Y-27632 Induces Neurite Outgrowth by Activating the NOX1-Mediated AKT and PAK1 Phosphorylation Cascades in PC12 Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
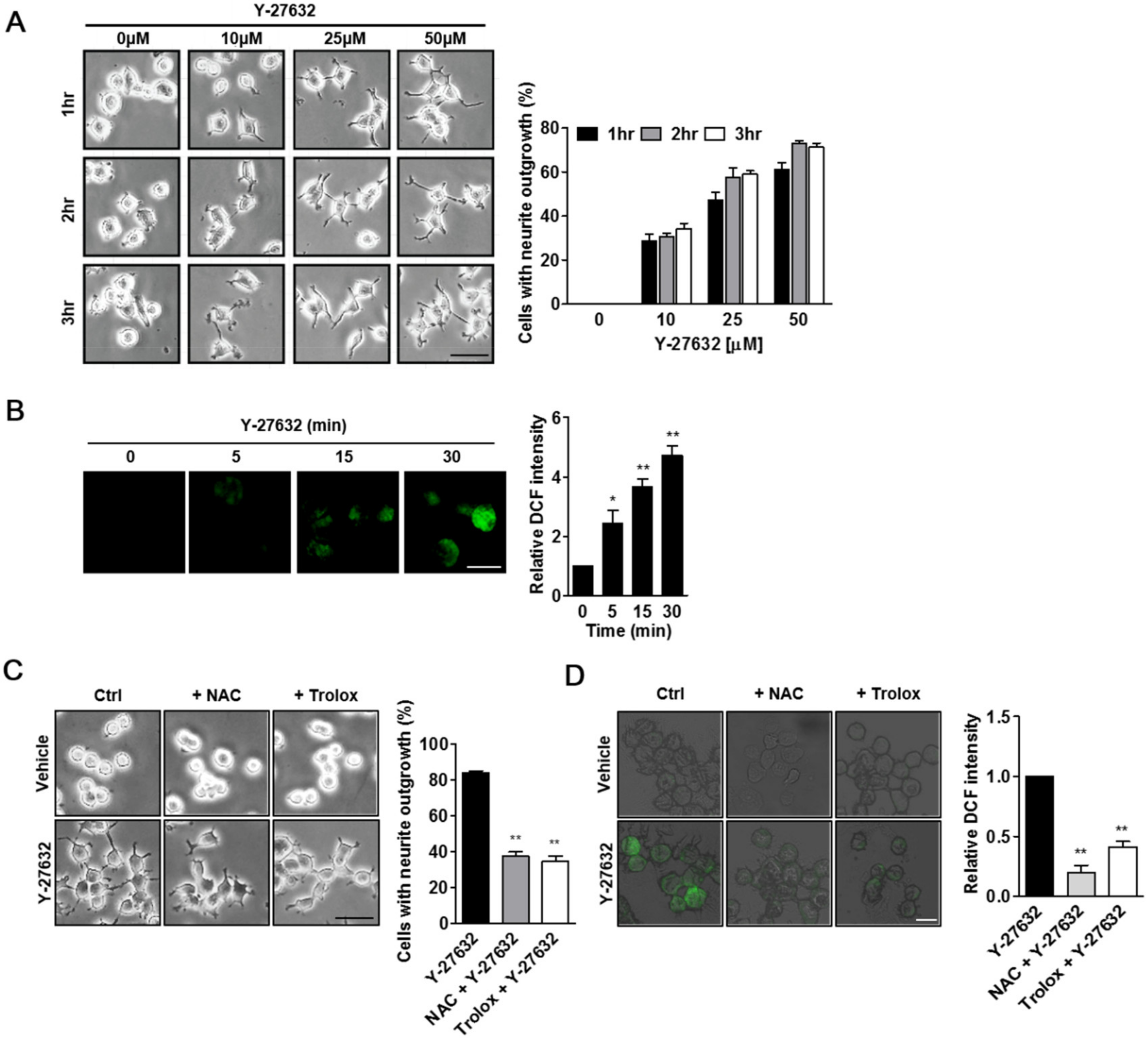
2.1. Role of ROS in Y-27632-Induced Neurite Outgrowth in PC12 Cells
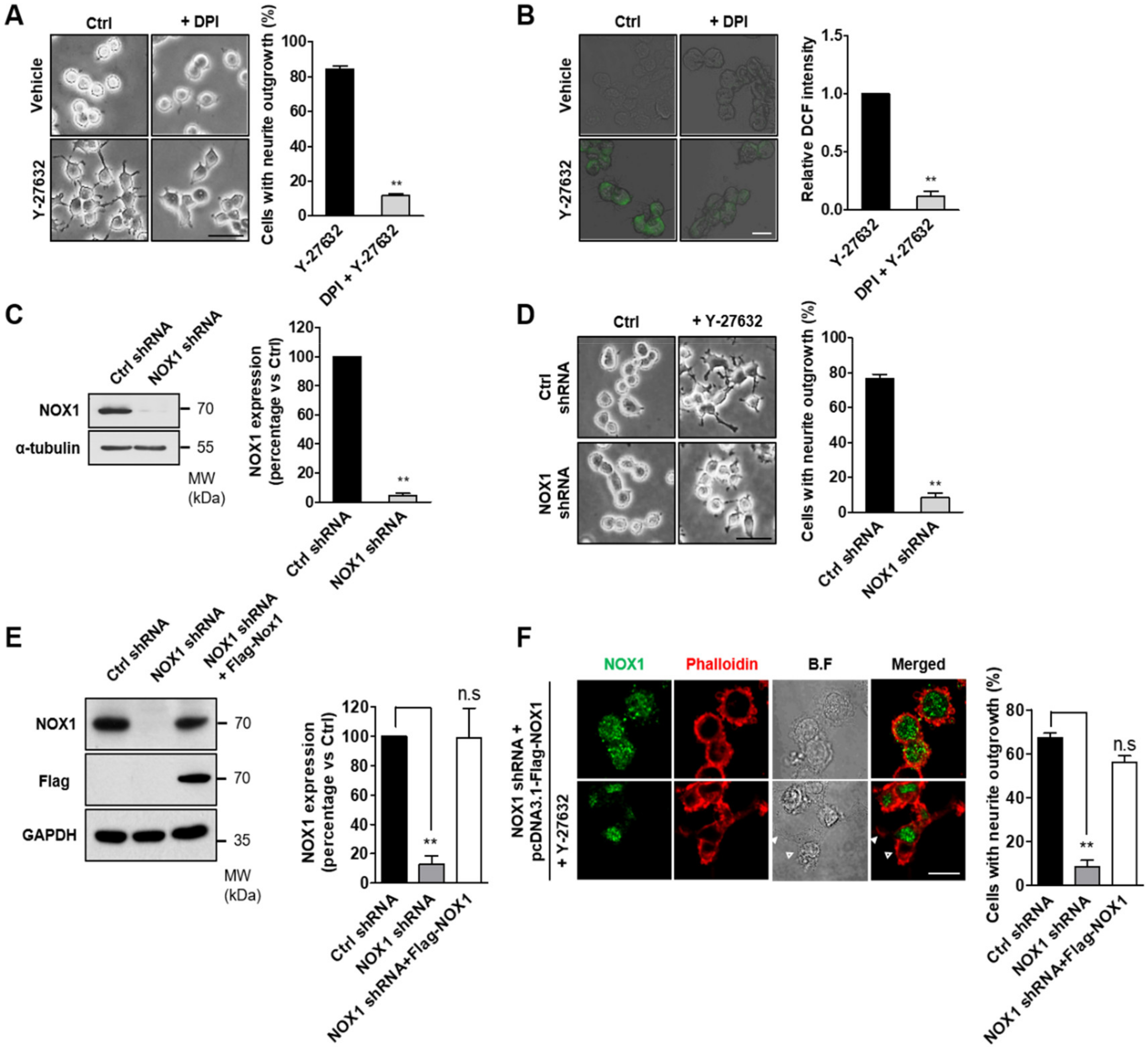
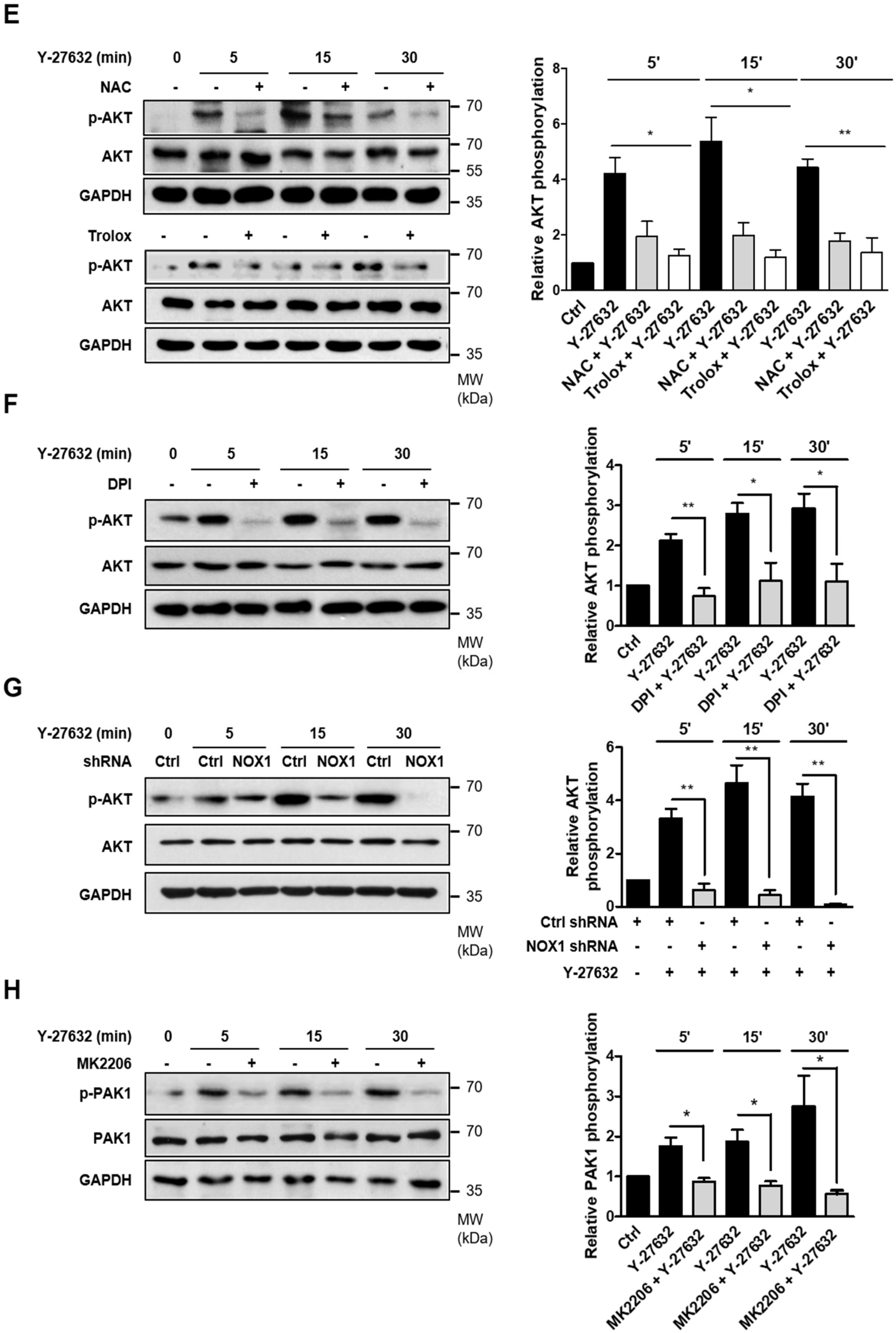
2.2. NOX-Dependent ROS Production during Y-27632-Induced Neurite Outgrowth
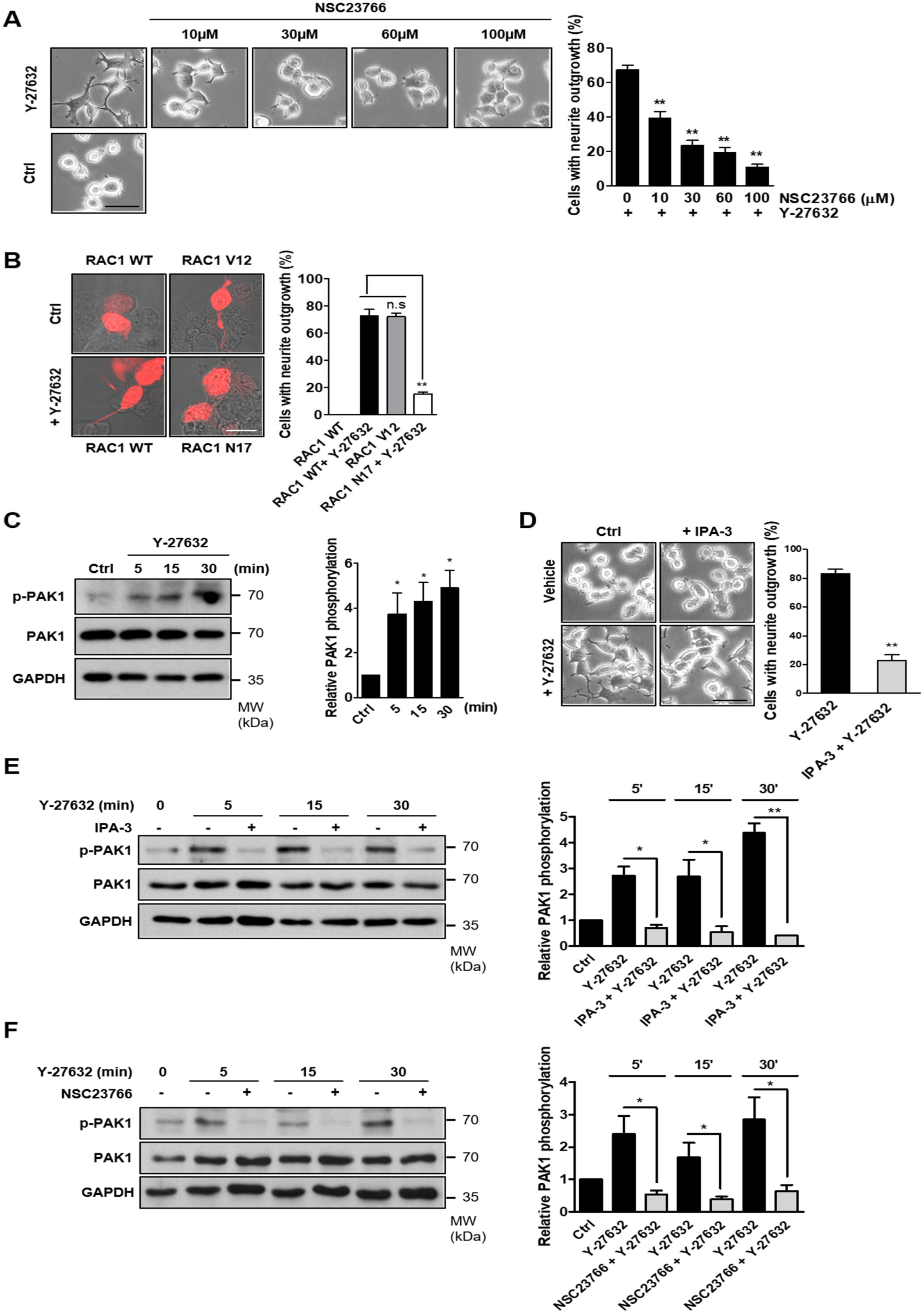
2.3. Involvement of Rac1-Mediated PAK1 Activation in Y-27632-Induced Neurite Outgrowth
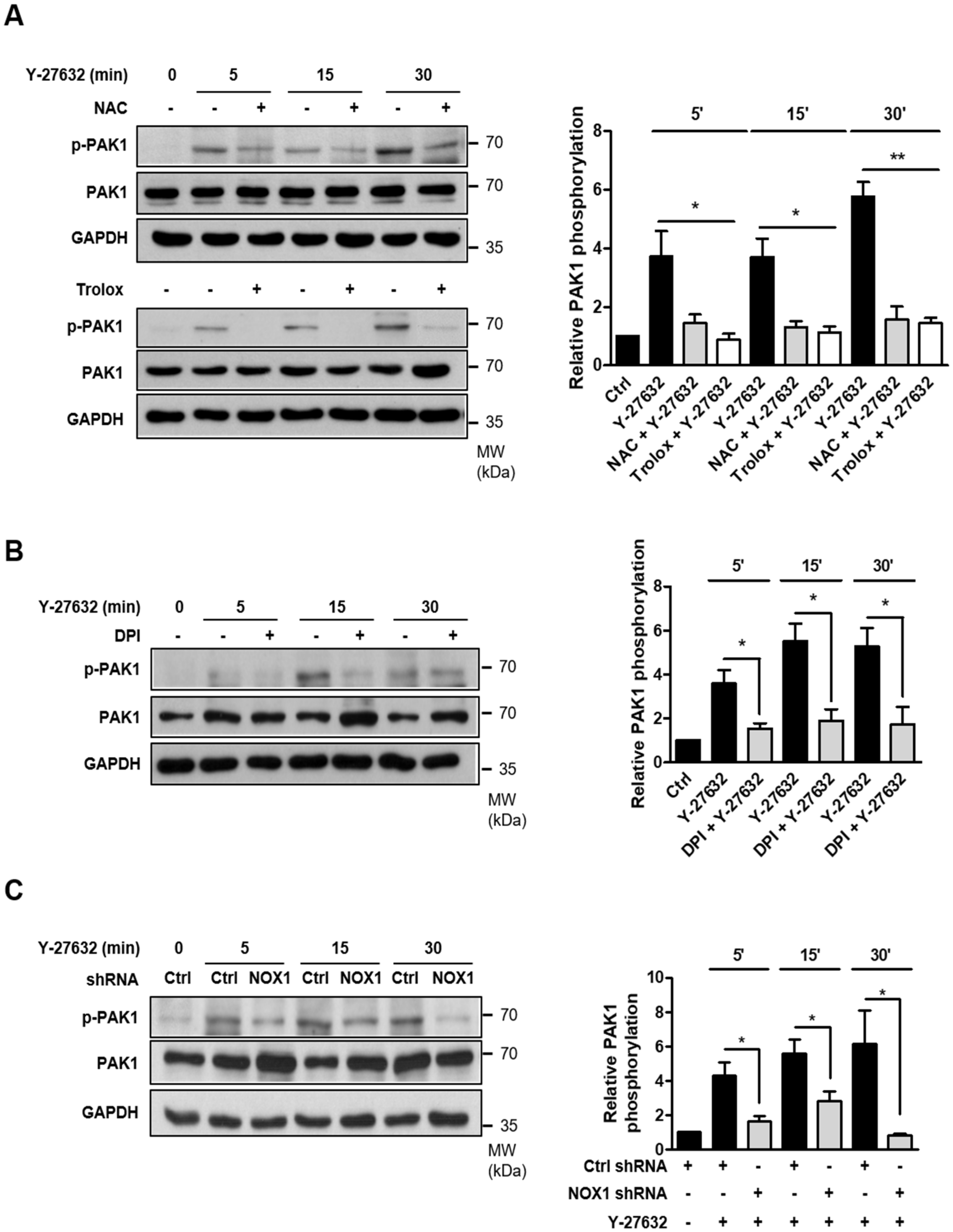
2.4. Activation of PAK1 Is Induced by NOX-Dependent ROS Generation in Y-27632-Stimulated PC12 Cells
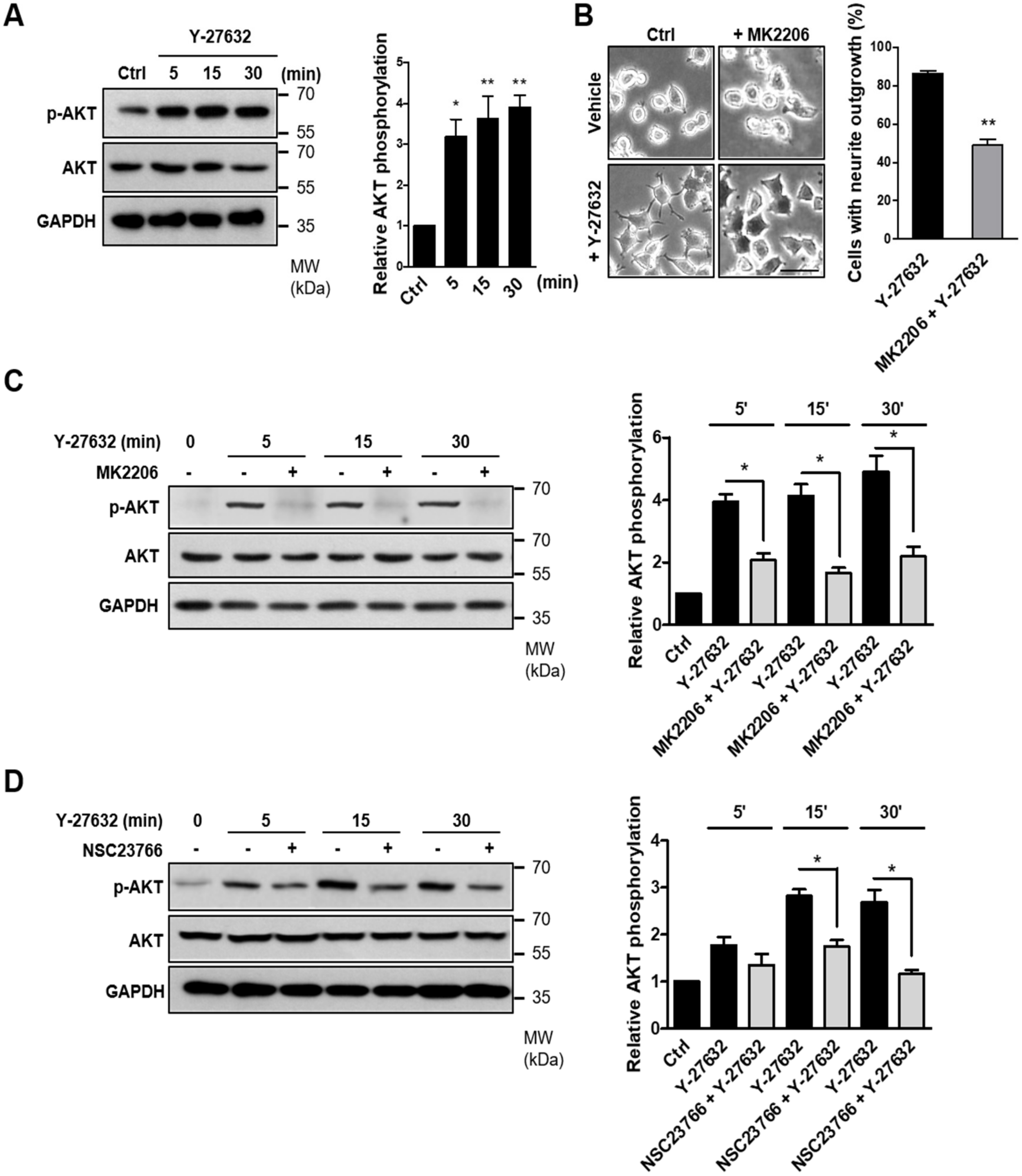
2.5. Involvement of the AKT-PAK1 Cascade in Y-27632-Induced Neurite Outgrowth
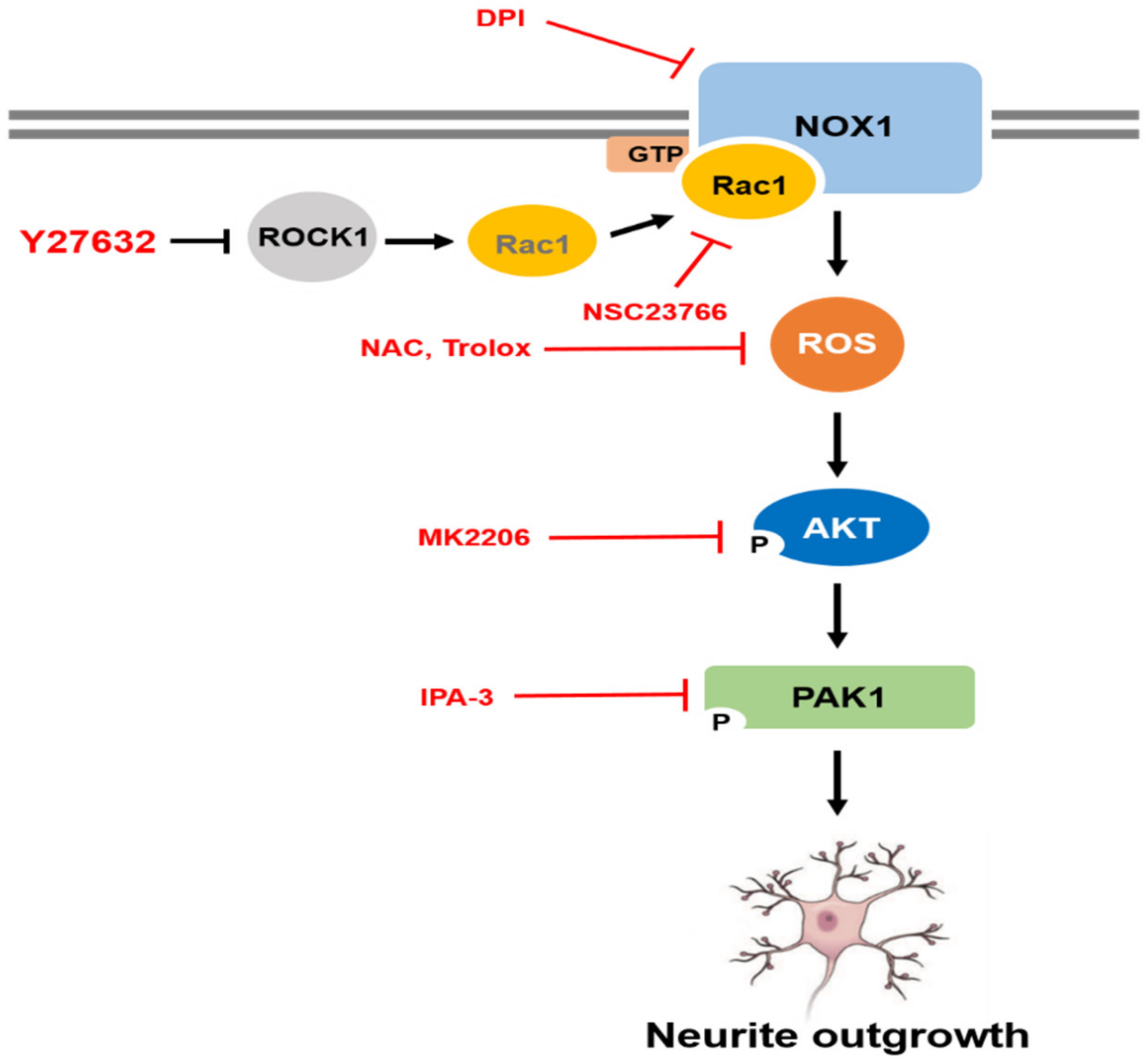
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Measurement of Neurite Outgrowth
4.3. Pharmacological Treatment
4.4. Measurement of Intracellular Reactive Oxygen Species (ROS)
4.5. Generation of Stable NOX1 shRNA-Expressing Cells
4.6. Western Blotting
4.7. Cell Transfection
4.8. Immunocytochemistry (ICC)
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ROCK | Rho-associated coiled coil-forming kinase |
| ROS | Reactive oxygen species |
| NGF | Nerve growth factor |
| NAC | N-acetyl cysteine |
| DPI | Diphenyleneiodonium |
| NOX | NADPH oxidase |
| PAK1 | p21-activated kinase 1 |
| GAPDH | Glyceraldehyde-3-Phosphate Dehydrogenase |
References
- Blanquie, O.; Bradke, F. Cytoskeleton dynamics in axon regeneration. Curr. Opin. Neurobiol. 2018, 51, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.E.; Suter, D.M. An Integrated Cytoskeletal Model of Neurite Outgrowth. Front. Cell. Neurosci. 2018, 12, 447. [Google Scholar] [CrossRef] [PubMed]
