PreS1 Mutations Alter the Large HBsAg Antigenicity of a Hepatitis B Virus Strain Isolated in Bangladesh




Abstract
1. Introduction
2. Results
2.1. Confirmation of HBV Infection/Replication
2.2. Successful Amplification and Cloning of Four Full Viral Genes
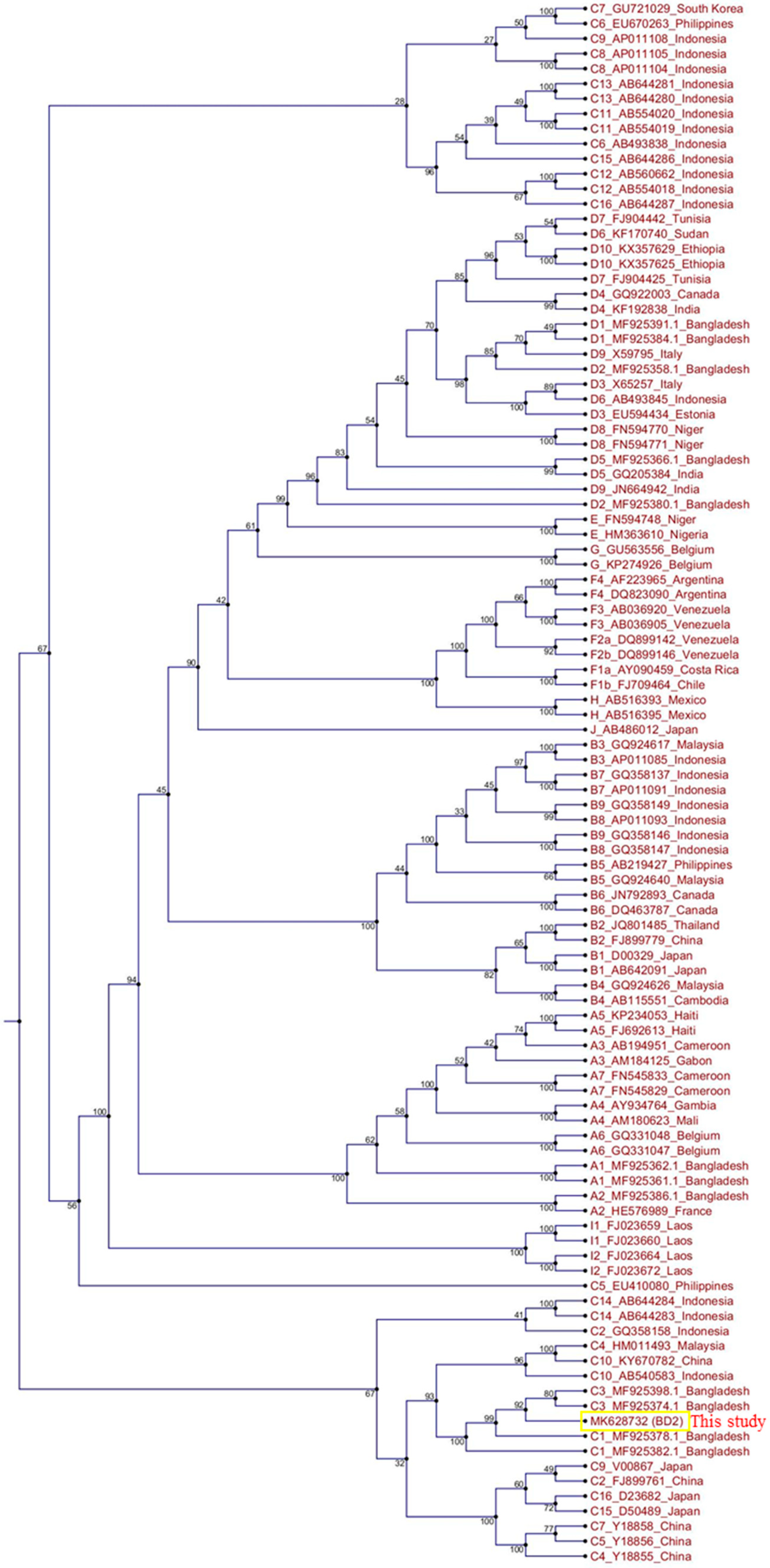
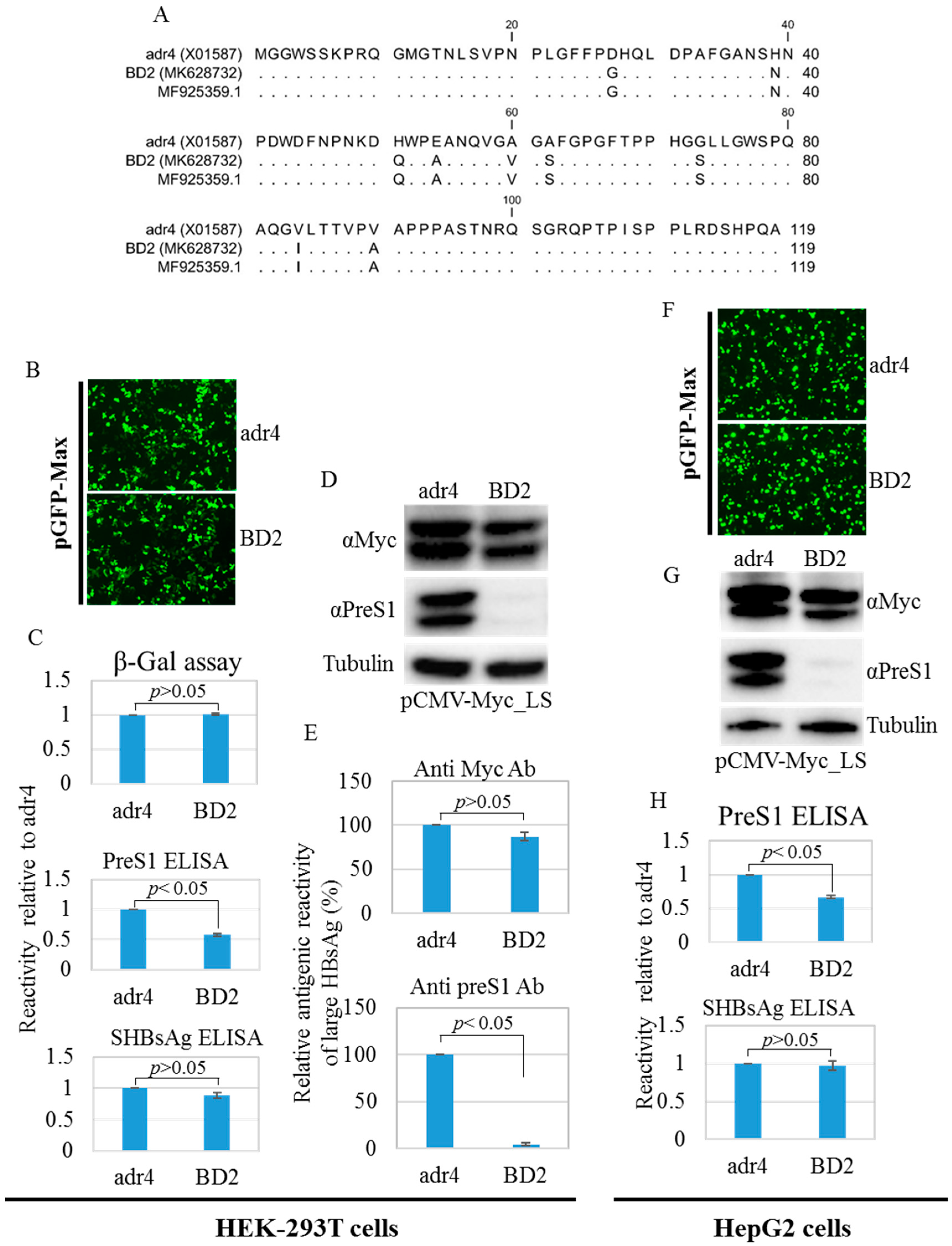
2.3. Characterization, Evolutionary Origin, and Mutational Analysis
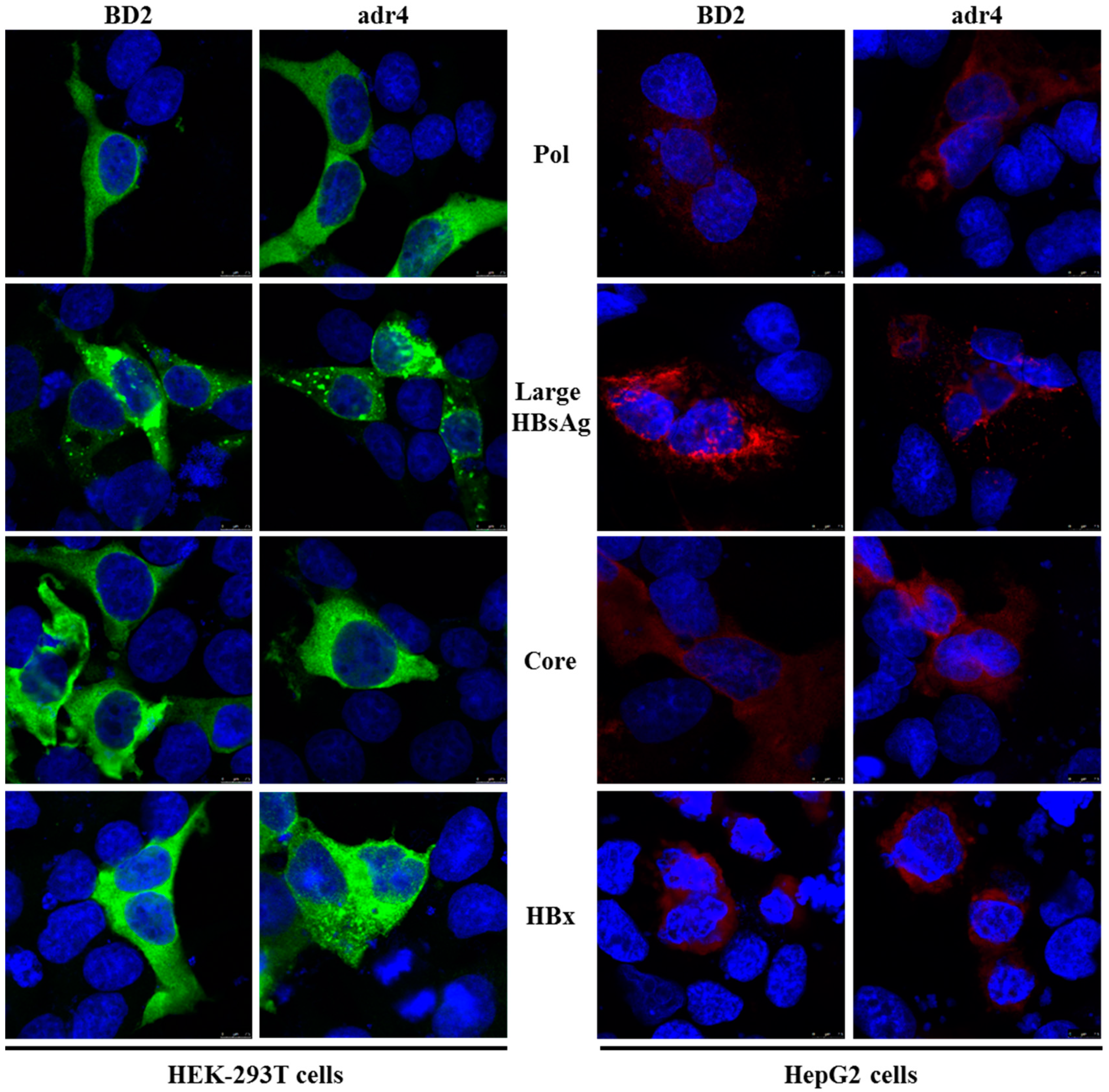
2.4. Expression Analysis of Different HBV Proteins
2.5. Antigenicity of Large HBsAg Is Altered due to Mutations in preS1 Region
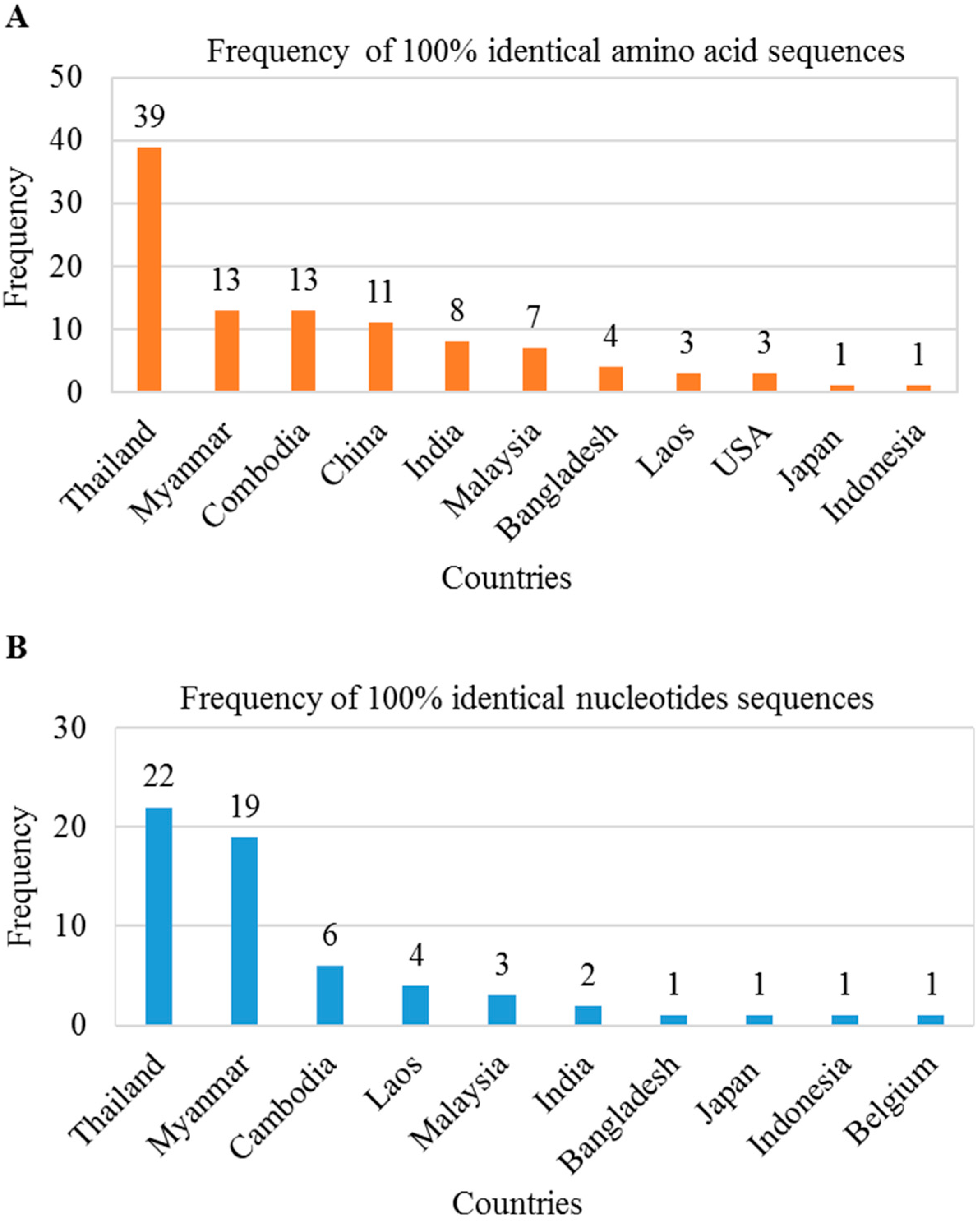
2.6. Silent Large HBsAg Containing Mutated HBV Is Circulating in Asiatic Countries
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. DNA Extraction, PCR Amplification, and Cloning
4.3. Real-Time Quantitative PCR
4.4. Sample Preparation for Sequencing
4.5. Sequence Annotation and Analysis and Phylogenetic and Mutational Analysis
4.6. Cell Lines and Transfection
4.7. Western Blot, Immunofluorescence Analysis, and ELISA
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HBV | Hepatitis B virus |
| HBsAg | Hepatitis B surface antigen |
| HCC | Hepatocellular carcinoma |
| NTCP | Sodium taurocholate co-transporting polypeptide |
| ORF | open reading frame |
References
- Locarnini, S. Molecular virology of hepatitis B virus. Semin. Liver Dis. 2004, 24, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Yano, Y.; Azuma, T.; Hayashi, Y. Variations and mutations in the hepatitis B virus genome and their associations with clinical characteristics. World J. Hepatol. 2015, 7, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, H.; Tsuda, F.; Sakugawa, H.; Sastrosoewignjo, R.I.; Imai, M.; Miyakawa, Y.; Mayumi, M. Typing hepatitis B virus by homology in nucleotide sequence: Comparison of surface antigen subtypes. J. Gen. Virol. 1988, 69, 2575–2583. [Google Scholar] [CrossRef] [PubMed]
- Glebe, D.; Urban, S. Viral and cellular determinants involved in hepadnaviral entry. World J. Gastroenterol. 2007, 13, 22–38. [Google Scholar] [CrossRef]
- Diab, A.; Foca, A.; Zoulim, F.; Durantel, D.; Andrisani, O. The diverse functions of the hepatitis B core/capsid protein (HBc) in the viral life cycle: Implications for the development of HBc-targeting antivirals. Antivir. Res. 2018, 149, 211–220. [Google Scholar] [CrossRef]
