Micro-RNAs: Crossroads between the Exposure to Environmental Particulate Pollution and the Obstructive Pulmonary Disease




Abstract
1. Introduction
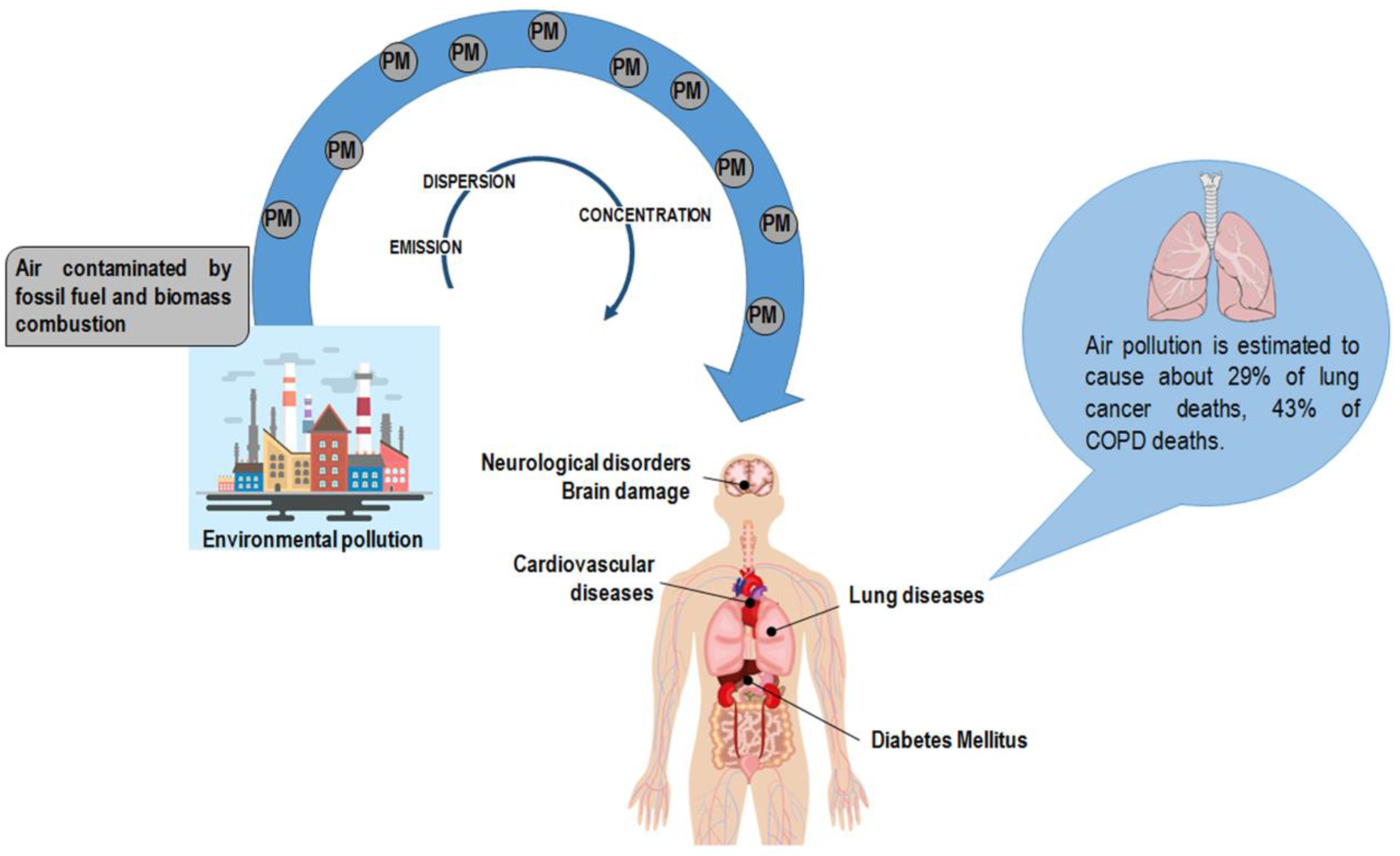
2. Air Pollution: The Role Played by Particulate Matter and Heavy Metals
3. The Exposome Concept
4. miRNA Response to Environmental Air Pollutants (PMs and Heavy Metals)
4.1. Particulate Matter Exposure and miRNAs
4.2. Arsenic Exposure and miRNAs
4.3. Cadmium Exposure and miRNAs
4.4. Lead Exposure and miRNAs
5. Co-Exposure Strategy in Environmental-Based Research and the Contribution of miRNAs
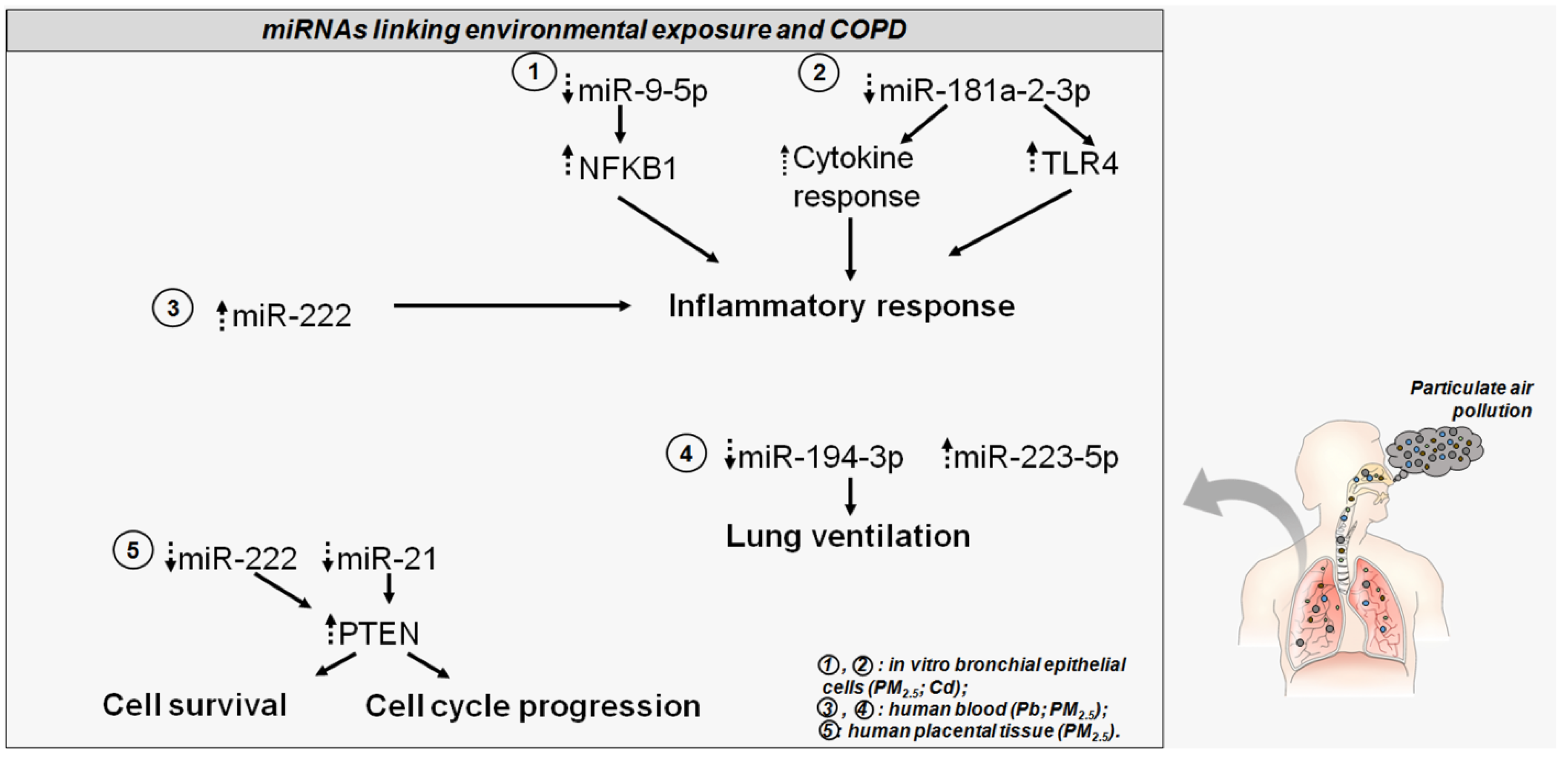
6. Air Pollution and COPD: The Role of miRNAs
7. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| As | Arsenic |
| cAMP | Adenylyl cyclase |
| Cd | Cadmium |
| CFTR | Cystic Fibrosis Transmembrane Conductance Regulator |
| COPD | Chronic Obstructive Pulmonary Disease |
| E2F1 | E2F transcription factor 1 |
| GCL | Glutamate–Cysteine Ligase |
| GCLC | Glutamate–Cysteine Ligase Catalytic Subunit |
| GCLM | Glutamate–Cysteine Ligase Modifier Subunit |
| GSH | Glutathione |
| HBECs | Human Bronchial Epithelial Cells |
| KRAS | Kirsten rat sarcoma viral oncogene homologue |
| Pb | Lead |
| lncRNA | long non-coding RNA |
| mTOR | Mammalian target of rapamycin |
| miRNAs | Micro-RNAs |
| RWPE-1 | Normal human prostate cells |
| NFKB1 | Nuclear Factor Kappa B Subunit 1 |
| NF-kB | Nuclear Factor Kappa-Light-Chain-Enhancer Of Activated B Cells |
| PM | Particulate matter |
| PTEN | Phosphatase and Tensin Homolog |
| PDGFB | Platelet-Derived Growth Factor Subunit B |
| ROS | Reactive Oxygen Species |
| RISC | RNA-induced silencing complex |
| WHO | The World Health Organization |
| TLR4 | Toll-Like Receptor 4 |
| VEGF | Vascular Endothelial Growth Factor |
| VCAM-1 | Vascular adhesion molecule 1 |
| Wnt | Wingless/Int |
References
- Kelly, F.J.; Fussell, J.C. Air pollution and public health: Emerging hazards and improved understanding of risk. Environ. Geochem. Health 2015, 37, 631–649. [Google Scholar] [CrossRef]
- Biswas, J.K.; Mondal, M.; Rinklebe, J.; Sarkar, S.K.; Chaudhuri, P.; Rai, M.; Shaheen, S.M.; Song, H.; Rizwan, M. Multi-metal resistance and plant growth promotion potential of a wastewater bacterium Pseudomonas aeruginosa and its synergistic benefits. Environ. Geochem. Health 2017, 39, 1583–1593. [Google Scholar] [CrossRef]
- Elleuch, B.; Bouhamed, F.; Elloussaief, M.; Jaghbir, M. Environmental sustainability and pollution prevention. Environ. Sci. Pollut. Res. Int. 2018, 25, 18223–18225. [Google Scholar] [CrossRef]
- Brockmeyer, S.; D’Angiulli, A. How air pollution alters brain development: The role of neuroinflammation. Transl. Neurosci. 2016, 7, 24–30. [Google Scholar] [CrossRef]
- Holland, N. Future of environmental research in the age of epigenomics and exposomics. Rev. Environ. Health 2017, 32, 45–54. [Google Scholar] [CrossRef]
- Manisalidis, I.; Stavropoulou, E.; Stavropoulos, A.; Bezirtzoglou, E. Environmental and Health Impacts of Air Pollution: A Review. Front. Public Health 2020, 8, 14. [Google Scholar] [CrossRef] [PubMed]