- Tönges, L.; Koch, J.C.; Bähr, M.; Lingor, P. ROCKing regeneration: Rho kinase inhibition as molecular target for neurorestoration. Front. Mol. Neurosci. 2011, 4, 39. [Google Scholar] [CrossRef] [PubMed]
- Boomkamp, S.D.; Riehle, M.O.; Wood, J.; Olson, M.; Barnett, S.C. The development of a rat in vitro model of spinal cord injury demonstrating the additive effects of rho and ROCK inhibitors on neurite outgrowth and myelination. Glia 2012, 60, 441–456. [Google Scholar] [CrossRef]
- Fournier, A.E.; Takizawa, B.T.; Strittmatter, S.M. Rho Kinase Inhibition Enhances Axonal Regeneration in the Injured CNS. J. Neurosci. 2003, 23, 1416–1423. [Google Scholar] [CrossRef]
- Fujita, A.; Hattori, Y.; Takeuchi, T.; Kamata, Y.; Hata, F. NGF induces neurite outgrowth via a decrease in phosphorylation of myosin light chain in PC12 cells. NeuroReport 2001, 12, 3599–3602. [Google Scholar] [CrossRef]
- Minase, T.; Ishima, T.; Itoh, K.; Hashimoto, N. Potentiation of nerve growth factor-induced neurite outgrowth by the ROCK inhibitor Y-27632: A possible role of IP3 receptors. Eur. J. Pharmacol. 2010, 648, 67–73. [Google Scholar] [CrossRef]
- Mueller, B.K.; Mack, H.; Teusch, N. Rho kinase, a promising drug target for neurological disorders. Nat. Rev. Drug Discov. 2005, 4, 387–398. [Google Scholar] [CrossRef]
- Ishizaki, T.; Uehata, M.; Tamechika, I.; Keel, J.; Nonomura, K.; Maekawa, M.; Narumiya, S. Pharmacological properties of Y-27632, a specific inhibitor of rho-associated kinases. Mol. Pharmacol. 2000, 57, 976–983. [Google Scholar]
- Uehata, M.; Ishizaki, T.; Satoh, H.; Ono, T.; Kawahara, T.; Morishita, T.; Tamakawa, H.; Yamagami, K.; Inui, J.; Maekawa, M.; et al. Calcium sensitization of smooth muscle mediated by a Rho-associated protein kinase in hypertension. Nat. Cell Biol. 1997, 389, 990–994. [Google Scholar] [CrossRef]
