Proteomics-Based Evidence for a Pro-Oncogenic Role of ESRP1 in Human Colorectal Cancer Cells

 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
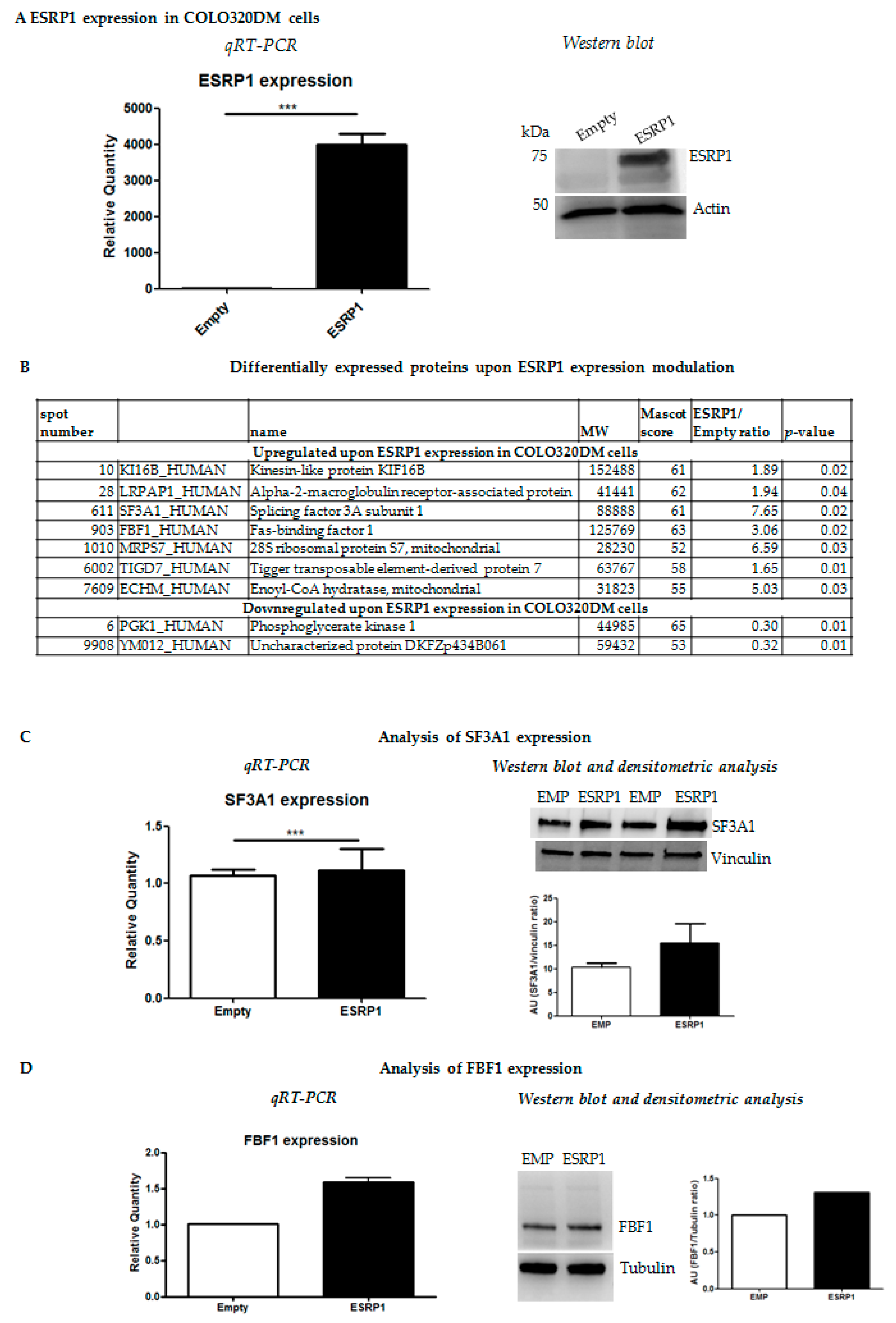
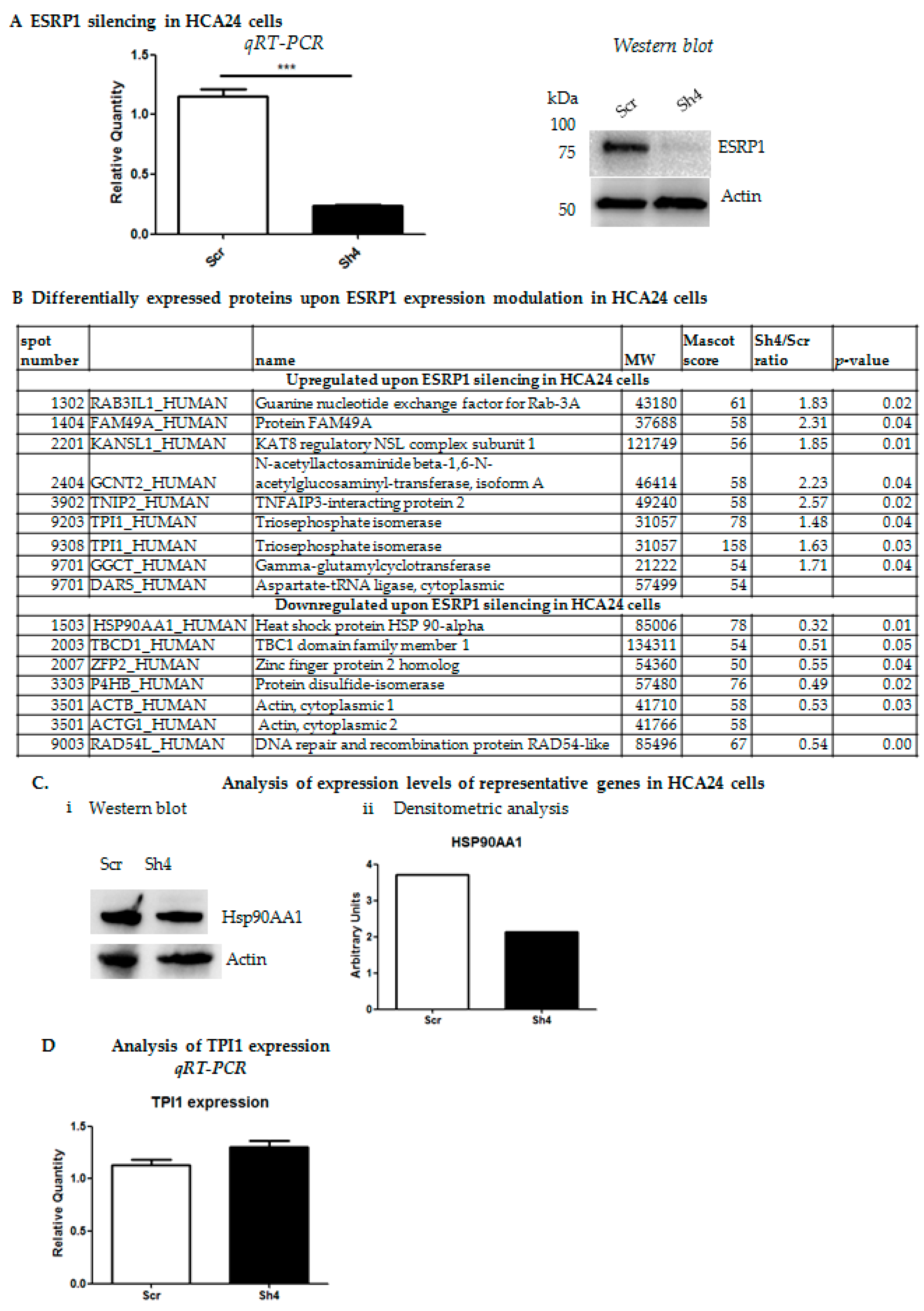
2.1. Proteomics Analysis Reveals Differential Expression of Several Cancer-Related Proteins upon Modulation of ESRP1 Expression in Human CRC Cells
Functional Characterization of Differentially Expressed Proteins Ranked Correlated Sets
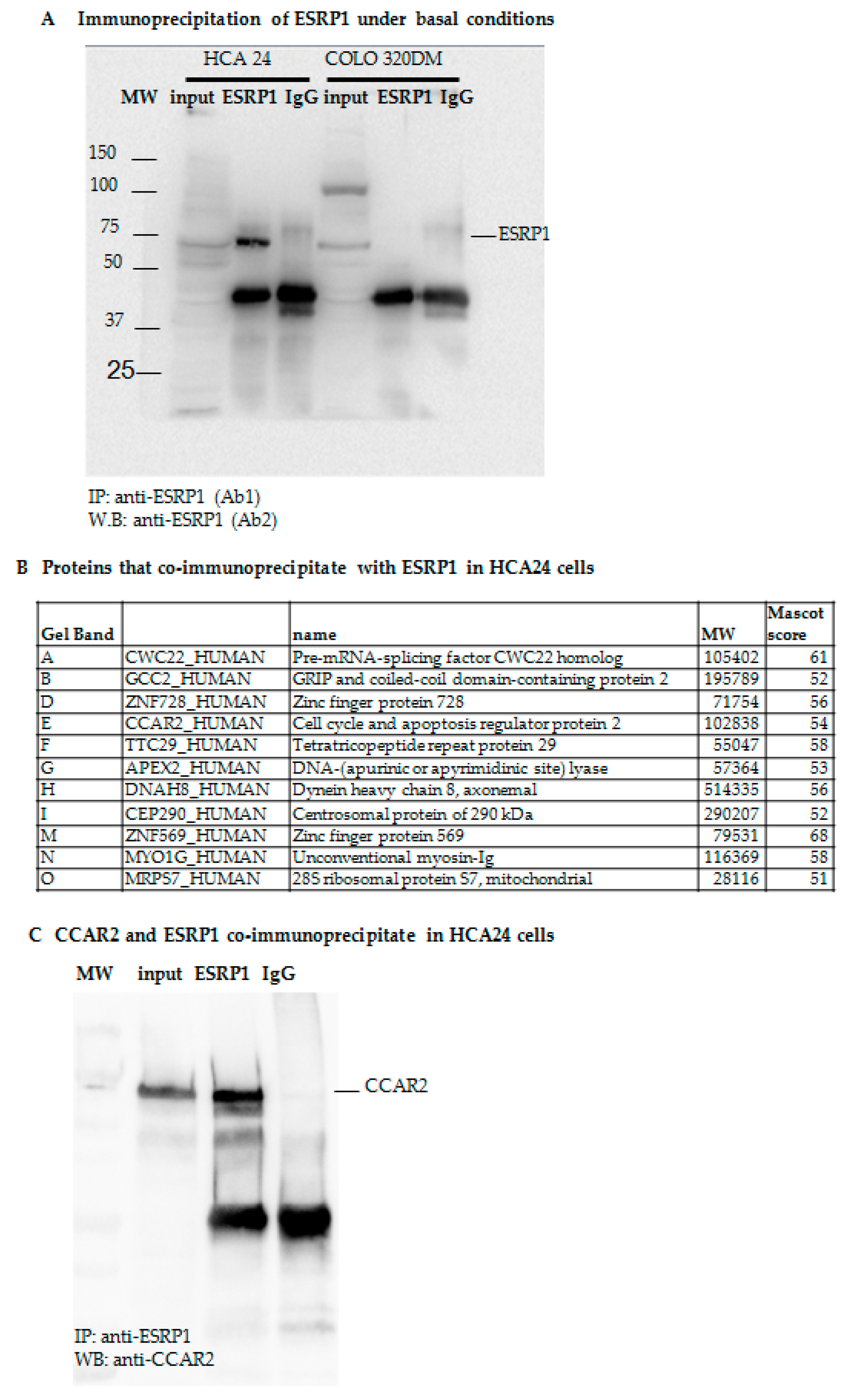
2.2. Proteomic Analysis of ESRP1-Interaction Network in CRC Cells
ESRP1-Interaction Network Ranked Correlated Sets Functional Characterization
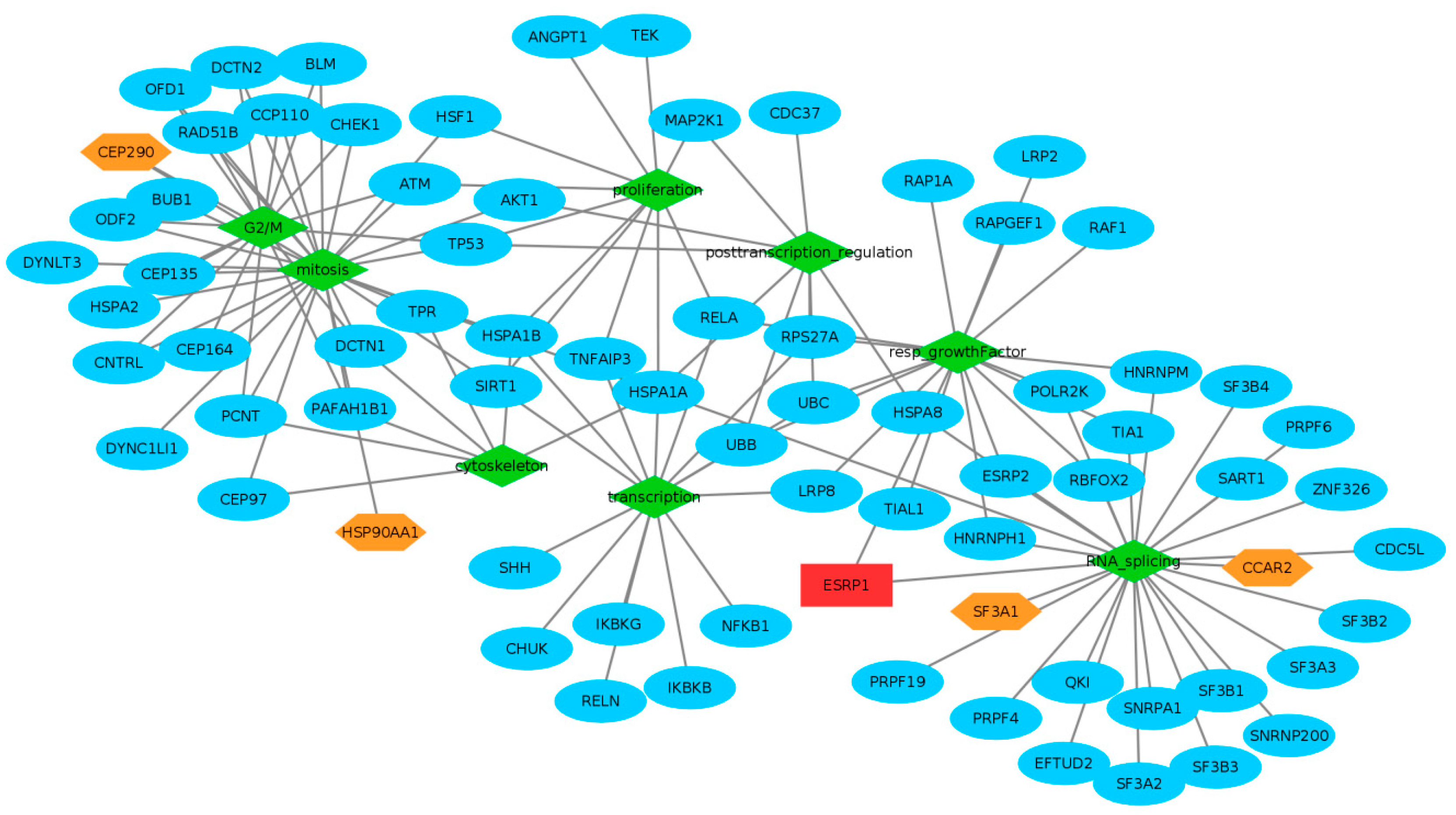
2.3. Global Tendencies of Enrichments
3. Discussion
4. Materials and Methods
4.1. CRC Cell Lines, Modulation of ESRP1 Expression
4.2. Chemicals and Reagents
4.3. RNA Extraction and Real-Time PCR
4.4. RNA-Immunoprecipitation
4.5. 2D Gel Electrophoresis and Mass Spectrometry Analysis
4.6. Western Blotting
4.7. Bioinformatics Analysis
4.7.1. Protein-Protein Interaction
4.7.2. Enrichment Analyses
4.7.3. Proteomics Data and Correlation Analysis
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ESRP1 | Epithelial Splicing Regulatory Protein 1 |
| CRC | ColoRectal Cancer |
| RIP | RNA Immuno Precipitation |
| RBP | RNA-Binding Protein |
| PPI | Protein-Protein Interaction |
| EMT | Epithelial to Mesenchymal Transition |
| GO | Gene Ontology |
| Scr | Scrambled |
| Sh | Short hairpin |
| MOI | Multiplicity of Infection |
| 2-DE | 2-dimensional gel electrophoresis |
| RCS | Ranked Correlated Sets |
References
