Survival of Lung Cancer Patients Dependent on the LOH Status for DMP1, ARF, and p53
by
 ,
,
Elizabeth A. Fry
1, 
Gloria E. Niehans
2,
Robert A. Kratzke
3,
Fumitake Kai
1 and
Kazushi Inoue
1,* 1
Dept. of Pathology, Wake Forest University School of Medicine, Medical Center Blvd., Winston-Salem, NC 27157, USA
2
Minneapolis VA Medical Center, Minneapolis, MN 55417, USA
3
Dept. of Medicine, University of Minnesota Medical Center, Masonic Cancer Institute, Minneapolis, MN 55455, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(21), 7971; https://doi.org/10.3390/ijms21217971
Submission received: 7 September 2020
/
Revised: 13 October 2020
/
Accepted: 19 October 2020
/
Published: 27 October 2020
(This article belongs to the Special Issue Cancer Prevention with Molecular Target Therapies)
Abstract
:Lung cancer is the leading cause of cancer deaths in the world, and accounts for more solid tumor deaths than any other carcinomas. The prognostic values of DMP1, ARF, and p53-loss are unknown in lung cancer. We have conducted survival analyses of non-small cell lung cancer (NSCLC) patients from the University of Minnesota VA hospital and those from the Wake Forest University Hospital. Loss of Heterozygosity (LOH) for hDMP1 was found in 26 of 70 cases (37.1%), that of the ARF/INK4a locus was found in 33 of 70 (47.1%), and that of the p53 locus in 43 cases (61.4%) in the University of Minnesota samples. LOH for hDMP1 was associated with favorable prognosis while that of p53 predicted worse prognosis. The survival was much shorter for ARF-loss than INK4a-loss, emphasizing the importance of ARF in human NSCLC. The adverse effect of p53 LOH on NSCLC patients’ survival was neutralized by simultaneous loss of the hDMP1 locus in NSCLC and breast cancer, suggesting the possible therapy of epithelial cancers with metastatic ability.
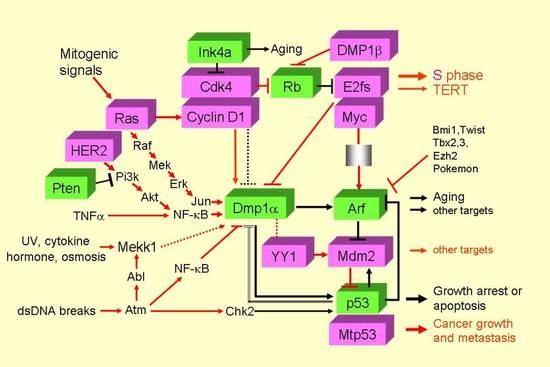
1. Introduction
Lung cancer is the leading cause of cancer deaths in the world, and accounts for more solid tumor deaths than any other carcinomas. More than 22,800 new cases are diagnosed each year in the United States alone, of which 135,700 will die 2020, representing 25% of all cancer deaths [1]. Lung cancer can be categorized into two major histopathological groups: Non small-cell lung cancer (NSCLC) and small-cell lung cancer (SCLC) [2], the latter of which shows neuroendocrine features. A total of 80%–85% of lung cancers are NSCLC [2], and they are subcategorized into adenocarcinomas (AC), squamous cell carcinomas (SCC), adenosquamous carcinomas, and large-cell carcinomas [3]. SCLC and NSCLC show major differences in histopathologic characteristics that can be explained by the distinct patterns of genetic alterations found in both tumor classes [4].
The ARF gene was cloned as the alternate reading frame gene for p16Ink4a from the mouse ARF/Ink4a locus [5,6,7,8,9,10]. Accumulating studies showed that the ARF tumor suppressor is a sensor for hyperproliferative oncogenic stimuli stemming from mutant Ras, c-Myc, E2F-1, and HER2 proteins [11,12,13,14], virtually induced by all the oncogenes stimulating p53 to induce cell cycle arrest, apoptosis, and autophagy [15]. p19ARF (p14ARF in humans) and p16Ink4a mRNAs are generated from separate and first exons 1β and 1α (19.4 kilo base pairs (kbp) apart in humans; 12.4 kbp apart in mice) which splice into two common exons 2 and 3 [5]. These two genes are different since p19ARF uses only exons 1 and 2 (also p14ARF) while p16INK4a uses all of the exons 1–3 for production of the protein [5,16]. This ARF-INK4a (CDKN2a) locus is located 11.5 kbp downstream of the genomic locus for CDKN2b that encodes for p15INK4 [9]. All of p15Ink4b, p19ARF, and p16Ink4a are tumor suppressor genes as proved by analyses of gene knockout mice [17,18,19,20]. Since RB is regulated by p16INK4a and p53 is regulated by p14ARF, the ARF/INK4a locus is very frequently inactivated in human cancers, second only to p53 [21,22]. ARF is a highly basic, insoluble protein (pI 11; [23]). Although human and mouse ARF differ in size (mouse 19 kDa, human 14 kDa) and show only 49% identity in amino acid sequences, the functions of the ARF proteins are well-conserved between the two species [6]. ARF is regulated at both transcriptional and protein levels by ULF, MKRN1, and SIVA1 [10].
Ectopic ARF arrests immortal rodent cell lines, such as NIH 3T3, as well as human cells from cancer [5,16,24]. The ability of ARF to inhibit cell cycle progression in a number of cell types suggested that ARF has powerful growth-inhibitory functions in cells, which stimulated researchers to study the in vivo activity of ARF to prevent tumors. ARF sequesters MDM2 in the nucleolus, thus preventing p53 degradation [25]. In addition, it inhibits the transcription factor E2F activity [26]. These activities lead to cell cycle arrest at G1 and G2 [16]. Itahana et al. studied the role of ARF in apoptosis and found that the mitochondrial protein p32/C1QBP bound to the ARF C-terminus, where p32 is required for ARF to localize to mitochondria to induce apoptosis, demonstrating the essential role of ARF in tumor suppression and programmed cell death [27]. Recent studies indicate that nuclear factor E2-related factor 2 is a major target of ARF in p53-independent tumor suppression [28].
The gene for the Dmp1 (cyclin D binding myb-like protein 1; Dmtf1) transcription factor was isolated in yeast two-hybrid screen of CTLL2 cell library with cyclin D2 bait [29]. Inoue and Sherr reported that gene expression and cell cycle arrest mediated by Dmp1 (Dmp1α) was antagonized by D-type cyclins through a Cdk-independent mechanism [30]. Importantly, Dmp1 directly binds to the Ets consensus sequence 5′ CCCGGATGC-3′ of the ARF promoter to activate its gene expression, thereby inducing p53-dependent cell cycle arrest ([29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47] for primary articles; [48,49,50,51] for reviews). The human DMP1 locus encodes DMP1α, and its splice variants DMP1β and γ [46]. DMP1α regulates the human ARF promoter, the activity of which is antagonized its splice variant DMP1β [46]. Similar mechanisms must be present in human ARF promoter since both E2F and DMP1-consensus sequences are found in the human version as well. Dmp1-deficient mice were prone to spontaneous tumor development, which was accelerated when the animals were neonatally treated with ionizing radiation or dimethylbenzanthracene [34,35]. Although Dmp1-deficient mice develop a broad spectrum of epithelial and non-epithelial tumors, lung tumors were the most frequently encountered neoplasms in both Dmp1-null and Dmp1-hetrozygous mice [34,35]. The wild type Dmp1 allele was retained and expressed in tumors arising from Dmp1+/- mice, demonstrating a haplo-insufficiency of Dmp1 in tumor suppression [35,51]. Tumors from Dmp1-/- or Dmp1+/- (Eμ-Myc, K-RasLA, HER2 mutant) mice rarely showed mutations, deletions, or silencing of p19ARF or p53, suggesting that Dmp1 is a critical regulator of the ARF-p53 tumor suppressor pathway in vivo [14,34,35,39,48,49,50,51].
Activation of the Dmp1 promoter by oncogenic Ras or mutant HER2 have been reported [11,14,36], indicating that it is a critical mediator in RAS or HER2 induced ARF, p53 cell cycle arrest to prevent incipient cancer cells. The Dmp1 promoter was also activated by an inflammatory cytokine TNFα mediated by NF-κB [38] as well as dsDNA breaks [41], indicating that Dmp1 is a mediator of a variety of stress signaling. We conducted GeneChip microarray using Dmp1+/+ and Dmp1-/- lungs and found that other transcriptional targets for Dmp1α include Areg, Thbs1, JunB, and Egr1 [40], suggesting that it is involved in signal transduction pathways involving cell proliferation, angiogenesis, and invasion/metastasis.
Dmp1 shows it tumor suppressive activity not only transactivating the ARF promoter in response to oncoprotein overexpression, but also through physical interaction with p53 in response to DNA damage response [41]. Our data indicate that acceleration of DNA-binding of p53 by Dmp1 is a critical process for Dmp1 to increase the p53 function in ARF-deficient cells [47].
Whether hDMP1 is involved in the pathogenesis in human cancer is a critical issue for research. We found that loss of heterozygosity (LOH) of hDMP1 was present in ~40% of non-small cell lung carcinomas (NSCLC), especially those that retain wild type INK4a/ARF and/or p53 [39]. In this study, we received specimen from the University Minnesota VA Hospital (UM) to study the survival of LOH for hDMP1, ARF/INK4a, p53; the impacts for overexpression of cyclin D1 and YY1 have also been studied on NSCLC survival, both progression-free survival (PFS) and total survival (TS).
The goals of this study are to clarify prognostic impact form the hDMP1 LOH, ARF/INK4a LOH, p53 LOH and immunohistochemistry (IHC), Cyclin D1 (IHC), and YY1 (genomic DNA amplification).