- Chen, K.; Zhang, W.; Chen, J.; Li, S.; Guo, G. Rho-associated protein kinase modulates neurite extension by regulating microtubule remodeling and vinculin distribution. Neural Regen. Res. 2013, 8, 3027–3035. [Google Scholar] [PubMed]
- Jia, X.-F.; Ye, F.; Wang, Y.-B.; Feng, D.-X. ROCK inhibition enhances neurite outgrowth in neural stem cells by upregulating YAP expression in vitro. Neural Regen. Res. 2016, 11, 983–987. [Google Scholar] [CrossRef] [PubMed]
- Feng, P.-L.; Wang, J.; Yang, Z.-J.; Liu, X.-H.; Zhong, Y. Effect of Y-27632 on the cultured retinal neurocytes of rats. Int. J. Ophthalmol. 2013, 6, 15–18. [Google Scholar] [PubMed]
- Roloff, F.; Scheiblich, H.; Dewitz, C.; Dempewolf, S.; Stern, M.; Bicker, G. Enhanced Neurite Outgrowth of Human Model (NT2) Neurons by Small-Molecule Inhibitors of Rho/ROCK Signaling. PLoS ONE 2015, 10, e0118536. [Google Scholar] [CrossRef]
- Schmandke, A.; Schmandke, A.; Strittmatter, S.M. ROCK and Rho: Biochemistry and Neuronal Functions of Rho-Associated Protein Kinases. Neuroscientist 2007, 13, 454–469. [Google Scholar] [CrossRef]
- Tang, B.L.; Liou, Y.C. Novel Modulators of Amyloid-Beta Precursor Protein Processing. J. Neurochem. 2007, 100, 314–323. [Google Scholar] [CrossRef]
- Shin, H.K.; Esalomone, S.; Ayata, C. Targeting cerebrovascular Rho-kinase in stroke. Expert Opin. Ther. Targets 2008, 12, 1547–1564. [Google Scholar] [CrossRef]
- Sung, J.K.; Miao, L.; Calvert, J.W.; Huang, L.; Harkey, H.L.; Zhang, J.H. A possible role of RhoA/Rho-kinase in experimental spinal cord injury in rat. Brain Res. 2003, 959, 29–38. [Google Scholar] [CrossRef]
- Zhou, Y.; Su, Y.; Li, B.; Liu, F.; Ryder, J.W.; Wu, X.; Gonzalez-DeWhitt, P.A.; Gelfanova, V.; Hale, J.E.; May, P.C.; et al. Nonsteroidal anti-inflammatory drugs can lower amyloidogenic Abeta42 by inhibiting Rho. Science 2003, 302, 1215–1217. [Google Scholar] [CrossRef]