- Dassi, E. Handshakes and fights: The regulatory interplay of rna-binding proteins. Front. Mol. Biosci. 2017, 4, 67. [Google Scholar] [CrossRef]
- Fagoonee, S.; Picco, G.; Orso, F.; Arrigoni, A.; Longo, D.L.; Forni, M.; Scarfo, I.; Cassenti, A.; Piva, R.; Cassoni, P.; et al. The rna-binding protein esrp1 promotes human colorectal cancer progression. Oncotarget 2017, 8, 10007–10024. [Google Scholar] [CrossRef]
- Pereira, B.; Billaud, M.; Almeida, R. Rna-binding proteins in cancer: Old players and new actors. Trends Cancer 2017, 3, 506–528. [Google Scholar] [CrossRef]
- Hong, S. Rna binding protein as an emerging therapeutic target for cancer prevention and treatment. J. Cancer Prev. 2017, 22, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, I.M.; Cheng, A.W.; Flytzanis, N.C.; Balsamo, M.; Condeelis, J.S.; Oktay, M.H.; Burge, C.B.; Gertler, F.B. An emt-driven alternative splicing program occurs in human breast cancer and modulates cellular phenotype. PLoS Genet. 2011, 7, e1002218. [Google Scholar] [CrossRef] [PubMed]
- Ueda, J.; Matsuda, Y.; Yamahatsu, K.; Uchida, E.; Naito, Z.; Korc, M.; Ishiwata, T. Epithelial splicing regulatory protein 1 is a favorable prognostic factor in pancreatic cancer that attenuates pancreatic metastases. Oncogene 2014, 33, 4485–4495. [Google Scholar] [CrossRef] [PubMed]
- Ishii, H.; Saitoh, M.; Sakamoto, K.; Kondo, T.; Katoh, R.; Tanaka, S.; Motizuki, M.; Masuyama, K.; Miyazawa, K. Epithelial splicing regulatory proteins 1 (ESRP1) and 2 (ESRP2) suppress cancer cell motility via different mechanisms. J. Biol. Chem. 2014, 289, 27386–27399. [Google Scholar] [CrossRef]