2. Results
2.1. Impacts of LOH for hDMP1, ARF/INK4a, and p53 on NSCLC Survival
Previous publication from WFU with samples of 51 NSCLC [39] showed that LOH of the hDMP1 gene was found in 33.3% of samples with 5′ primer, 36.1% with 3′ primer (average 35%), but signs of biallelic involvement (promoter hypermethylation or complete loss) was extremely rare (the former 2.2% and 0% for the latter) in human lung cancer indicating that hDMP1 was haploinsufficient for tumor suppression [35,51]. Now, we have studied different set of samples (n = 70) with survival data (PFS and TS) from the University of Minnesota VA hospital (UM) [48,49].
With the 5′ set of hDMP1 primers (#92465), 10 of 70 cases (14.3%) were positive for LOH; with the 3′ set of hDMP1 primers (#198004, #176671), 24 of 70 cases were positive (34.3%) (average 24.3%). 26 of 70 cases (37.1%) were positive for either of these (Table 1). The number was close to those published in WFU samples (35%; 39). Eight of 70 cases (11.4%) were positive for LOH with both of these hDMP1 primer sets (Table 1), suggesting that gene deletion extends the entire DMP1 locus in these samples. LOH for hDMP1 was associated with longer survival in both PFS (p = 0.0029, χ2 = 8.8993) and total survival (p = 0.0040, χ2 = 8.3026) in the UM samples (Figure 1A,B). Consistently, LOH for hDMP1 was associated with favorable survival in TS in our previous WFU samples analyzed (p = 0.0324, χ2 = 4.5230, n = 42, Supplementary Figure S1A) [39]. The tendency was more prominent is squamous cell carcinoma than adenocarcinoma (Supplementary Figure S2).
Then the same NSCLC samples pairs were studied for LOH of ARF/INK4a and p53. With ARF/INK4a primers, 23 of 70 cases (32.9%) were positive for LOH 5′ primers close to the ARF locus, 19 of 70 cases (27.1%) were positive for LOH 3′ primers close to the INK4a locus (average 30%), and 33 of 70 showed LOH for either one of these (47.1%). Nine of 70 cases (12.9%) showed LOH with both sets of the ARF/INK4a primers (Table 1). Importantly, LOH for the ARF/INK4a locus was not associated with survival PFS (p = 0.1718, χ2= 1.8670) or TFS (p = 0.1721, χ2= 1.8649) although there was a trend that it was associated with worse prognosis (Figure 1C,D). The trend was the same when AC and SCC were separately analyzed (Supplementary Figure S3). The LOH for the ARF/INK4a locus was not associated with TS in previous WFU specimens analyzed (p = 0.7707, χ2 = 0.08495, n = 43; Supplementary Figure S1B). Forty-seven of 53 cases (88.7%) showed mutually exclusive loss of the hDMP1 and the ARF-INK4a loci (p = 0.0023; χ2 = 9.330; 95% confidence interval, 80.1%–97.2%; Table 1). We found that LOH for ARF was much worse prognostic factor than that of INK4a (Figure 1E,F) in both PFS and TS; as a matter of fact none of the NSCLC relapsed and died within 1800 (PFS) and 3000 days (TS) of observation for INK4a LOH, whereas NSCLC with ARF deletion relapsed and died in our survival analyses (Figure 1E,F; p = 0.0430 for PFS and p = 0.0448 for TS), suggesting that ARF LOH has much stronger impact on NSCLC survival than that of INK4a. When the survival for LOH for both of ARF and INK4a was studied, it was shorter in patients with the locus involvement than those without although data were not statistically significant (Supplementary Figure S4).
LOH of p53 (incl. biallelic gene deletion) was found in 37.1% (26/70) with 5′ primers, 47.1% (33/70) with 3′ primers (average 42.1%), and 61.4% (43/70) with either primers in UM NSCLC samples (Table 1). LOH of p53 was associated with worse prognosis in both PFS (p = 0.0472, χ2 = 3.9372) and TS (p = 0.0302, χ2 = 4.6963) in the UM samples (Supplementary Figure S5A,B). Consistently, LOH for p53 had the trend for worse prognosis in WFU NSCLC samples (p = 0.1079, χ2 = 2.5849, n = 42; Supplementary Figure S1C). Forty-eight of 58 cases (82.8%) showed mutually exclusive loss of the hDMP1 and the p53 loci (p = 0.0128; χ2 = 6.195; 95% confidence interval, 75.6–93.9%; Table 1). The p values did not become smaller when AC and SCC were analyzed separately since N became smaller (Supplementary Figure S6).
We also conducted IHC analysis for p53 to study the impact of p53 expression on patients’ survival (Supplementary Figure S5C,D). High expression of p53 protein (level 2, intense staining) was associated with shorter survival in PFS (p = 0.0340, χ2 = 4.4944, n = 62) and TS (p = 0.0537, χ2 = 3.7270) suggesting that it is an indicator worse prognosis (Supplementary Figure S5C,D). The p vales were relative large because there were significant number of samples where p53 IHC was not performed, and because there were samples that showed weak p53 IHC (grade 1; these cases were excluded from the study). Like the relation of LOH for hDMP1 and p53 loci, LOH of hDMP1 and overexpression of the p53 protein did not to overlap each other (29/33 = 87.9% mutually exclusive; 95% confidence interval: 76.7%–99.0%; Table 1).
2.2. The Impact of hDMP1 LOH on NSCLC Survival with p53 LOH
The fact that LOH for hDMP1 was mutually exclusive for that of p53 in 82.8% in NSCLC samples (48/58, Table 1) means that LOH for hDMP1 was overlapping with that of p53 in 17.2% of cases (10/58, 12 cases showed no LOH for DMP1 and p53). We, therefore, studied the impact of hDMP1 LOH in p53 LOH(+) UM samples (Figure 2). We found that hDMP1 LOH dramatically improved the survival (both PFS and TS) of p53 LOH(+) in NSCLC patients’ survival in NSCLC (Figure 2A,B). As a matter of fact, the survival of p53 LOH(+); hDMP1 LOH(+) was comparable to or even better than that of p53 LOH(-) in UM samples (p = 0.0013, χ2 = 13.2841 in PFS; p = 0.0012, χ2 = 13.5258 in TS, triple survival assay), indicating that one locus hDMP1 deletion neutralized the negative effect of p53 LOH in NSCLC (3000 day survival of 12.0% became 87.5% in PFS (7.3 fold improvement), Figure 2A; 3000 day survival of 19.8% became 87.5% in TS (4.2 fold), Figure 2B). The same phenomenon was also observed in previous WFU NSCLC specimens as analyzed for the total survival (Supplementary Figure S7A, p = 0.0714, χ2 = 5.2800, n = 42). Indeed, no SCLC patient with double LOH for p53 and hDMP1 relapsed in the observation period of 1900 days. The p value was relatively large because two Wake Forest University Health Sciences (WFUHS) NSCLC samples with double hDMP1; p53 LOH (1990-10, 2005-308 in ref. [39]) did not have any survival data. The same trend was also found in breast cancer samples in WFUHS [42] since none of the 10 patients that showed dual LOH for p53 and hDMP1 relapsed during the 2100 days of observation period (p = 0.0189, χ2 = 7.9382, n = 105) (Figure 2C). Conversely, the LOH for the ARF/INK4a locus did not improve the negative effect of p53 LOH in NSCLC survival (Supplementary Figure S7B,C). These results show that the adverse prognostic impact of p53 LOH in epithelial tumors (i.e., NSCLC and breast carcinoma) is greatly improved with simultaneous loss of the hDMP1, the result of which are consistent with favorable impact of hDMP1 LOH in NSCLC (this study) and breast cancer [42].
2.3. Cyclin D1 Overexpression and YY1 Amplification and NSCLC Survival
Finally, we examined the impact of Cyclin D1 [52,53] overexpression at protein levels and YY1 [54,55] expression at genomic DNA level. Overexpression of the Cyclin D1 protein (grade 2 in IHC) was found in 26 of 52 cases while 26 samples had no expression of the protein in NSCLC samples. Samples with Cyclin D1 overexpression had the trend to be associated with favorable prognosis in PFS and TS, but the data were not statistically significant (p = 0.0875, χ2 = 2.9191 in PFS; p = 0.0754, χ2 = 3.1603 in TS) (Figure 3A,B). Although both hDMP1 LOH and Cyclin D1 overexpression were associated with favorable prognosis, the Cyclin D1 protein overexpression was not related to the LOH of hDMP1 (p > 0.20) suggesting that these two events happened independently. When we conducted quadruple analysis of NSCLC for Cyclin D1 and hDMP1 LOH, we found significantly shorter survival for the group without Cyclin D1 expression without LOH for hDMP1 (p = 0.0031, χ2 = 13.8559 for PFS; p = 0.0020, χ2 = 14.8269 for TS; Figure 3C,D). As a matter of fact, none of the patients lived more than 1400 days in this group where clinicians are alerted for early relapse.
The MDM2 stimulator [54] and epigenetic modifier [55]. YY1 gene amplification (>3 folds) was found in 16 of 68 NSCLC samples (23.5%) examined. The gene amplification seemed to predict favorable outcome of patients, but neither of them (PFS: p = 0.2833, χ2 = 1.1590; TS: p = 0.0973, χ2 = 2.7490) was statistically significant (Figure 4A,B). Amplification of YY1 genomic locus was independent of LOH for hDMP1 (p > 0.50). Then we conducted quadruple analysis of NSCLC for YY1 amplification and hDMP1 LOH. We found shorter survival for the group without YY1 amplification expression without LOH for hDMP1 (p = 0.0109, χ2 = 11.1565 for PFS, p = 0.0024, χ2 = 14.4367 for TS; Figure 4C,D), indicating that YY1 amplification (-); hDMP1 LOH (-) genotype is an ominous sign of NSCLC.
In summary, both Cyclin D1 protein overexpression and YY1 genomic amplification seem to predict favorable outcome of NSCLC patients; these do not overlap with LOH for hDMP1 which is another favorable prognostic factor. When these are absent without LOH for hDMP1, NSCLC patients will not live long.
