukin 4 |
| kDa | Kilodalton |
| KDN | 3-Deoxy-D-glycero-D-galacto-2-nonulosonic acid or 2-keto-3-deoxy-D-glycero-D-galacto-nononic acid |
| MeCN | Acetonitrile |
| MeOH | Methanol |
| MS | Mass spectrometry |
| NADH | Nicotinamide adenine dinucleotide, reduced |
| NaOH | Sodium hydroxide |
| NaOMe | Sodium methoxide |
| NBS | N-Bromosuccinimide |
| Neu | 5-Amino-3,5-dideoxy-D-glycero-D-galacto-2-nonulosonic acid or Neuraminic acid |
| Neu5Ac | N-acetylneuraminic acid |
| Neu5Ac1,7L | N-acetylneuraminic acid 1,7-lactone |
| Neu5Gc | N-glycolylneuraminic acid |
| NMR | Nuclear magnetic resonance |
| RBC | Red blood cell |
| Sias | Sialic acids |
| THF | Tetrahydrofuran |
| TMG | 1,1,3,3–Tetramethylguanidine |
References
- Schauer, R.; Kamerling, J.P. Exploration of the sialic acid world. Adv. Carbohydr. Chem. Biochem. 2018, 75, 1–213. [Google Scholar] [CrossRef] [PubMed]
- Schauer, R. Sialic acids as regulators of molecular and cellular interactions. Curr. Opin. Struct. Biol. 2009, 19, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Schauer, R. Sialic acids: Fascinating sugars in higher animals and man. Zoology 2004, 107, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Varki, A.; Cummings, R.D.; Esko, J.D.; Stanley, P.; Hart, G.W.; Aebi, M.; Darvill, A.G.; Kinoshita, T.; Packer, N.H.; Prestegard, J.H.; et al. Essentials of Glycobiology, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2015. [Google Scholar]
- Angata, T.; Varki, A. Chemical diversity in the sialic acids and related alpha-keto acids: An evolutionary perspective. Chem. Rev. 2002, 102, 439–469. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Varki, A. Advances in the biology and chemistry of sialic acids. ACS Chem. Biol. 2010, 5, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Schauer, R.; Srinivasan, G.V.; Wipfler, D.; Kniep, B.; Schwartz-Albiez, R. O-Acetylated Sialic Acids and Their Role in Immune Defense. In The Molecular Immunology of Complex Carbohydrates-3. Advances in Experimental Medicine and Biology; Wu, A., Ed.; Springer: Boston, MA, USA, 2011; Volume 705, pp. 525–548. [Google Scholar] [CrossRef]
- Schauer, R. Achievements and challenges of sialic acid research. Glycoconj. J. 2000, 17, 485–499. [Google Scholar] [CrossRef]
- Kooner, A.S.; Yu, H.; Chen, X. Synthesis of N-glycolylneuraminic acid (Neu5Gc) and its glycosides. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Varki, A. Loss of N-glycolylneuraminic acid in humans: Mechanisms, consequences, and implications for hominid evolution. Am. J. Phys. Anthropol. 2001, 116, 54–69. [Google Scholar] [CrossRef]
- Kawano, T.; Koyama, S.; Takematsu, H.; Kozutsumi, Y.; Kawasaki, H.; Kawashima, S.; Kawasaki, T.; Suzuki, A. Molecular cloning of cytidine monophospho-N-acetylneuraminic acid hydroxylase. Regulation of species- and tissue-specific expression of N-glycolylneuraminic acid. J. Biol. Chem. 1995, 270, 16458–16463. [Google Scholar] [CrossRef] [Green Version]
- Shaw, L.; Schneckenburger, P.; Carlsen, J.; Christiansen, K.; Schauer, R. Mouse liver cytidine-5′-monophosphate-N-acetylneuraminic acid hydroxylase. Eur. J. Biochem. 1992, 206, 269–277. [Google Scholar] [CrossRef]
- Samraj, A.; Läubli, H.; Varki, N.; Varki, A. Involvement of a non-human sialic acid in human cancer. Front. Oncol. 2014, 4. [Google Scholar] [CrossRef] [Green Version]