- Voena, C.; Varesio, L.M.; Zhang, L.; Menotti, M.; Poggio, T.; Panizza, E.; Wang, Q.; Minero, V.G.; Fagoonee, S.; Compagno, M.; et al. Oncogenic alk regulates emt in non-small cell lung carcinoma through repression of the epithelial splicing regulatory protein 1. Oncotarget 2016, 7, 33316–33330. [Google Scholar] [CrossRef]
- Yae, T.; Tsuchihashi, K.; Ishimoto, T.; Motohara, T.; Yoshikawa, M.; Yoshida, G.J.; Wada, T.; Masuko, T.; Mogushi, K.; Tanaka, H.; et al. Alternative splicing of cd44 mrna by esrp1 enhances lung colonization of metastatic cancer cell. Nat. Commun. 2012, 3, 883. [Google Scholar] [CrossRef]
- Marzese, D.M.; Liu, M.; Huynh, J.L.; Hirose, H.; Donovan, N.C.; Huynh, K.T.; Kiyohara, E.; Chong, K.; Cheng, D.; Tanaka, R.; et al. Brain metastasis is predetermined in early stages of cutaneous melanoma by cd44v6 expression through epigenetic regulation of the spliceosome. Pigment Cell Melanoma Res. 2015, 28, 82–93. [Google Scholar] [CrossRef]
- Jeong, H.M.; Han, J.; Lee, S.H.; Park, H.J.; Lee, H.J.; Choi, J.S.; Lee, Y.M.; Choi, Y.L.; Shin, Y.K.; Kwon, M.J. ESRP1 is overexpressed in ovarian cancer and promotes switching from mesenchymal to epithelial phenotype in ovarian cancer cells. Oncogenesis 2017, 6, e391. [Google Scholar] [CrossRef] [PubMed]
- Leontieva, O.V.; Ionov, Y. Rna-binding motif protein 35a is a novel tumor suppressor for colorectal cancer. Cell Cycle 2009, 8, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Guo, C.; Cao, J.; Burnett, J.; Yang, Z.; Ran, Y.; Sun, D. Over-expression of alpha-enolase as a prognostic biomarker in patients with pancreatic cancer. Int. J. Med Sci. 2017, 14, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.; Huhn, S.; Shen, Z. Complex roles of filamin-a mediated cytoskeleton network in cancer progression. Cell Biosci. 