Antagonistic Autoantibodies to Insulin-Like Growth Factor-1 Receptor Associate with Poor Physical Strength

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
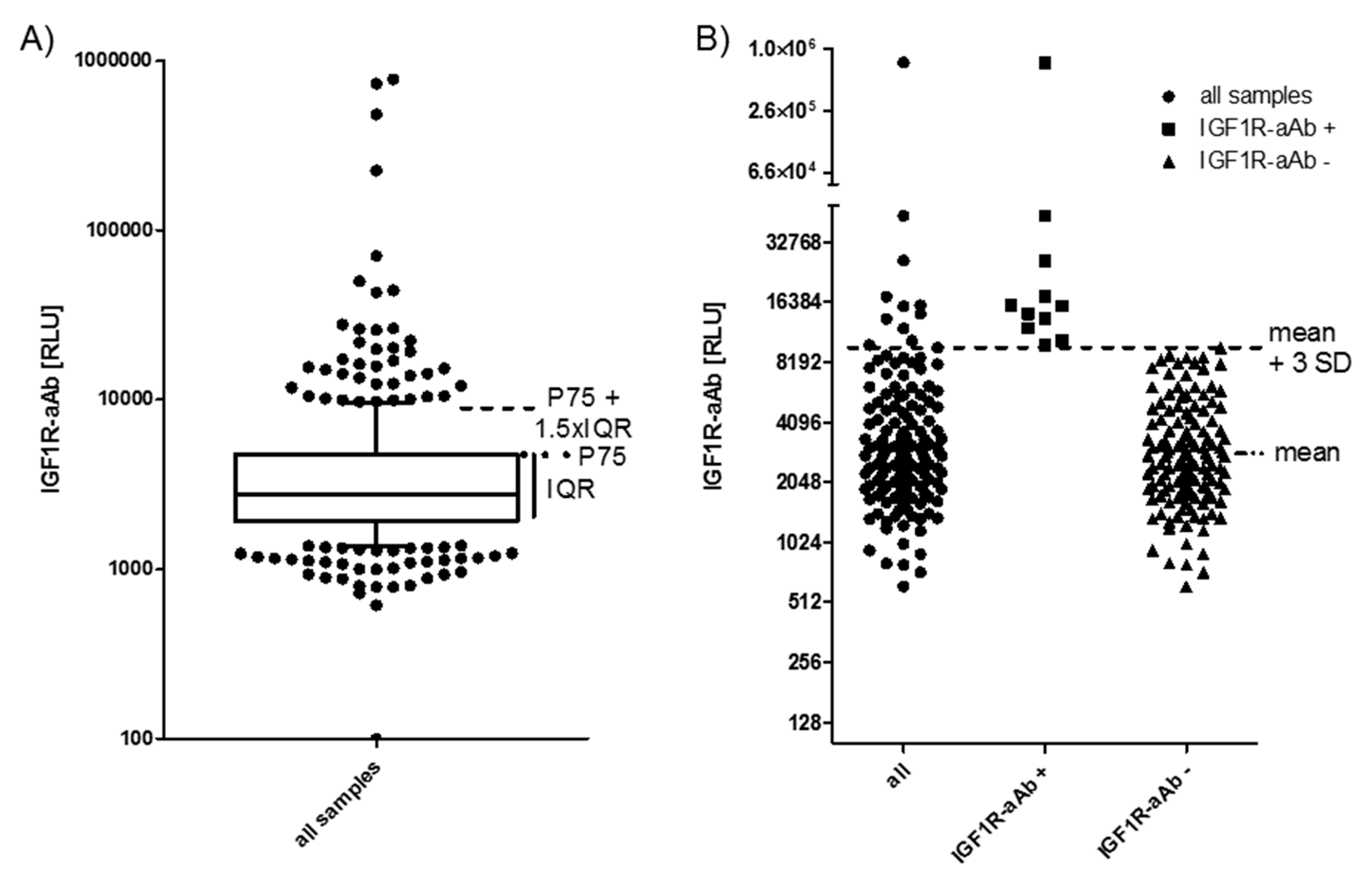
2.1. Prevalence of IGF1R-aAb in Overweight Young Subjects
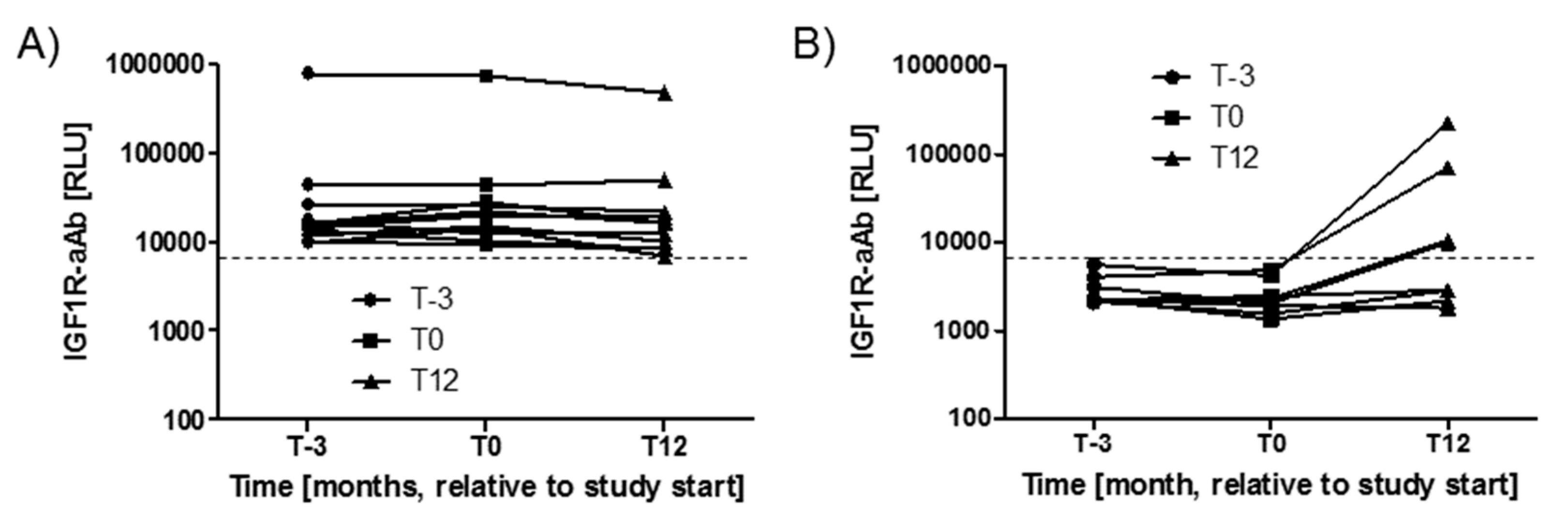
2.2. Stability of IGF1R-aAb Concentrations during the Study Period
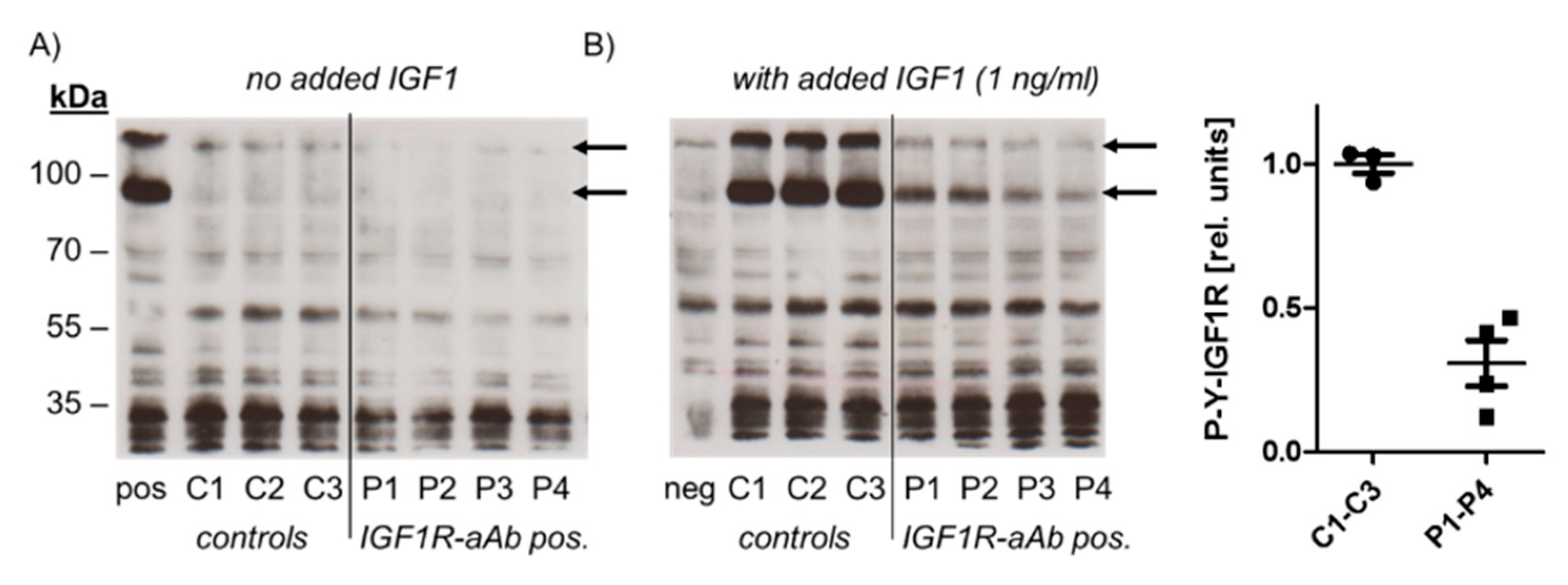
2.3. Molecular Characterization of IGF1R-Ab
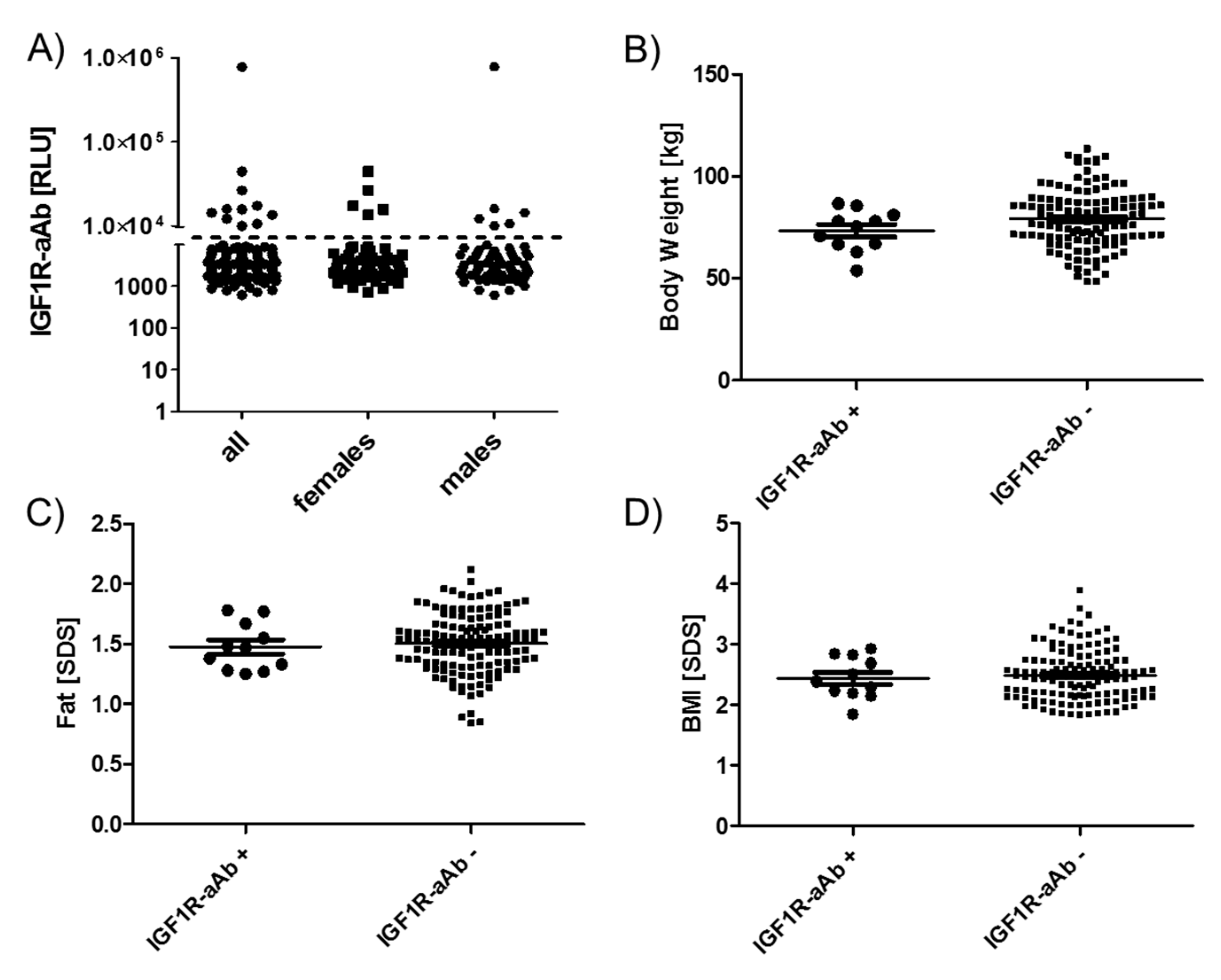
2.4. Comparison of Anthropometric Parameters in Relation to IGF1R-Ab
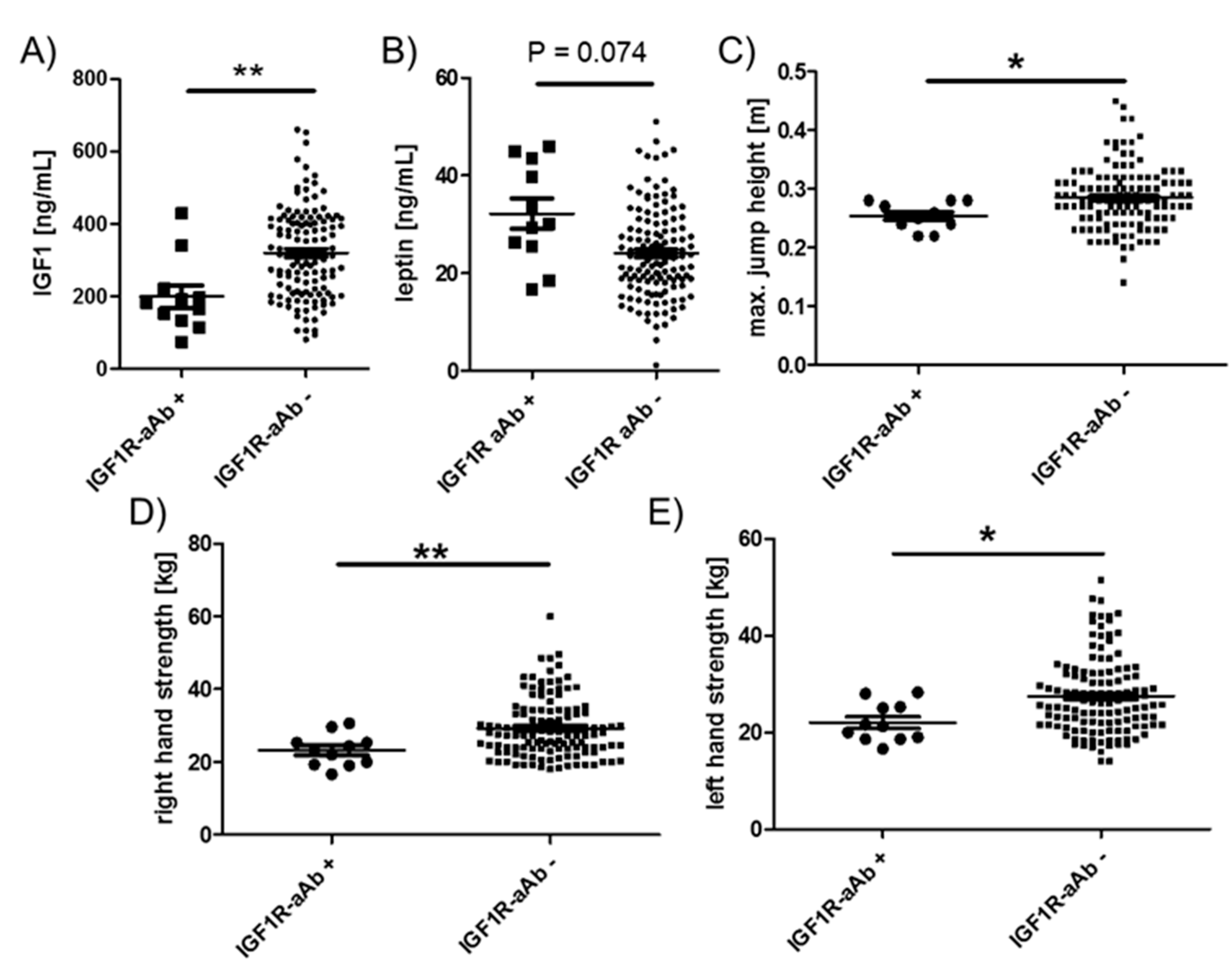
2.5. Comparison of Endocrine and Physical Strength Parameters in Relation to IGF1R-aAb
3. Discussion
4. Materials and Methods
4.1. Recombinant Expression of IGF1R as Luciferase Fusion Protein
4.2. Molecular Characterization of IGF1R Autoantibodies
4.3. Serum Samples from Children Enrolled in the Intervention Trial
4.4. Quantification of IGF1R Autoantibodies
4.5. Anthropometric Analyses
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BRCA1 | BReast CAncer 1 |
| BRCA2 | BReast CAncer 2 |
| PALB2 | PArtner and Localizer of BRCA2 |
| PDAC | pancreatic ductal adenocarcinoma |
| TMAs | Tissue microarrays |
| IHC | immunohistochemistry |
| OS | Overall survival |
| FANCN | Fanconi anemia complementation |
| TCGA | The Cancer Genome Atlas |
| GSEA | gene set enrichment analysis |
| CIs | confidence intervals |
| HR | hazard ratio |
| T | primary tumor site |
| N | regional lymph node involvement |