- Gunther, R.; Saal, K.A.; Suhr, M.; Scheer, D.; Koch, J.C.; Bahr, M.; Lingor, P.; Tonges, L. The rho kinase inhibitor Y-27632 improves motor performance in male SOD1G93A mice. Front. Neurosci. 2014, 8, 304. [Google Scholar] [CrossRef]
- Bai, J.; Nakamura, H.; Kwon, Y.-W.; Hattori, I.; Yamaguchi, Y.; Kim, Y.-C.; Kondo, N.; Oka, S.-I.; Ueda, S.; Masutani, H.; et al. Critical Roles of Thioredoxin in Nerve Growth Factor-Mediated Signal Transduction and Neurite Outgrowth in PC12 Cells. J. Neurosci. 2003, 23, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Horstkorte, R.; Reinke, S.; Bauer, C.; Reutter, W.; Kontou, M. N-Propionylmannosamine-induced over-expression and secretion of thioredoxin leads to neurite outgrowth of PC12 cells. Biochem. Biophys. Res. Commun. 2010, 395, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; An, J.M.; Gil Lee, H.; Seo, S.R.; Kim, S.S.; Kim, J.Y.; Kang, J.-W.; Bae, Y.S.; Seo, J.T. Activation of Rac1-dependent redox signaling is critically involved in staurosporine-induced neurite outgrowth in PC12 cells. Free Radic. Res. 2013, 47, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Brandes, R.P.; Weissmann, N.; Schröder, K. Nox family NADPH oxidases: Molecular mechanisms of activation. Free Radic. Biol. Med. 2014, 76, 208–226. [Google Scholar] [CrossRef]
- Cheng, G.; Diebold, B.A.; Hughes, Y.; Lambeth, J.D. Nox1-dependent Reactive Oxygen Generation Is Regulated by Rac1. J. Biol. Chem. 2006, 281, 17718–17726. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Katoh, H.; Yasui, H.; Mori, K.; Negishi, M. RhoA Inhibits the Nerve Growth Factor-induced Rac1 Activation through Rho-associated Kinase-dependent Pathway. J. Biol. Chem. 2001, 276, 18977–18983. [Google Scholar] [CrossRef]