3. Discussion
We have analyzed LOH values for hDMP1, ARF/INK4a, and p53 loci from NSCLC specimen obtained from a different institution. The percentage is a bit different—37.1% of hDMP1 locus, 47.1% for ARF/INK4a, and 61.4% for p53 in the UM VA hospital samples collected in 1991–2000 while it was 40.8%, 36.0%, and 47.9% from the WFU hospital, 1999–2006, because of the difference of demographic distribution. Importantly, the hDMP1 locus had positive impact on NSCLC survival in both institutions (p = 0.0029 in PFS, p = 0.0040 in TS in UM samples; p = 0.03551 in WFU sample) [39]. LOH for the ARF/INK4a locus did not have prognostic values in both institutions (p = 0.1718 in PFS, p = 0.1721 in TS in UM specimens), although there was a trend that LOH for ARF/INK4a locus with either of the two primers was associated with worse prognosis in UM samples. Current data show that LOH for the ARF/INK4a locus is mutually exclusive for that for the hDMP1 (88.7%) in NSCLC. Published studies show that Dmp1 activates both ARF-p53 and Ink4a-Rb pathways in mice for tumor suppression [43]. These human and mouse data are very consistent indicating that DMP1 is in the upstream of these pathways, both having binding sites for transactivation [43].
Since ARF/INK4a locus 5′ probe #33647 was close to ARF exon 1β and #27251 was close to INK4a exon 1α, LOH was evaluated separately in this study. Of note, we noticed that #33647 LOH (representing the ARF locus; LOH, 23/70, 32.9%) had much worse impact on NSCLC survival than that of #27251 (representing the INK4a locus; 19/70, LOH 27.1%). This is quite unexpected because INK4a has been considered to be much more important tumor suppressor than ARF in human cancer [21,22]. This is possibly because the ARF gene is inactivated in human cancer by gene deletion, splicing alteration than INK4a, which is inactivated mainly by promoter methylation or coding exon point mutations [5,16]; for splicing errors for ARF, see ref. [2]). Frameshift caused by nucleotide insertion or deletion affects both of these genes at equal frequency [16]. Since the frequency of LOH is similar for ARF and INK4a, it is highly possible that ARF plays an important role as INK4a in suppressing tumor development in NSCLC. Importantly, LOH for ARF/INK4a was mutually exclusive of that for the hDMP1 locus in UM samples (this study) as well as WFU samples [39,42] confirming that DMP1 is in the same pathway as that of ARF-p53 signaling.
In good contrast to LOH for hDMP1, LOH for p53 had negative impact on patients’ survival (Supplementary Figure S5) in both PFS and TS in UM NSCLC samples. Likewise, IHC study shows that p53 protein overexpression is associated with worse prognosis in lung cancer. Again IHC staining for p53 was mutually exclusive for that of hDMP1 in UM specimen for NSCLC (29/33). The data are consistent our findings that Dmp1 binds directly to the p53 protein, esp. when cells receive DNA damage [41,47]. IHC for p53 (grade 2) tend not to overlap LOH for hDMP1 in our study (87.9% exclusive), indicating that p53 overexpression is a sign of p53 mutation(s) that happen(s) with LOH for p53.
We also conducted survival analysis of hDMP1; p53 double LOH cases (n = 10 in UM samples) although the analysis was difficult due to mutual exclusiveness of LOH for hDMP1 and p53. We expected that when inactivation of two tumor suppressors overlap, the prognosis of patients will be even worse than p53 LOH alone; however, the negative effect of p53 loss was strikingly improved by simultaneous loss of hDMP1 (Figure 2). The trend was the same regardless of the origin of NSCLC samples (UM or WFU) or the cancer types (lung cancer or breast cancer). The improvement of the negative effects of p53 LOH in NSCLC was specific to hDMP1 LOH because it was not found in LOH for ARF/INK4a (Supplementary Figure S7B,C). The quenching of poor prognosis of p53 LOH by simultaneous loss of hDMP1 is consistent with relatively good prognosis of NSCLC with LOH for hDMP1 in both institutions. Although the molecular mechanism(s) underlying this phenomenon is not clear at this moment, we speculate that it is a generalized tendency of carcinoma. Our data show that Dmp1α stimulates the p53 pathway through direct physical interaction with p53 in response to dsDNA breaks [41,47]. The human DMP1 locus encodes DMP1α, and its splice variants DMP1β and γ [46]. Our preliminary study suggests that Dmp1α binds to both wt and mutant p53 although the affinity is much higher for the latter. Thus, DMP1α is either tumor suppressive (wt p53) or oncogeic (mutant p53) dependent on the p53 status of cells. DMP1β/γ do not interact with p53, is always oncogenic [45]. DMP1α regulates the human ARF promoter, the activity of which is antagonized its splice variant DMP1β [46]. Whatever the mechanism is, small molecule inhibitor screening should be performed to disrupt DMP1α and mutant p53 interaction for future cancer therapy.
The present study shows that Cyclin D1 overexpression as detected by IHC is associated with longer survival/better prognosis of NSCLC in both PFS and TS. The p values were more than 0.05 because we cannot include low level of Cyclin D1 expression (grade 1) and because not all samples were stained for Cyclin D1 in the current study. The current study shows that both hDMP1 LOH and Cyclin D1 overexpression are favorable prognostic factors associated with longer survival (Figure 1 and Figure 4). Thus we conducted quadruple analysis of NSCLC patients and found that samples that show Cyclin D1 IHC zero and hDMP1 LOH (-) have much worse prognosis than other three groups (Figure 4C,D). It is expected that tumor cells without LOH for hDMP1 and Cyclin D1, low express high hDMP1. Whatever the situation is, hDMP1 overexpression combined with no Cyclin D1 expression in IHC is associated with worse prognosis of NSCLC as shown in Figure 3C,D, delineating special group of NSCLC patients who will relapse early. Dmp1α behaves as a tumor suppressor when the p53 is wild type since it binds and stabilizes p53, but it will behave like as an oncogene when p53 is mutant. Whatever the situation is, molecular studies should be performed in the near future to explain the oncogenic role of hDMP1 in human cancer. Of note, mouse genome produces only Dmp1α, but not β or γ isoforms, explaining the difference between mice and humans.
We also studied the effects of YY1 genomic amplification in NSCLC. Its amplification again had positive effects in NSCLC survival although the data were not statistically significant. It has been reported that YY1 augments HDM2-mediated p53 polyubiquitination, and thus be oncogenic [54]. Our study shows that Dmp1α antagonizes p53’s ubiquitination by MDM2 both in vitro and in cell, and restores p53’s nuclear localization that had been lost with MDM2 expression [41]; theoretically Dmp1α will antagonize the action of YY1 to show its tumor-suppressive activity. Then the LOH for hDMP1 will nullify the effects of YY1 in NSCLC survival. As a matter of fact, the survival of YY1-amplified lung cancer almost the same between YY1 amplified cases and YY1 amplified; hDMP1 LOH cases in PFS and TS of NSCLC. We identified a group of bad prognosis in NSCLC, i.e., NSCLC without YY1 amplification without LOH for hDMP1 had the worse prognosis than the other three groups.
We will analyze mRNA levels in the future.
4. Materials and Methods
4.1. Human Lung Cancer Samples
Seventy pairs of frozen human lung cancer tissues (36 cases of adenocarcinoma, 25 cases of squamous cell carcinoma, 5 cases of large cell carcinoma, and 4 cases of adenosquamous carcinoma) and their normal counterparts were obtained from the Tissue Procurement Core Facility at the Minnesota Veteran’s hospital (UM VA hospital [56,57]). Fifty-one pairs of frozen human lung cancer tissues (33 adenocarcinoma, 16 squamous cell carcinoma, and 2 adenosquamous carcinoma) and their normal counterparts were obtained from the Tissue Procurement Core Facility at the Wake Forest University Comprehensive Cancer Center (WFU [39]). The samples had already been resected from patients with informed consent and had been stored in liquid nitrogen in both cases. The samples do not contain any subject identifiers. The human protocol had been approved by the Institutional Review Board.
4.2. Loss of Heterozygosity (LOH) and Sequencing Analyses of Human Lung Cancer Specimen
LOH assays have been conducted as described previously by PCR [39,42]. PCR products were visualized on a 1.2% agarose gel. Genotypes were identified by peak analysis of the fluorescent signal detected on an ABI 3700 DNA analyzer (Applied Biosystems). LOH was assessed if the qLOH value was found to be >2.0 or <0.5 [39,42]. The third set of primers (#176671; Table 1) was used to examine the status of hDMP1 gene 3′ when LOH study of hDMP1 locus was single with those for #198004. Real-time PCR was also conducted to study the genomic status (shown as deletion, no deletion in the Table 1).
4.3. Statistical Analyses for Mutual Exclusiveness of LOH
Chi square analyses and confidence interval assays were performed as previously described [39]. IHC studies were also performed for Cyclin D1 [43,52,53] and gene amplification study was performed for exon 4 of human YY1 [54,55]. The cut-off level for YY1 was 3.0 folds over neighbor tissues in survival analyses.
The progression-free survival (PFS) and total survival (TS) of UM NSCLC specimen was analyzed by using MedCalc software (Ostend, Belgium; 42). The TS of WFU specimens were also analyzed by the MedCalc software.
5. Conclusions
We have done survival analyses of NSCLC patients in UM and WFU samples, the data of which were obtained by LOH and IHC analyses. LOH for hDMP1 was associated with favorable prognosis while that of p53 with worse prognosis. LOH for ARF had much negative effects than INK4a loss in NSCLC survival, showing the role of ARF tumor suppressor by gene deletion. Our data also show that the adverse effect of p53 LOH was neutralized by simultaneous loss of the hDMP1 locus. We are currently unaware of the mechanism(s), which will allow more bench works employing DMP1 and p53.