| NES | normalized enrichment score |
| EMT | epithelial to mesenchymal transition |
References
- Cabail, M.Z.; Li, S.; Lemmon, E.; Bowen, M.E.; Hubbard, S.R.; Miller, W.T. The insulin and IGF1 receptor kinase domains are functional dimers in the activated state. Nat. Commun. 2015, 6, 6406. [Google Scholar] [CrossRef] [PubMed]
- Blakesley, V.A.; Scrimgeour, A.; Esposito, D.; Le Roith, D. Signaling via the insulin-like growth factor-I receptor: Does it differ from insulin receptor signaling? Cytokine Growth Factor Rev. 1996, 7, 153–159. [Google Scholar] [CrossRef]
- Baker, J.; Liu, J.P.; Robertson, E.J.; Efstratiadis, A. Role of insulin-like growth factors in embryonic and postnatal growth. Cell 1993, 75, 73–82. [Google Scholar] [CrossRef]
- Abuzzahab, M.J.; Schneider, A.; Goddard, A.; Grigorescu, F.; Lautier, C.; Keller, E.; Kiess, W.; Klammt, J.; Kratzsch, J.; Osgood, D.; et al. IGF-I receptor mutations resulting in intrauterine and postnatal growth retardation. N. Engl. J. Med. 2003, 349, 2211–2222. [Google Scholar] [CrossRef]
- Walenkamp, M.J.; Losekoot, M.; Wit, J.M. Molecular IGF-1 and IGF-1 receptor defects: From genetics to clinical management. Endocr Dev. 2013, 24, 128–137. [Google Scholar]
- LeRoith, D.; Roberts, C.T., Jr. The insulin-like growth factor system and cancer. Cancer Lett. 2003, 195, 127–137. [Google Scholar] [CrossRef]
- McLachlan, S.M.; Rapoport, B. Thyrotropin-blocking autoantibodies and thyroid-stimulating autoantibodies: Potential mechanisms involved in the pendulum swinging from hypothyroidism to hyperthyroidism or vice versa. Thyroid 2013, 23, 14–24. [Google Scholar] [CrossRef]