- Gianicolo, E.A.; Bruni, A.; Rosati, E.; Sabina, S.; Guarino, R.; Padolecchia, G.; Leo, C.; Vigotti, M.A.; Andreassi, M.G.; Latini, G. Congenital anomalies among live births in a polluted area. A ten-year retrospective study. BMC Pregnancy Childbirth 2012, 12, 165. [Google Scholar] [CrossRef] [PubMed]
- Alfano, R.; Herceg, Z.; Nawrot, T.S.; Chadeau-Hyam, M.; Ghantous, A.; Plusquin, M. The Impact of Air Pollution on Our Epigenome: How Far Is the Evidence? (A Systematic Review). Curr. Environ. Health Rep. 2018, 5, 544–578. [Google Scholar] [CrossRef]
- Fiorito, G.; Vlaanderen, J.; Polidoro, S.; Gulliver, J.; Galassi, C.; Ranzi, A.; Krogh, V.; Grioni, S.; Agnoli, C.; Sacerdote, C.; et al. Oxidative stress and inflammation mediate the effect of air pollution on cardio- and cerebrovascular disease: A prospective study in nonsmokers. Environ. Mol. Mutagen. 2018, 59, 234–246. [Google Scholar] [CrossRef]
- Hamra, G.B.; Guha, N.; Cohen, A.; Laden, F.; Raaschou-Nielsen, O.; Samet, J.M.; Vineis, P.; Forastiere, F.; Saldiva, P.; Yorifuji, T.; et al. Outdoor particulate matter exposure and lung cancer: A systematic review and meta-analysis. Environ. Health Perspect. 2014, 122, 906–911. [Google Scholar] [CrossRef]
- Hoek, G.; Krishnan, R.M.; Beelen, R.; Peters, A.; Ostro, B.; Brunekreef, B.; Kaufman, J.D. Long-term air pollution exposure and cardio- respiratory mortality: A review. Environ. Health 2013, 12, 43. [Google Scholar] [CrossRef]
- Forte, A.; Finicelli, M.; Grossi, M.; Vicchio, M.; Alessio, N.; Sante, P.; De Feo, M.; Cotrufo, M.; Berrino, L.; Rossi, F.; et al. DNA damage and repair in a model of rat vascular injury. Clin. Sci. 2010, 118, 473–485. [Google Scholar] [CrossRef][Green Version]
- Mirbahai, L.; Chipman, J.K. Epigenetic memory of environmental organisms: A reflection of lifetime stressor exposures. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2014, 764, 10–17. [Google Scholar] [CrossRef]
- Li, J.; Wang, T.; Wang, Y.; Xu, M.; Zhang, L.; Li, X.; Liu, Z.; Gao, S.; Jia, Q.; Fan, Y.; et al. Particulate matter air pollution and the expression of microRNAs and pro-inflammatory genes: Association and mediation among children in Jinan, China. J. Hazard. Mater. 2020, 389, 121843. [Google Scholar] [CrossRef]
- Wang, F.; Liu, J.; Zeng, H. Interactions of particulate matter and pulmonary surfactant: Implications for human health. Adv. Colloid Interface Sci. 2020, 284, 102244. [Google Scholar] [CrossRef]
- Popoola, L.T.; Adebanjo, S.A.; Adeoye, B.K. Assessment of atmospheric particulate matter and heavy metals: A critical review. Int. J. Environ. Sci. Technol. 2018, 15, 935–948. [Google Scholar] [CrossRef]
- WHO. Health Effects of Particulate Matter; WHO: Copenhagen Ø, Denmark, 2013. [Google Scholar]
- Schraufnagel, D.E.; Balmes, J.R.; Cowl, C.T.; De Matteis, S.; Jung, S.H.; Mortimer, K.; Perez-Padilla, R.; Rice, M.B.; Riojas-Rodriguez, H.; Sood, A.; et al. Air Pollution and Noncommunicable Diseases: A Review by the Forum of International Respiratory Societies’ Environmental Committee, Part 1: The Damaging Effects of Air Pollution. Chest 2019, 155, 409–416. [Google Scholar] [CrossRef]
- WHO. Ambient Air Pollution: Health Impacts. Available online: https://www.who.int/airpollution/ambient/health-impacts/en/ (accessed on 24 September 2020).