- Sohn, S.Y.; Kim, S.B.; Kim, J.; Ahn, B.Y. Negative regulation of hepatitis B virus replication by cellular Hsp40/DnaJ proteins through destabilization of viral core and X proteins. J. Gen. Virol. 2006, 87, 1883–1891. [Google Scholar] [CrossRef]
- Hayashi, M.; Imanaka-Yoshida, K.; Yoshida, T.; Wood, M.; Fearns, C.; Tatake, R.J.; Lee, J.D. A crucial role of mitochondrial Hsp40 in preventing dilated cardiomyopathy. Nat. Med. 2006, 12, 128–132. [Google Scholar] [CrossRef]
- Bouchard, M.J.; Schneider, R.J. The enigmatic X gene of hepatitis B virus. J. Virol. 2004, 78, 12725–12734. [Google Scholar] [CrossRef]
- Rawat, S.; Clippinger, A.J.; Bouchard, M.J. Modulation of apoptotic signaling by the hepatitis B virus X protein. Viruses 2012, 4, 2945–2972. [Google Scholar] [CrossRef]
- Kim, K.H.; Kim, N.D.; Seong, B.L. Discovery and development of anti-HBV agents and their resistance. Molecules 2010, 15, 5878–5908. [Google Scholar] [CrossRef]
- Tacke, F.; Liedtke, C.; Bocklage, S.; Manns, M.P.; Trautwein, C. CREB/PKA sensitive signalling pathways activate and maintain expression levels of the hepatitis B virus pre-S2/S promoter. Gut 2005, 54, 1309–1317. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.G.; Ueda, K. Investigation of a novel hepatitis B virus surface antigen (HBsAg) escape mutant affecting immunogenicity. PLoS ONE 2017, 12, e0167871. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Li, M.; Huang, B.; Liu, J.; Yu, L.; Chen, G. Detection of hepatitis B virus PreS1 antigen using a time-resolved fluoroimmunoassay. J. Immunoass. Immunochem. 2012, 33, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Ge, S.; Xiong, J.; Yan, Q.; Li, Z.; Hao, X.; Tian, D.; Niu, J.; Su, Z.; Chen, C.; et al. A novel immunoassay for PreS1 and/or core-related antigens for detection of HBsAg variants. J. Virol. Methods 2010, 168, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Ge, S.X.; Yan, Q.; Zhao, Y.; Xiong, J.H.; Zhang, J.; Xia, N.S. Establishment of a new combined enzyme immunoassay for detection of HBV preS1 and core antigens and the consistency with HBV DNA test. Bing Du Xue Bao Chin. J. Virol. 2007, 23, 252–257. [Google Scholar]
- Wei, J.; Wang, Y.Q.; Lu, Z.M.; Li, G.D.; Wang, Y.; Zhang, Z.C. Detection of anti-preS1 antibodies for recovery of hepatitis B patients by immunoassay. World J. Gastroenterol. 2002, 8, 276–281. [Google Scholar] [CrossRef]