- Diana, T.; Krause, J.; Olivo, P.D.; Konig, J.; Kanitz, M.; Decallonne, B.; Kahaly, G.J. Prevalence and clinical relevance of thyroid stimulating hormone receptor-blocking antibodies in autoimmune thyroid disease. Clin. Exp. Immunol. 2017, 189, 304–309. [Google Scholar] [CrossRef]
- Eckstein, A.K.; Plicht, M.; Lax, H.; Neuhauser, M.; Mann, K.; Lederbogen, S.; Heckmann, C.; Esser, J.; Morgenthaler, N.G. Thyrotropin receptor autoantibodies are independent risk factors for Graves’ ophthalmopathy and help to predict severity and outcome of the disease. J. Clin. Endocrinol. Metab. 2006, 91, 3464–3470. [Google Scholar] [CrossRef]
- Wiersinga, W.M. Autoimmunity in Graves’ ophthalmopathy: The result of an unfortunate marriage between TSH receptors and IGF-1 receptors? J. Clin. Endocrinol. Metab. 2011, 96, 2386–2394. [Google Scholar] [CrossRef]
- Bahn, R.S. Graves’ ophthalmopathy. N. Engl. J. Med. 2010, 362, 726–738. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.J. TSH-receptor-expressing fibrocytes and thyroid-associated ophthalmopathy. Nat. Rev. Endocrinol. 2015, 11, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Krieger, C.C.; Place, R.F.; Bevilacqua, C.; Marcus-Samuels, B.; Abel, B.S.; Skarulis, M.C.; Kahaly, G.J.; Neumann, S.; Gershengorn, M.C. TSH/IGF-1 Receptor Cross Talk in Graves’ Ophthalmopathy Pathogenesis. J. Clin. Endocrinol. Metab. 2016, 101, 2340–2347. [Google Scholar] [CrossRef] [PubMed]