- Lu, F.; Xu, D.; Cheng, Y.; Dong, S.; Guo, C.; Jiang, X.; Zheng, X. Systematic review and meta-analysis of the adverse health effects of ambient PM2.5 and PM10 pollution in the Chinese population. Environ. Res. 2015, 136, 196–204. [Google Scholar] [CrossRef]
- Fiordelisi, A.; Piscitelli, P.; Trimarco, B.; Coscioni, E.; Iaccarino, G.; Sorriento, D. The mechanisms of air pollution and particulate matter in cardiovascular diseases. Heart Fail. Rev. 2017, 22, 337–347. [Google Scholar] [CrossRef]
- Leepe, K.A.; Li, M.; Fang, X.; Hiyoshi, A.; Cao, Y. Acute effect of daily fine particulate matter pollution on cerebrovascular mortality in Shanghai, China: A population-based time series study. Environ. Sci. Pollut. Res. Int. 2019, 26, 25491–25499. [Google Scholar] [CrossRef]
- Martins, V.; Faria, T.; Diapouli, E.; Manousakas, M.I.; Eleftheriadis, K.; Viana, M.; Almeida, S.M. Relationship between indoor and outdoor size-fractionated particulate matter in urban microenvironments: Levels, chemical composition and sources. Environ. Res. 2020, 183, 109203. [Google Scholar] [CrossRef] [PubMed]
- Sahu, D.; Kannan, G.M.; Vijayaraghavan, R. Carbon black particle exhibits size dependent toxicity in human monocytes. Int. J. Inflam. 2014, 2014, 827019. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, M.; Ahmadi, A.; Baradaran, A.; Masoudipoor, N.; Frouzandeh, S. Modeling of the relationship between the environmental air pollution, clinical risk factors, and hospital mortality due to myocardial infarction in Isfahan, Iran. J. Res. Med. Sci. 2015, 20, 757–762. [Google Scholar] [CrossRef] [PubMed]
- Ghorani-Azam, A.; Riahi-Zanjani, B.; Balali-Mood, M. Effects of air pollution on human health and practical measures for prevention in Iran. J. Res. Med. Sci. 2016, 21, 65. [Google Scholar] [CrossRef]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef]
- Yu, R.; He, L.; Cai, R.; Li, B.; Li, Z.; Yang, K. Heavy metal pollution and health risk in China. GHJ 2017, 1, 47–55. [Google Scholar] [CrossRef]
- Anyanwu, B.O.; Ezejiofor, A.N.; Igweze, Z.N.; Orisakwe, O.E. Heavy Metal Mixture Exposure and Effects in Developing Nations: An Update. Toxics 2018, 6, 65. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Ilahi, I. Environmental Chemistry and Ecotoxicology of Hazardous Heavy Metals: Environmental Persistence, Toxicity, and Bioaccumulation. J. Chem. 2019. [Google Scholar] [CrossRef]
- Alghamdi, M.A. Characteristics and Risk Assessment of Heavy Metals in Airborne PM10 from a Residential Area of Northern Jeddah City, Saudi Arabia. Pol. J. Environ. Stud. 2016, 25, 939–949. [Google Scholar] [CrossRef]
- Li, H.; Qian, X.; Wang, Q. Heavy metals in atmospheric particulate matter: A comprehensive understanding is needed for monitoring and risk mitigation. Environ. Sci. Technol. 2013, 47, 13210–13211. [Google Scholar] [CrossRef]
- Di Vaio, P.; Magli, E.; Caliendo, G.; Corvino, A.; Fiorino, F.; Frecentese, F.; Saccone, I.; Santagada, V.; Severino, B.; Onorati, G.; et al. Heavy Metals Size Distribution in PM10 and Environmental-Sanitary Risk Analysis in Acerra (Italy). Atmosphere 2018, 9, 58. [Google Scholar] [CrossRef]
- WHO. 2011. Available online: https://www.who.int/water_sanitation_health/dwq/chemicals/arsenic.pdf (accessed on 22 September 2020).