- Lin, C.L.; Kao, J.H. Hepatitis B virus genotypes and variants. Cold Spring Harb. Perspect. Med. 2015, 5, a021436. [Google Scholar] [CrossRef]
- El-Mowafy, M.; Elgaml, A.; El-Mesery, M.; Elegezy, M. Molecular analysis of Hepatitis B virus sub-genotypes and incidence of preS1/preS2 region mutations in HBV-infected Egyptian patients from Mansoura. J. Med. Virol. 2017, 89, 1559–1566. [Google Scholar] [CrossRef]
- Lee, S.A.; Kim, K.J.; Kim, D.W.; Kim, B.J. Male-specific W4P/R mutation in the pre-S1 region of hepatitis B virus, increasing the risk of progression of liver diseases in chronic patients. J. Clin. Microbiol. 2013, 51, 3928–3936. [Google Scholar] [CrossRef]
- Yan, H.; Zhong, G.; Xu, G.; He, W.; Jing, Z.; Gao, Z.; Huang, Y.; Qi, Y.; Peng, B.; Wang, H.; et al. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. Elife 2012, 1, e00049. [Google Scholar] [CrossRef]
- Hu, W.G.; Wei, J.; Yang, X.X.; Xia, H.C.; Li, F.; Zhang, Z.C. Expression of overlapping PreS1 fragment recombinant proteins for the determination of immunogenic domains in HBsAg PreS1 region. Acta Biochim. Biophys. Sin. 2004, 36, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Bian, Y.; Zhang, Z.; Sun, Z.; Zhao, J.; Zhu, D.; Wang, Y.; Fu, S.; Guo, J.; Liu, L.; Su, L.; et al. Vaccines targeting preS1 domain overcome immune tolerance in hepatitis B virus carrier mice. Hepatology 2017, 66, 1067–1082. [Google Scholar] [CrossRef] [PubMed]
- Noor-E-Alam, S.M. Management of hepatocellular carcinoma: Bangladesh perspective. Euroasian J. Hepato gastroenterol. 2018, 8, 52. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Hakim, F.; Ahmed, M.; Ahsan, C.R.; Nessa, J.; Yasmin, M. Prevalence of genotypes and subtypes of hepatitis B viruses in Bangladeshi population. SpringerPlus 2016, 5, 278. [Google Scholar] [CrossRef] [PubMed]
- Munshi, S.U.; Tran, T.T.T.; Vo, T.N.T.; Tabassum, S.; Sultana, N.; Nguyen, T.H.; Jahan, M.; Le, C.N.; Baker, S.; Rahman, M. Molecular characterization of hepatitis B virus in Bangladesh reveals a highly recombinant population. PLoS ONE 2017, 12, e0188944. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.G.; Ueda, K. A meta-analysis on genetic variability of RT/HBsAg overlapping region of hepatitis B virus (HBV) isolates of Bangladesh. Infect. Agents Cancer 2019, 14, 33. [Google Scholar] [CrossRef]