Supplementary Materials
Supplementary Materials can be found at https://www.mdpi.com/1422-0067/21/21/7971/s1. Supplementary Figure S1. Total survival (TS) of patients of human non-small cell lung cancer obtained from the Wake Forest Baptist hospital dependent on LOH for hDMP1 (A), ARF/INK4a (B), and p53 loci (C). Kaplan-Meier analyses have been conducted to study the impact for of loss of each locus on non-small cell lung cancer (NSCLC) patients’ total survival up to 3500 days. The MedCalc software (Mariakerke, Belgium) was used to analyze the specimens. LOH for hDMP1 (A, n = 42) has significantly positive impact on patient’s relapse-free survival. On the other hand, LOH for ARF/INK4a did not have significant impact on NSCLC survival (the TS indicate either of the two primers; B, n = 43). p53 LOH had statistically significant negative impact on NSCLC patients’ survival (C, n = 42); Supplementary Figure S2. PFS and TS of patients of human AC and SCC of the lung cancer obtained from the Minnesota VA Hospital on LOH for hDMP1. The quadruple analyses; Supplementary Figure S3. PFS and TS of patients of human AC and SCC of the lung cancer obtained from the Minnesota VA Hospital on LOH for ARF/INK4a (either). The quadruple analyses; Supplementary Figure S4. PFS and TS of patients of human non-small cell lung cancer obtained from the Minnesota VA Hospital on LOH for ARF/INK4a (both). PFS and TS were determined by the MedCalc software. Two groups comparison was made between that of ARF/INK4a LOH (-) and ARF/INK4a LOH, both (+) (A, PFS; B, TS); Supplementary Figure S5. PFS and TS of human NSCLC obtained from UM dependent on LOH and IHC for p53. Kaplan-Meier analyses have been conducted to study the impact for the impact of loss of the p53 locus on NSCLC patients’ disease-free survival up to 5000 days. The same software was used to calculate the p and chi-square values. p53 LOH had statistically significant negative impact on NSCLC patients’ survival (A,B). Similar analyses were conducted with IHC data for the p53 protein which often overlap with LOH for p53 (C,D). The p values were relatively larger because not all samples were stained for p53, and because grade 1 staining was excluded from the study; Supplementary Figure S6. PFS and TS of patients of human AC and SCC of the lung cancer obtained from the Minnesota VA Hospital on LOH for p53 (either). The quadruple analyses; Supplementary Figure S7. The triple analysis of cancer survival for p53 in WFU (NSCLC, A), UM (NSCLC; B,C) samples. The impact of DMP1 LOH on p53 LOH was analyzed by Medcalc software for WFU and UM NSCLC samples. Loss of DMP1 neutralized the negative effect of p53 LOH in NSCLC by moving the survival curves from p53 LOH (+) to p53 LOH (-) levels or better in WFU samples. On the other hand, both PFS and TS of p53 LOH became worse by simultaneous LOH for ARF/INK4a (B,C).
Author Contributions
Conception and design: G.E.N., R.A.K. and K.I. Development of methodology: E.A.F., G.E.N., R.A.K. and F.K. Acquisition of data: E.A.F., G.E.N., R.A.K. and F.K. Analysis and interpretation of data: G.E.N., R.A.K and K.I. Writing, review and revision of the manuscript: E.A.F., G.E.N., R.A.K. and K.I., Administrative, technical, or material support: G.E.N., R.A.K. and K.I. Acquisition for grants: K.I. All authors have read and agreed to the published version of the manuscript.
Funding
K.I. was supported by NIH/NCI 2R01CA106314, ACS RSG-07-207-01-MGO, and KG080179.
Acknowledgments
We thank Gregory Kucera for total survival data for NSCLC at WFUHS hospital. We also thank Deb Sumitra for providing mutant p53 cDNAs, all other members of Inoue’s lab for sharing unpublished research data.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Oser, M.G.; Niederst, M.J.; Sequist, L.V.; Engelman, J.A. Transformation from non-small-cell lung cancer to small-cell lung cancer: Molecular drivers and cells of origin. Lancet Oncol. 2015, 16, e165–e172. [Google Scholar] [CrossRef] [Green Version]
- Satoh, Y.; Matsuo, Y.; Kuba, T.; Yamashita, K.; Sawano, M.; Tozaka, S.; Yamazaki, H.; Sonoda, D.; Mikubo, M.; Naito, M.; et al. EGFR mutation genotyping and ALK status determination in liquid-based cytology samples of non-small cell lung cancer. Virchows Arch. 2020, 476, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.; Di Maio, M.; Chiodini, P.; Rudd, R.M.; Okamoto, H.; Skarlos, D.V.; Fruh, M.; Qian, W.; Tamura, T.; Samantas, E.; et al. Carboplatin- or cisplatin-based chemotherapy in first-line treatment of small-cell lung cancer: The COCIS meta-analysis of individual patient data. J. Clin. Oncol. 2012, 30, 1692. [Google Scholar] [CrossRef] [PubMed]
- Quelle, D.E.; Zindy, F.; Ashmun, R.A.; Sherr, C.J. Alternative reading frames of the INK4a tumor suppressor gene encode two unrelated proteins capable of inducing cell cycle arrest. Cell 1995, 83, 993–1000. [Google Scholar] [PubMed] [Green Version]
- Maggi, L.B., Jr.; Winkeler, C.L.; Miceli, A.P.; Apicelli, A.J.; Brady, S.N.; Kuchenreuther, M.J.; Weber, J.D. ARF tumor suppression in the nucleolus. Biochim. Biophys. Acta 2014, 1842, 831–839. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Murphy, E. Genetic Modifiers of the p53 Pathway. Cold Spring Harb. Perspect. Med. 2016, 6, a026302. [Google Scholar] [CrossRef]
- Carrasco-Garcia, E.; Moreno, M.; Moreno-Cugnon, L.; Mathew, A. Increased Arf/p53 activity in stem cells, aging and cancer. Aging Cell 2017, 16, 219–225. [Google Scholar] [CrossRef]
- Inoue, K.; Fry, E.A. Aberrant expression of p16INK4a in human cancer—A new biomarker? Cancer Rep. Rev. 2018, 2, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Fry, E.A. Overexpression of ARF in human cancer—A new biomarker? Tumor Microenviron. 2018, 1, 37–44. [Google Scholar] [CrossRef]
- Palmero, I.; Pantoja, C.; Serrano, M. p19ARF links the tumour suppressor p53 to ras. Nature 1998, 395, 125–126. [Google Scholar] [CrossRef] [PubMed]
- Zindy, F.; Eischen, C.M.; Randle, D.H.; Kamijo, T.; Cleveland, J.L.; Sherr, C.J.; Roussel, M.F. Myc signaling via the ARF tumor suppressor regulates p53-dependent apoptosis and immortalization. Genes Dev. 1998, 12, 2424–2433. [Google Scholar] [CrossRef] [Green Version]
- Bates, S.; Phillips, A.C.; Clarke, P.A.; Stott, F.; Peters, G.; Ludwig, R.L.; Vousden, K.H. p14ARF links tumor suppressors RB and p53. Nature 1998, 395, 124–125. [Google Scholar] [CrossRef]
- Taneja, P.; Maglic, D.; Kai, F.; Sugiyama, T.; Kendig, R.D.; Frazier, D.P.; Willingham, M.C.; Inoue, K. Critical role of Dmp1 in HER2/neu-p53 signaling and breast carcinogenesis. Cancer Res. 2010, 70, 9084–9094. [Google Scholar] [CrossRef] [Green Version]
- Sherr, C.J. Autophagy by ARF: A short story. Mol. Cell 2006, 22, 436–437. [Google Scholar] [CrossRef] [PubMed]
- Quelle, D.E.; Cheng, M.; Ashmun, R.A.; Sherr, C.J. Cancer-associated mutations at the INK4a locus cancel cell cycle arrest by p16INK4a but not by the alternative reading frame protein p19ARF. Proc. Natl. Acad. Sci. USA 1997, 94, 669–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krimpenfort, P.; Quon, K.C.; Mooi, W.J.; Loonstra, A.; Berns, A. Loss of p16Ink4a confers susceptibility to metastatic melanoma in mice. Nature 2001, 413, 83–86. [Google Scholar] [CrossRef]
- Krimpenfort, P.; Ijpenberg, A.; Song, J.Y.; van der Valk, M.; Nawijn, M.; Zevenhoven, J.; Berns, A. p15Ink4b is a critical tumour suppressor in the absence of p16Ink4a. Nature 2007, 448, 943–946. [Google Scholar] [CrossRef]
- Kamijo, T.; Zindy, F.; Roussel, M.F.; Quelle, D.E.; Downing, J.R.; Ashmun, R.A.; Grosveld, G.; Sherr, C.J. Tumor suppression at the mouse INK4a locus mediated by the alternative reading frame product p19ARF. Cell 1997, 91, 649–655. [Google Scholar] [CrossRef] [Green Version]
- Sharpless, N.E.; Bardeesy, N.; Lee, K.H.; Carrasco, D.; Castrillon, D.H.; Aguirre, A.J.; Wu, E.A.; Horner, J.W.; DePinho, R.A. Loss of p16Ink4a with retention of p19Arf predisposes mice to tumorigenesis. Nature 2001, 413, 86–91. [Google Scholar] [CrossRef]
- Ruas, M.; Peters, G. The p16INK4a/CDKN2A tumor suppressor and its relatives. Biochim. Biophys. Acta 1998, 1378, F115–F177. [Google Scholar] [CrossRef]
- Gil, J.; Peters, G. Regulation of the INK4b-ARF-INK4a tumour suppressor locus: All for one or one for all. Nat. Rev. Mol. Cell Biol. 2006, 7, 667–677. [Google Scholar] [CrossRef]