- Paul, S.; Giri, A.K. Epimutagenesis: A prospective mechanism to remediate arsenic-induced toxicity. Environ. Int. 2015, 81, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Berg, C.J.; An, L.C.; Thomas, J.L.; Lust, K.A.; Sanem, J.R.; Swan, D.W.; Ahluwalia, J.S. Smoking patterns, attitudes and motives: Unique characteristics among 2-year versus 4-year college students. Health Educ. Res. 2011, 26, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Maud, J.; Rumsby, P. A Review of the Toxicity of Arsenic in Air; SC020104/SR4; Environment Agency: Bristol, UK, 2008. [Google Scholar]
- Smith, A.H.; Lingas, E.O.; Rahman, M. Contamination of drinking-water by arsenic in Bangladesh: A public health emergency. Bull. World Health Organ. 2000, 78, 1093–1103. [Google Scholar]
- Rosado, J.L.; Ronquillo, D.; Kordas, K.; Rojas, O.; Alatorre, J.; Lopez, P.; Garcia-Vargas, G.; Del Carmen Caamano, M.; Cebrian, M.E.; Stoltzfus, R.J. Arsenic exposure and cognitive performance in Mexican schoolchildren. Environ. Health Perspect. 2007, 115, 1371–1375. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Manchester, L.C.; Tan, D.X. Neurotoxins: Free radical mechanisms and melatonin protection. Curr. Neuropharmacol. 2010, 8, 194–210. [Google Scholar] [CrossRef]
- Chen, P.H.; Ko, Y.C.; Yang, Y.H.; Lin, Y.C.; Shieh, T.Y.; Chen, C.H.; Tsai, C.C. Important prognostic factors of long-term oropharyngeal carcinoma survivors in Taiwan. Oral Oncol. 2004, 40, 847–855. [Google Scholar] [CrossRef]
- Simeonova, P.P.; Luster, M.I. Arsenic and atherosclerosis. Toxicol. Appl. Pharmacol. 2004, 198, 444–449. [Google Scholar] [CrossRef]
- Willerson, J.T.; Ridker, P.M. Inflammation as a cardiovascular risk factor. Circulation 2004, 109, II2-10. [Google Scholar] [CrossRef]
- Wauson, E.M.; Langan, A.S.; Vorce, R.L. Sodium arsenite inhibits and reverses expression of adipogenic and fat cell-specific genes during in vitro adipogenesis. Toxicol. Sci. 2002, 65, 211–219. [Google Scholar] [CrossRef]
- Sabir, S.; Akash, M.S.H.; Fiayyaz, F.; Saleem, U.; Mehmood, M.H.; Rehman, K. Role of cadmium and arsenic as endocrine disruptors in the metabolism of carbohydrates: Inserting the association into perspectives. Biomed. Pharmacother. 2019, 114, 108802. [Google Scholar] [CrossRef]
- Schwerdtle, T.; Walter, I.; Mackiw, I.; Hartwig, A. Induction of oxidative DNA damage by arsenite and its trivalent and pentavalent methylated metabolites in cultured human cells and isolated DNA. Carcinogenesis 2003, 24, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, J.S.; Chang, E.T.; Gentry, P.R.; Clewell, H.J.; Boffetta, P.; Cohen, S.M. Dose-response for assessing the cancer risk of inorganic arsenic in drinking water: The scientific basis for use of a threshold approach. Crit. Rev. Toxicol. 2019, 49, 36–84. [Google Scholar] [CrossRef] [PubMed]
- Schoen, A.; Beck, B.; Sharma, R.; Dube, E. Arsenic toxicity at low doses: Epidemiological and mode of action considerations. Toxicol. Appl. Pharmacol. 2004, 198, 253–267. [Google Scholar] [CrossRef]
- Rehman, K.; Fatima, F.; Waheed, I.; Akash, M.S.H. Prevalence of exposure of heavy metals and their impact on health consequences. J. Cell. Biochem. 2018, 119, 157–184. [Google Scholar] [CrossRef]
- Williams, C.R.; Harrison, R.M. Cadmium in the atmosphere. Experientia 1984, 40, 29–36. [Google Scholar] [CrossRef]
- Tarakina, N.V.; Verberck, B. A portrait of cadmium. Nat. Chem. 2016, 9, 96. [Google Scholar] [CrossRef]
- Satarug, S.; Garrett, S.H.; Sens, M.A.; Sens, D.A. Cadmium, environmental exposure, and health outcomes. Ciencia Saude Coletiva 2011, 16, 2587–2602. [Google Scholar] [CrossRef] [PubMed]
- Henson, M.C.; Chedrese, P.J. Endocrine disruption by cadmium, a common environmental toxicant with paradoxical effects on reproduction. Exp. Biol. Med. 2004, 229, 383–392. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Y.; Tan, M.; Liu, J.; Bao, L.; Zhang, G.; Li, Y.; Iida, A. Atmospheric lead pollution in fine particulate matter in Shanghai, China. J. Environ. Sci. 2009, 21, 1118–1124. [Google Scholar] [CrossRef]
- Gerhardsson, L.; Dahlin, L.; Knebel, R.; Schutz, A. Blood lead concentration after a shotgun accident. Environ. Health Perspect 2002, 110, 115–117. [Google Scholar] [CrossRef]
- Wani, A.L.; Ara, A.; Usmani, J.A. Lead toxicity: A review. Interdiscip. Toxicol. 2015, 8, 55–64. [Google Scholar] [CrossRef]
- Mener, D.J.; Garcia-Esquinas, E.; Navas-Acien, A.; Dietert, R.R.; Shargorodsky, J.; Lin, S.Y. Lead exposure and increased food allergic sensitization in U.S. children and adults. Int. Forum Allergy Rhinol. 2015, 5, 214–220. [Google Scholar] [CrossRef]
- Kumar, S. Occupational and Environmental Exposure to Lead and Reproductive Health Impairment: An Overview. Indian J. Occup. Environ. Med. 2018, 22, 128–137. [Google Scholar] [CrossRef]
- WHO. Lead Poisoning and Health. Available online: https://www.who.int/news-room/fact-sheets/detail/lead-poisoning-and-health (accessed on 24 September 2020).