- Takefuji, M.; Mori, K.; Morita, Y.; Arimura, N.; Nishimura, T.; Nakayama, M.; Hoshino, M.; Iwamatsu, A.; Murohara, T.; Kaibuchi, K.; et al. Rho-kinase modulates the function of STEF, a Rac GEF, through its phosphorylation. Biochem. Biophys. Res. Commun. 2007, 355, 788–794. [Google Scholar] [CrossRef]
- Manser, E.; Huang, H.Y.; Loo, T.H.; Chen, X.Q.; Dong, J.M.; Leung, T.; Lim, L. Expression of constitutively active alpha-PAK reveals effects of the kinase on actin and focal complexes. Mol. Cell. Biol. 1997, 17, 1129–1143. [Google Scholar] [CrossRef]
- Daniels, R.H.; Hall, P.S.; Bokoch, G.M. Membrane targeting of p21-activated kinase 1 (PAK1) induces neurite outgrowth from PC12 cells. EMBO J. 1998, 17, 754–764. [Google Scholar] [CrossRef]
- Tang, Y.; Zhou, H.; Chen, A.I.; Pittman, R.N.; Field, J. The Akt Proto-oncogene Links Ras to Pak and Cell Survival Signals. J. Biol. Chem. 2000, 275, 9106–9109. [Google Scholar] [CrossRef]
- Read, D.E.; Gorman, A.M. Involvement of Akt in neurite outgrowth. Cell. Mol. Life Sci. 2009, 66, 2975–2984. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Seger, R.; Babu, C.V.S.; Hwang, S.-Y.; Yoo, Y.S. A positive role of the PI3-K/Akt signaling pathway in PC12 cell differentiation. Mol. Cells 2004, 18, 353–359. [Google Scholar] [PubMed]
- Tsatmali, M.; Walcott, E.C.; Makarenkova, H.; Crossin, K.L. Reactive oxygen species modulate the differentiation of neurons in clonal cortical cultures. Mol. Cell. Neurosci. 2006, 33, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Estrada, C.P.; Covacu, R.; Sankavaram, S.R.; Svensson, M.; Brundin, L. Oxidative Stress Increases Neurogenesis and Oligodendrogenesis in Adult Neural Progenitor Cells. Stem Cells Dev. 2014, 23, 2311–2327. [Google Scholar] [CrossRef]