- Kozutsumi, Y.; Kawano, T.; Kawasaki, H.; Suzuki, K.; Yamakawa, T.; Suzuki, A. Reconstitution of CMP-N-acetylneuraminic acid hydroxylation activity using a mouse liver cytosol fraction and soluble cytochrome b5 purified from horse erythrocytes. J. Biochem. 1991, 110, 429–435. [Google Scholar] [CrossRef]
- Rota, P.; Cirillo, F.; Piccoli, M.; Gregorio, A.; Tettamanti, G.; Allevi, P.; Anastasia, L. Synthesis and biological evaluation of several dephosphonated analogues of CMP-Neu5Ac as inhibitors of GM3-synthase. Chem. Eur. J. 2015, 21, 14614–14629. [Google Scholar] [CrossRef]
- Derevitskaya, V.A.; Kalinevich, V.M.; Kochetkov, N.K. Lactone of N-acetylneuraminic acid. Dokl. Akad. Nauk SSSR 1966, 169, 1087–1090. [Google Scholar]
- Colombo, R.; Anastasia, M.; Rota, P.; Allevi, P. The first synthesis of N-acetylneuraminic acid 1,7-lactone. Chem. Commun. 2008, 43, 5517–5519. [Google Scholar] [CrossRef]
- Allevi, P.; Anastasia, M.; Costa, M.L.; Rota, P. Two procedures for the syntheses of labeled sialic acids and their 1,7-lactones. Tetrahedron Asymmetry 2011, 22, 338–344. [Google Scholar] [CrossRef]
- Allevi, P.; Rota, P.; Scaringi, R.; Colombo, R.; Anastasia, M. Chemoselective synthesis of sialic acid 1,7-lactones. J. Org. Chem. 2010, 75, 5542–5548. [Google Scholar] [CrossRef] [PubMed]
- Colombo, R.; Anastasia, M.; Rota, P.; Allevi, P. Correction: The first synthesis of N-acetylneuraminic acid 1,7-lactone. Chem. Commun. 2015, 51, 4719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allevi, P.; Rota, P.; Scaringi, R.; Colombo, R.; Anastasia, M. Correction to “chemoselective synthesis of sialic acid 1,7-lactones”. J. Org. Chem. 2015, 80, 3357. [Google Scholar] [CrossRef] [PubMed]
- Mitsuoka, C.; Ohmori, K.; Kimura, N.; Kanamori, A.; Komba, S.; Ishida, H.; Kiso, M.; Kannagi, R. Regulation of selectin binding activity by cyclization of sialic acid moiety of carbohydrate ligands on human leukocytes. Proc. Natl. Acad. Sci. USA 1999, 96, 1597–1602. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, H.; Ando, H.; Ishida, H.; Kiso, M.; Ishihara, H.; Koketsu, M. Synthetic study on α(2→8)-linked oligosialic acid employing 1,5-lactamization as a key step. Tetrahedron Lett. 2009, 50, 4478–4481. [Google Scholar] [CrossRef]
- Hamada, T.; Hirota, H.; Yokoyama, S.; Otsubo, N.; Ishida, H.; Kiso, M.; Kanamori, A.; Kannagi, R. NMR analysis of novel ganglioside GM4 analogues containing de-N-acetyl and lactamized sialic acid: Probes for searching new ligand structures for human L-selectin. Magn. Reson. Chem. 2002, 40, 517–523. [Google Scholar] [CrossRef]
- Ando, H.; Koike, Y.; Koizumi, S.; Ishida, H.; Kiso, M. 1,5-lactamized sialyl acceptors for various disialoside syntheses: Novel method for the synthesis of glycan portions of Hp-s6 and HLG-2 gangliosides. Angew. Chem. Int. Ed. Engl. 2005, 44, 6759–6763. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.F.; Maliakel, B.P.; Zbiral, E. Formation of a lactam derivative on direct benzylation of N-acetylneuraminic acid alpha-methyl ketoside. Synlett 1992, 7, 561–562. [Google Scholar] [CrossRef]
- Meindl, P.; Bodo, G.; Palese, P.; Schulman, J.; Tuppy, H. Inhibition of neuraminidase activity by derivatives of 2-deoxy-2,3-dehydro-N-acetylneuraminic acid. Virology 1974, 58, 457–463. [Google Scholar] [CrossRef]
- Meindl, P.; Tuppy, H. 2-deoxy-2.3-dehydro-Sialic Acids. I. synthesis and properties of 2-deoxy-2.3-dehydro-N-acyl-neuraminic acids and their methyl esters. Mon. Chem. 1969, 100, 1295–1306. [Google Scholar] [CrossRef]