- Yabe, J.; Nakayama, S.M.; Nakata, H.; Toyomaki, H.; Yohannes, Y.B.; Muzandu, K.; Kataba, A.; Zyambo, G.; Hiwatari, M.; Narita, D.; et al. Current trends of blood lead levels, distribution patterns and exposure variations among household members in Kabwe, Zambia. Chemosphere 2020, 243, 125412. [Google Scholar] [CrossRef]
- Bressler, J.P.; Goldstein, G.W. Mechanisms of lead neurotoxicity. Biochem. Pharmacol. 1991, 41, 479–484. [Google Scholar] [CrossRef]
- Ahmad, F.; Salahuddin, M.; Alsamman, K.; AlMulla, A.A.; Salama, K.F. Developmental lead (Pb)-induced deficits in hippocampal protein translation at the synapses are ameliorated by ascorbate supplementation. Neuropsychiatr. Dis. Treat. 2018, 14, 3289–3298. [Google Scholar] [CrossRef]
- Khalil, N.; Morrow, L.A.; Needleman, H.; Talbott, E.O.; Wilson, J.W.; Cauley, J.A. Association of cumulative lead and neurocognitive function in an occupational cohort. Neuropsychology 2009, 23, 10–19. [Google Scholar] [CrossRef]
- Ma, T.; Wu, X.; Cai, Q.; Wang, Y.; Xiao, L.; Tian, Y.; Li, H. Lead Poisoning Disturbs Oligodendrocytes Differentiation Involved in Decreased Expression of NCX3 Inducing Intracellular Calcium Overload. Int. J. Mol. Sci. 2015, 16, 19096–19110. [Google Scholar] [CrossRef]
- Lentini, P.; Zanoli, L.; Granata, A.; Signorelli, S.S.; Castellino, P.; Dell’Aquila, R. Kidney and heavy metals—The role of environmental exposure (Review). Mol. Med. Rep. 2017, 15, 3413–3419. [Google Scholar] [CrossRef]
- Hoffman, J.B.; Hennig, B. Protective influence of healthful nutrition on mechanisms of environmental pollutant toxicity and disease risks. Ann. N. Y. Acad. Sci. 2017, 1398, 99–107. [Google Scholar] [CrossRef]
- Sun, Q.; Wang, A.; Jin, X.; Natanzon, A.; Duquaine, D.; Brook, R.D.; Aguinaldo, J.G.; Fayad, Z.A.; Fuster, V.; Lippmann, M.; et al. Long-term air pollution exposure and acceleration of atherosclerosis and vascular inflammation in an animal model. JAMA 2005, 294, 3003–3010. [Google Scholar] [CrossRef]
- Flora, S.J.; Mittal, M.; Mehta, A. Heavy metal induced oxidative stress & its possible reversal by chelation therapy. Indian J. Med. Res. 2008, 128, 501–523. [Google Scholar]
- Wild, C.P. Complementing the genome with an “exposome”: The outstanding challenge of environmental exposure measurement in molecular epidemiology. Cancer Epidemiol. Biomarkers Prev. 2005, 14, 1847–1850. [Google Scholar] [CrossRef]
- Siroux, V.; Agier, L.; Slama, R. The exposome concept: A challenge and a potential driver for environmental health research. Eur. Respir. Rev. 2016, 25, 124–129. [Google Scholar] [CrossRef]
- Miller, G.W.; Jones, D.P. The nature of nurture: Refining the definition of the exposome. Toxicol. Sci. 2014, 137, 1–2. [Google Scholar] [CrossRef]
- Baccarelli, A.; Bollati, V. Epigenetics and environmental chemicals. Curr. Opin. Pediatr. 2009, 21, 243–251. [Google Scholar] [CrossRef]
- Wei, J.W.; Huang, K.; Yang, C.; Kang, C.S. Non-coding RNAs as regulators in epigenetics (Review). Oncol. Rep. 2017, 37, 3–9. [Google Scholar] [CrossRef]
- Fraga, M.F.; Ballestar, E.; Paz, M.F.; Ropero, S.; Setien, F.; Ballestar, M.L.; Heine-Suner, D.; Cigudosa, J.C.; Urioste, M.; Benitez, J.; et al. Epigenetic differences arise during the lifetime of monozygotic twins. Proc. Natl. Acad. Sci. USA 2005, 102, 10604–10609. [Google Scholar] [CrossRef]
- Christensen, B.C.; Houseman, E.A.; Marsit, C.J.; Zheng, S.; Wrensch, M.R.; Wiemels, J.L.; Nelson, H.H.; Karagas, M.R.; Padbury, J.F.; Bueno, R.; et al. Aging and environmental exposures alter tissue-specific DNA methylation dependent upon CpG island context. PLoS Genet. 2009, 5, e1000602. [Google Scholar] [CrossRef]
- Langevin, S.M.; Houseman, E.A.; Christensen, B.C.; Wiencke, J.K.; Nelson, H.H.; Karagas, M.R.; Marsit, C.J.; Kelsey, K.T. The influence of aging, environmental exposures and local sequence features on the variation of DNA methylation in blood. Epigenetics 2011, 6, 908–919. [Google Scholar] [CrossRef]
- Gronniger, E.; Weber, B.; Heil, O.; Peters, N.; Stab, F.; Wenck, H.; Korn, B.; Winnefeld, M.; Lyko, F. Aging and chronic sun exposure cause distinct epigenetic changes in human skin. PLoS Genet. 2010, 6, e1000971. [Google Scholar] [CrossRef]
- Feil, R.; Fraga, M.F. Epigenetics and the environment: Emerging patterns and implications. Nat. Rev. Genet. 2012, 13, 97–109. [Google Scholar] [CrossRef]
- Wang, J.; Cui, Q. Specific Roles of MicroRNAs in Their Interactions with Environmental Factors. J. Nucleic Acids 2012, 2012, 978384. [Google Scholar] [CrossRef]
- Tumolo, M.R.; Panico, A.; De Donno, A.; Mincarone, P.; Leo, C.G.; Guarino, R.; Bagordo, F.; Serio, F.; Idolo, A.; Grassi, T.; et al. The expression of microRNAs and exposure to environmental contaminants related to human health: A review. Int. J. Environ. Health Res. 2020, 1–23. [Google Scholar] [CrossRef]
- Kaikkonen, M.U.; Lam, M.T.; Glass, C.K. Non-coding RNAs as regulators of gene expression and epigenetics. Cardiovasc. Res. 2011, 90, 430–440. [Google Scholar] [CrossRef]
- Krauskopf, J.; Verheijen, M.; Kleinjans, J.C.; de Kok, T.M.; Caiment, F. Development and regulatory application of microRNA biomarkers. Biomark. Med. 2015, 9, 1137–1151. [Google Scholar] [CrossRef]