- Kuo, M.L.; den Besten, W.; Bertwistle, D.; Roussel, M.F.; Sherr, C.J. N-terminal polyubiquitination and degradation of the Arf tumor suppressor. Genes Dev. 2004, 18, 1862–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stott, F.J.; Bates, S.; James, M.C.; McConnell, B.B.; Starborg, M.; Brookes, S.; Palmero, I.; Ryan, K.; Hara, E.; Vousden, K.H.; et al. The alternative product from the human CDKN2A locus, p14(ARF), participates in a regulatory feedback loop with p53 and MDM2. EMBO J. 1998, 17, 5001–5014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, J.D.; Taylor, L.J.; Roussel, M.F.; Sherr, C.J.; Bar-Sagi, D. Nucleolar Arf sequesters Mdm2 and activates p53. Nat. Cell Biol. 1999, 1, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Martelli, F.; Hamilton, T.; Silver, D.P.; Sharpless, N.E.; Bardeesy, N.; Rokas, M.; DePinho, R.A.; Livingston, D.M.; Grossman, S.R. p19ARF targets certain E2F species for degradation. Proc. Natl. Acad. Sci. USA 2001, 98, 4455–4460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itahana, K.; Zhang, Y. Mitochondrial p32 is a critical mediator of ARF-induced apoptosis. Cancer Cell 2008, 13, 542–553. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Tavana, O.; Chu, B.; Erber, L.; Chen, Y.; Baer, R.; Gu, W. NRF2 is a major target of ARF in p53-independent tumor Suppression. Mol. Cell 2017, 68, 224–332. [Google Scholar] [CrossRef]
- Hirai, H.; Sherr, C.J. Interaction of D-type cyclins with a novel myb-like transcription factor, DMP1. Mol. Cell. Biol. 1996, 16, 6457–6467. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Sherr, C.J. Gene expression and cell cycle arrest mediated by transcription factor DMP1 is antagonized by D-type cyclins through a cyclin-dependent-kinase-independent mechanism. Mol. Cell. Biol. 1998, 18, 1590–1600. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Sherr, C.J.; Shapiro, L.H. Regulation of the CD13/aminopeptidase N gene by DMP1, a transcription factor antagonized by D-type cyclins. J. Biol. Chem. 1998, 273, 29188–29194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, K.; Roussel, M.F.; Sherr, C.J. Induction of ARF tumor suppressor gene expression and cell cycle arrest by transcription factor DMP1. Proc. Natl. Acad. Sci. USA 1999, 96, 3993–3998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodner, S.M.; Naeve, C.W.; Rakestraw, K.M.; Jones, B.G.; Valentine, V.A.; Valentine, M.B.; Luthardt, F.W.; Willman, C.L.; Raimondi, S.C.; Downing, J.R.; et al. Cloning and chromosomal localization of the gene encoding human cyclin D-binding Myb-like protein (hDMP1). Gene 1999, 229, 223–229. [Google Scholar] [CrossRef]
- Inoue, K.; Wen, R.; Rehg, J.E.; Adachi, M.; Cleveland, J.L.; Roussel, M.F.; Sherr, C.J. Disruption of the ARF transcriptional activator DMP1 facilitates cell immortalization, Ras transformation, and tumorigenesis. Genes Dev. 2000, 14, 1797–1809. [Google Scholar]
- Inoue, K.; Zindy, F.; Randle, D.H.; Rehg, J.E.; Sherr, C.J. Dmp1 is haplo-insufficient for tumor suppression and modifies the frequencies of Arf and p53 mutations in Myc-induced lymphomas. Genes Dev. 2001, 15, 2934–2939. [Google Scholar] [CrossRef] [Green Version]
- Sreeramaneni, R.; Chaudhry, A.; McMahon, M.; Sherr, C.J.; Inoue, K. Ras-Raf-Arf signaling critically depends on Dmp1 transcription factor. Mol. Cell. Biol. 2005, 25, 220–232. [Google Scholar] [CrossRef] [Green Version]
- Mallakin, A.; Taneja, P.; Matise, L.A.; Willingham, M.C.; Inoue, K. Expression of Dmp1 in specific differentiated, nonproliferating cells and its repression by E2Fs. Oncogene 2006, 25, 7703–7713. [Google Scholar] [CrossRef] [Green Version]
- Taneja, P.; Mallakin, A.; Matise, L.A.; Frazier, D.P.; Choudhary, M.; Inoue, K. Repression of Dmp1 and Arf transcription by anthracyclins: Critical roles of the NF-kappaB subunit p65. Oncogene 2007, 26, 7457–7466. [Google Scholar] [CrossRef] [Green Version]
- Mallakin, A.; Sugiyama, T.; Taneja, P.; Matise, L.A.; Frazier, D.P.; Choudhary, M.; Hawkins, G.A.; D’Agostino, R.B., Jr.; Willingham, M.C.; Inoue, K. Mutually exclusive inactivation of DMP1 and ARF/p53 in lung cancer. Cancer Cell 2007, 12, 381–394. [Google Scholar] [CrossRef] [Green Version]
- Mallakin, A.; Sugiyama, T.; Kai, F.; Taneja, P.; Kendig, R.D.; Frazier, D.P.; Maglic, D.; Matise, L.A.; Willingham, M.C.; Inoue, K. The Arf-inducing transcription factor Dmp1 encodes transcriptional activator of amphiregulin, thrombospondin-1, JunB and Egr1. Int. J. Cancer 2010, 126, 1403–1416. [Google Scholar] [CrossRef] [Green Version]
- Frazier, D.P.; Kendig, R.D.; Kai, F.; Maglic, D.; Sugiyama, T.; Morgan, R.L.; Fry, E.A.; Lagedrost, S.J.; Sui, G.; Inoue, K. Dmp1 physically interacts with p53 and positively regulates p53’s stabilization, nuclear localization, and function. Cancer Res. 2012, 72, 1740–1750. [Google Scholar] [CrossRef] [Green Version]
- Maglic, D.; Zhu, S.; Taneja, P.; Fry, E.A.; Kai, F.; Kendig, R.D.; Sugiyama, T.; Miller, L.D.; Willingham, M.C.; Inoue, K. Prognostic value of the hDMP1-ARF-Hdm2-p53 pathway in breast cancer. Oncogene 2013, 32, 4120–4129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Mott, R.T.; Fry, E.A.; Taneja, P.; Kulik, G.; Sui, G.; Inoue, K. Cooperation between cyclin D1 expression and Dmp1-loss in breast cancer. Am. J. Pathol. 2013, 183, 1339–1350. [Google Scholar] [CrossRef] [Green Version]
- Fry, E.A.; Taneja, P.; Maglic, D.; Zhu, S.; Sui, G.; Inoue, K. Dmp1α inhibits HER2/neu-induced mammary tumorigenesis. PLoS ONE 2013, 8, e77870. [Google Scholar] [CrossRef] [Green Version]
- Maglic, D.; Stovall, D.B.; Cline, J.M.; Fry, E.A.; Mallakin, A.; Taneja, P.; Caudell, D.L.; Willingham, M.C.; Sui, G.; Inoue, K. DMP1β, a splice isoform of the tumor suppressor DMP1 locus, induces proliferation and progression of breast cancer. J. Pathol. 2015, 236, 90–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tschan, M.P.; Federzoni, E.A.; Haimovici, A.; Britschgi, C.; Moser, B.A.; Jin, J.; Reddy, V.A.; Sheeter, D.A.; Fischer, K.M.; Sun, P.; et al. Human DMTF1β antagonizes DMTF1α regulation of the p14(ARF) tumor suppressor and promotes cellular proliferation. Biochim. Biophys. Acta 2015, 1849, 1198–1208. [Google Scholar] [CrossRef] [Green Version]
- Kendig, R.D.; Kai, F.; Fry, E.A.; Inoue, K. Stabilization of the p53-DNA complex by the nuclear protein Dmp1α. Cancer Investig. 2017, 35, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Fry, E.A.; Frazier, D.P. Transcription factors that interact with p53 and Mdm2. Int. J. Cancer 2016, 138, 1577–1585. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Fry, E.A. Aberrant splicing of the DMP1-INK4a/ARF-MDM2-p53 pathway in cancer. Int. J. Cancer 2016, 139, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fry, E.A.; Taneja, P.; Inoue, K. Oncogenic and tumor-suppressive mouse models for breast cancer employing HER2/neu. Int. J. Cancer 2017, 140, 495–503. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Fry, E.A. Haplo-insufficient tumor suppressor genes. In Advances in Medicine and Biology; Nova Science Publishers, Inc.: Suite N Hauppauge, NY, USA, 2017; Volume 118, Chapter 6. [Google Scholar]
- Qie, S.; Diehl, J.A. Cyclin D1, cancer progression, and opportunities in cancer treatment. J. Mol. Med. (Berl.) 2016, 94, 1313–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, S.C.; Lee, S.S.; Abraham, J. Mechanisms of therapeutic CDK4/6 inhibition in breast cancer. Semin. Oncol. 2017, 44, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Sui, G.; Affar, B.; Shi, Y.; Brignone, C.; Wall, N.R.; Yin, P.; Donohoe, M.; Luke, M.P.; Calvo, D.; Grossman, S.R.; et al. Yin Yang 1 is a negative regulator of p53. Cell 2004, 117, 859–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Stovall, D.B.; Inoue, K.; Sui, G. The oncogenic role of Yin Yang 1. Crit. Rev. Oncog. 2011, 16, 163–197. [Google Scholar] [CrossRef]
- Kratzke, R.A.; Greatens, T.M.; Rubins, J.B.; Maddaus, M.A.; Niewoehner, D.E.; Niehans, G.A.; Geradts, J. Rb and p16INK4a expression in resected non-small cell lung tumors. Cancer Res. 1996, 56, 3415–3420. [Google Scholar]
- Zhou, J.X.; Niehans, G.A.; Shar, A.; Rubins, J.B.; Frizelle, S.P.; Kratzke, R.A. Mechanisms of G1 checkpoint loss in resected early stage non-small cell lung cancer. Lung Cancer 2001, 32, 27–38. [Google Scholar] [CrossRef]
Figure 1.