2013, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Principe, M.; Borgoni, S.; Cascione, M.; Chattaragada, M.S.; Ferri-Borgogno, S.; Capello, M.; Bulfamante, S.; Chapelle, J.; Di Modugno, F.; Defilippi, P.; et al. Alpha-enolase (eno1) controls alpha v/beta 3 integrin expression and regulates pancreatic cancer adhesion, invasion, and metastasis. J. Hematol. Oncol. 2017, 10, 16. [Google Scholar] [CrossRef]
- Capello, M.; Ferri-Borgogno, S.; Cappello, P.; Novelli, F. Alpha-enolase: A promising therapeutic and diagnostic tumor target. FEBS J. 2011, 278, 1064–1074. [Google Scholar] [CrossRef]
- Boulagnon-Rombi, C.; Schneider, C.; Leandri, C.; Jeanne, A.; Grybek, V.; Bressenot, A.M.; Barbe, C.; Marquet, B.; Nasri, S.; Coquelet, C.; et al. Lrp1 expression in colon cancer predicts clinical outcome. Oncotarget 2018, 9, 8849–8869. [Google Scholar] [CrossRef]
- Tian, J.; Liu, Y.; Zhu, B.; Tian, Y.; Zhong, R.; Chen, W.; Lu, X.; Zou, L.; Shen, N.; Qian, J.; et al. Sf3a1 and pancreatic cancer: New evidence for the association of the spliceosome and cancer. Oncotarget 2015, 6, 37750–37757. [Google Scholar] [CrossRef]
- Sun, S.; Liang, X.; Zhang, X.; Liu, T.; Shi, Q.; Song, Y.; Jiang, Y.; Wu, H.; Lu, X.; Pang, D. Phosphoglycerate kinase-1 is a predictor of poor survival and a novel prognostic biomarker of chemoresistance to paclitaxel treatment in breast cancer. Br. J. Cancer 2015, 112, 1332–1339. [Google Scholar] [CrossRef]