- Vrijens, K.; Bollati, V.; Nawrot, T.S. MicroRNAs as potential signatures of environmental exposure or effect: A systematic review. Environ. Health Perspect. 2015, 123, 399–411. [Google Scholar] [CrossRef]
- Siddeek, B.; Inoubli, L.; Lakhdari, N.; Rachel, P.B.; Fussell, K.C.; Schneider, S.; Mauduit, C.; Benahmed, M. MicroRNAs as potential biomarkers in diseases and toxicology. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2014, 764, 46–57. [Google Scholar] [CrossRef]
- Yu, H.W.; Cho, W.C. The role of microRNAs in toxicology. Arch. Toxicol. 2015, 89, 319–325. [Google Scholar] [CrossRef]
- Schraml, E.; Hackl, M.; Grillari, J. MicroRNAs and toxicology: A love marriage. Toxicol. Rep. 2017, 4, 634–636. [Google Scholar] [CrossRef]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The microRNA spectrum in 12 body fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef]
- Wang, K. The Ubiquitous Existence of MicroRNA in Body Fluids. Clin. Chem. 2017, 63, 784–785. [Google Scholar] [CrossRef]
- Izzotti, A.; Calin, G.A.; Arrigo, P.; Steele, V.E.; Croce, C.M.; De Flora, S. Downregulation of microRNA expression in the lungs of rats exposed to cigarette smoke. FASEB J. 2009, 23, 806–812. [Google Scholar] [CrossRef]
- Krauskopf, J.; Caiment, F.; van Veldhoven, K.; Chadeau-Hyam, M.; Sinharay, R.; Chung, K.F.; Cullinan, P.; Collins, P.; Barratt, B.; Kelly, F.J.; et al. The human circulating miRNome reflects multiple organ disease risks in association with short-term exposure to traffic-related air pollution. Environ. Int. 2018, 113, 26–34. [Google Scholar] [CrossRef]
- Quezada-Maldonado, E.M.; Sanchez-Perez, Y.; Chirino, Y.I.; Vaca-Paniagua, F.; Garcia-Cuellar, C.M. miRNAs deregulation in lung cells exposed to airborne particulate matter (PM10) is associated with pathways deregulated in lung tumors. Environ. Pollut. 2018, 241, 351–358. [Google Scholar] [CrossRef]
- Rodosthenous, R.S.; Coull, B.A.; Lu, Q.; Vokonas, P.S.; Schwartz, J.D.; Baccarelli, A.A. Ambient particulate matter and microRNAs in extracellular vesicles: A pilot study of older individuals. Part. Fibre Toxicol. 2016, 13, 13. [Google Scholar] [CrossRef]
- Chen, R.; Li, H.; Cai, J.; Wang, C.; Lin, Z.; Liu, C.; Niu, Y.; Zhao, Z.; Li, W.; Kan, H. Fine Particulate Air Pollution and the Expression of microRNAs and Circulating Cytokines Relevant to Inflammation, Coagulation, and Vasoconstriction. Environ. Health Perspect. 2018, 126, 017007. [Google Scholar] [CrossRef]
- Chen, C.; Yang, Q.; Wang, D.; Luo, F.; Liu, X.; Xue, J.; Yang, P.; Xu, H.; Lu, J.; Zhang, A.; et al. MicroRNA-191, regulated by HIF-2alpha, is involved in EMT and acquisition of a stem cell-like phenotype in arsenite-transformed human liver epithelial cells. Toxicol. In Vitro 2018, 48, 128–136. [Google Scholar] [CrossRef]
- Ren, X.; Gaile, D.P.; Gong, Z.; Qiu, W.; Ge, Y.; Zhang, C.; Huang, C.; Yan, H.; Olson, J.R.; Kavanagh, T.J.; et al. Arsenic responsive microRNAs in vivo and their potential involvement in arsenic-induced oxidative stress. Toxicol. Appl. Pharmacol. 2015, 283, 198–209. [Google Scholar] [CrossRef]
- Perez-Vazquez, M.S.; Ochoa-Martinez, A.C.; RuIz-Vera, T.; Araiza-Gamboa, Y.; Perez-Maldonado, I.N. Evaluation of epigenetic alterations (mir-126 and mir-155 expression levels) in Mexican children exposed to inorganic arsenic via drinking water. Environ. Sci. Pollut. Res. Int. 2017, 24, 28036–28045. [Google Scholar] [CrossRef]
- Ngalame, N.N.; Waalkes, M.P.; Tokar, E.J. Silencing KRAS Overexpression in Cadmium-Transformed Prostate Epithelial Cells Mitigates Malignant Phenotype. Chem. Res. Toxicol. 2016, 29, 1458–1467. [Google Scholar] [CrossRef]
- Hassan, F.; Nuovo, G.J.; Crawford, M.; Boyaka, P.N.; Kirkby, S.; Nana-Sinkam, S.P.; Cormet-Boyaka, E. MiR-101 and miR-144 regulate the expression of the CFTR chloride channel in the lung. PLoS ONE 2012, 7, e50837. [Google Scholar] [CrossRef]
- Fay, M.J.; Alt, L.A.C.; Ryba, D.; Salamah, R.; Peach, R.; Papaeliou, A.; Zawadzka, S.; Weiss, A.; Patel, N.; Rahman, A.; et al. Cadmium Nephrotoxicity Is Associated with Altered MicroRNA Expression in the Rat Renal Cortex. Toxics 2018, 6, 16. [Google Scholar] [CrossRef]
- Bollati, V.; Marinelli, B.; Apostoli, P.; Bonzini, M.; Nordio, F.; Hoxha, M.; Pegoraro, V.; Motta, V.; Tarantini, L.; Cantone, L.; et al. Exposure to metal-rich particulate matter modifies the expression of candidate microRNAs in peripheral blood leukocytes. Environ. Health Perspect. 2010, 118, 763–768. [Google Scholar] [CrossRef]
- An, J.; Cai, T.; Che, H.; Yu, T.; Cao, Z.; Liu, X.; Zhao, F.; Jing, J.; Shen, X.; Liu, M.; et al. The changes of miRNA expression in rat hippocampus following chronic lead exposure. Toxicol. Lett. 2014, 229, 158–166. [Google Scholar] [CrossRef]
- Xu, M.; Yu, Z.; Hu, F.; Zhang, H.; Zhong, L.; Han, L.; An, Y.; Zhu, B.; Zhang, H. Identification of differential plasma miRNA profiles in Chinese workers with occupational lead exposure. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef]
- Hou, L.; Wang, D.; Baccarelli, A. Environmental chemicals and microRNAs. Mutat. Res. 2011, 714, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Li, S.; Liu, J.; Ni, B. MicroRNA-193b modulates proliferation, migration, and invasion of non-small cell lung cancer cells. Acta Biochimica Biophysica Sinica 2012, 44, 424–430. [Google Scholar] [CrossRef]