- Rota, P.; La Rocca, P.; Piccoli, M.; Montefiori, M.; Cirillo, F.; Olsen, L.; Orioli, M.; Allevi, P.; Anastasia, L. Potent inhibitors against newcastle disease virus hemagglutinin-neuraminidase. ChemMedChem 2018, 13, 236–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rota, P.; La Rocca, P.; Cirillo, F.; Piccoli, M.; Allevi, P.; Anastasia, L. The acidic hydrolysis of N-acetylneuraminic 4,5-oxazoline allows a direct functionalization of the C5 position of Neu5Ac2en (DANA). RSC Adv. 2020, 10, 162–165. [Google Scholar] [CrossRef] [Green Version]
- Rota, P.; Papini, N.; La Rocca, P.; Montefiori, M.; Cirillo, F.; Piccoli, M.; Scurati, R.; Olsen, L.; Allevi, P.; Anastasia, L. Synthesis and chemical characterization of several perfluorinated sialic acid glycals and evaluation of their in vitro antiviral activity against Newcastle disease virus. MedChemComm 2017, 8, 1505–1513. [Google Scholar] [CrossRef]
- Allevi, P.; Rota, P.; Agnolin, I.S.; Gregorio, A.; Anastasia, M. A Simple synthetic access to differently 4-substituted Neu5Ac2en glycals combining elements of molecules with anti-neuraminidase activity. Eur. J. Org. Chem. 2013, 2013, 4065–4077. [Google Scholar] [CrossRef]
- Rota, P.; Allevi, P.; Agnolin, I.S.; Mattina, R.; Papini, N.; Anastasia, M. A simple synthesis of N-perfluoroacylated and N-acylated glycals of neuraminic acid with a cyclic aminic substituent at the 4alpha position as possible inhibitors of sialidases. Org. Biomol. Chem. 2012, 10, 2885–2894. [Google Scholar] [CrossRef] [PubMed]
- Agnolin, I.S.; Rota, P.; Allevi, P.; Gregorio, A.; Anastasia, M. Simple and rapid procedures for the synthesis of 5-acylated 4β-acylamido- and 4β-acetoxyneuraminic acid glycals. Eur. J. Org. Chem. 2012, 2012, 6537–6547. [Google Scholar] [CrossRef]
- Rota, P.; Agnolin, I.S.; Allevi, P.; Anastasia, M. Facile diastereoselective entry to 4β-acylamidation of Neu5Ac2en glycals using their N-perfluoroacylated congeners as key tools. Eur. J. Org. Chem. 2012, 2012, 2508–2510. [Google Scholar] [CrossRef]
- Cheng, M.-C.; Lin, C.-H.; Lin, H.-J.; Yu, Y.-P.; Wu, S.-H. Hydrolysis, lactonization, and identification of α(2 → 8)/α(2 → 9) alternatively linked tri-, tetra-, and polysialic acids. Glycobiology 2004, 14, 147–155. [Google Scholar] [CrossRef]
- Lifely, M.R.; Gilbert, A.S.; Moreno, C. Rate, mechanism, and immunochemical studies of lactonisation in serogroup B and C polysaccharides of Neisseria meningitidis. Carbohydr. Res. 1984, 134, 229–243. [Google Scholar] [CrossRef]
- Lifely, M.R.; Gilbert, A.S.; Moreno, C. Sialic acid polysaccharide antigens of Neisseria meningitidis and Escherichia coli: Esterification between adjacent residues. Carbohydr. Res. 1981, 94, 193–203. [Google Scholar] [CrossRef]
- Ando, S.; Yu, R.K.; Scarsdale, J.N.; Kusunoki, S.; Prestegard, J.H. High resolution proton NMR studies of gangliosides. Structure of two types of GD3 lactones and their reactivity with monoclonal antibody R24. J. Biol. Chem. 1989, 264, 3478–3483. [Google Scholar]
- Kielczynski, W.; Bartholomeusz, R.K.; Harrison, L. Characterization of ganglioside associated with the thyrotrophin receptor. Glycobiology 1994, 4, 791–796. [Google Scholar] [CrossRef]
- Fronza, G.; Kirschner, G.; Acquotti, D.; Sonnino, S. Synthesis, structure, and conformation of the dilactone derivative of Gd1b ganglioside. Carbohydr. Res. 1989, 195, 51–58. [Google Scholar] [CrossRef]
- Maggio, B.; Ariga, T.; Yu, R.K. Ganglioside GD3 lactones: Polar head group mediated control of the intermolecular organization. Biochemistry 1990, 29, 8729–8734. [Google Scholar] [CrossRef]