- Tangkijvanich, P.; Mahachai, V.; Komolmit, P.; Fongsarun, J.; Theamboonlers, A.; Poovorawan, Y. Hepatitis B virus genotypes and hepatocellular carcinoma in Thailand. World J. Gastroenterol. 2005, 11, 2238–2243. [Google Scholar] [CrossRef]
- Orito, E.; Mizokami, M. Differences of HBV genotypes and hepatocellular carcinoma in Asian countries. Hepatol. Res. 2007, 37, S33–S35. [Google Scholar] [CrossRef]
- Chan, H.L.; Hui, A.Y.; Wong, M.L.; Tse, A.M.; Hung, L.C.; Wong, V.W.; Sung, J.J. Genotype C hepatitis B virus infection is associated with an increased risk of hepatocellular carcinoma. Gut 2004, 53, 1494–1498. [Google Scholar] [CrossRef]
- Shaha, M.; Hoque, S.A.; Rahman, S.R. Molecular epidemiology of hepatitis B virus isolated from Bangladesh. SpringerPlus 2016, 5, 1513. [Google Scholar] [CrossRef]
- Shaha, M.; Sarker, P.K.; Hossain, M.S.; Das, K.C.; Jahan, M.; Dey, S.K.; Tabassum, S.; Hashem, A.; Salimullah, M. Analysis of the complete genome of hepatitis B virus subgenotype C2 isolate NHB17965 from a HBV infected patient. F1000Research 2018, 7, 1023. [Google Scholar] [CrossRef] [PubMed]
- Sicheritz-Pontén, T.; Andersson, S.G. A phylogenomic approach to microbial evolution. Nucleic Acids Res. 2001, 29, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Luongo, M.; Critelli, R.; Grottola, A.; Gitto, S.; Bernabucci, V.; Bevini, M.; Vecchi, C.; Montagnani, G.; Villa, E. Acute hepatitis B caused by a vaccine-escape HBV strain in vaccinated subject: Sequence analysis and therapeutic strategy. J. Clin. Virol. 2015, 62, 89–91. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.M.; Jow, G.M.; Mu, S.C.; Chen, B.F. Naturally occurring hepatitis B virus B-cell and T-cell epitope mutants in hepatitis B vaccinated children. Sci. World J. 2013, 2013, 571875. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Xu, Y.; Zhang, Z.; Meng, Z.; Qin, L.; Lu, M.; Yang, D. The amino Acid residues at positions 120 to 123 are crucial for the antigenicity of hepatitis B surface antigen. J. Clin. Microbiol. 2007, 45, 2971–2978. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.M.; Li, X.H.; Mom, V.; Lu, Z.H.; Liao, X.W.; Han, Y.; Pichoud, C.; Gong, Q.M.; Zhang, D.H.; Zhang, Y.; et al. N-glycosylation mutations within hepatitis B virus surface major hydrophilic region contribute mostly to immune escape. J. Hepatol. 2014, 60, 515–522. [Google Scholar] [CrossRef]