- Chao, C.C.; Wu, P.H.; Huang, H.C.; Chung, H.Y.; Chou, Y.C.; Cai, B.H.; Kannagi, R. Downregulation of mir-199a/b-5p is associated with gcnt2 induction upon epithelial-mesenchymal transition in colon cancer. FEBS Lett. 2017, 591, 1902–1917. [Google Scholar] [CrossRef]
- Majumdar, I.; Ahuja, V.; Paul, J. Altered expression of tumor necrosis factor alpha—Induced protein 3 correlates with disease severity in ulcerative colitis. Sci. Rep. 2017, 7, 9420. [Google Scholar] [CrossRef] [PubMed]
- Moser, C.; Lang, S.A.; Stoeltzing, O. Heat-shock protein 90 (hsp90) as a molecular target for therapy of gastrointestinal cancer. Anticancer Res. 2009, 29, 2031–2042. [Google Scholar] [PubMed]
- Lee, E.; Lee, D.H. Emerging roles of protein disulfide isomerase in cancer. BMB Rep. 2017, 50, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Rubie, C.; Kempf, K.; Hans, J.; Su, T.; Tilton, B.; Georg, T.; Brittner, B.; Ludwig, B.; Schilling, M. Housekeeping gene variability in normal and cancerous colorectal, pancreatic, esophageal, gastric and hepatic tissues. Mol. Cell. Probes 2005, 19, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Warzecha, C.C.; Sato, T.K.; Nabet, B.; Hogenesch, J.B.; Carstens, R.P. Esrp1 and Esrp2 are epithelial cell-type-specific regulators of fgfr2 splicing. Mol. Cell 2009, 33, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Bebee, T.W.; Park, J.W.; Sheridan, K.I.; Warzecha, C.C.; Cieply, B.W.; Rohacek, A.M.; Xing, Y.; Carstens, R.P. The splicing regulators esrp1 and esrp2 direct an epithelial splicing program essential for mammalian development. eLife 2015, 4, e08954. [Google Scholar] [CrossRef]