- Yu, R.K.; Koerner, T.A.W.; Ando, S.; Yohe, H.C.; Prestegard, J.H. High-resolution proton NMR studies of gangliosides. III. Elucidation of the structure of ganglioside GM3 lactone. J. Biochem. 1985, 98, 1367–1373. [Google Scholar] [CrossRef] [PubMed]
- Terabayashi, T.; Kawanishi, Y. Naturally occurring ganglioside lactones in Minke whale brain. Carbohydr. Res. 1998, 307, 281–290. [Google Scholar] [CrossRef]
- Terabayashi, T.; Ogawa, T.; Kawanishi, Y. Characterization of Ganglioside GM4 Lactones isolated from the whale brain. J. Biochem. 1990, 107, 868–871. [Google Scholar] [CrossRef]
- Sato, T.; Ishii, M.; Ohtake, F.; Nagata, K.; Terabayashi, T.; Kawanishi, Y.; Okahata, Y. Binding affinity of GM3 lactone for influenza virus. Glycoconj. J. 1999, 16, 223–227. [Google Scholar] [CrossRef]
- Cheng, M.-C.; Lin, C.-H.; Wang, H.-Y.; Lin, H.-R.; Wu, S.-H. Regioselective lactonization of tetrasialic acid. Angew. Chem. Int. Ed. Engl. 2000, 39, 772–776. [Google Scholar] [CrossRef]
- Yu, Y.-P.; Cheng, M.-C.; Lin, H.-R.; Lin, C.-H.; Wu, S.-H. Acid-catalyzed hydrolysis and lactonization of α2,8-linked oligosialic acids. J. Org. Chem. 2001, 66, 5248–5251. [Google Scholar] [CrossRef]
- Bratosin, D.; Palii, C.; Moicean, A.D.; Zanetta, J.-P.; Montreuil, J. Reduced diversity of the human erythrocyte membrane sialic acids in polycythemia vera. Absence of N-glycolylneuraminic acid and characterisation of N-acetylneuraminic acid 1,7 lactone. Biochimie 2007, 89, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Rinninger, A.; Richet, C.; Pons, A.; Kohla, G.; Schauer, R.; Bauer, H.C.; Zanetta, J.-P.; Vlasak, R. Localisation and distribution of O-acetylated N-acetylneuraminic acids, the endogenous substrates of the hemagglutinin-esterases of murine coronaviruses, in mouse tissue. Glycoconj. J. 2006, 23, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Zanetta, J.-P.; Srinivasan, V.; Schauer, R. Analysis of monosaccharides, fatty constituents and rare O-acetylated sialic acids from gonads of the starfish Asterias rubens. Biochimie 2006, 88, 171–178. [Google Scholar] [CrossRef]
- Bohin, A.; Bouchart, F.; Richet, C.; Kol, O.; Leroy, Y.; Timmerman, P.; Huet, G.; Bohin, J.-P.; Zanetta, J.-P. GC/MS identification and quantification of constituents of bacterial lipids and glycoconjugates obtained after methanolysis as heptafluorobutyrate derivatives. Anal. Biochem. 2005, 340, 231–244. [Google Scholar] [CrossRef]
- Pons, A.; Richet, C.; Robbe, C.; Herrmann, A.; Timmerman, P.; Huet, G.; Leroy, Y.; Carlstedt, I.; Capon, C.; Zanetta, J.-P. Sequential GC/MS Analysis of sialic acids, monosaccharides, and amino acids of glycoproteins on a single sample as heptafluorobutyrate derivatives. Biochemistry 2003, 42, 8342–8353. [Google Scholar] [CrossRef] [PubMed]
- Bulai, T.; Bratosin, D.; Pons, A.; Montreuil, J.; Zanetta, J.-P. Diversity of the human erythrocyte membrane sialic acids in relation with blood groups. FEBS Lett. 2003, 534, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Robbe, C.; Capon, C.; Maes, E.; Rousset, M.; Zweibaum, A.; Zanetta, J.-P.; Michalski, J.C. Evidence of regio-specific glycosylation in human intestinal mucins: Presence of an acidic gradient along the intestinal tract. J. Biol. Chem. 2003, 278, 46337–46348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cebo, C.; Vergoten, G.; Zanetta, J.-P. Lectin activities of cytokines: Functions and putative carbohydrate-recognition domains. Biochim. Biophys. Acta 2002, 1572, 422–434. [Google Scholar] [CrossRef]