- Huang, C.H.; Yuan, Q.; Chen, P.J.; Zhang, Y.L.; Chen, C.R.; Zheng, Q.B.; Yeh, S.H.; Yu, H.; Xue, Y.; Chen, Y.X.; et al. Influence of mutations in hepatitis B virus surface protein on viral antigenicity and phenotype in occult HBV strains from blood donors. J. Hepatol. 2012, 57, 720–729. [Google Scholar] [CrossRef]
- Shi, Y.; Wei, F.; Hu, D.; Li, Q.; Smith, D.; Li, N.; Chen, D. Mutations in the major hydrophilic region (MHR) of hepatitis B virus genotype C in North China. J. Med. Virol. 2012, 84, 1901–1906. [Google Scholar] [CrossRef]
- Dos Santos, M.; Pacheco, S.R.; Stocker, A.; Schinoni, M.I.; Parana, R.; Reis, M.G.; Silva, L.K. Mutations associated with drug resistance and prevalence of vaccine escape mutations in patients with chronic hepatitis B infection. J. Med. Virol. 2017, 89, 1811–1816. [Google Scholar] [CrossRef]
- Wu, C.C.; Chen, Y.S.; Cao, L.; Chen, X.W.; Lu, M.J. Hepatitis B virus infection: Defective surface antigen expression and pathogenesis. World J. Gastroenterol. 2018, 24, 3488–3499. [Google Scholar] [CrossRef]
- Zhou, L.Y.; Chen, E.Q.; Wang, M.L.; Chen, L.L.; Liu, C.P.; Zeng, F.; Tang, H. Biological characteristics comparison of HBV rtA181T mutants with truncated or substituted HBsAg expression in vitro and in vivo model systems. Sci. Rep. 2016, 6, 39260. [Google Scholar] [CrossRef] [PubMed]
- Li, M.S.; Lau, T.C.K.; Chan, S.K.P.; Wong, C.H.; Ng, P.K.S.; Sung, J.J.Y.; Chan, H.L.Y.; Tsui, S.K.W. The G1613A mutation in the HBV genome affects HBeAg expression and viral replication through altered core promoter activity. PLoS ONE 2011, 6, e21856. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.M.; Lee, S.Y.; Kim, B.J. Naturally occurring hepatitis B virus mutations leading to endoplasmic reticulum stress and their contribution to the progression of hepatocellular Carcinoma. Int. J. Mol. Sci. 2019, 20, 597. [Google Scholar] [CrossRef] [PubMed]
- Tassopoulos, N.C.; Papaevangelou, G.J.; Roumeliotou-Karayiannis, A.; Sjogren, M.H.; Engle, R.; Karpodini, E.; Purcell, R.H. Serologic markers of hepatitis B virus (HBV) and hepatitis D virus infection in carriers of hepatitis B surface antigen who are frequently exposed to HBV. Hepato-Gastroenterology 1986, 33, 151–154. [Google Scholar]