Progression-free Survival (PFS) and total survival (TS) of 70 cases of human non-small cell lung cancer obtained from the University of Minnesota VA hospital (UM) dependent on LOH for hDMP1 (A,B) and ARF/INK4a loci (C–F). Kaplan-Meier analyses have been conducted to study the impact for the impact of loss of each locus on non-small cell lung cancer (NSCLC) patients’ disease-free survival up to 5000 days. The Med Calc software (Mariakerke, Belgium) was used to analyze the specimens. LOH for hDMP1 (A,B) has significantly positive impact on patient’s relapse-free survival. On the other hand, LOH for ARF/INK4a did not have significant impact on NSCLC survival although there was a trend for association of worse prognosis (the survival indicate either of the two primers; C,D). When the ARF/INK4a locus was studied separately, the prognosis of patients with ARF LOH was much worse than that of INK4a only, indicating much stronger prognostic power for the former than the latter (E,F).
Figure 1.
Progression-free Survival (PFS) and total survival (TS) of 70 cases of human non-small cell lung cancer obtained from the University of Minnesota VA hospital (UM) dependent on LOH for hDMP1 (A,B) and ARF/INK4a loci (C–F). Kaplan-Meier analyses have been conducted to study the impact for the impact of loss of each locus on non-small cell lung cancer (NSCLC) patients’ disease-free survival up to 5000 days. The Med Calc software (Mariakerke, Belgium) was used to analyze the specimens. LOH for hDMP1 (A,B) has significantly positive impact on patient’s relapse-free survival. On the other hand, LOH for ARF/INK4a did not have significant impact on NSCLC survival although there was a trend for association of worse prognosis (the survival indicate either of the two primers; C,D). When the ARF/INK4a locus was studied separately, the prognosis of patients with ARF LOH was much worse than that of INK4a only, indicating much stronger prognostic power for the former than the latter (E,F).
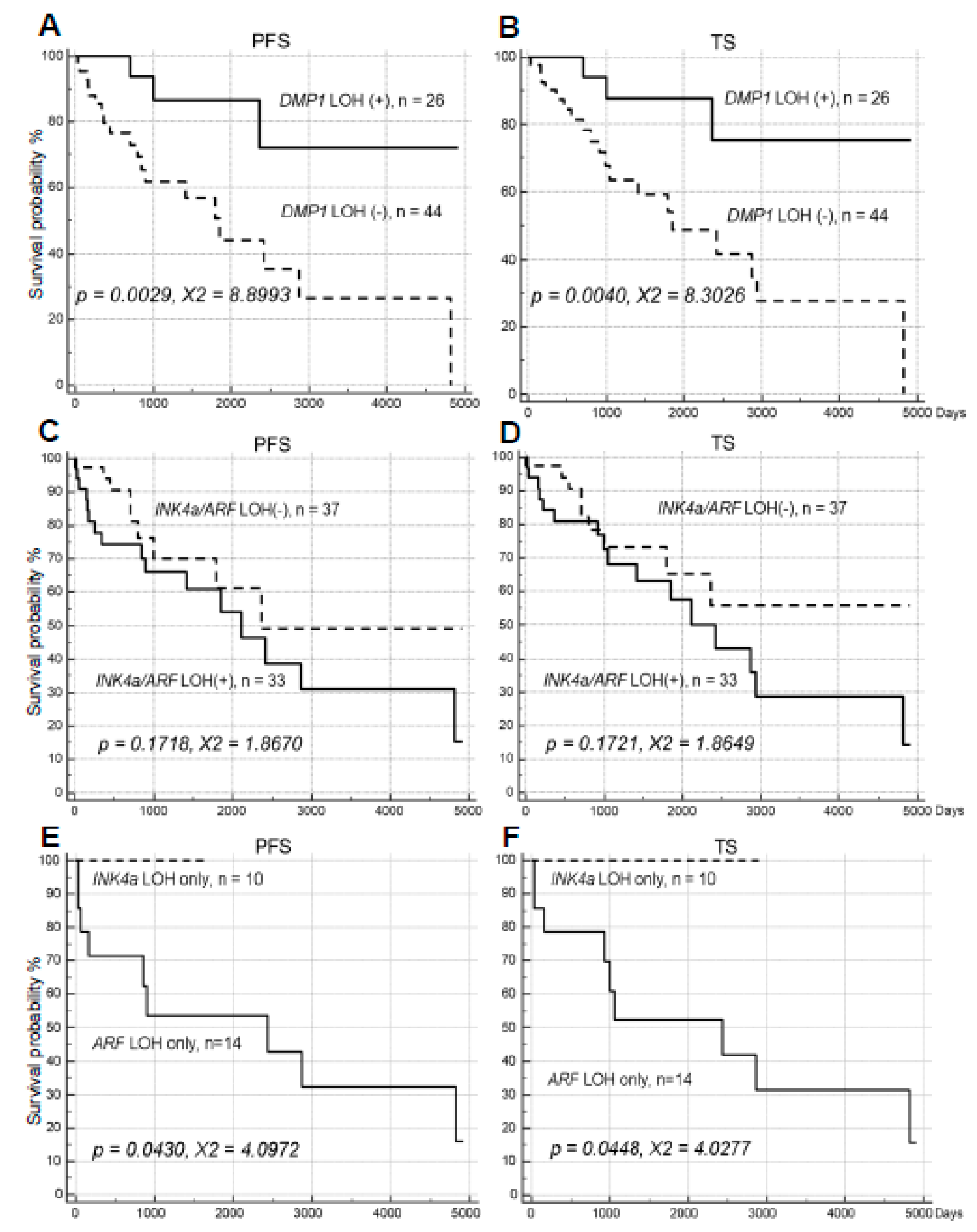
Figure 2.
The triple analysis of cancer survival for p53 in University of Minnesota (UM) (NSCLC), and Wake Forest University (WFU) (breast cancer) samples. The impact of DMP1 LOH on p53 LOH was analyzed by Medcalc software for UM (NSCLC) and WFU (breast cancer) samples. Loss of DMP1 neutralized the negative effect of p53 LOH in NSCLC by moving the survival curves from p53 LOH (+) to p53 LOH(-) levels in both PFS and TS in UM samples. The same trend was observed in statistically significant fashion in WFU NSCLC samples (n = 44) or WFU breast cancer samples (n = 105) suggesting that the improvement of cancer survival of p53 LOH samples with loss of the DMP1 locus is a generalized phenomenon. (A) PFS of lung cancer (UM); (B) TS of lung cancer (UM), (C) DFS of breast cancer (BC, WFUHS).
Figure 2.
The triple analysis of cancer survival for p53 in University of Minnesota (UM) (NSCLC), and Wake Forest University (WFU) (breast cancer) samples. The impact of DMP1 LOH on p53 LOH was analyzed by Medcalc software for UM (NSCLC) and WFU (breast cancer) samples. Loss of DMP1 neutralized the negative effect of p53 LOH in NSCLC by moving the survival curves from p53 LOH (+) to p53 LOH(-) levels in both PFS and TS in UM samples. The same trend was observed in statistically significant fashion in WFU NSCLC samples (n = 44) or WFU breast cancer samples (n = 105) suggesting that the improvement of cancer survival of p53 LOH samples with loss of the DMP1 locus is a generalized phenomenon. (A) PFS of lung cancer (UM); (B) TS of lung cancer (UM), (C) DFS of breast cancer (BC, WFUHS).
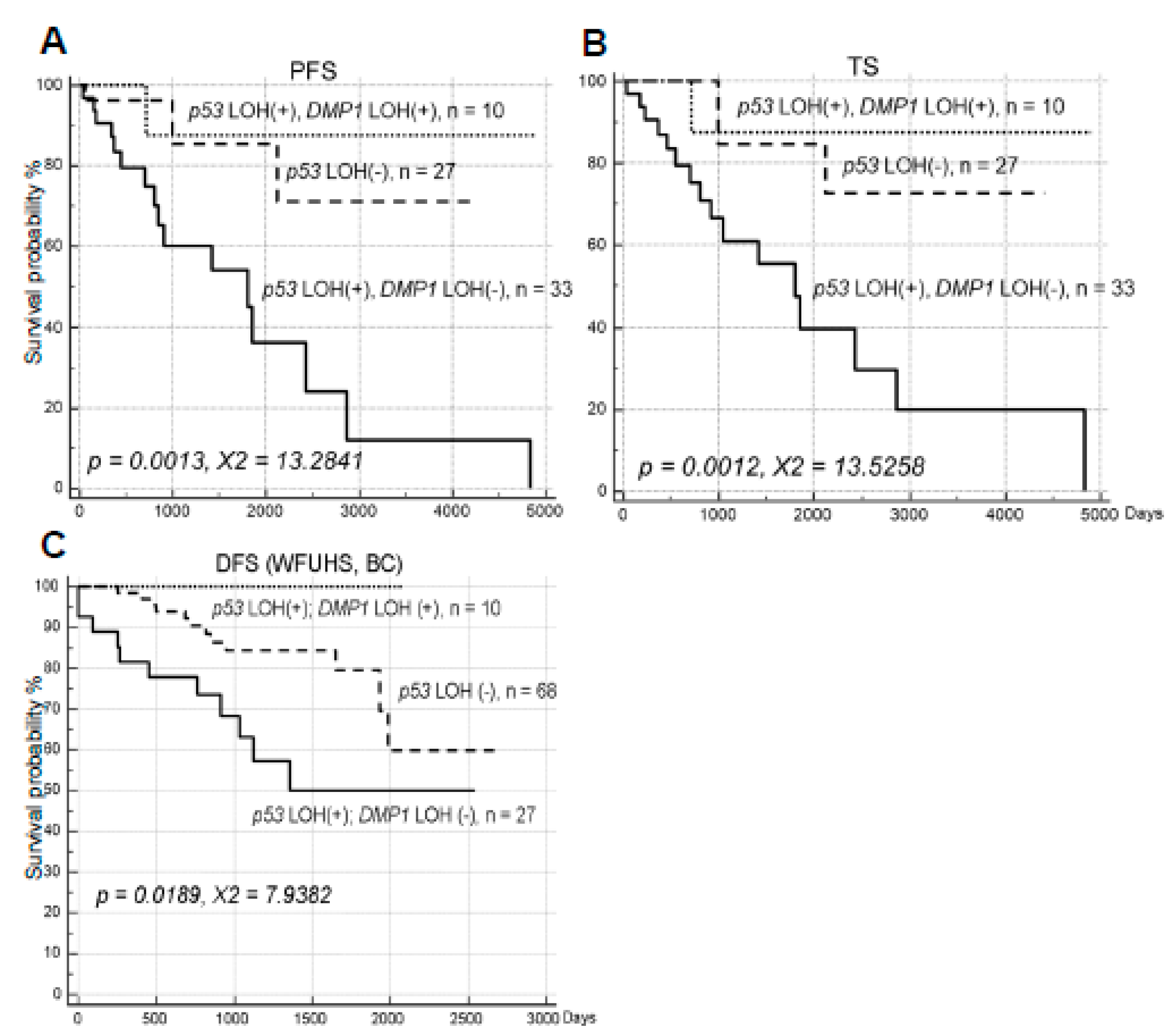
Figure 3.