- Marcus-Samuels, B.; Krieger, C.C.; Boutin, A.; Kahaly, G.J.; Neumann, S.; Gershengorn, M.C. Evidence That Graves’ Ophthalmopathy Immunoglobulins Do Not Directly Activate IGF-1 Receptors. Thyroid 2018, 28, 650–655. [Google Scholar] [CrossRef]
- Minich, W.B.; Dehina, N.; Welsink, T.; Schwiebert, C.; Morgenthaler, N.G.; Kohrle, J.; Eckstein, A.; Schomburg, L. Autoantibodies to the IGF1 receptor in Graves’ orbitopathy. J. Clin. Endocrinol. Metab. 2013, 98, 752–760. [Google Scholar] [CrossRef]
- Bau, A.M.; Ernert, A.; Krude, H.; Wiegand, S. Hormonal regulatory mechanisms in obese children and adolescents after previous weight reduction with a lifestyle intervention: Maintain-paediatric part-a RCT from 2009-15. BMC Obes. 2016, 3, 29. [Google Scholar] [CrossRef]
- Beeson, P.B. Age and Sex Associations of 40 Autoimmune-Diseases. Am. J. Med. 1994, 96, 457–462. [Google Scholar] [CrossRef]
- Marino, M.; Rotondo Dottore, G.; Ionni, I.; Lanzolla, G.; Sabini, E.; Ricci, D.; Sframeli, A.; Mazzi, B.; Menconi, F.; Latrofa, F.; et al. Serum antibodies against the insulin-like growth factor-1 receptor (IGF-1R) in Graves’ disease and Graves’ orbitopathy. J. Endocrinol. Investig. 2019, 42, 471–480. [Google Scholar] [CrossRef]
- Chazenbalk, G.D.; Pichurin, P.; McLachlan, S.M.; Rapoport, B. A direct binding assay for thyrotropin receptor autoantibodies. Thyroid 1999, 9, 1057–1061. [Google Scholar] [CrossRef]