- Tsatmali, M.; Walcott, E.C.; Crossin, K.L. Newborn neurons acquire high levels of reactive oxygen species and increased mitochondrial proteins upon differentiation from progenitors. Brain Res. 2005, 1040, 137–150. [Google Scholar] [CrossRef]
- Suzukawa, K.; Miura, K.; Mitsushita, J.; Resau, J.; Hirose, K.; Crystal, R.; Kamata, T. Nerve Growth Factor-induced Neuronal Differentiation Requires Generation of Rac1-regulated Reactive Oxygen Species. J. Biol. Chem. 2000, 275, 13175–13178. [Google Scholar] [CrossRef]
- Munnamalai, V.; Suter, D.M. Reactive oxygen species regulate F-actin dynamics in neuronal growth cones and neurite outgrowth. J. Neurochem. 2009, 108, 644–661. [Google Scholar] [CrossRef]
- Ridley, A.J.; Hall, A. The Small GTP-Binding Protein Rho Regulates the Assembly of Focal Adhesions and Actin Stress Fibers in Response to Growth Factors. Cell 1992, 70, 389–399. [Google Scholar] [CrossRef]
- Etienne-Manneville, S.; Hall, A. Rho GTPases in cell biology. Nat. Cell Biol. 2002, 420, 629–635. [Google Scholar] [CrossRef]
- Kozma, R.; Sarner, S.; Ahmed, S.; Lim, L. Rho family GTPases and neuronal growth cone remodelling: Relationship between increased complexity induced by Cdc42Hs, Rac1, and acetylcholine and collapse induced by RhoA and lysophosphatidic acid. Mol. Cell. Biol. 1997, 17, 1201–1211. [Google Scholar] [CrossRef]
- Manser, E.; Leung, T.; Salihuddin, H.; Zhao, Z.-S.; Lim, L. A brain serine/threonine protein kinase activated by Cdc42 and Rac1. Nat. Cell Biol. 1994, 367, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.-L.; Zhuo, Y.; King, C.C.; Fryer, B.H.; Bokoch, G.M.; Field, J. Akt Phosphorylation of Serine 21 on Pak1 Modulates Nck Binding and Cell Migration. Mol. Cell. Biol. 2003, 23, 8058–8069. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Du, J.; Hu, Z.; Liu, J.; Tian, Y.; Zhu, Y.; Wang, L.; Gu, L. Activation of Rac1-PI3K/Akt is required for epidermal growth factor-induced PAK1 activation and cell migration in MDA-MB-231 breast cancer cells. J. Biomed. Res. 2011, 25, 237–245. [Google Scholar] [CrossRef]