- Pons, A.; Timmerman, P.; Leroy, Y.; Zanetta, J.-P. Gas-chromatography/mass-spectrometry analysis of human skin constituents as heptafluorobutyrate derivatives with special reference to long-chain bases. J. Lipid. Res. 2002, 43, 794–804. [Google Scholar]
- Zanetta, J.-P.; Cebo, C.; Vergoten, G. Lectin activities of cytokines: A new concept in immunology. Trends Glycosci. Glycotechnol. 2002, 14, 303–318. [Google Scholar] [CrossRef]
- Cebo, C.; Dambrouck, T.; Maes, E.; Laden, C.; Strecker, G.; Michalski, J.C.; Zanetta, J.-P. Recombinant human interleukins IL-1alpha, IL-1beta, IL-4, IL-6, and IL-7 show different and specific calcium-independent carbohydrate-binding properties. J. Biol. Chem. 2001, 276, 5685–5691. [Google Scholar] [CrossRef] [Green Version]
- Zanetta, J.-P.; Pons, A.; Iwersen, M.; Mariller, C.; Leroy, Y.; Timmerman, P.; Schauer, R. Diversity of sialic acids revealed using gas chromatography/mass spectrometry of heptafluorobutyrate derivatives. Glycobiology 2001, 11, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Pons, A.; Popa, J.; Portoukalian, J.; Bodennec, J.; Ardail, D.; Kol, O.; Martin-Martin, M.J.; Hueso, P.; Timmerman, P.; Leroy, Y.; et al. Single-step gas chromatography-mass spectrometry analysis of glycolipid constituents as heptafluorobutyrate derivatives with a special reference to the lipid portion. Anal. Biochem. 2000, 284, 201–216. [Google Scholar] [CrossRef]
- Zanetta, J.P.; Timmerman, P.; Leroy, Y. Gas-liquid chromatography of the heptafluorobutyrate derivatives of the O-methyl-glycosides on capillary columns: A method for the quantitative determination of the monosaccharide composition of glycoproteins and glycolipids. Glycobiology 1999, 9, 255–266. [Google Scholar] [CrossRef]
- Horne, C.R.; Kind, L.; Davies, J.S.; Dobson, R.C.J. On the structure and function of escherichia coli YjhC: An oxidoreductase involved in bacterial sialic acid metabolism. Proteins 2020, 88, 654–668. [Google Scholar] [CrossRef] [PubMed]
- Vimr, E.R. Unified theory of bacterial sialometabolism: How and why bacteria metabolize host sialic acids. ISRN Microbiol. 2013, 2013, 816713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popa, I.; Zanetta, J.P.; Portoukalian, J.; Thomas, L. GM3 ganglioside of human melanoma tumors contains a lactonized sialic that is a ligand of interleukin-4. J. Investig. Dermatol. 2002, 118, 903. [Google Scholar]
- Rota, P.; Anastasia, L.; Allevi, P. Elucidation of several neglected reactions in the GC-MS identification of sialic acids as heptafluorobutyrates calls for an urgent reassessment of previous claims. Org. Biomol. Chem. 2015, 13, 4931–4939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gervay, J.; Ramamoorthy, P.S.; Mamuya, N.N. Ring opening of sialyllactones with glycine esters: Synthesis of selectively protected glycinyl-NeuAc saccharopeptides. Tetrahedron 1997, 53, 11039–11048. [Google Scholar] [CrossRef]
- Gervay, J.; Mamuya, N.N.; Andrew Barber, R. Synthesis of 13C enriched sialyllactones and their characterization using isotope edited inverse detected NMR spectroscopy. Tetrahedron Lett. 1997, 38, 1865–1868. [Google Scholar] [CrossRef]
- Parrill, A.L.; Mamuya, N.; Dolata, D.P.; Gervay, J. Computational studies of sialyllactones: Methods and uses. Glycoconj. J. 1997, 14, 523–529. [Google Scholar] [CrossRef]