- Giuliani, C.; Saji, M.; Bucci, I.; Napolitano, G. Bioassays for TSH Receptor Autoantibodies, from FRTL-5 Cells to TSH Receptor-LH/CG Receptor Chimeras: The Contribution of Leonard D. Kohn. Front. Endocrinol. 2016, 7, 103. [Google Scholar] [CrossRef]
- Tozzoli, R.; Bagnasco, M.; Giavarina, D.; Bizzaro, N. TSH receptor autoantibody immunoassay in patients with Graves’ disease: Improvement of diagnostic accuracy over different generations of methods. Systematic review and meta-analysis. Autoimmun. Rev. 2012, 12, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Haberland, A.; Muller, J.; Wallukat, G.; Wenzel, K. Antigen-free control wells in an ELISA set-up for the determination of autoantibodies against G protein-coupled receptors-a requisite for correct data evaluation. Anal. Bioanal. Chem. 2018, 410, 5101–5105. [Google Scholar] [CrossRef] [PubMed]
- Boege, F.; Westenfeld, R.; Jahns, R. Beta1AAb Determined by Peptide ELISA: A Signal in the Noise? J. Am. Coll. Cardiol. 2017, 70, 807–808. [Google Scholar] [CrossRef] [PubMed]
- Diana, T.; Kanitz, M.; Lehmann, M.; Li, Y.; Olivo, P.D.; Kahaly, G.J. Standardization of a bioassay for thyrotropin receptor stimulating autoantibodies. Thyroid 2015, 25, 169–175. [Google Scholar] [CrossRef]
- Joshi-Barr, S.; Haberland, A.; Bartel, S.; Muller, J.; Choi, T.; Wallukat, G. High throughput bioassay for beta1-adrenoceptor autoantibody detection. Int. J. Cardiol. 2016, 219, 98–104. [Google Scholar] [CrossRef]
- Jahns, R.; Boivin, V.; Schwarzbach, V.; Ertl, G.; Lohse, M.J. Pathological autoantibodies in cardiomyopathy. Autoimmunity 2008, 41, 454–461. [Google Scholar] [CrossRef]