- Fagoonee, S.; Di Cunto, F.; Vozzi, D.; Volinia, S.; Pellegrino, M.; Gasparini, P.; Silengo, L.; Altruda, F.; Tolosano, E. Microarray and large-scale in silico—Based identification of genes functionally related to haptoglobin and/or hemopexin. DNA Cell Biol. 2006, 25, 323–330. [Google Scholar] [CrossRef]
- Chen, Z.H.; Jing, Y.J.; Yu, J.B.; Jin, Z.S.; Li, Z.; He, T.T.; Su, X.Z. Esrp1 induces cervical cancer cell g1-phase arrest via regulating cyclin a2 mrna stability. Int. J. Mol. Sci. 2019, 20, 3705. [Google Scholar] [CrossRef]
- Yao, J.; Caballero, O.L.; Huang, Y.; Lin, C.; Rimoldi, D.; Behren, A.; Cebon, J.S.; Hung, M.C.; Weinstein, J.N.; Strausberg, R.L.; et al. Altered expression and splicing of esrp1 in malignant melanoma correlates with epithelial-mesenchymal status and tumor-associated immune cytolytic activity. Cancer Immunol. Res. 2016, 4, 552–561. [Google Scholar] [CrossRef]
- Garcia-Cardenas, J.M.; Guerrero, S.; Lopez-Cortes, A.; Armendariz-Castillo, I.; Guevara-Ramirez, P.; Perez-Villa, A.; Yumiceba, V.; Zambrano, A.K.; Leone, P.E.; Paz, Y.M.C. Post-transcriptional regulation of colorectal cancer: A focus on rna-binding proteins. Front. Mol. Biosci. 2019, 6, 65. [Google Scholar] [CrossRef]
- Wang, J.; Mouradov, D.; Wang, X.; Jorissen, R.N.; Chambers, M.C.; Zimmerman, L.J.; Vasaikar, S.; Love, C.G.; Li, S.; Lowes, K.; et al. Colorectal cancer cell line proteomes are representative of primary tumors and predict drug sensitivity. Gastroenterology 2017, 153, 1082–1095. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, A.; Saitoh, M.; Miyazawa, K. Dual roles for epithelial splicing regulatory proteins 1 (esrp1) and 2 (esrp2) in cancer progression. Adv. Exp. Med. Biol. 2017, 925, 33–40. [Google Scholar] [PubMed]