- Yao, R.; Cooper, G. Requirement for phosphatidylinositol-3 kinase in the prevention of apoptosis by nerve growth factor. Science 1995, 267, 2003–2006. [Google Scholar] [CrossRef] [PubMed]
- Namikawa, K.; Honma, M.; Abe, K.; Takeda, M.; Mansur, K.; Obata, T.; Miwa, A.; Okado, H.; Kiyama, H. Akt/Protein Kinase B Prevents Injury-Induced Motoneuron Death and Accelerates Axonal Regeneration. J. Neurosci. 2000, 20, 2875–2886. [Google Scholar] [CrossRef]
- Andjelkovic, M.; Suidan, H.S.; Meier, R.; Frech, M.; Alessi, D.R.; Hemmings, B.A. Nerve growth factor promotes activation of the alpha, beta and gamma isoforms of protein kinase B in PC12 pheochromocytoma cells. Eur. J. Biol. Inorg. Chem. 1998, 251, 195–200. [Google Scholar] [CrossRef]
- Liu, L.-Z.; Hu, X.-W.; Xia, C.; He, J.; Zhou, Q.; Shi, X.; Fang, J.; Jiang, B.-H. Reactive oxygen species regulate epidermal growth factor-induced vascular endothelial growth factor and hypoxia-inducible factor-1α expression through activation of AKT and P70S6K1 in human ovarian cancer cells. Free Radic. Biol. Med. 2006, 41, 1521–1533. [Google Scholar] [CrossRef]
- Hervera, A.; De Virgiliis, F.; Palmisano, I.; Zhou, L.; Tantardini, E.; Kong, G.; Hutson, T.; Danzi, M.C.; Perry, R.B.-T.; Santos, C.X.C.; et al. Reactive oxygen species regulate axonal regeneration through the release of exosomal NADPH oxidase 2 complexes into injured axons. Nat. Cell Biol. 2018, 20, 307–319. [Google Scholar] [CrossRef]