- Vicco, M.H.; Ferini, F.; Rodeles, L.; Cardona, P.; Bontempi, I.; Lioi, S.; Beloscar, J.; Nara, T.; Marcipar, I.; Bottasso, O.A. Assessment of Cross-reactive Host-pathogen Antibodies in Patients with Different Stages of Chronic Chagas Disease. Rev. Esp. De Cardiol. 2013, 66, 791–796. [Google Scholar] [CrossRef]
- Klubo-Gwiezdzinska, J.; Lange, M.; Cochran, E.; Semple, R.K.; Gewert, C.; Brown, R.J.; Gorden, P. Combined Immunosuppressive Therapy Induces Remission in Patients with Severe Type B Insulin Resistance: A Prospective Cohort Study. Diabetes Care 2018, 41, 2353–2360. [Google Scholar] [CrossRef]
- Chikani, V.; Ho, K.K. Action of GH on skeletal muscle function: Molecular and metabolic mechanisms. J. Mol. Endocrinol. 2014, 52, R107–R123. [Google Scholar] [CrossRef]
- Walenkamp, M.J.; van der Kamp, H.J.; Pereira, A.M.; Kant, S.G.; van Duyvenvoorde, H.A.; Kruithof, M.F.; Breuning, M.H.; Romijn, J.A.; Karperien, M.; Wit, J.M. A variable degree of intrauterine and postnatal growth retardation in a family with a missense mutation in the insulin-like growth factor I receptor. J. Clin. Endocrinol. Metab. 2006, 91, 3062–3070. [Google Scholar] [CrossRef]
- Labarta, J.I.; Barrio, E.; Audi, L.; Fernandez-Cancio, M.; Andaluz, P.; de Arriba, A.; Puga, B.; Calvo, M.T.; Mayayo, E.; Carrascosa, A.; et al. Familial short stature and intrauterine growth retardation associated with a novel mutation in the IGF-I receptor (IGF1R) gene. Clin. Endocrinol. 