- Bleck, B.; Grunig, G.; Chiu, A.; Liu, M.; Gordon, T.; Kazeros, A.; Reibman, J. MicroRNA-375 regulation of thymic stromal lymphopoietin by diesel exhaust particles and ambient particulate matter in human bronchial epithelial cells. J. Immunol. 2013, 190, 3757–3763. [Google Scholar] [CrossRef]
- Miguel, V.; Cui, J.Y.; Daimiel, L.; Espinosa-Diez, C.; Fernandez-Hernando, C.; Kavanagh, T.J.; Lamas, S. The Role of MicroRNAs in Environmental Risk Factors, Noise-Induced Hearing Loss, and Mental Stress. Antioxid. Redox Signal. 2018, 28, 773–796. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.D.; Yosim, A.; Fry, R.C. Incorporating epigenetic data into the risk assessment process for the toxic metals arsenic, cadmium, chromium, lead, and mercury: Strategies and challenges. Front. Genet. 2014, 5, 201. [Google Scholar] [CrossRef]
- Hou, L.; Zhang, X.; Wang, D.; Baccarelli, A. Environmental chemical exposures and human epigenetics. Int. J. Epidemiol. 2012, 41, 79–105. [Google Scholar] [CrossRef]
- Wild, C.P.; Scalbert, A.; Herceg, Z. Measuring the exposome: A powerful basis for evaluating environmental exposures and cancer risk. Environ. Mol. Mutagen. 2013, 54, 480–499. [Google Scholar] [CrossRef] [PubMed]
- Boogaard, H.; Walker, K.; Cohen, A.J. Air pollution: The emergence of a major global health risk factor. Int. Health 2019, 11, 417–421. [Google Scholar] [CrossRef]
- Jadhav, S.H.; Sarkar, S.N.; Tripathit, H.C. Cytogenetic effects of a mixture of selected metals following subchronic exposure through drinking water in male rats. Indian J. Exp. Biol. 2006, 44, 997–1005. [Google Scholar] [PubMed]
- Hernandez-Garcia, A.; Romero, D.; Gomez-Ramirez, P.; Maria-Mojica, P.; Martinez-Lopez, E.; Garcia-Fernandez, A.J. In vitro evaluation of cell death induced by cadmium, lead and their binary mixtures on erythrocytes of Common buzzard (Buteo buteo). Toxicol. In Vitro 2014, 28, 300–306. [Google Scholar] [CrossRef]
- Bagchi, D.; Vuchetich, P.J.; Bagchi, M.; Hassoun, E.A.; Tran, M.X.; Tang, L.; Stohs, S.J. Induction of oxidative stress by chronic administration of sodium dichromate [chromium VI] and cadmium chloride [cadmium II] to rats. Free Radic. Biol. Med. 1997, 22, 471–478. [Google Scholar] [CrossRef]
- Santra, A.; Maiti, A.; Das, S.; Lahiri, S.; Charkaborty, S.K.; Mazumder, D.N. Hepatic damage caused by chronic arsenic toxicity in experimental animals. J. Toxicol. Clin. Toxicol. 2000, 38, 395–405. [Google Scholar] [CrossRef]
- Aykin-Burns, N.; Laegeler, A.; Kellogg, G.; Ercal, N. Oxidative effects of lead in young and adult Fisher 344 rats. Arch. Environ. Contam. Toxicol. 2003, 44, 417–420. [Google Scholar] [CrossRef]
- Jadhav, S.H.; Sarkar, S.N.; Patil, R.D.; Tripathi, H.C. Effects of subchronic exposure via drinking water to a mixture of eight water-contaminating metals: A biochemical and histopathological study in male rats. Arch. Environ. Contam. Toxicol. 2007, 53, 667–677. [Google Scholar] [CrossRef]
- Ankley, G.T.; Bennett, R.S.; Erickson, R.J.; Hoff, D.J.; Hornung, M.W.; Johnson, R.D.; Mount, D.R.; Nichols, J.W.; Russom, C.L.; Schmieder, P.K.; et al. Adverse outcome pathways: A conceptual framework to support ecotoxicology research and risk assessment. Environ. Toxicol. Chem. 2010, 29, 730–741. [Google Scholar] [CrossRef]
- Ho, S.M.; Johnson, A.; Tarapore, P.; Janakiram, V.; Zhang, X.; Leung, Y.K. Environmental epigenetics and its implication on disease risk and health outcomes. ILAR J. 2012, 53, 289–305. [Google Scholar] [CrossRef]
- Ji, X.; Zhang, Y.; Ku, T.; Yun, Y.; Li, G.; Sang, N. MicroRNA-338-5p modulates pulmonary hypertension-like injuries caused by SO2, NO2 and PM2.5 co-exposure through targeting the HIF-1alpha/Fhl-1 pathway. Toxicol. Res. 2016, 5, 1548–1560. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Pacheco, M.; Hidalgo-Miranda, A.; Romero-Cordoba, S.; Valverde, M.; Rojas, E. MRNA and miRNA expression patterns associated to pathways linked to metal mixture health effects. Gene 2014, 533, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.; Dai, X.; Feng, W.; Huang, S.; Yuan, Y.; Xiao, Y.; Zhang, Z.; Deng, N.; Deng, H.; Zhang, X.; et al. Co-exposure to metals and polycyclic aromatic hydrocarbons, microRNA expression, and early health damage in coke oven workers. Environ. Int. 2019, 122, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Ristovski, Z.D.; Miljevic, B.; Surawski, N.C.; Morawska, L.; Fong, K.M.; Goh, F.; Yang, I.A. Respiratory health effects of diesel particulate matter. Respirology 2012, 17, 201–212. [Google Scholar] [CrossRef]
- Jang, A.S. Particulate Air Pollutants and Respiratory Diseases. In Air Pollution—A Comprehensive Perspective; Haryanto, B., Ed.; Intech Open: London, UK, 2012; pp. 153–174. [Google Scholar] [CrossRef]
- Balmes, J.R. Household air pollution from domestic combustion of solid fuels and health. J. Allergy Clin. Immunol. 2019, 143, 1979–1987. [Google Scholar] [CrossRef]
- WHO. The Top 10 Causes of Death. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 24 September 2020).