- Kim, W.; Pyo, J.; Noh, B.J.; Jeong, J.W.; Lee, J.; Kim, J.E. Ccar2 negatively regulates il-8 production in cervical cancer cells. Oncotarget 2018, 9, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Close, P.; East, P.; Dirac-Svejstrup, A.B.; Hartmann, H.; Heron, M.; Maslen, S.; Chariot, A.; Soding, J.; Skehel, M.; Svejstrup, J.Q. Dbird complex integrates alternative mrna splicing with rna polymerase II transcript elongation. Nature 2012, 484, 386–389. [Google Scholar] [CrossRef] [PubMed]
- Restelli, M.; Magni, M.; Ruscica, V.; Pinciroli, P.; De Cecco, L.; Buscemi, G.; Delia, D.; Zannini, L. A novel crosstalk between ccar2 and akt pathway in the regulation of cancer cell proliferation. Cell Death Dis. 2016, 7, e2453. [Google Scholar] [CrossRef]
- Abdel-Wahab, O.; Levine, R. The spliceosome as an indicted conspirator in myeloid malignancies. Cancer Cell 2011, 20, 420–423. [Google Scholar] [CrossRef]
- Medico, E.; Russo, M.; Picco, G.; Cancelliere, C.; Valtorta, E.; Corti, G.; Buscarino, M.; Isella, C.; Lamba, S.; Martinoglio, B.; et al. The molecular landscape of colorectal cancer cell lines unveils clinically actionable kinase targets. Nat. Commun. 2015, 6, 7002. [Google Scholar] [CrossRef]
- Mandili, G.; Marini, C.; Carta, F.; Zanini, C.; Prato, M.; Khadjavi, A.; Turrini, F.; Giribaldi, G. Identification of phosphoproteins as possible differentiation markers in all-trans-retinoic acid-treated neuroblastoma cells. PLoS ONE 2011, 6, e18254. [Google Scholar] [CrossRef]
- STRING: Functional Protein Association Networks. Available online: https://string-db.org/ (accessed on 14 January 2020).