Progression-free survival (PFS) and total survival (TS) analysis of human NSCLC obtained from UM dependent on Cyclin D1. (A,B) PFS and TS analysis of human NSCLC obtained from UM dependent on Cyclin D1 in human NSCLC (n = 62). High expression of Cyclin D1 protein tended to be associated with longer survival. (C,D) The quadruple analysis of NSCLC samples on Cyclin D1 expression and LOH for DMP1. Quadruple analysis was conducted in UM NSCLC samples for Cyclin D1 high (2+); DMP1 LOH (+) and (-), and Cyclin D1 no expression; DMP1 LOH (+) and (-). Both Cyclin D1 expression and LOH for DMP1 are associated with favorable prognosis; we could identify NSCLC patients with worst prognosis in the group of Cyclin D1, no expression; DMP1 LOH (-) group.
Figure 3.
Progression-free survival (PFS) and total survival (TS) analysis of human NSCLC obtained from UM dependent on Cyclin D1. (A,B) PFS and TS analysis of human NSCLC obtained from UM dependent on Cyclin D1 in human NSCLC (n = 62). High expression of Cyclin D1 protein tended to be associated with longer survival. (C,D) The quadruple analysis of NSCLC samples on Cyclin D1 expression and LOH for DMP1. Quadruple analysis was conducted in UM NSCLC samples for Cyclin D1 high (2+); DMP1 LOH (+) and (-), and Cyclin D1 no expression; DMP1 LOH (+) and (-). Both Cyclin D1 expression and LOH for DMP1 are associated with favorable prognosis; we could identify NSCLC patients with worst prognosis in the group of Cyclin D1, no expression; DMP1 LOH (-) group.
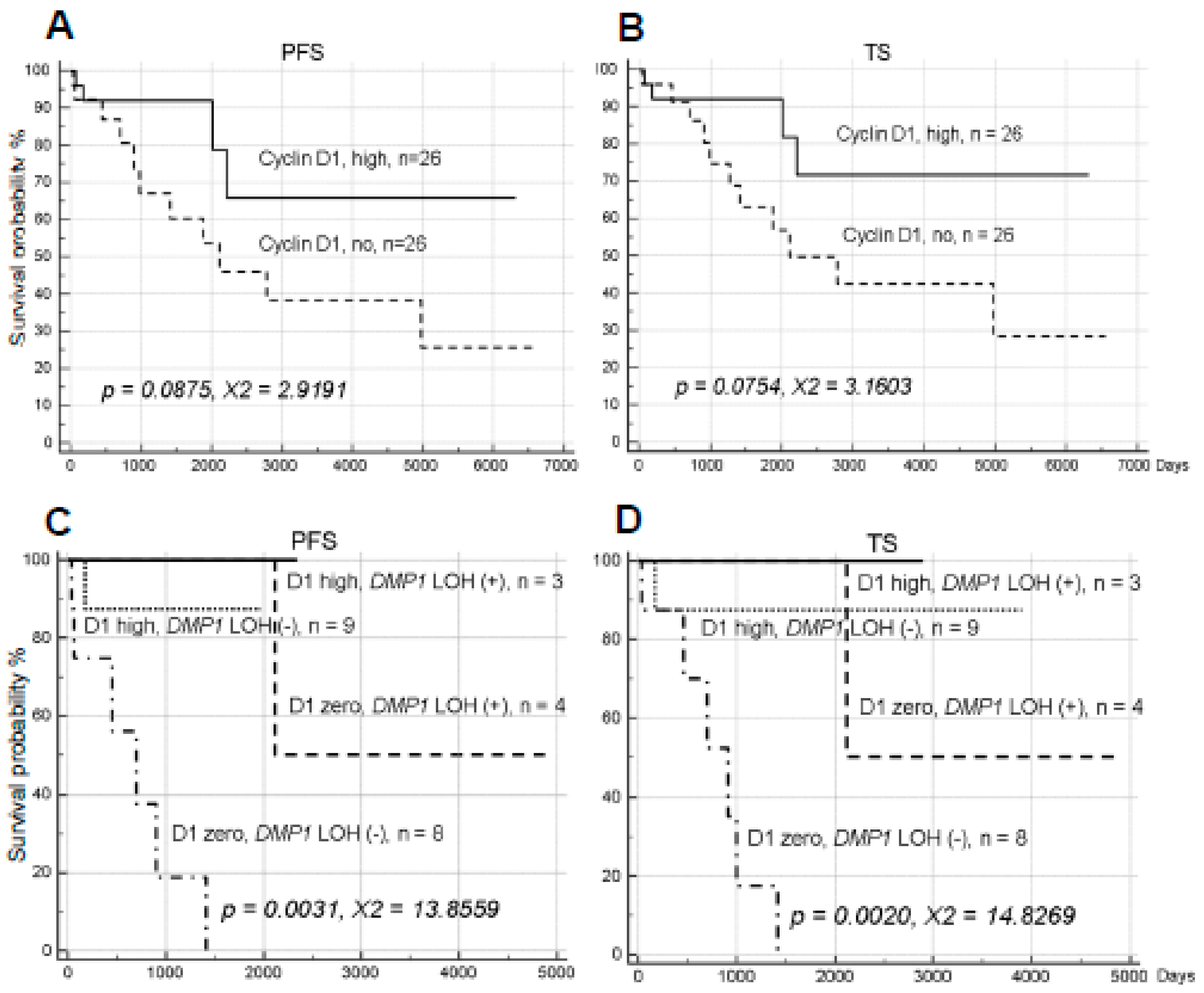
Figure 4.
PFS and TS analysis of human NSCLC obtained from UM dependent on YY1. (A,B) PFS and TS analysis of human NSCLC obtained from UM dependent on YY1 in human NSCLC. (C,D) The quadruple analysis of NSCLC samples on YY1 amplification and LOH for DMP1. Quadruple analysis was conducted in UM NSCLC samples for YY1 amplification (>3 folds); DMP1 LOH (+) and (-); YY1 no-amplification; DMP1 LOH (+) and (-); YY1 amplification. Both YY1 amplification and LOH for DMP1 are associated with favorable prognosis; we could identify NSCLC patients with worst prognosis in the group of YY1 no-amplification; DMP1 LOH (-) group.
Figure 4.
PFS and TS analysis of human NSCLC obtained from UM dependent on YY1. (A,B) PFS and TS analysis of human NSCLC obtained from UM dependent on YY1 in human NSCLC. (C,D) The quadruple analysis of NSCLC samples on YY1 amplification and LOH for DMP1. Quadruple analysis was conducted in UM NSCLC samples for YY1 amplification (>3 folds); DMP1 LOH (+) and (-); YY1 no-amplification; DMP1 LOH (+) and (-); YY1 amplification. Both YY1 amplification and LOH for DMP1 are associated with favorable prognosis; we could identify NSCLC patients with worst prognosis in the group of YY1 no-amplification; DMP1 LOH (-) group.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
NSCLC samples of UM VA hospital. The LOH values of each genomic locus is shown. When both loci shows a single peak, real-time PCR was conducted to determine gene deletion. NME: Non-mutually exclusive, ME: Mutually exclusive. IHC: Immunohistochemistry. Not only chi square tests, but also 95% confidence interval assays were conducted to evaluate mutual exclusiveness of LOH for genomic loci. Mutually exclusive cases for LOH and immunohistochemistry (IHC), the results of confidence interval assays are shaded.
Table 1.
NSCLC samples of UM VA hospital. The LOH values of each genomic locus is shown. When both loci shows a single peak, real-time PCR was conducted to determine gene deletion. NME: Non-mutually exclusive, ME: Mutually exclusive. IHC: Immunohistochemistry. Not only chi square tests, but also 95% confidence interval assays were conducted to evaluate mutual exclusiveness of LOH for genomic loci. Mutually exclusive cases for LOH and immunohistochemistry (IHC), the results of confidence interval assays are shaded.