- Huang, X.; Zhu, Z.; Guo, X.; Kong, X. The roles of microRNAs in the pathogenesis of chronic obstructive pulmonary disease. Int. Immunopharmacol. 2019, 67, 335–347. [Google Scholar] [CrossRef]
- Pope, C.A., 3rd; Kanner, R.E. Acute effects of PM10 pollution on pulmonary function of smokers with mild to moderate chronic obstructive pulmonary disease. Am. Rev. Respir. Dis. 1993, 147, 1336–1340. [Google Scholar] [CrossRef]
- Zhao, J.; Pu, J.; Hao, B.; Huang, L.; Chen, J.; Hong, W.; Zhou, Y.; Li, B.; Ran, P. LncRNA RP11-86H7.1 promotes airway inflammation induced by TRAPM2.5 by acting as a ceRNA of miRNA-9-5p to regulate NFKB1 in HBECS. Sci. Rep. 2020, 10, 11587. [Google Scholar] [CrossRef]
- Assad, N.A.; Balmes, J.; Mehta, S.; Cheema, U.; Sood, A. Chronic obstructive pulmonary disease secondary to household air pollution. Semin. Respir. Crit. Care Med. 2015, 36, 408–421. [Google Scholar] [CrossRef]
- Li, R.; Kou, X.; Xie, L.; Cheng, F.; Geng, H. Effects of ambient PM2.5 on pathological injury, inflammation, oxidative stress, metabolic enzyme activity, and expression of c-fos and c-jun in lungs of rats. Environ. Sci. Pollut. Res. Int. 2015, 22, 20167–20176. [Google Scholar] [CrossRef] [PubMed]
- Duan, R.R.; Hao, K.; Yang, T. Air pollution and chronic obstructive pulmonary disease. Chronic. Dis. Transl. Med. 2020. [Google Scholar] [CrossRef]
- Tan, B.W.Q.; Sim, W.L.; Cheong, J.K.; Kuan, W.S.; Tran, T.; Lim, H.F. MicroRNAs in chronic airway diseases: Clinical correlation and translational applications. Pharmacol. Res. 2020, 160, 105045. [Google Scholar] [CrossRef] [PubMed]
- Yue, P.; Jing, L.; Zhao, X.; Zhu, H.; Teng, J. Down-regulation of taurine-up-regulated gene 1 attenuates inflammation by sponging miR-9-5p via targeting NF-kappaB1/p50 in multiple sclerosis. Life Sci. 2019, 233, 116731. [Google Scholar] [CrossRef]
- Tsamou, M.; Vrijens, K.; Madhloum, N.; Lefebvre, W.; Vanpoucke, C.; Nawrot, T.S. Air pollution-induced placental epigenetic alterations in early life: A candidate miRNA approach. Epigenetics 2018, 13, 135–146. [Google Scholar] [CrossRef]
- Zhou, T.; Yu, Q.; Sun, C.; Wang, Y.; Zhong, Y.; Wang, G. A pilot study of blood microRNAs and lung function in young healthy adults with fine particulate matter exposure. J. Thorac. Dis. 2018, 10, 7073–7080. [Google Scholar] [CrossRef]
- Molteni, M.; Gemma, S.; Rossetti, C. The Role of Toll-Like Receptor 4 in Infectious and Noninfectious Inflammation. Mediators Inflamm. 2016, 2016, 6978936. [Google Scholar] [CrossRef]
- Kim, J.; Kim, D.Y.; Heo, H.R.; Choi, S.S.; Hong, S.H.; Kim, W.J. Role of miRNA-181a-2-3p in cadmium-induced inflammatory responses of human bronchial epithelial cells. J. Thorac. Dis. 2019, 11, 3055–3069. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Chemical/Source | Model | miRNA Modification | Effect | Reference |
|---|---|---|---|---|
| Particulate matter (PM10; PM2.5) | In vivo | ↑ miR-25-3p, ↑miR-30d-5p and ↑miR-107 | Systemic biomarkers for traffic-related air pollution | [90] |
| ↓ miR133a-3p, ↓miR-499a-5p | Biomarkers for traffic-related air pollution effect in heart | |||
| ↓ miR-433-3p | Biomarkers for traffic-related air pollution effect in brain | |||
| ↑ miR-1224-5p | Biomarkers for traffic-related air pollution effect in pancreas | |||
| In vitro | ↑ miR-4448, ↑miR-6808-5p, ↑miR-3147,↑ miR-1298-3p and ↑miR-125a-3p | Regulatory effect on pathway strictly related to cancer development | [91] | |
| In vivo | ↓ miR-8063, ↓miR-4674, ↓miR-6790-5p, ↓miR-1469 and ↓miR-663a | Regulation of molecular pathways involved in cardiovascular diseases | [92] | |
| ↑ let-7g-5p, ↑miR-126-3p, ↑miR-130a-3p,↑miR-146a-5p,↑miR-150-5p, ↑miR-191-5p, and ↑miR-23a-3p. | ||||
| In vivo | ↓ miR-21-5p, ↓miR-187-3p, ↓miR-146a-5p, ↓miR-1-3p and ↓miR-199a-5p | Modifications of genes regulating immune response, vasoconstriction and coagulation | [93] | |
| Arsenic | In vitro | ↑ miR-191 | Malignant transformation of L-02 cells | [94] |
| In vivo | ↑miR-151;↑miR-183 | Oxidative stress | [95] | |
| ↓ miR26a, ↓miR-423 and ↓miR-148b | ||||
| In vivo | ↓ miR-126 | Increased risk for cardiovascular illnesses | [96] | |
| Cadmium | In vitro | ↑ miR-96 and ↑miR-9 | Malignant transformation of RWPE-1cells | [97] |
| ↓ miR-205, ↓miR-155, ↓miR-373, ↓miR-138 ↓miR-222 and ↑miR-134 | ||||
| In vitro | ↑miR101 and↑ miR-144 | Increased risk of pulmonary disease (e.g.,COPD) | [98] | |
| In vivo | ↑ miR-21-5p, ↑miR-34a-5p, ↑miR-146b-5p, ↑miR-149-3p, ↑miR-224-5p and ↑miR-451-5p | Nephrotoxic phenotype | [99] | |
| ↓ miR-193b-3p, ↓miR-455-3p, and ↓miR-342-3p | ||||
| In vivo | ↑ miR-146a | Increased risk for cancer and cardiovascular diseases | [100] | |
| Lead | In vivo | ↑ miR-204, ↑ miR-211, ↑miR-448, ↑miR-449a, ↑miR-34b, and ↑miR-34 | alteration of neurophysiological pathways and risk of neurodegenerative disease | [101] |
| In vivo | ↑ miR-572 | Systemic biomarkers for Pb-exposure | [102] | |
| ↓ miR-520c-3p, ↓miR-211, and ↓miR-148a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Finicelli, M.; Squillaro, T.; Galderisi, U.; Peluso, G. Micro-RNAs: Crossroads between the Exposure to Environmental Particulate Pollution and the Obstructive Pulmonary Disease. Int. J. Mol. Sci. 2020, 21, 7221. https://doi.org/10.3390/ijms21197221
Finicelli M, Squillaro T, Galderisi U, Peluso G. Micro-RNAs: Crossroads between the Exposure to Environmental Particulate Pollution and the Obstructive Pulmonary Disease. International Journal of Molecular Sciences. 2020; 21(19):7221. https://doi.org/10.3390/ijms21197221
Chicago/Turabian StyleFinicelli, Mauro, Tiziana Squillaro, Umberto Galderisi, and Gianfranco Peluso. 2020. "Micro-RNAs: Crossroads between the Exposure to Environmental Particulate Pollution and the Obstructive Pulmonary Disease" International Journal of Molecular Sciences 21, no. 19: 7221. https://doi.org/10.3390/ijms21197221
APA StyleFinicelli, M., Squillaro, T., Galderisi, U., & Peluso, G. (2020). Micro-RNAs: Crossroads between the Exposure to Environmental Particulate Pollution and the Obstructive Pulmonary Disease. International Journal of Molecular Sciences, 21(19), 7221. https://doi.org/10.3390/ijms21197221