- Sato, S.; Furuhata, K.; Ogura, H. Studies on Sialic Acids.XIV. Lactone derivatives of N-acetylneuraminic acid. Chem. Pharm. Bull. 1988, 36, 4678–4688. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, N.; Sugai, K.E.I.; Yamada, N.; Goto, M.; Ban, C.; Furuhata, K.; Takayanagi, H.; Ogura, H. Studies on Sialic Acids. IX: Formation of a 1,7-lactone derivative by direct acetylation of N-acetylneuraminic acid. Chem. Pharm. Bull. 1988, 36, 1147–1152. [Google Scholar] [CrossRef] [Green Version]
- Kirchner, E.; Thiem, F.; Dernick, R.; Heukeshoven, J.; Thieina, J. Studies on the glycosylation of N-acetylneuraminic acid. J. Carbohydr. Chem. 1988, 7, 453–486. [Google Scholar] [CrossRef]
- Khorlin, A.Y.; Privalova, I.M. Acylation of N-acetylneuraminic acid. Chem. Nat. Compd. 1967, 3, 159–162. [Google Scholar] [CrossRef]
- Horn, E.J.; Saludes, J.P.; Gervay-Hague, J. Investigation into an efficient synthesis of 2,3-dehydro-N-acetyl neuraminic acid leads to three decarboxylated sialic acid dimers. Carbohydr. Res. 2008, 343, 936–940. [Google Scholar] [CrossRef]
- Tsvetkov, Y.E.; Schmidt, R.R. 8-O-Sialylation of derivatives of neuraminic acid 1,7-lactone unusual stereoselectivity. Tetrahedron Lett. 1994, 35, 8583–8586. [Google Scholar] [CrossRef]
- La Rocca, P.; Rota, P.; Piccoli, M.; Cirillo, F.; Orioli, M.; Ravelli, A.; Allevi, P.; Anastasia, L. Lactonization method to assign the anomeric configuration of the 3,4-unsaturated congeners of N-acetylneuraminic acid. J. Org. Chem. 2019, 84, 5460–5470. [Google Scholar] [CrossRef]
- Compound 4a (N-Acetyl-beta-neuraminic Acid 1,7-Lactone CAS: 335592-15-9) Is Commercially Available from Atomax Chemicals. Available online: http://en.atomaxchem.com (accessed on 29 October 2020).
- Rota, P.; Allevi, P.; Anastasia, L. The sialic acids waltz: Novel stereoselective isomerization of the 1,7-lactones of N-acetylneuraminic acids into the corresponding γ-lactones and back to the free sialic acids. Asian J. Org. Chem. 2015, 4, 1315–1321. [Google Scholar] [CrossRef]
- Nakamura, M.; Takayanagi, H.; Furuhata, K.; Ogura, H. synthesis and characterization of furanose and pyranose derivatives of 3-deoxy-D-glycero-D-galacto-2-nonulosonic acid (KDN). Chem. Pharm. Bull. 1992, 40, 879–885. [Google Scholar] [CrossRef] [Green Version]
- David, S.; Malleron, A.; Cavayé, B. Some derivatives of 3-deoxy-D-glycero-D-galacto-non-2-ulosonic acid (KDN). Carbohydr. Res. 1994, 260, 233–241. [Google Scholar] [CrossRef]
- Kuhn, R.; Baschang, G. Die konfiguration der sialinsäuren am C-atom 4. Chem. Ber. 1962, 95, 2384–2385. [Google Scholar] [CrossRef]
- Kuhn, R.; Brossmer, R. The configuration of sialic acids. Angew. Chem. Int. Ed. 1962, 1, 218. [Google Scholar] [CrossRef]
- Kuhn, R.; Baschang, G. Aminozucker-Synthesen, XXV. Synthese der lactaminsäure. Justus Liebigs Ann. Chem. 1962, 659, 156–163. [Google Scholar] [CrossRef]
- Faillard, H.; Blohm, M. Synthesis of N-glycolyneuraminic acid. Hoppe Seyler Z. Physiol. Chem. 1965, 341, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Sawada, Y.; Nakatsuka, S. Synthesis of gamma-lactone-type sialic acid, an isomer of 2,3-dehydrosialic acid. Biosci. Biotechnol. Biochem. 2010, 74, 1106–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khorlin, A.Y.; Privalova, I.M. Synthesis of N-acetylneuraminic acid 8-methyl ether. Carbohydr. Res. 1970, 13, 373–377. [Google Scholar] [CrossRef]