- Kao, J.H. Diagnosis of hepatitis B virus infection through serological and virological markers. Expert Rev. Gastroenterol. Hepatol. 2008, 2, 553–562. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Wu, C.C.; Chen, X.W.; Li, X.; Li, J.; Lu, M.J. Genetic variation of hepatitis B virus and its significance for pathogenesis. World J. Gastroenterol. 2016, 22, 126–144. [Google Scholar] [CrossRef]
- Yang, H.; Westland, C.E.; Delaney, W.E.; Heathcote, E.J.; Ho, V.; Fry, J.; Brosgart, C.; Gibbs, C.S.; Miller, M.D.; Xiong, S. Resistance surveillance in chronic hepatitis B patients treated with adefovir dipivoxil for up to 60 weeks. Hepatology 2002, 36, 464–473. [Google Scholar] [CrossRef]
- Choi, Y.M.; Lee, S.Y.; Kim, B.J. Naturally occurring hepatitis B virus reverse transcriptase mutations related to potential antiviral drug resistance and liver disease progression. World J. Gastroenterol. 2018, 24, 1708–1724. [Google Scholar] [CrossRef]
- Unchwaniwala, N.; Sherer, N.M.; Loeb, D.D. Hepatitis B virus polymerase localizes to the mitochondria, and its terminal protein domain contains the mitochondrial targeting signal. J. Virol. 2016, 90, 8705–8719. [Google Scholar] [CrossRef]
- Jazayeri, M.S.; Dornan, E.S.; Boner, W.; Fattovich, G.; Hadziyannis, S.; Carman, W.F. Intracellular distribution of hepatitis B virus core protein expressed in vitro depends on the sequence of the isolate and the serologic pattern. J. Infect. Dis. 2004, 189, 1634–1645. [Google Scholar] [CrossRef][Green Version]
- Patzer, E.J.; Nakamura, G.R.; Simonsen, C.C.; Levinson, A.D.; Brands, R. Intracellular assembly and packaging of hepatitis B surface antigen particles occur in the endoplasmic reticulum. J. Virol. 1986, 58, 884–892. [Google Scholar] [CrossRef] [PubMed]
- Patient, R.; Hourioux, C.; Sizaret, P.-Y.; Trassard, S.; Sureau, C.; Roingeard, P. Hepatitis B virus subviral envelope particle morphogenesis and intracellular trafficking. J. Virol. 2007, 81, 3842–3851. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Candotti, D.; Allain, J.P. Specific amino acid substitutions in the S protein prevent its excretion in vitro and may contribute to occult hepatitis B virus infection. J. Virol. 2013, 87, 7882–7892. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.A.; Lee, C. Hepatitis B virus X gene and hepatocarcinogenesis. J. Gastroenterol. 2011, 46, 974–990. [Google Scholar] [CrossRef]