- Lee, S.-R.; Yang, K.-S.; Kwon, J.; Lee, C.; Jeong, W.; Rhee, S.G. Reversible Inactivation of the Tumor Suppressor PTEN by H2O2. J. Biol. Chem. 2002, 277, 20336–20342. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Z.; Liu, W.; Ai, Z. ROS-generating oxidase NOX1 promotes the self-renewal activity of CD133+ thyroid cancer cells through activation of the Akt signaling. Cancer Lett. 2019, 447, 154–163. [Google Scholar] [CrossRef]
- Li, L.; Liu, B. ROCK inhibitor Y-27632 protects rats against cerebral ischemia/reperfusion-induced behavioral deficits and hippocampal damage. Mol. Med. Rep. 2019, 20, 3395–3405. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Rashid, H.; Fujita, R.; Contos, J.J.A.; Chun, J.; Ueda, H. Initiation of neuropathic pain requires lysophosphatidic acid receptor signaling. Nat. Med. 2004, 10, 712–718. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.Y.; An, J.M.; Seo, J.T.; Seo, S.R. Y-27632 Induces Neurite Outgrowth by Activating the NOX1-Mediated AKT and PAK1 Phosphorylation Cascades in PC12 Cells. Int. J. Mol. Sci. 2020, 21, 7679. https://doi.org/10.3390/ijms21207679
Park SY, An JM, Seo JT, Seo SR. Y-27632 Induces Neurite Outgrowth by Activating the NOX1-Mediated AKT and PAK1 Phosphorylation Cascades in PC12 Cells. International Journal of Molecular Sciences. 2020; 21(20):7679. https://doi.org/10.3390/ijms21207679
Chicago/Turabian StylePark, So Yeong, Jeong Mi An, Jeong Taeg Seo, and Su Ryeon Seo. 2020. "Y-27632 Induces Neurite Outgrowth by Activating the NOX1-Mediated AKT and PAK1 Phosphorylation Cascades in PC12 Cells" International Journal of Molecular Sciences 21, no. 20: 7679. https://doi.org/10.3390/ijms21207679
APA StylePark, S. Y., An, J. M., Seo, J. T., & Seo, S. R. (2020). Y-27632 Induces Neurite Outgrowth by Activating the NOX1-Mediated AKT and PAK1 Phosphorylation Cascades in PC12 Cells. International Journal of Molecular Sciences, 21(20), 7679. https://doi.org/10.3390/ijms21207679