2013, 78, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Laron, Z.; Kauli, R.; Lapkina, L.; Werner, H. IGF-I deficiency, longevity and cancer protection of patients with Laron syndrome. Mutat. Res. Rev. Mutat. Res. 2017, 772, 123–133. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwiebert, C.; Kühnen, P.; Becker, N.-P.; Welsink, T.; Keller, T.; Minich, W.B.; Wiegand, S.; Schomburg, L. Antagonistic Autoantibodies to Insulin-Like Growth Factor-1 Receptor Associate with Poor Physical Strength. Int. J. Mol. Sci. 2020, 21, 463. https://doi.org/10.3390/ijms21020463
Schwiebert C, Kühnen P, Becker N-P, Welsink T, Keller T, Minich WB, Wiegand S, Schomburg L. Antagonistic Autoantibodies to Insulin-Like Growth Factor-1 Receptor Associate with Poor Physical Strength. International Journal of Molecular Sciences. 2020; 21(2):463. https://doi.org/10.3390/ijms21020463
Chicago/Turabian StyleSchwiebert, Christian, Peter Kühnen, Niels-Peter Becker, Tim Welsink, Theresa Keller, Waldemar B. Minich, Susanna Wiegand, and Lutz Schomburg. 2020. "Antagonistic Autoantibodies to Insulin-Like Growth Factor-1 Receptor Associate with Poor Physical Strength" International Journal of Molecular Sciences 21, no. 2: 463. https://doi.org/10.3390/ijms21020463
APA StyleSchwiebert, C., Kühnen, P., Becker, N.-P., Welsink, T., Keller, T., Minich, W. B., Wiegand, S., & Schomburg, L. (2020). Antagonistic Autoantibodies to Insulin-Like Growth Factor-1 Receptor Associate with Poor Physical Strength. International Journal of Molecular Sciences, 21(2), 463. https://doi.org/10.3390/ijms21020463