| Patient ID | hDMP1 | INK4a/ARF | INK4a/ARF | I/A LOH ME | p53 | p53 LOH ME | p53 IHC | p53 IHC ME | ||
|---|---|---|---|---|---|---|---|---|---|---|
| #92465 | #198004 | #33647 | #27251 | #182 SE | #377 SE | |||||
| Group 1 | 5′ | 3′ | 5′ | 3′ | 5′ | 3′ | ||||
| 307 | 0.68 | 0.26 | 0.15 | 0.98 | NME | 2.01 | 2.04 | NME | ND | |
| 308 | 0.70 | 1.04 | 3.38 | 0.41 | ME | Deletion | 27.20 | ME | ND | |
| 309 | 1.21 | 2.51 | 1.42 | 1.11 | ME | 1.06 | 0.54 | ME | ND | |
| 310 | 0.06 | 4.31 | 0.68 | 0.93 | ME | 1.06 | 0.92 | ME | ND | |
| 311 | 1.05 | 1.08 | 1.31 | 1.26 | None | 1.50 | 0.84 | None | ND | |
| 313 | 0.94 | 0.47 | 1.46 | 1.32 | ME | 1.23 | 0.96 | ME | ND | |
| 314 | 0.46 | 6.09 | 2.41 | 0.63 | NME | 0.43 | 0.30 | NME | ND | |
| 315 | 1.09 | 1.87 | 1.12 | 0.98 | None | 9.97 | 0.28 | ME | ND | |
| 317 | 0.61 | 2.26 | 1.56 | 0.91 | ME | Deletion | 1.40 | NME | ND | |
| 318 | 1.23 | 0.90 | 5.48 | 2.21 | ME | 0.42 | 0.38 | ME | ND | |
| 319 | 1.86 | 2.29 | 1.30 | 0.82 | ME | 2.16 | 3.57 | NME | ND | |
| 321 | 0.77 | 0.29 | 0.69 | 1.03 | ME | 1.26 | 1.74 | None | ND | |
| 322 | 1.12 | 2.01 | 0.97 | 0.93 | ME | 1.16 | 0.07 | NME | ND | |
| 323 | 0.86 | 0.52 | 0.75 | 1.06 | None | 1.08 | 1.56 | None | ND | |
| 324 | 0.64 | 2.96 | 0.77 | 0.16 | NME | 2.31 | 0.31 | NME | ND | |
| 325 | 1.10 | 1.51 | 2.13 | 0.45 | ME | 0.34 | 0.29 | ME | ND | |
| 326 | 1.00 | 0.59 | 0.28 | 0.60 | ME | 2.37 | 0.62 | ME | ND | |
| 327 | 1.32 | >10.0 | 1.04 | 1.03 | ME | 1.80 | No deletion | ME | ND | |
| 328 | 2.33 | 4.97 | 1.53 | 1.34 | ME | No deletion | No deletion | ME | 2 | NME |
| 329 | 0.74 | 0.80 | 1.02 | 2.69 | ME | 0.25 | 0.9 | ME | 2 | ME |
| 330 | 0.87 | 0.72 | 0.96 | 0.64 | None | 0.88 | 13.97 | ME | 1 | None |
| 331 | 1.11 | 1.08 | 0.10 | 4.66 | ME | 1.34 | 0.48 | ME | 0 | None |
| 332 | 0.82 | 0.52 | 0.68 | 0.74 | None | 0.97 | 0.40 | ME | 2 | ME |
| 333 | 1.15 | 0.48 | 0.89 | 1.06 | ME | 1.89 | 1.09 | ME | 1 | ME |
| 277 | 0.41 | 0.41 | 1.19 | 1.02 | ME | 0.61 | No deletion | ME | 1 | ME |
| 295 | 0.74 | 0.13 | 1.24 | 1.01 | ME | 0.99 | 1.02 | ME | 1 | ME |
| 299 | 0.32 | 2.11 | 1.18 | 0.79 | ME | 0.87 | 0.51 | ME | 1 | ME |
| 338 | 1.73 | 1.27 | 1.54 | 0.04 | ME | 13.1 | 4.06 | ME | 0 | None |
| 349 | 2.95 | 20.00 | 0.93 | 1.21 | ME | 1.73 | 1.62 | ME | 1 | ME |
| 351 | 1.69 | 0.41 | No deletion | No deletion | ME | 0.83 | 1.08 | ME | 1 | ME |
| 379 | 0.98 | 0.18 | 0.76 | 0.64 | ME | 0.92 | 0.94 | ME | 2 | NME |
| 380 | 0.33 | 0.33 | 0.67 | 0.80 | ME | 0.55 | 1.82 | ME | ND | |
| 271 | 0.66 | 0.59 | 0.49 | 0.91 | ME | 0.65 | 3.39 | ME | 2 | ME |
| 272 | 0.87 | 0.92 | 0.78 | 1.93 | None | 0.92 | 0.82 | None | 0 | None |
| 273 | 0.57 | 0.41 | 0.32 | 0.83 | NME | 0.23 | Deletion | NME | 2 | NME |
| 274 | 1.02 | 0.87 | 0.70 | 0.95 | None | 0.69 | 0.17 | ME | 1 | None |
| 275 | 0.55 | 0.90 | 0.18 | 1.04 | ME | 0.35 | Deletion | ME | 2 | ME |
| 278 | 0.95 | No deletion | 0.76 | 3.11 | ME | 0.56 | 14.00 | ME | 2 | ME |
| 282 | 1.12 | 1.02 | 1.03 | 0.95 | None | 0.93 | 4.01 | ME | 2 | ME |
| 284 | 1.02 | 1.02 | 0.35 | 0.92 | ME | 2.27 | 10.40 | ME | 0 | None |
| 290 | 1.03 | 0.92 | 2.48 | 1.03 | ME | 0.96 | 0.92 | None | 2 | ME |
| 293 | 0.94 | 0.99 | 0.60 | 0.73 | None | 1.86 | 0.47 | ME | 0 | None |
| Group 2 | ||||||||||
| 269 | 0.74 | 0.74 | 0.76 | 0.15 | ME | 0.71 | 0.33 | ME | 2 | ME |
| 270 | 1.08 | 1.16 | 1.10 | 0.99 | None | 0.73 | 1.06 | None | 2 | ME |
| 276 | 0.85 | 1.07 | 4.00 | 1.41 | ME | 0.15 | 3.08 | ME | 2 | ME |
| 300 | 1.49 | 0.96 | 1.30 | 0.66 | None | 2.23 | 0.57 | ME | 2 | ME |
| 334 | 0.80 | 0.61 | 0.35 | 0.94 | ME | 1.05 | 4.73 | ME | 2 | ME |
| 337 | 1.06 | 0.81 | 1.09 | 0.88 | None | 1.66 | 2.07 | ME | 2 | ME |
| 339 | 0.66 | 1.75 | 1.27 | 0.07 | ME | 0.93 | 0.95 | None | 2 | ME |
| 340 | 3.90 | 0.91 | 1.30 | 1.25 | ME | 1.71 | No deletion | ME | 1 | ME |
| 353 | 4.28 | 0.37 | 1.20 | 0.59 | ME | 3.62 | No deletion | NME | 1 | ME |
| 356 | 1.68 | 1.03 | 1.65 | 0.59 | None | No deletion | No deletion | None | 0 | None |
| 357 | 1.17 | 1.26 | 1.88 | 0.18 | ME | 2.59 | 2.55 | ME | 1 | None |
| 358 | 1.07 | 1.23 | 2.91 | 1.45 | ME | 2.01 | 0.65 | ME | 1 | None |
| 360 | 0.95 | 1.02 | 0.25 | 0.41 | ME | 0.45 | 3.11 | ME | 2 | ME |
| 362 | 1.64 | No deletion | 1.33 | 1.03 | None | 0.88 | 0.22 | ME | 2 | ME |
| 363 | 0.84 | 0.88 | 0.27 | 1.22 | ME | 0.35 | 0.18 | ME | 1 | None |
| 364 | 1.04 | 1.07 | 0.11 | 0.06 | ME | 0.75 | 0.16 | ME | 0 | None |
| 365 | No deletion | 0.93 | 1.29 | 0.95 | None | 0.75 | 0.93 | None | 1 | None |
| 366 | 0.87 | 1.26 | 0.44 | 0.99 | ME | 1.04 | 0.98 | None | 2 | ME |
| 367 | 0.92 | 1.04 | 0.78 | 2.15 | ME | 0.70 | 1.20 | None | 0 | None |
| 368 | 0.97 | 0.48 | 0.35 | 0.27 | NME | 1.52 | 0.91 | ME | 1 | ME |
| 369 | 1.49 | 1.06 | 1.40 | 1.27 | None | 3.42 | No deletion | ME | 0 | None |
| 370 | 0.55 | 1.11 | 1.76 | 0.26 | ME | 0.47 | No deletion | ME | 2 | ME |
| 371 | 1.49 | 0.56 | 8.11 | 2.46 | ME | 0.71 | 0.25 | ME | 0 | None |
| 372 | 2.50 | 1.72 | 0.51 | 0.57 | ME | 0.27 | No deletion | NME | 2 | NME |
| 381 | 1.30 | 0.99 | 1.02 | 1.19 | None | 0.55 | 0.49 | ME | 0 | None |
| 384 | 1.63 | 0.80 | 2.53 | 0.48 | ME | 1.54 | 1.38 | None | 2 | ME |
| 387 | 1.23 | 0.26 | 0.39 | 1.99 | NME | 2.04 | 1.80 | NME | 0 | ME |
| 389 | 1.14 | 1.23 | 1.47 | 0.17 | ME | 1.16 | 0.42 | ME | 1 | None |
| Percentage | 26/70 = 37.1% | 33/70 = 47.1% | 47/53 = 88.7% | 43/70 = 61.4% | 48/58 = 82.8% | 29/33 = 87.9% | ||||
| p = 0.0023 | p = 0.0128 | |||||||||
| X2 = 9.330 | X2 = 6.195 | |||||||||
| 95% CI | 80.1–97.2% | 95% CI | 75.6–93.9% | 76.7–99.0% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fry, E.A.; Niehans, G.E.; Kratzke, R.A.; Kai, F.; Inoue, K. Survival of Lung Cancer Patients Dependent on the LOH Status for DMP1, ARF, and p53. Int. J. Mol. Sci. 2020, 21, 7971. https://doi.org/10.3390/ijms21217971
AMA Style
Fry EA, Niehans GE, Kratzke RA, Kai F, Inoue K. Survival of Lung Cancer Patients Dependent on the LOH Status for DMP1, ARF, and p53. International Journal of Molecular Sciences. 2020; 21(21):7971. https://doi.org/10.3390/ijms21217971
Chicago/Turabian StyleFry, Elizabeth A., Gloria E. Niehans, Robert A. Kratzke, Fumitake Kai, and Kazushi Inoue. 2020. "Survival of Lung Cancer Patients Dependent on the LOH Status for DMP1, ARF, and p53" International Journal of Molecular Sciences 21, no. 21: 7971. https://doi.org/10.3390/ijms21217971
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.