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. String v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. Clusterprofiler: An R package for comparing biological themes among gene clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef]
- GSEA/MSigDB—Broad Institute. Available online: http://software.broadinstitute.org/gsea/msigdb/index.jsp (accessed on 14 January 2020).





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ala, U.; Manco, M.; Mandili, G.; Tolosano, E.; Novelli, F.; Provero, P.; Altruda, F.; Fagoonee, S. Proteomics-Based Evidence for a Pro-Oncogenic Role of ESRP1 in Human Colorectal Cancer Cells. Int. J. Mol. Sci. 2020, 21, 575. https://doi.org/10.3390/ijms21020575
Ala U, Manco M, Mandili G, Tolosano E, Novelli F, Provero P, Altruda F, Fagoonee S. Proteomics-Based Evidence for a Pro-Oncogenic Role of ESRP1 in Human Colorectal Cancer Cells. International Journal of Molecular Sciences. 2020; 21(2):575. https://doi.org/10.3390/ijms21020575
Chicago/Turabian StyleAla, Ugo, Marta Manco, Giorgia Mandili, Emanuela Tolosano, Francesco Novelli, Paolo Provero, Fiorella Altruda, and Sharmila Fagoonee. 2020. "Proteomics-Based Evidence for a Pro-Oncogenic Role of ESRP1 in Human Colorectal Cancer Cells" International Journal of Molecular Sciences 21, no. 2: 575. https://doi.org/10.3390/ijms21020575
APA StyleAla, U., Manco, M., Mandili, G., Tolosano, E., Novelli, F., Provero, P., Altruda, F., & Fagoonee, S. (2020). Proteomics-Based Evidence for a Pro-Oncogenic Role of ESRP1 in Human Colorectal Cancer Cells. International Journal of Molecular Sciences, 21(2), 575. https://doi.org/10.3390/ijms21020575