- Holmquist, L. 7,9-O-ethylidene and 7,9-O-benzylidene derivatives of 5-acetamido-3,5-dideoxy-D-glycero-D-galacto-nonulosonic acid (N-acetylneuraminic acid). Acta Chem. Scand. 1971, 25, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Zbiral, E.; Schmid, W. Structural modifications on N-acetylneuraminic Acid. 4. Transformations on the diethyldithioketal of N-acetylneuraminic acid-gamma-lactone. Mon. Chem. 1985, 116, 253–262. [Google Scholar] [CrossRef]
- Furuhata, K.; Sato, S.; Anazawa, K.; Goto, M.; Takayanagi, H.; Ogura, H. Studies on sialic acids. VIII. The 1,4-lactone derivatives of N-acetylneuraminic acid. Chem. Pharm. Bull. 1987, 35, 3609–3614. [Google Scholar] [CrossRef] [Green Version]
- Warwel, M.; Fessner, W.-D. Indium-mediated chain-extension: An improved protocol for the concise, diastereospecific synthesis of KDN and other sialic acids. Synlett 2000, 2000, 865–867. [Google Scholar] [CrossRef]
- Rota, P.; Allevi, P.; Costa, M.L.; Anastasia, M. High yielding N-transacylation of secondary amides in acids labile molecules by the action of perfluorinated anhydrides in the presence of a mild base. Tetrahedron Asymmetry 2010, 21, 2681–2686. [Google Scholar] [CrossRef]
- Rota, P.; Allevi, P.; Mattina, R.; Anastasia, M. Reaction of N-acetylneuraminic acid derivatives with perfluorinated anhydrides: A short access to N-perfluoracylated glycals with antiviral properties. Org. Biomol. Chem. 2010, 8, 3771–3776. [Google Scholar] [CrossRef]
- Monticelli, E.; Aman, C.S.; Costa, M.L.; Rota, P.; Bogdan, D.; Allevi, P.; Cighetti, G. Simultaneous free and glycosylated pyridinium crosslink determination in urine: Validation of an HPLC-fluorescence method using a deoxypyridinoline homologue as internal standard. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2011, 879, 2764–2771. [Google Scholar] [CrossRef] [PubMed]
- Rota, P.; Allevi, P.; Colombo, R.; Costa, M.L.; Anastasia, M. General and chemoselective N-transacylation of secondary amides by means of perfluorinated anhydrides. Angew. Chem. Int. Ed. 2010, 49, 1850–1853. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | Reagent | T (°C) | Step A | Step B | ||
|---|---|---|---|---|---|---|
| t (h) | Yield (%) of 33 | t (h) | Yield (%) of 1a | |||
| entry 1 | Dowex H+ a | 25 | 1 | 86 | 120 | 87 |
| entry 2 | Dowex H+ a | 80 | - | - | 8 | 81 |
| entry 3 | 1M HCl a | 80 | - | - | 1 | 65 |
| entry 4 | 2M AcOH a | 25 | 24 | 83 | - | - |
| entry 5 | 2M AcOH a | 80 | 0.25 | 78 | 3 | 50 |
| entry 6 | 0.5M NaOMe b | 30 | 0.5 | 92 | - | - |
| entry 7 | 0.05M TMG b | 25 | 0.15 | 91 | 72 | 78 |
| entry 8 | 0.05M TMG b | 60 | - | - | 3 | 97 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rota, P.; La Rocca, P.; Allevi, P.; Pappone, C.; Anastasia, L. Intramolecular Lactones of Sialic Acids. Int. J. Mol. Sci. 2020, 21, 8098. https://doi.org/10.3390/ijms21218098
Rota P, La Rocca P, Allevi P, Pappone C, Anastasia L. Intramolecular Lactones of Sialic Acids. International Journal of Molecular Sciences. 2020; 21(21):8098. https://doi.org/10.3390/ijms21218098
Chicago/Turabian StyleRota, Paola, Paolo La Rocca, Pietro Allevi, Carlo Pappone, and Luigi Anastasia. 2020. "Intramolecular Lactones of Sialic Acids" International Journal of Molecular Sciences 21, no. 21: 8098. https://doi.org/10.3390/ijms21218098
APA StyleRota, P., La Rocca, P., Allevi, P., Pappone, C., & Anastasia, L. (2020). Intramolecular Lactones of Sialic Acids. International Journal of Molecular Sciences, 21(21), 8098. https://doi.org/10.3390/ijms21218098