- Keasler, V.V.; Hodgson, A.J.; Madden, C.R.; Slagle, B.L. Hepatitis B virus HBx protein localized to the nucleus restores HBx-deficient virus replication in HepG2 cells and in vivo in hydrodynamically-injected mice. Virology 2009, 390, 122–129. [Google Scholar] [CrossRef]
- Yin, J.; Xie, J.; Zhang, H.; Shen, Q.; Han, L.; Lu, W.; Han, Y.; Li, C.; Ni, W.; Wang, H.; et al. Significant association of different preS mutations with hepatitis B-related cirrhosis or hepatocellular carcinoma. J. Gastroenterol. 2010, 45, 1063–1071. [Google Scholar] [CrossRef]
- Chen, P.J.; Wu, T.C. One step closer to an experimental infection system for Hepatitis B Virus?—The identification of sodium taurocholate cotransporting peptide as a viral receptor. Cell Biosci. 2013, 3, 2. [Google Scholar] [CrossRef]
- Meier, A.; Mehrle, S.; Weiss, T.S.; Mier, W.; Urban, S. Myristoylated PreS1-domain of the hepatitis B virus L-protein mediates specific binding to differentiated hepatocytes. Hepatology 2013, 58, 31–42. [Google Scholar] [CrossRef]
- Dobrica, M.O.; Lazar, C.; Paruch, L.; Skomedal, H.; Steen, H.; Haugslien, S.; Tucureanu, C.; Caras, I.; Onu, A.; Ciulean, S.; et al. A novel chimeric Hepatitis B virus S/preS1 antigen produced in mammalian and plant cells elicits stronger humoral and cellular immune response than the standard vaccine-constituent, S protein. Antivir. Res. 2017, 144, 256–265. [Google Scholar] [CrossRef]
- Pourkarim, M.R.; Amini-Bavil-Olyaee, S.; Verbeeck, J.; Lemey, P.; Zeller, M.; Rahman, M.; Maes, P.; Nevens, F.; Van Ranst, M. Molecular evolutionary analysis and mutational pattern of full-length genomes of hepatitis B virus isolated from Belgian patients with different clinical manifestations. J. Med. Virol. 2010, 82, 379–389. [Google Scholar] [CrossRef]
- Hellström, U.B.; Madalinski, K.; Sylvan, S.P. PreS1 epitope recognition in newborns after vaccination with the third-generation Sci-B-Vac vaccine and their relation to the antibody response to hepatitis B surface antigen. Virol. J. 2009, 6, 7. [Google Scholar]
- Hu, W.G.; Wei, J.; Xia, H.C.; Yang, X.X.; Li, F.; Li, G.D.; Wang, Y.; Zhang, Z.C. Identification of the immunogenic domains in HBsAg preS1 region using overlapping preS1 fragment fusion proteins. World J. Gastroenterol. 2005, 11, 2088–2094. [Google Scholar] [CrossRef]
- Hossain, M.G.; Ohsaki, E.; Honda, T.; Ueda, K. Importance of promyelocytic leukema protein (PML) for Kaposi’s sarcoma-associated herpesvirus lytic replication. Front. Microbiol. 2018, 9, 2324. [Google Scholar] [CrossRef] [PubMed]
- Magnius, L.O.; Norder, H. Subtypes, genotypes and molecular epidemiology of the hepatitis B virus as reflected by sequence variability of the S-gene. Intervirology 1995, 38, 24–34. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes or Purpose | Primer Names | Sequences (5′–3′) |
|---|---|---|
| P | EcoRI-Pol-1 | GTGGAATTCGGATGCCCCTATCTTATCAACAC |
| Pol-stop-SalI | CACGTCGACTCACGGTGGTCTCCATGCGAC | |
| S | EcoRI-HBs preS1 | GTGGAATTCGGATGGGAGGTTGGTCTTCCAAAC |
| HBs-stop-SalI | CACGTCGACTTAAATGTATACCCAAAGAC | |
| C | EcoRI-Core-1 | GTGGAATTCGGATGGACATTGACCCGTATAAAG |
| Core-stop-SalI | CACGTCGACTAACATTGAGATTCCCGAG | |
| X | EcoRI-HBx-1 | GTGGAATTCGGATGGCTGCTAGGGTGTGCTG |
| HBx-stop-SalI | CACGTCGACTTAGGCAGAGGTGAAAAAGTTG | |
| pCMV_Myc plasmid | CMV Pro | ACGGTGGGAGGTCTATATAAGC |
| Real-time quantitative PCR | HBs F2 | CTTCATCCTGCTGCTATGCCT |
| HBsR2 | AAAGCCCAGGATGATGGGAT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, M.G.; Mahmud, M.M.; Nazir, K.H.M.N.H.; Ueda, K. PreS1 Mutations Alter the Large HBsAg Antigenicity of a Hepatitis B Virus Strain Isolated in Bangladesh. Int. J. Mol. Sci. 2020, 21, 546. https://doi.org/10.3390/ijms21020546
Hossain MG, Mahmud MM, Nazir KHMNH, Ueda K. PreS1 Mutations Alter the Large HBsAg Antigenicity of a Hepatitis B Virus Strain Isolated in Bangladesh. International Journal of Molecular Sciences. 2020; 21(2):546. https://doi.org/10.3390/ijms21020546
Chicago/Turabian StyleHossain, Md. Golzar, Md. Muket Mahmud, K. H. M. Nazmul Hussain Nazir, and Keiji Ueda. 2020. "PreS1 Mutations Alter the Large HBsAg Antigenicity of a Hepatitis B Virus Strain Isolated in Bangladesh" International Journal of Molecular Sciences 21, no. 2: 546. https://doi.org/10.3390/ijms21020546
APA StyleHossain, M. G., Mahmud, M. M., Nazir, K. H. M. N. H., & Ueda, K. (2020). PreS1 Mutations Alter the Large HBsAg Antigenicity of a Hepatitis B Virus Strain Isolated in Bangladesh. International Journal of Molecular Sciences, 21(2), 546. https://doi.org/10.3390/ijms21020546